Water–Air Interface to Mimic In Vitro Tumoral Cell Migration in Complex Micro-Environments

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
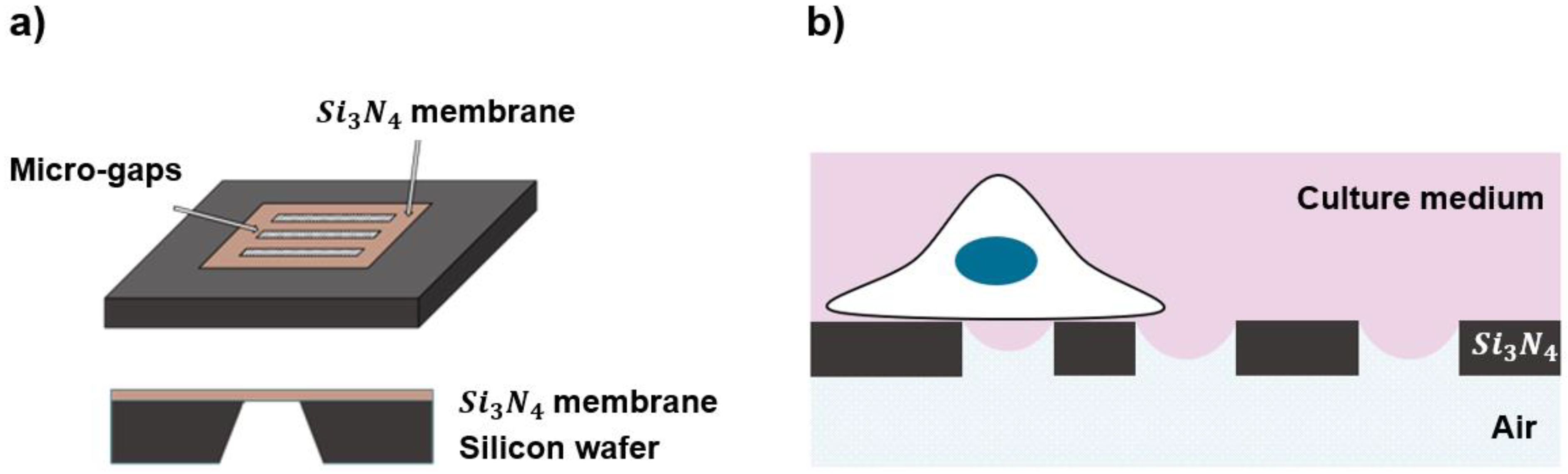
2.1. Fabrication of the Substrates
2.2. Half-Wetting Assembly
2.3. Functionalization of the Substrates
2.4. Cell Culture
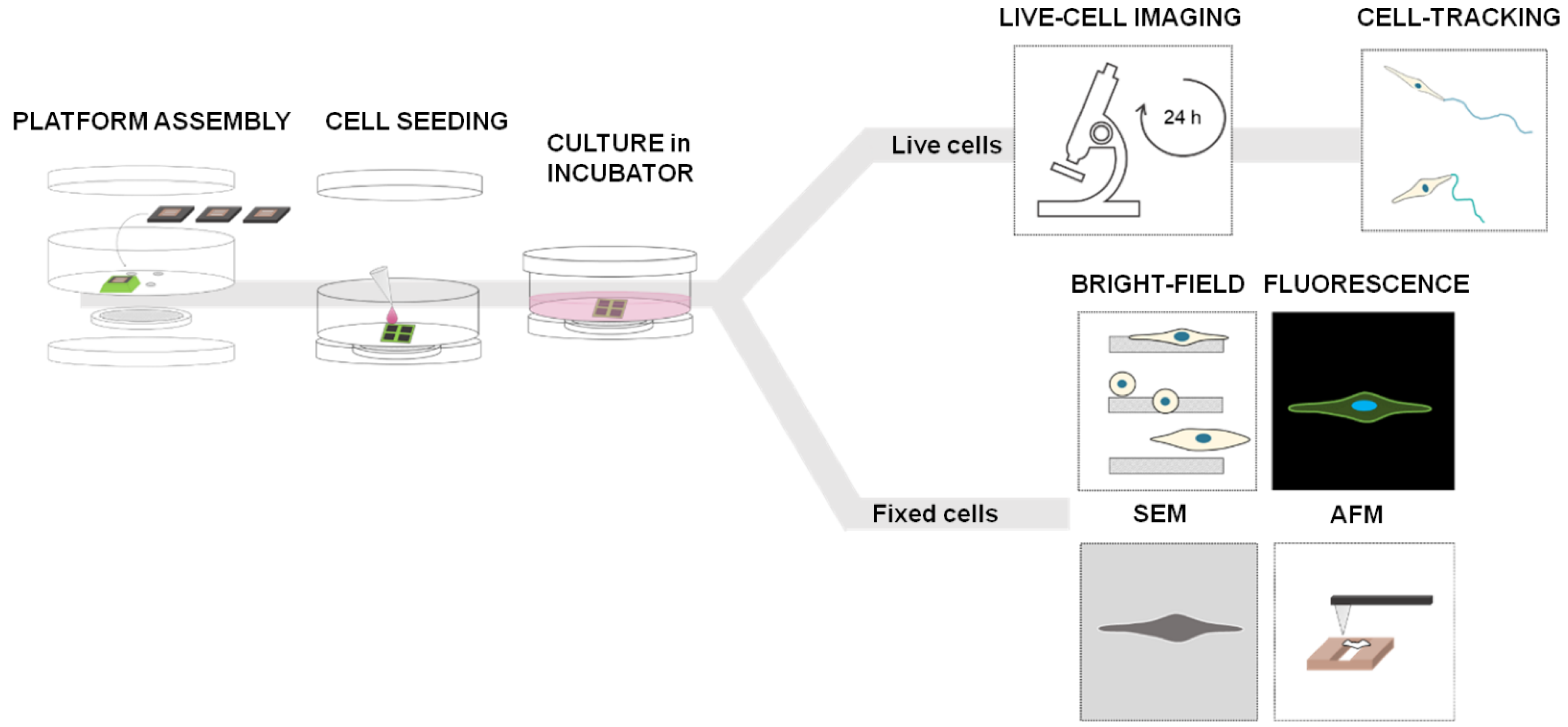
2.5. Experimental Procedure
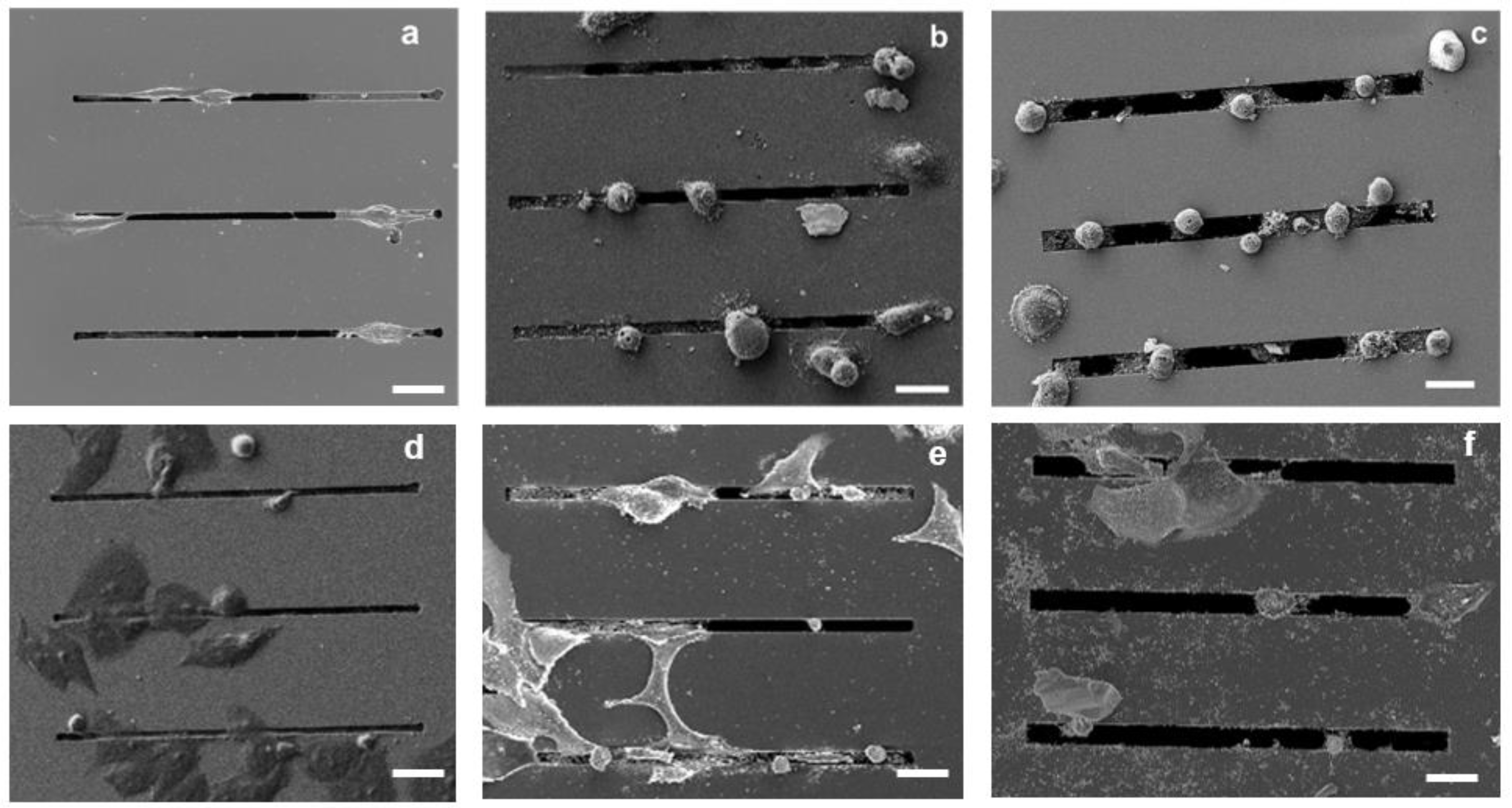
2.6. SEM Imaging
2.7. AFM Imaging
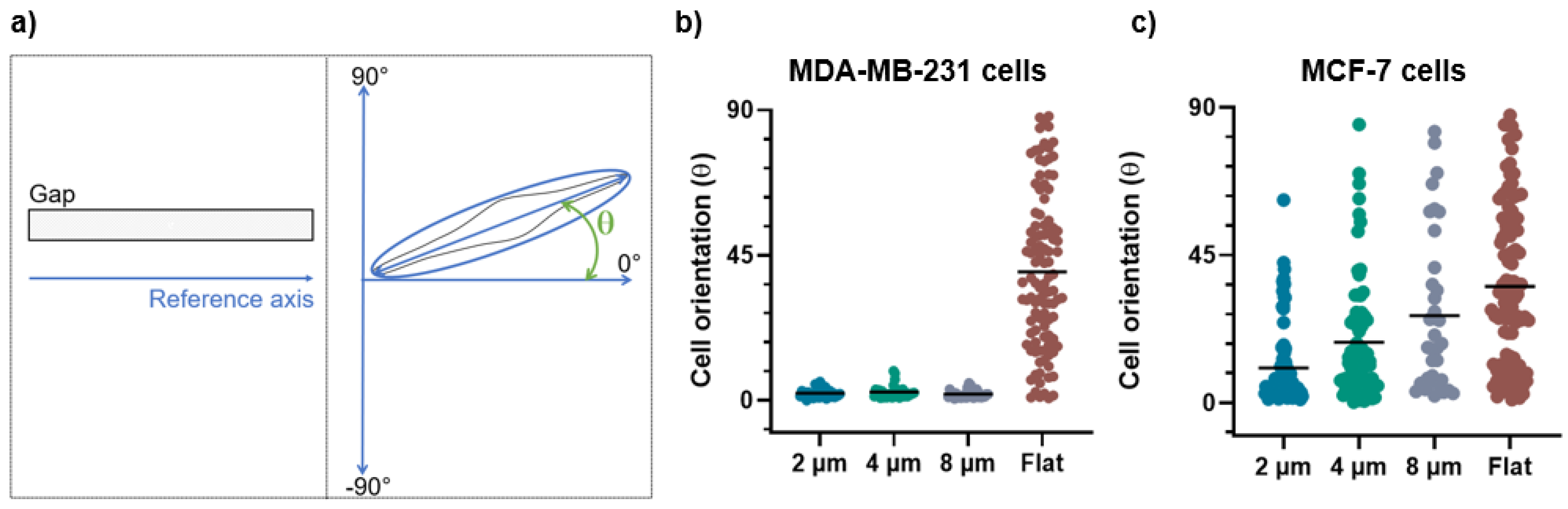
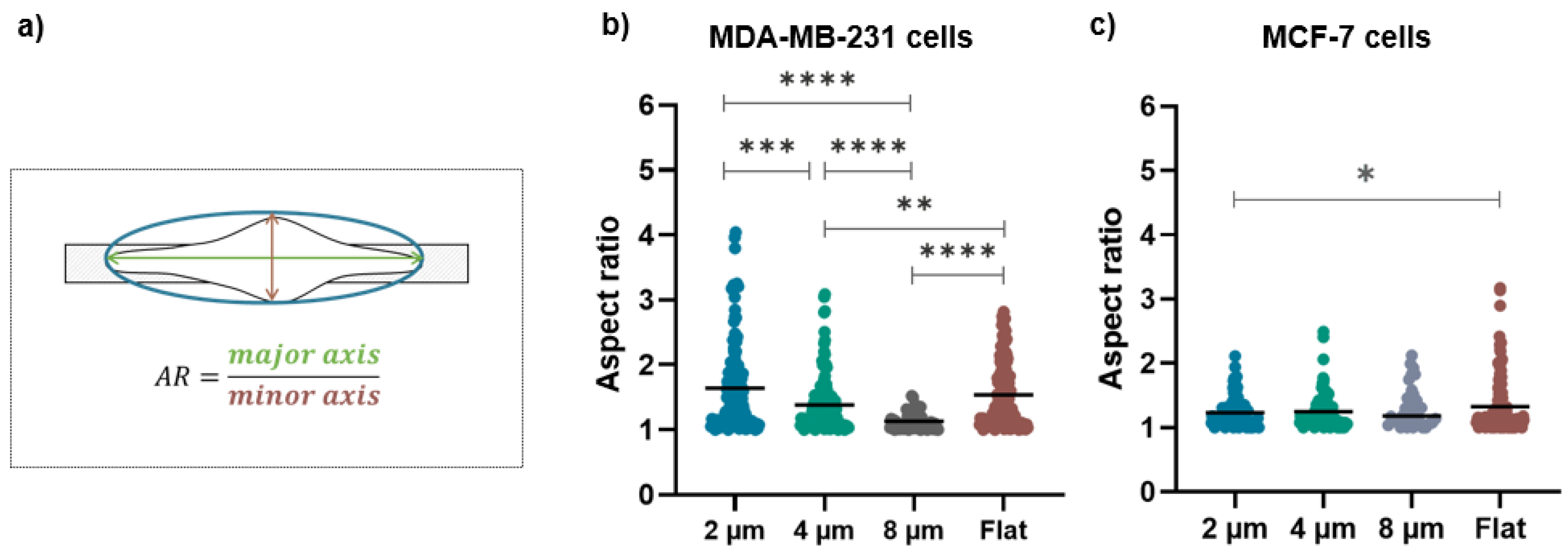
2.8. Morphological Analysis
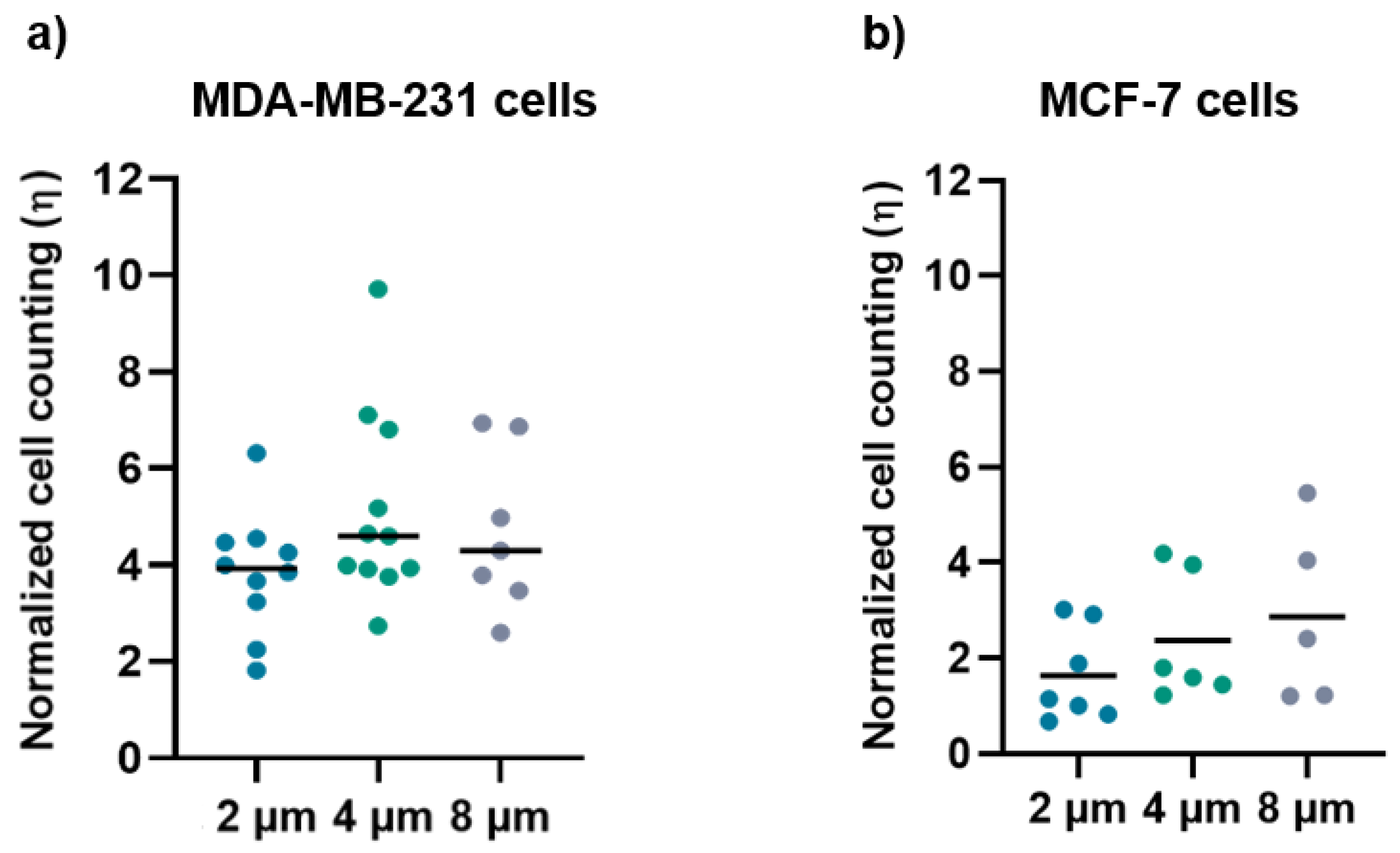
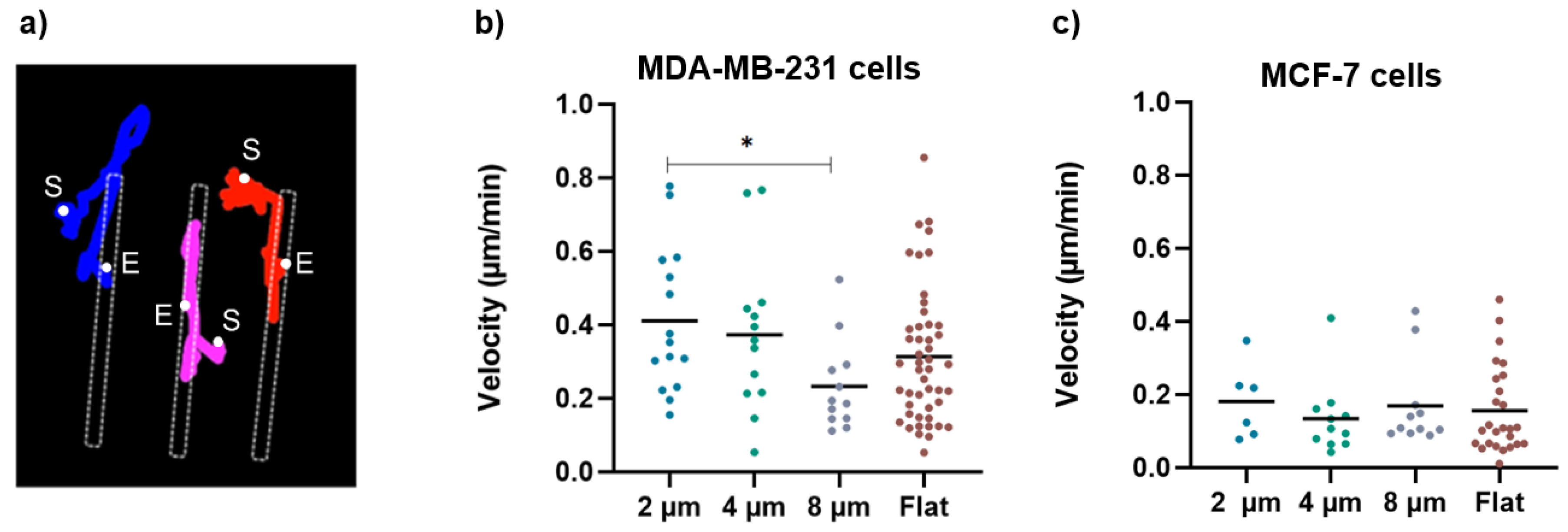
2.9. Migration and Density Analysis
2.10. Statistical Analysis
3. Results
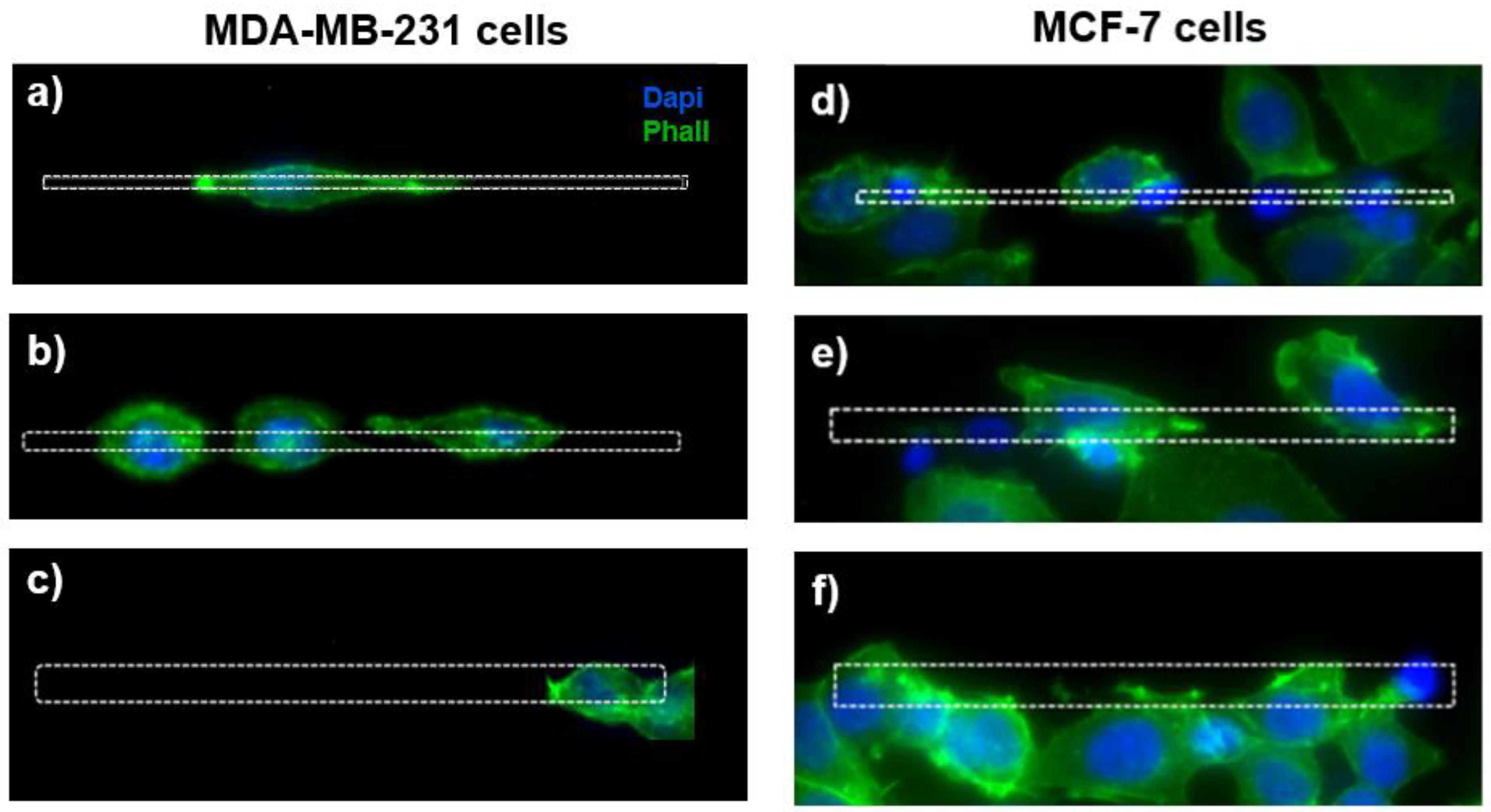
3.1. Morphological Evaluations
3.2. Migration and Cell Density Analysis
3.3. Matrix Deposition
4. Discussion
4.1. Design and Fabrication of the 2D Micro-Pattern Assembly
4.2. Cell Morphology as a Function of the Micro-Gap Size
4.3. Cell Migration in Micro-Gaps
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Galarza, S.; Kim, H.; Atay, N.; Peyton, S.R.; Munson, J.M. 2D or 3D? How Cell Motility Measurements Are Conserved across Dimensions in Vitro and Translate In Vivo. Bioeng. Transl. Med. 2019, 5, e10148. [Google Scholar] [CrossRef]
