Genetically Encoded Fluorescent Indicators for Imaging Brain Chemistry

Abstract
1. Introduction
2. Advantages of Genetically Encoded Fluorescent Indicators
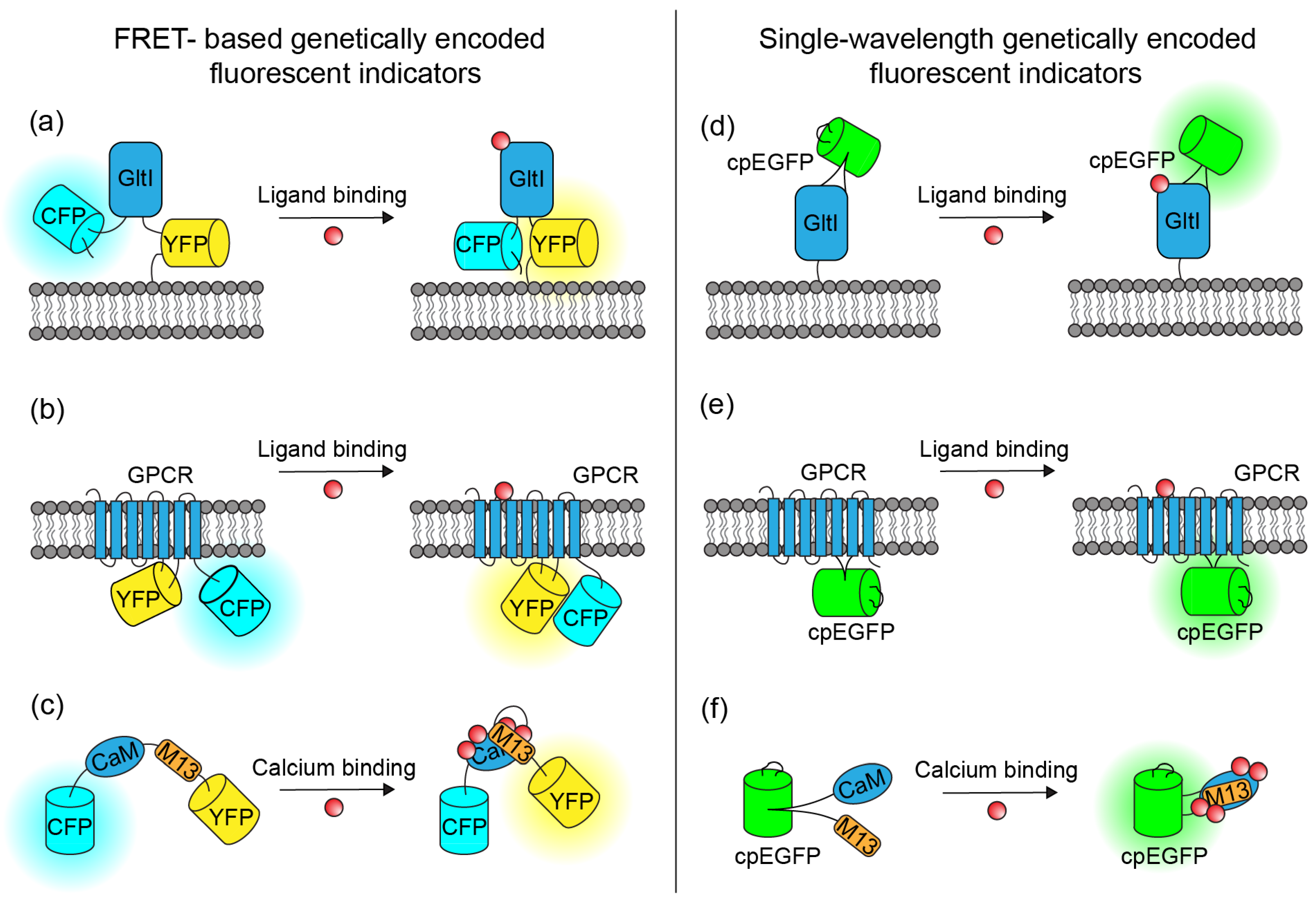
3. Development and Architectures of Genetically Encoded Fluorescent Indicators
4. Genetically Encoded Neurotransmitter Indicators
4.1. PBP-Based Indicators
4.2. GPCR-Based Indicators
5. Genetically Encoded Calcium Indicators
5.1. FRET-Based GECIs

{kind=link}
| Genetically Encoded Calcium Indicator | Ca2+-Binding Domain | Reporter Elements | Dynamic Range ΔR/R0 or ΔF/F0 (In Vitro Unless Otherwise Noted) | Affinity (Kd) (In Vitro Unless Otherwise Noted) | Kinetics |
|---|---|---|---|---|---|
| FRET-based GECIs | |||||
| YC 2 [44] | CaM | ECFP/EYFP | 1.8 | 100 nM | τd = 83 ms |
| YC 2.6 [45] | CaM | ECFP/cpVenus | 6.6 | 95 nM | rise T1/2 = 185 ms|decay T1/2 = 2.31 s (in neuron) |
| YC 3.6 [45] | CaM | ECFP/cpVenus | 5.6 | 250 nM | rise T1/2 = 214 ms|decay T1/2 = 0.4 s (in neuron) |
| YC-Nano15 [48] | CaM | ECFP/cpVenus | 14.5 | 15.8 nM | rise T1/2 = 159 ms|decay T1/2 = 2.38 s (in neuron) |
| iGECI [51] | CaM | miRFP670/miRFP720 | 6 | 15 nM/890 nM | rise T = 0.70 s|decay T = 14 s |
| D3cpv [52] | CaM | ECFP/cpVenus | 5.1 | 600 nM | rise T1/2 = 108 ms|decay T1/2 = 9.5s (in neuron) |
| D4cpv [52] | CaM | ECFP/cpVenus | 3.8 | 60 µM | ND |
| TN-XXL [55] | TnC | ECFP/cpCitrine | 3.3 | 800 nM | rise T1/2 = 80 ms|decay T1/2 = 1.6s (in neuron) |
| Twitch-2B [56] | TnC | mCerulean3/cpVenus | 8 | 200 nM | decay T1/2 = 2.1 s (in neurons) |
| Single-fluorophore GECIs | |||||
| GCaMP 1 [57] | CaM | cpEGFP | 4.5 | 240 nM | τd = 200 ms |
| GCaMP 1.6 [20] | CaM | cpEGFP | 5 | 146 nM | τd = 260 ms |
| GCaMP 2 [58] | CaM | cpEGFP | 5 | 840 nM | rise T1/2 = 95 ms|decay T1/2 = 480 ms (in brain slice) |
| GCaMP 3 [58] | CaM | cpEGFP | 13.5 | 660 nM | rise T1/2 = 95 ms|decay T1/2 = 650 ms (in neuron) |
| GCaMP-HS [59] | CaM | Superfolder GFP | 4 (in matured motor neurons) | 102 nM | decay T1/2 = 0.92 s (in neuron) |
| Fast-GCaMP-EF20 [60] | CaM | cpEGFP | 18 | 6.12 µM | decay T1/2 = 35 ms |
| GCaMP 5D [61] | CaM | cpEGFP | 22 | 730 nM | kon = 7.8 s−1 (measured at 670 nM) |