- Pijuan, J.; Barceló, C.; Moreno, D.F.; Maiques, O.; Sisó, P.; Marti, R.M.; Macià, A.; Panosa, A. In Vitro Cell Migration, Invasion, and Adhesion Assays: From Cell Imaging to Data Analysis. Front. Cell Dev. Biol. 2019, 7, 107. [Google Scholar] [CrossRef] [PubMed]
- Ridley, A.J.; Schwartz, M.A.; Burridge, K.; Firtel, R.A.; Ginsberg, M.H.; Borisy, G.; Parsons, J.T.; Horwitz, A.R. Cell Migration: Integrating Signals from Front to Back. Science 2003, 302, 1704–1709. [Google Scholar] [CrossRef]
- Leclech, C.; Villard, C. Cellular and Subcellular Contact Guidance on Microfabricated Substrates. Front. Bioeng. Biotechnol. 2020, 8, 551505. [Google Scholar] [CrossRef] [PubMed]
- Paul, C.D.; Mistriotis, P.; Konstantopoulos, K. Cancer Cell Motility: Lessons from Migration in Confined Spaces. Nat. Rev. Cancer 2017, 17, 131–140. [Google Scholar] [CrossRef]
- Kramer, N.; Walzl, A.; Unger, C.; Rosner, M.; Krupitza, G.; Hengstschläger, M.; Dolznig, H. In Vitro Cell Migration and Invasion Assays. Mutat. Res. Mutat. Res. 2013, 752, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Ashby, W.J.; Zijlstra, A. Established and Novel Methods of Interrogating Two-Dimensional Cell Migration. Integr. Biol. Quant. Biosci. Nano Macro 2012, 4, 1338–1350. [Google Scholar] [CrossRef]