| GCaMP 5G [61] | CaM | cpEGFP | 33 | 460 nM | kon = 6.5 s−1 (measured at 670 nM) |
| GCaMP 6s [62] | CaM | cpEGFP | 63 | 140 nM | kon = 4.30 × 106 M−1s−1|koff = 0.69 s−1 |
| GCaMP 6m [62] | CaM | cpEGFP | 38 | 170 nM | koff = 2.06 s−1 |
| GCaMP 6f [62] | CaM | cpEGFP | 52 | 380 nM | rise T1/2 = 74 ms|decay T1/2 = 400 ms (in neuron)|kon = 9.44 × 106 M−1s−1|koff = 4.01 s−1 |
| jGCaMP 7f [63] | CaM | cpEGFP | 30.2 | 174 nM | kon = 1.34 × 107 M−1s−1|koff = 5.86 s−1 |
| jGCaMP 7s [63] | CaM | cpEGFP | 40.4 | 68 nM | kon = 2.15 × 107 M−1s−1|koff = 2.87 s−1 (fast) 0.27 s−1 (slow) |
| jGCaMP 7c [63] | CaM | cpEGFP | 145.6 | 298 nM | kon = 3.56 × 106 M−1s−1|koff = 2.79 s−1 |
| jGCaMP 7b [63] | CaM | cpEGFP | 22.1 | 82 nM | kon = 1.6 × 107 M−1s−1|koff = 4.48 s−1 |
| jGCaMP 8f [64] | CaM | cpEGFP | 78.8 | 334 nM | koff = 37.03 s−1 |
| jGCaMP 8m [64] | CaM | cpEGFP | 45.7 | 108 nM | koff = 18.25 s−1 |
| jGCaMP 8s [64] | CaM | cpEGFP | 49.5 | 46 nM | koff = 3.68 s−1 |
| G-GECO 1.1 [65] | CaM | cpEGFP | 26 | 482 nM | kon = 8.17 × 1015 M−ns−1|koff = 0.675 s−1 (n = 2.6) |
| G-GECO 1.2 [65] | CaM | cpEGFP | 24 | 1.15 µM | kon = 8.55 × 1017 M−ns−1|koff = 0.7 s−1 (n = 3.0) |
| R-GECO 1 [65] | CaM | cpmApple | 16 | 150 nM | kon = 9.52 × 109 M−ns−1|koff = 0.752 s−1 (n = 1.6) |
| B-GECO 1 [65] | CaM | BFP | 7 | 160 nM | kon = 4.68 × 1012 M−ns−1|koff = 0.490 s−1 (n = 2.0) |
| NIR-GECO [66] | CaM | mIFP | 8 | 215 nM | rise T = 1.5 s|decay T = 4.0 s |
| XCaMP-G [67] | CaM | cpEGFP | 80 | 200 nM | rise T = 80 ms|decay T = 190 ms |
| XCaMP-Y [67] | CaM | cpmVenus | 115 | 81 nM | rise T = 65 ms|decay T = 210 ms |
| XCaMP-R [67] | CaM | cpmApple | 20 | 97 nM | rise T = 30 ms|decay T = 200 ms |
5.2. Single-Fluorophore GECIs
5.3. Light-and-Calcium-Gated Transcription Factor System
6. In Vivo Applications of Protein Indicators of Neurochemistry
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lodish, H.; Berk, A.; Zipursky, S.L.; Matsudaira, P.; Baltimore, D.; Darnell, J. Neurotransmitters, Synapses, and Impulse Transmission. In Molecular Cell Biology, 4th ed.; 2000; Available online: https://www.ncbi.nlm.nih.gov/books/NBK21521/ (accessed on 23 November 2020).