- Decaestecker, C.; Debeir, O.; Van Ham, P.; Kiss, R. Can Anti-Migratory Drugs Be Screened in Vitro? A Review of 2D and 3D Assays for the Quantitative Analysis of Cell Migration. Med. Res. Rev. 2007, 27, 149–176. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, X.; Cheng, Y.; Cao, X. Dendritic Cell Migration in Inflammation and Immunity. Cell. Mol. Immunol. 2021, 18, 2461–2471. [Google Scholar] [CrossRef]
- Fritzsch, B.; Elliott, K.L.; Pavlinkova, G.; Duncan, J.S.; Hansen, M.R.; Kersigo, J.M. Neuronal Migration Generates New Populations of Neurons That Develop Unique Connections, Physiological Properties and Pathologies. Front. Cell Dev. Biol. 2019, 7, 59. [Google Scholar] [CrossRef]
- Friedl, P.; Alexander, S. Cancer Invasion and the Microenvironment: Plasticity and Reciprocity. Cell 2011, 147, 992–1009. [Google Scholar] [CrossRef] [PubMed]
- Conklin, M.W.; Eickhoff, J.C.; Riching, K.M.; Pehlke, C.A.; Eliceiri, K.W.; Provenzano, P.P.; Friedl, A.; Keely, P.J. Aligned Collagen Is a Prognostic Signature for Survival in Human Breast Carcinoma. Am. J. Pathol. 2011, 178, 1221–1232. [Google Scholar] [CrossRef] [PubMed]
- Gandalovičová, A.; Rosel, D.; Fernandes, M.; Veselý, P.; Heneberg, P.; Čermák, V.; Petruželka, L.; Kumar, S.; Sanz-Moreno, V.; Brábek, J. Migrastatics—Anti-Metastatic and Anti-Invasion Drugs: Promises and Challenges. Trends Cancer 2017, 3, 391–406. [Google Scholar] [CrossRef] [PubMed]