- Beck, C.; Zhang, D.; Gong, Y. Enhanced genetically encoded voltage indicators advance their applications in neuroscience. Curr. Opin. Biomed. Eng. 2019, 12, 111–117. [Google Scholar] [CrossRef]
- Shevtsova, Z.; Malik, J.M.I.; Michel, U.; Bähr, M.; Kügler, S. Promoters and serotypes: Targeting of adeno-associated virus vectors for gene transfer in the rat central nervous system in vitro and in vivo. Exp. Physiol. 2005, 90, 53–59. [Google Scholar] [CrossRef]
- Nieuwenhuis, B.; Haenzi, B.; Hilton, S.; Carnicer-Lombarte, A.; Hobo, B.; Verhaagen, J.; Fawcett, J.W. Optimization of adeno-associated viral vector-mediated transduction of the corticospinal tract: Comparison of four promoters. Gene Ther. 2021, 28, 56–74. [Google Scholar] [CrossRef] [PubMed]
- Kwan, K.M.; Fujimoto, E.; Grabher, C.; Mangum, B.D.; Hardy, M.E.; Campbell, D.S.; Parant, J.M.; Yost, H.J.; Kanki, J.P.; Chien, C.-B. The Tol2kit: A multisite gateway-based construction kit for Tol2 transposon transgenesis constructs. Dev. Dyn. 2007, 236, 3088–3099. [Google Scholar] [CrossRef] [PubMed]
- Jenett, A.; Rubin, G.M.; Ngo, T.-T.B.; Shepherd, D.; Murphy, C.; Dionne, H.; Pfeiffer, B.D.; Cavallaro, A.; Hall, D.; Jeter, J.; et al. A GAL4-Driver Line Resource for Drosophila Neurobiology. Cell Rep. 2012, 2, 991–1001. [Google Scholar] [CrossRef] [PubMed]
- Sauer, B.; Henderson, N. Site-specific DNA recombination in mammalian cells by the Cre recombinase of bacteriophage P1. Proc. Natl. Acad. Sci. USA 1988, 85, 5166–5170. [Google Scholar] [CrossRef]
- Kim, H.; Kim, M.; Im, S.-K.; Fang, S. Mouse Cre-LoxP system: General principles to determine tissue-specific roles of target genes. Lab. Anim. Res. 2018, 34, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Sadowski, P.D. The Flp recombinase of the 2-microns plasmid of Saccharomyces cerevisiae. Prog. Nucleic Acid. Res. Mol. Biol. 1995, 51, 53–91. [Google Scholar]
- Madisen, L.; Garner, A.R.; Shimaoka, D.; Chuong, A.S.; Klapoetke, N.C.; Li, L.; van der Bourg, A.; Niino, Y.; Egolf, L.; Monetti, C.; et al. Transgenic mice for intersectional targeting of neural sensors and effectors with high specificity and performance. Neuron 2015, 85, 942–958. [Google Scholar] [CrossRef]
- Song, C.; Do, Q.B.; Antic, S.D.; Knöpfel, T. Transgenic Strategies for Sparse but Strong Expression of Genetically Encoded Voltage and Calcium Indicators. Int. J. Mol. Sci. 2017, 18, 1461. [Google Scholar] [CrossRef]
- Mizrahi, A.; Crowley, J.C.; Shtoyerman, E.; Katz, L.C. High-Resolution In Vivo Imaging of Hippocampal Dendrites and Spines. J. Neurosci. 2004, 24, 3147–3151. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Kleiber, S.; Schmid, L.; Nebeling, F.; Chamoun, M.; Steffen, J.; Wagner, J.; Fuhrmann, M. Long-Term In Vivo Imaging of Dendritic Spines in the Hippocampus Reveals Structural Plasticity. J. Neurosci. 2014, 34, 13948–13953. [Google Scholar] [CrossRef] [PubMed]
- Lecoq, J.; Orlova, N.; Grewe, B.F. Wide. Fast. Deep: Recent Advances in Multiphoton Microscopy of In Vivo Neuronal Activity. J. Neurosci. 2019, 39, 9042–9052. [Google Scholar] [CrossRef] [PubMed]
- Wekselblatt, J.B.; Flister, E.D.; Piscopo, D.M.; Niell, C.M. Large-scale imaging of cortical dynamics during sensory perception and behavior. J. Neurophysiol. 2016, 115, 2852–2866. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, K.K.; Burns, L.D.; Cocker, E.D.; Nimmerjahn, A.; Ziv, Y.; Gamal, A.E.; Schnitzer, M.J. Miniaturized integration of a fluorescence microscope. Nat. Methods 2011, 8, 871–878. [Google Scholar] [CrossRef]
- Ouzounov, D.G.; Wang, T.; Wang, M.; Feng, D.D.; Horton, N.G.; Cruz-Hernández, J.C.; Cheng, Y.-T.; Reimer, J.; Tolias, A.S.; Nishimura, N.; et al. In vivo three-photon imaging of activity of GCaMP6-labeled neurons deep in intact mouse brain. Nat. Methods 2017, 14, 388–390. [Google Scholar] [CrossRef]