- De Ninno, A.; Bertani, F.R.; Gerardino, A.; Schiavoni, G.; Musella, M.; Galassi, C.; Mattei, F.; Sistigu, A.; Businaro, L. Microfluidic Co-Culture Models for Dissecting the Immune Response in in Vitro Tumor Microenvironments. J. Vis. Exp. JoVE 2021, 170, e61895. [Google Scholar] [CrossRef]
- Shellard, A.; Mayor, R. All Roads Lead to Directional Cell Migration. Trends Cell Biol. 2020, 30, 852–868. [Google Scholar] [CrossRef]
- Pieuchot, L.; Marteau, J.; Guignandon, A.; Dos Santos, T.; Brigaud, I.; Chauvy, P.-F.; Cloatre, T.; Ponche, A.; Petithory, T.; Rougerie, P.; et al. Curvotaxis Directs Cell Migration through Cell-Scale Curvature Landscapes. Nat. Commun. 2018, 9, 3995. [Google Scholar] [CrossRef]
- Lämmermann, T.; Sixt, M. Mechanical Modes of ‘Amoeboid’ Cell Migration. Curr. Opin. Cell Biol. 2009, 21, 636–644. [Google Scholar] [CrossRef]
- Bodor, D.L.; Pönisch, W.; Endres, R.G.; Paluch, E.K. Of Cell Shapes and Motion: The Physical Basis of Animal Cell Migration. Dev. Cell 2020, 52, 550–562. [Google Scholar] [CrossRef]
- Luciano, M.; Xue, S.-L.; De Vos, W.H.; Redondo-Morata, L.; Surin, M.; Lafont, F.; Hannezo, E.; Gabriele, S. Cell Monolayers Sense Curvature by Exploiting Active Mechanics and Nuclear Mechanoadaptation. Nat. Phys. 2021, 17, 1382–1390. [Google Scholar] [CrossRef]