- McHenry, J.A.; Otis, J.M.; Rossi, M.A.; Robinson, J.E.; Kosyk, O.; Miller, N.W.; McElligott, Z.A.; Budygin, E.A.; Rubinow, D.R.; Stuber, G.D. Hormonal gain control of a medial preoptic area social reward circuit. Nat. Neurosci. 2017, 20, 449–458. [Google Scholar] [CrossRef]
- Wu, J.; Liang, Y.; Chen, S.; Hsu, C.-L.; Chavarha, M.; Evans, S.W.; Shi, D.; Lin, M.Z.; Tsia, K.K.; Ji, N. Kilohertz two-photon fluorescence microscopy imaging of neural activity in vivo. Nat. Methods 2020, 17, 287–290. [Google Scholar] [CrossRef]
- Ohkura, M.; Matsuzaki, M.; Kasai, H.; Imoto, K.; Nakai, J. Genetically Encoded Bright Ca2+ Probe Applicable for Dynamic Ca2+ Imaging of Dendritic Spines. Anal. Chem. 2005, 77, 5861–5869. [Google Scholar] [CrossRef]
- Akerboom, J.; Carreras Calderón, N.; Tian, L.; Wabnig, S.; Prigge, M.; Tolö, J.; Gordus, A.; Orger, M.B.; Severi, K.E.; Macklin, J.J.; et al. Genetically Encoded Calcium Indicators for Multi-Color Neural Activity Imaging and Combination with Optogenetics. Available online: https://pubmed.ncbi.nlm.nih.gov/23459413/ (accessed on 4 December 2020).
- Jing, M.; Li, Y.; Zeng, J.; Huang, P.; Skirzewski, M.; Kljakic, O.; Peng, W.; Qian, T.; Tan, K.; Zou, J.; et al. An optimized acetylcholine sensor for monitoring in vivo cholinergic activity. Nat. Methods 2020, 17, 1139–1146. [Google Scholar] [CrossRef]
- Jing, M.; Zhang, P.; Wang, G.; Feng, J.; Mesik, L.; Zeng, J.; Jiang, H.; Wang, S.; Looby, J.C.; Guagliardo, N.A.; et al. A genetically encoded fluorescent acetylcholine indicator for in vitro and in vivo studies. Nat. Biotechnol. 2018, 36, 726–737. [Google Scholar] [CrossRef] [PubMed]
- Bedbrook, C.N.; Yang, K.K.; Robinson, J.E.; Mackey, E.D.; Gradinaru, V.; Arnold, F.H. Machine learning-guided channelrhodopsin engineering enables minimally invasive optogenetics. Nat. Methods 2019, 16, 1176–1184. [Google Scholar] [CrossRef]
- Senior, A.W.; Evans, R.; Jumper, J.; Kirkpatrick, J.; Sifre, L.; Green, T.; Qin, C.; Žídek, A.; Nelson, A.W.R.; Bridgland, A.; et al. Improved protein structure prediction using potentials from deep learning. Nature 2020, 577, 706–710. [Google Scholar] [CrossRef] [PubMed]
- Okumoto, S.; Looger, L.L.; Micheva, K.D.; Reimer, R.J.; Smith, S.J.; Frommer, W.B. Detection of glutamate release from neurons by genetically encoded surface-displayed FRET nanosensors. Proc. Natl. Acad. Sci. USA 2005, 102, 8740–8745. [Google Scholar] [CrossRef]
- Tsien, R.Y. Building and breeding molecules to spy on cells and tumors. FEBS Lett. 2005, 579, 927–932. [Google Scholar] [CrossRef]
- Hires, S.A.; Zhu, Y.; Tsien, R.Y. Optical measurement of synaptic glutamate spillover and reuptake by linker optimized glutamate-sensitive fluorescent reporters. Proc. Natl. Acad. Sci. USA 2008, 105, 4411–4416. [Google Scholar] [CrossRef] [PubMed]
- Marvin, J.S.; Borghuis, B.G.; Tian, L.; Cichon, J.; Harnett, M.T.; Akerboom, J.; Gordus, A.; Renninger, S.L.; Chen, T.-W.; Bargmann, C.I.; et al. An optimized fluorescent probe for visualizing glutamate neurotransmission. Nat. Methods 2013, 10, 162–170. [Google Scholar] [CrossRef]
- Wu, J.; Abdelfattah, A.S.; Zhou, H.; Ruangkittisakul, A.; Qian, Y.; Ballanyi, K.; Campbell, R.E. Genetically Encoded Glutamate Indicators with Altered Color and Topology. ACS Chem. Biol. 2018, 13, 1832–1837. [Google Scholar] [CrossRef] [PubMed]
- Marvin, J.S.; Shimoda, Y.; Magloire, V.; Leite, M.; Kawashima, T.; Jensen, T.P.; Kolb, I.; Knott, E.L.; Novak, O.; Podgorski, K.; et al. A genetically encoded fluorescent sensor for in vivo imaging of GABA. Nat. Methods 2019, 16, 763–770. [Google Scholar] [CrossRef]
- Borden, P.M.; Zhang, P.; Shivange, A.V.; Marvin, J.S.; Cichon, J.; Dan, C.; Podgorski, K.; Figueiredo, A.; Novak, O.; Tanimoto, M.; et al. A Fast Genetically Encoded Fluorescent Sensor for Faithful In Vivo Acetylcholine Detection in Mice, Fish, Worms and Flies; Social Science Research Network: Rochester, NY, USA, 2020. [Google Scholar]