- Wang, K.; Wu, F.; Seo, B.R.; Fischbach, C.; Chen, W.; Hsu, L.; Gourdon, D. Breast Cancer Cells Alter the Dynamics of Stromal Fibronectin-Collagen Interactions. Matrix Biol. 2017, 60–61, 86–95. [Google Scholar] [CrossRef]
- Paul, C.D.; Shea, D.J.; Mahoney, M.R.; Chai, A.; Laney, V.; Hung, W.-C.; Konstantopoulos, K. Interplay of the Physical Microenvironment, Contact Guidance, and Intracellular Signaling in Cell Decision Making. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2016, 30, 2161–2170. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, P.K.; Pan, C.Q.; Low, B.C.; Lim, C.T. Topography Induces Differential Sensitivity on Cancer Cell Proliferation via Rho-ROCK-Myosin Contractility. Sci. Rep. 2016, 6, 19672. [Google Scholar] [CrossRef]
- Holle, A.W.; Govindan Kutty Devi, N.; Clar, K.; Fan, A.; Saif, T.; Kemkemer, R.; Spatz, J.P. Cancer Cells Invade Confined Microchannels via a Self-Directed Mesenchymal-to-Amoeboid Transition. Nano Lett. 2019, 19, 2280–2290. [Google Scholar] [CrossRef]
- Ambrosetti, E.; Bernardinelli, G.; Hoffecker, I.; Hartmanis, L.; Kiriako, G.; de Marco, A.; Sandberg, R.; Högberg, B.; Teixeira, A.I. A DNA-Nanoassembly-Based Approach to Map Membrane Protein Nanoenvironments. Nat. Nanotechnol. 2021, 16, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Ambrosetti, E.; Conti, M.; Teixeira, A.I.; Zilio, S.D. Patterned Carboxymethyl-Dextran Functionalized Surfaces Using Organic Mixed Monolayers for Biosensing Applications. ACS Appl. Bio Mater. 