- Unger, E.K.; Keller, J.P.; Altermatt, M.; Liang, R.; Matsui, A.; Dong, C.; Hon, O.J.; Yao, Z.; Sun, J.; Banala, S.; et al. Directed Evolution of a Selective and Sensitive Serotonin Sensor via Machine Learning. Cell 2020, 183, 1986–2002.e26. [Google Scholar] [CrossRef]
- Vilardaga, J.-P.; Bünemann, M.; Krasel, C.; Castro, M.; Lohse, M.J. Measurement of the millisecond activation switch of G protein-coupled receptors in living cells. Nat. Biotechnol. 2003, 21, 807–812. [Google Scholar] [CrossRef]
- Patriarchi, T.; Cho, J.R.; Merten, K.; Howe, M.W.; Marley, A.; Xiong, W.-H.; Folk, R.W.; Broussard, G.J.; Liang, R.; Jang, M.J.; et al. Ultrafast neuronal imaging of dopamine dynamics with designed genetically encoded sensors. Science 2018, 360. [Google Scholar] [CrossRef]
- Patriarchi, T.; Mohebi, A.; Sun, J.; Marley, A.; Liang, R.; Dong, C.; Puhger, K.; Mizuno, G.O.; Davis, C.M.; Wiltgen, B.; et al. An expanded palette of dopamine sensors for multiplex imaging in vivo. Nat. Methods 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Zeng, J.; Jing, M.; Zhou, J.; Feng, J.; Owen, S.F.; Luo, Y.; Li, F.; Wang, H.; Yamaguchi, T.; et al. A Genetically Encoded Fluorescent Sensor Enables Rapid and Specific Detection of Dopamine in Flies, Fish, and Mice. Cell 2018, 174, 481–496.e19. [Google Scholar] [CrossRef]
- Feng, J.; Zhang, C.; Lischinsky, J.E.; Jing, M.; Zhou, J.; Wang, H.; Zhang, Y.; Dong, A.; Wu, Z.; Wu, H.; et al. A Genetically Encoded Fluorescent Sensor for Rapid and Specific In Vivo Detection of Norepinephrine. Neuron 2019, 102, 745–761.e8. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Peng, W.; Li, X.; Qian, T.; Song, K.; Zeng, J.; Deng, F.; Hao, S.; Feng, J.; Zhang, P.; et al. A genetically encoded GRAB sensor for measuring serotonin dynamics in vivo. bioRxiv 2020. [Google Scholar] [CrossRef]
- Miyawaki, A.; Llopis, J.; Heim, R.; McCaffery, J.M.; Adams, J.A.; Ikura, M.; Tsien, R.Y. Fluorescent indicators for Ca 2+ based on green fluorescent proteins and calmodulin. Nature 1997, 388, 882–887. [Google Scholar] [CrossRef]
- Miyawaki, A. Development of Probes for Cellular Functions Using Fluorescent Proteins and Fluorescence Resonance Energy Transfer. Annu. Rev. Biochem. 2011, 80, 357–373. [Google Scholar] [CrossRef] [PubMed]
- Palmer, A.E.; Qin, Y.; Park, J.G.; McCombs, J.E. Design and application of genetically encoded biosensors. Trends Biotechnol. 2011, 29, 144–152. [Google Scholar] [CrossRef]
- Griesbeck, O.; Baird, G.S.; Campbell, R.E.; Zacharias, D.A.; Tsien, R.Y. Reducing the Environmental Sensitivity of Yellow Fluorescent Protein: Mechanism and applications. J. Biol. Chem. 2001, 276, 29188–29194. [Google Scholar] [CrossRef] [PubMed]
- Nagai, T.; Ibata, K.; Park, E.S.; Kubota, M.; Mikoshiba, K.; Miyawaki, A. A variant of yellow fluorescent protein with fast and efficient maturation for cell-biological applications. Nat. Biotechnol. 2002, 20, 87–90. [Google Scholar] [CrossRef]
- Nagai, T.; Yamada, S.; Tominaga, T.; Ichikawa, M.; Miyawaki, A. Expanded dynamic range of fluorescent indicators for Ca(2+) by circularly permuted yellow fluorescent proteins. Proc. Natl. Acad. Sci. USA 2004, 101, 10554–10559. [Google Scholar] [CrossRef] [PubMed]
- Lütcke, H.; Murayama, M.; Hahn, T.; Margolis, D.J.; Astori, S.; Meyer, S.; Göbel, W.; Yang, Y.; Tang, W.; Kügler, S.; et al. Optical recording of neuronal activity with a genetically-encoded calcium indicator in anesthetized and freely moving mice. Front. Neural Circuits 2010, 4. [Google Scholar] [CrossRef] [PubMed]
- Miyawaki, A.; Tsien, R.Y. Monitoring protein conformations and interactions by fluorescence resonance energy transfer between mutants of green fluorescent protein. Methods Enzymol. 2000, 327, 472–500. [Google Scholar] [PubMed]