2022, 5, 3310–3319. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.-X.; Yue, G.G.-L.; Dong, J.-R.; Lam, C.W.-K.; Wong, C.-K.; Qiu, M.-H.; Lau, C.B.-S. Actein Inhibits the Proliferation and Adhesion of Human Breast Cancer Cells and Suppresses Migration In Vivo. Front. Pharmacol. 2018, 9, 1466. [Google Scholar] [CrossRef]
- Yadav, A.R.; Sriram, R.; Carter, J.A.; Miller, B.L. Comparative Study of Solution–Phase and Vapor–Phase Deposition of Aminosilanes on Silicon Dioxide Surfaces. Mater. Sci. Eng. C 2014, 35, 283–290. [Google Scholar] [CrossRef]
- Lee, M.H.; Brass, D.A.; Morris, R.; Composto, R.J.; Ducheyne, P. The Effect of Non-Specific Interactions on Cellular Adhesion Using Model Surfaces. Biomaterials 2005, 26, 1721–1730. [Google Scholar] [CrossRef]
- Moore, M.J.; Sebastian, J.A.; Kolios, M.C. Determination of Cell Nucleus-to-Cytoplasmic Ratio Using Imaging Flow Cytometry and a Combined Ultrasound and Photoacoustic Technique: A Comparison Study. J. Biomed. Opt. 2019, 24, 106502. [Google Scholar] [CrossRef]
- Ray, A.; Lee, O.; Win, Z.; Edwards, R.M.; Alford, P.W.; Kim, D.-H.; Provenzano, P.P. Anisotropic Forces from Spatially Constrained Focal Adhesions Mediate Contact Guidance Directed Cell Migration. Nat. Commun. 2017, 8, 14923. [Google Scholar] [CrossRef]