- Horikawa, K.; Yamada, Y.; Matsuda, T.; Kobayashi, K.; Hashimoto, M.; Matsu-ura, T.; Miyawaki, A.; Michikawa, T.; Mikoshiba, K.; Nagai, T. Spontaneous network activity visualized by ultrasensitive Ca2+ indicators, yellow Cameleon-Nano. Nat. Methods 2010, 7, 729–732. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Michikawa, T.; Hashimoto, M.; Horikawa, K.; Nagai, T.; Miyawaki, A.; Häusser, M.; Mikoshiba, K. Quantitative comparison of genetically encoded Ca indicators in cortical pyramidal cells and cerebellar Purkinje cells. Front. Cell Neurosci. 2011, 5, 18. [Google Scholar] [CrossRef]
- Bajar, B.T.; Wang, E.S.; Lam, A.J.; Kim, B.B.; Jacobs, C.L.; Howe, E.S.; Davidson, M.W.; Lin, M.Z.; Chu, J. Improving brightness and photostability of green and red fluorescent proteins for live cell imaging and FRET reporting. Sci. Rep. 2016, 6, 20889. [Google Scholar] [CrossRef] [PubMed]
- Shemetov, A.A.; Monakhov, M.V.; Zhang, Q.; Canton-Josh, J.E.; Kumar, M.; Chen, M.; Matlashov, M.E.; Li, X.; Yang, W.; Nie, L.; et al. A near-infrared genetically encoded calcium indicator for in vivo imaging. Nat. Biotechnol. 2021, 39, 368–377. [Google Scholar] [CrossRef]
- Palmer, A.E.; Giacomello, M.; Kortemme, T.; Hires, S.A.; Lev-Ram, V.; Baker, D.; Tsien, R.Y. Ca2+ Indicators Based on Computationally Redesigned Calmodulin-Peptide Pairs. Chem. Biol. 2006, 13, 521–530. [Google Scholar] [CrossRef]
- Heim, N.; Griesbeck, O. Genetically Encoded Indicators of Cellular Calcium Dynamics Based on Troponin C and Green Fluorescent Protein. J. Biol. Chem. 2004, 279, 14280–14286. [Google Scholar] [CrossRef]
- Mank, M.; Reiff, D.F.; Heim, N.; Friedrich, M.W.; Borst, A.; Griesbeck, O. A FRET-Based Calcium Biosensor with Fast Signal Kinetics and High Fluorescence Change. Biophys. J. 2006, 90, 1790–1796. [Google Scholar] [CrossRef] [PubMed]
- Mank, M.; Santos, A.F.; Direnberger, S.; Mrsic-Flogel, T.D.; Hofer, S.B.; Stein, V.; Hendel, T.; Reiff, D.F.; Levelt, C.; Borst, A.; et al. A genetically encoded calcium indicator for chronic in vivo two-photon imaging. Nat. Methods 2008, 5, 805–811. [Google Scholar] [CrossRef]
- Thestrup, T.; Litzlbauer, J.; Bartholomäus, I.; Mues, M.; Russo, L.; Dana, H.; Kovalchuk, Y.; Liang, Y.; Kalamakis, G.; Laukat, Y.; et al. Optimized ratiometric calcium sensors for functional in vivo imaging of neurons and T lymphocytes. Nat. Methods 2014, 11, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Nakai, J.; Ohkura, M.; Imoto, K. A high signal-to-noise Ca(2+) probe composed of a single green fluorescent protein. Nat. Biotechnol. 2001, 19, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Tallini, Y.N.; Ohkura, M.; Choi, B.-R.; Ji, G.; Imoto, K.; Doran, R.; Lee, J.; Plan, P.; Wilson, J.; Xin, H.-B.; et al. Imaging cellular signals in the heart in vivo: Cardiac expression of the high-signal Ca2+ indicator GCaMP2. Proc. Natl. Acad. Sci. USA 2006, 103, 4753–4758. [Google Scholar] [CrossRef] [PubMed]
- Muto, A.; Ohkura, M.; Kotani, T.; Higashijima, S.; Nakai, J.; Kawakami, K. Genetic visualization with an improved GCaMP calcium indicator reveals spatiotemporal activation of the spinal motor neurons in zebrafish. Proc. Natl. Acad. Sci. USA 2011, 108, 5425–5430. [Google Scholar] [CrossRef]
- Sun, X.R.; Badura, A.; Pacheco, D.A.; Lynch, L.A.; Schneider, E.R.; Taylor, M.P.; Hogue, I.B.; Enquist, L.W.; Murthy, M.; Wang, S.S.-H. Fast GCaMPs for improved tracking of neuronal activity. Nat. Commun. 2013, 4, 2170. [Google Scholar] [CrossRef]
- Akerboom, J.; Chen, T.-W.; Wardill, T.J.; Tian, L.; Marvin, J.S.; Mutlu, S.; Calderón, N.C.; Esposti, F.; Borghuis, B.G.; Sun, X.R.; et al. Optimization of a GCaMP Calcium Indicator for Neural Activity Imaging. J. Neurosci. 2012, 32, 13819–13840. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.-W.; Wardill, T.J.; Sun, Y.; Pulver, S.R.; Renninger, S.L.; Baohan, A.; Schreiter, E.R.; Kerr, R.A.; Orger, M.B.; Jayaraman, V.; et al. Ultrasensitive fluorescent proteins for imaging neuronal activity. Nature 2013, 499, 295–300. [Google Scholar] [CrossRef]
- Dana, H.; Sun, Y.; Mohar, B.; Hulse, B.K.; Kerlin, A.M.; Hasseman, J.P.; Tsegaye, G.; Tsang, A.; Wong, A.; Patel, R.; et al. High-performance calcium sensors for imaging activity in neuronal populations and microcompartments. Nat. Methods 2019, 16, 649–657. [Google Scholar] [CrossRef]
- Zhang, Y.; Rózsa, M.; Bushey, D.; Zheng, J.; Reep, D.; Liang, Y.; Broussard, G.J.; Tsang, A.; Tsegaye, G.; Patel, R.; et al. jGCaMP8 Fast Genetically Encoded Calcium Indicators. Janelia Res. Campus 2020. [Google Scholar] [CrossRef]
- Zhao, Y.; Araki, S.; Wu, J.; Teramoto, T.; Chang, Y.-F.; Nakano, M.; Abdelfattah, A.S.; Fujiwara, M.; Ishihara, T.; Nagai, T.; et al. An Expanded Palette of Genetically Encoded Ca2+ Indicators. Science 2011, 333, 1888–1891. [Google Scholar] [CrossRef]
- Qian, Y.; Piatkevich, K.D.; Mc Larney, B.; Abdelfattah, A.S.; Mehta, S.; Murdock, M.H.; Gottschalk, S.; Molina, R.S.; Zhang, W.; Chen, Y.; et al. A genetically encoded near-infrared fluorescent calcium ion indicator. Nat. Methods 2019, 16, 171–174. [Google Scholar] [CrossRef]
- Inoue, M.; Takeuchi, A.; Manita, S.; Horigane, S.; Sakamoto, M.; Kawakami, R.; Yamaguchi, K.; Otomo, K.; Yokoyama, H.; Kim, R.; et al. Rational Engineering of XCaMPs, a Multicolor GECI Suite for In Vivo Imaging of Complex Brain Circuit Dynamics. Cell 2019, 177, 1346–1360.e24. [Google Scholar] [CrossRef]
- Baird, G.S.; Zacharias, D.A.; Tsien, R.Y. Circular permutation and receptor insertion within green fluorescent proteins. Proc. Natl. Acad. Sci. USA 1999, 96, 11241–11246. [Google Scholar] [CrossRef] [PubMed]
- Nagai, T.; Sawano, A.; Park, E.S.; Miyawaki, A. Circularly permuted green fluorescent proteins engineered to sense Ca2+. Proc. Natl. Acad. Sci. USA 2001, 98, 3197–3202. [Google Scholar] [CrossRef]
- Souslova, E.A.; Belousov, V.V.; Lock, J.G.; Strömblad, S.; Kasparov, S.; Bolshakov, A.P.; Pinelis, V.G.; Labas, Y.A.; Lukyanov, S.; Mayr, L.M.; et al. Single fluorescent protein-based Ca2+ sensors with increased dynamic range. BMC Biotechnol. 2007, 7, 37. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Hires, S.A.; Mao, T.; Huber, D.; Chiappe, M.E.; Chalasani, S.H.; Petreanu, L.; Akerboom, J.; McKinney, S.A.; Schreiter, E.R.; et al. Imaging neural activity in worms, flies and mice with improved GCaMP calcium indicators. Nat. Methods 2009, 6, 875–881. [Google Scholar] [CrossRef] [PubMed]
- Fosque, B.F.; Sun, Y.; Dana, H.; Yang, C.-T.; Ohyama, T.; Tadross, M.R.; Patel, R.; Zlatic, M.; Kim, D.S.; Ahrens, M.B.; et al. Labeling of active neural circuits in vivo with designed calcium integrators. Science 2015, 347, 755–760. [Google Scholar] [CrossRef]
- Moeyaert, B.; Holt, G.; Madangopal, R.; Perez-Alvarez, A.; Fearey, B.C.; Trojanowski, N.F.; Ledderose, J.; Zolnik, T.A.; Das, A.; Patel, D.; et al. Improved methods for marking active neuron populations. Nat. Commun. 2018, 9, 4440. [Google Scholar] [CrossRef] [PubMed]
- Edwards, K.A.; Hoppa, M.B.; Bosco, G. The Photoconvertible Fluorescent Probe, CaMPARI, Labels Active Neurons in Freely-Moving Intact Adult Fruit Flies. Front. Neural Circuits 2020, 14. [Google Scholar] [CrossRef]
- Trojanowski, N.F.; Bottorff, J.; Turrigiano, G.G. Activity labeling in vivo using CaMPARI2 reveals intrinsic and synaptic differences between neurons with high and low firing rate set points. Neuron 2021, 109, 663–676.e5. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wildes, C.P.; Pattarabanjird, T.; Sanchez, M.I.; Glober, G.F.; Matthews, G.A.; Tye, K.M.; Ting, A.Y. A light- and calcium-gated transcription factor for imaging and manipulating activated neurons. Nat. Biotechnol. 2017, 35, 864–871. [Google Scholar] [CrossRef] [PubMed]
- Sabatini, B.L.; Tian, L. Imaging Neurotransmitter and Neuromodulator Dynamics In Vivo with Genetically Encoded Indicators. Neuron 2020, 108, 17–32. [Google Scholar] [CrossRef]