- Andolfi, L.; Murello, A.; Cassese, D.; Ban, J.; Zilio, S.D.; Lazzarino, M. High Aspect Ratio Silicon Nanowires Control Fibroblast Adhesion and Cytoskeleton Organization. Nanotechnology 2017, 28, 155102. [Google Scholar] [CrossRef] [PubMed]
- Ansardamavandi, A.; Tafazzoli-Shadpour, M.; Shokrgozar, M.A. Behavioral Remodeling of Normal and Cancerous Epithelial Cell Lines with Differing Invasion Potential Induced by Substrate Elastic Modulus. Cell Adhes. Migr. 2018, 12, 472–488. [Google Scholar] [CrossRef] [PubMed]
- Yaginuma, T.; Kushiro, K.; Takai, M. Unique Cancer Migratory Behaviors in Confined Spaces of Microgroove Topography with Acute Wall Angles. Sci. Rep. 2020, 10, 6110. [Google Scholar] [CrossRef]
- Wolf, K.; Mazo, I.; Leung, H.; Engelke, K.; von Andrian, U.H.; Deryugina, E.I.; Strongin, A.Y.; Bröcker, E.-B.; Friedl, P. Compensation Mechanism in Tumor Cell Migration: Mesenchymal–Amoeboid Transition after Blocking of Pericellular Proteolysis. J. Cell Biol. 2003, 160, 267–277. [Google Scholar] [CrossRef]
- Juste-Lanas, Y.; Guerrero, P.; Camacho-Gomez, D.; Hervas-Raluy, S.; Garcia Aznar, J.M.; Gomez-Benito, M.J. Confined Cell Migration and Asymmetric Hydraulic Environments to Evaluate The Metastatic Potential of Cancer Cells. J. Biomech. Eng. 2021, 144, 074502. [Google Scholar] [CrossRef] [PubMed]
- Malandrino, A.; Mak, M.; Kamm, R.D.; Moeendarbary, E. Complex Mechanics of the Heterogeneous Extracellular Matrix in Cancer. Extreme Mech. Lett. 2018, 21, 25–34. [Google Scholar] [CrossRef]
- Papalazarou, V.; Salmeron-Sanchez, M.; Machesky, L.M. Tissue Engineering the Cancer Microenvironment—Challenges and Opportunities. Biophys. Rev. 2018, 10, 1695–1711. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conti, M.; Bolzan, I.; Dal Zilio, S.; Parisse, P.; Andolfi, L.; Lazzarino, M. Water–Air Interface to Mimic In Vitro Tumoral Cell Migration in Complex Micro-Environments. Biosensors 2022, 12, 822. https://doi.org/10.3390/bios12100822
Conti M, Bolzan I, Dal Zilio S, Parisse P, Andolfi L, Lazzarino M. Water–Air Interface to Mimic In Vitro Tumoral Cell Migration in Complex Micro-Environments. Biosensors. 2022; 12(10):822. https://doi.org/10.3390/bios12100822
Chicago/Turabian StyleConti, Martina, Ilaria Bolzan, Simone Dal Zilio, Pietro Parisse, Laura Andolfi, and Marco Lazzarino. 2022. "Water–Air Interface to Mimic In Vitro Tumoral Cell Migration in Complex Micro-Environments" Biosensors 12, no. 10: 822. https://doi.org/10.3390/bios12100822
APA StyleConti, M., Bolzan, I., Dal Zilio, S., Parisse, P., Andolfi, L., & Lazzarino, M. (2022). Water–Air Interface to Mimic In Vitro Tumoral Cell Migration in Complex Micro-Environments. Biosensors, 12(10), 822. https://doi.org/10.3390/bios12100822