- Wang, J.W.; Wong, A.M.; Flores, J.; Vosshall, L.B.; Axel, R. Two-Photon Calcium Imaging Reveals an Odor-Evoked Map of Activity in the Fly Brain. Cell 2003, 112, 271–282. [Google Scholar] [CrossRef]
- Peled, E.S.; Isacoff, E.Y. Optical quantal analysis of synaptic transmission in wild-type and rab3-mutant Drosophila melanogaster motor axons. Nat. Neurosci. 2011, 14, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, V.; Ji, N.; Wang, X.; Clark, C.; Mitchell, J.K.; Klein, M.; Tabone, C.J.; Florman, J.; Ji, H.; Greenwood, J.; et al. Pan-neuronal imaging in roaming Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2016, 113, E1082–E1088. [Google Scholar] [CrossRef]
- Ahrens, M.B.; Li, J.M.; Orger, M.B.; Robson, D.N.; Schier, A.F.; Engert, F.; Portugues, R. Brain-wide neuronal dynamics during motor adaptation in zebrafish. Nature 2012, 485, 471–477. [Google Scholar] [CrossRef]
- Peters, A.J.; Chen, S.X.; Komiyama, T. Emergence of reproducible spatiotemporal activity during motor learning. Nature 2014, 510, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Lovett-Barron, M.; Kaifosh, P.; Kheirbek, M.A.; Danielson, N.; Zaremba, J.D.; Reardon, T.R.; Turi, G.F.; Hen, R.; Zemelman, B.V.; Losonczy, A. Dendritic inhibition in the hippocampus supports fear learning. Science 2014, 343, 857–863. [Google Scholar] [CrossRef]
- Sun, W.; Tan, Z.; Mensh, B.D.; Ji, N. Thalamus provides layer 4 of primary visual cortex with orientation- and direction-tuned inputs. Nat. Neurosci. 2016, 19, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Melom, J.E.; Akbergenova, Y.; Gavornik, J.P.; Littleton, J.T. Spontaneous and Evoked Release Are Independently Regulated at Individual Active Zones. J. Neurosci. 2013, 33, 17253–17263. [Google Scholar] [CrossRef]
- Cichon, J.; Gan, W.-B. Branch-specific dendritic Ca2+ spikes cause persistent synaptic plasticity. Nature 2015, 520, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Sheffield, M.E.J.; Dombeck, D.A. Calcium transient prevalence across the dendritic arbour predicts place field properties. Nature 2015, 517, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Sofroniew, N.J.; Flickinger, D.; King, J.; Svoboda, K. A large field of view two-photon mesoscope with subcellular resolution for in vivo imaging. eLife 2016, 5, e14472. [Google Scholar] [CrossRef] [PubMed]
- Peron, S.P.; Freeman, J.; Iyer, V.; Guo, C.; Svoboda, K. A Cellular Resolution Map of Barrel Cortex Activity during Tactile Behavior. Neuron 2015, 86, 783–799. [Google Scholar] [CrossRef] [PubMed]
- Stirman, J.N.; Smith, I.T.; Kudenov, M.W.; Smith, S.L. Wide field-of-view, multi-region, two-photon imaging of neuronal activity in the mammalian brain. Nat. Biotechnol. 2016, 34, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Wallace, D.J.; Meyer zum Alten Borgloh, S.; Astori, S.; Yang, Y.; Bausen, M.; Kügler, S.; Palmer, A.E.; Tsien, R.Y.; Sprengel, R.; Kerr, J.N.D.; et al. Single-spike detection in vitro and in vivo with a genetic Ca2+ sensor. Nat. Methods 2008, 5, 797–804. [Google Scholar] [CrossRef]
- Hendel, T.; Mank, M.; Schnell, B.; Griesbeck, O.; Borst, A.; Reiff, D.F. Fluorescence Changes of Genetic Calcium Indicators and OGB-1 Correlated with Neural Activity and Calcium In Vivo and In Vitro. J. Neurosci. 2008, 28, 7399–7411. [Google Scholar] [CrossRef]

| Genetically Encoded Neurotransmitter Indicator | Ligand | Reporter Element | Dynamic Range ΔR/R0 or ΔF/F0 (In Vitro Unless Otherwise Noted) | Affinity (Kd) (In Vitro Unless Otherwise Noted) | On|Off Kinetics |
|---|---|---|---|---|---|
| PBP-based neurotransmitter indicators | |||||
| FLIPE [26] | Glutamate | CFP/Venus | ND | 0.6 µM | kon = 10.0 × 107 M−1s−1|koff = 60 s−1 |
| GluSnFR [27] | Glutamate | CFP/YFP | 0.07 | 150 nM | ND |
| SuperGluSnFR [28] | Glutamate | CFP/Citrine | 0.44 | 2.5 µM | kon = 3.0 × 107 M−1s−1|koff = 75 s−1 |
| iGluSnFR [29] | Glutamate | cpEGFP | 4.5 | 110 µM | τon = ~5 ms|τoff = ~92 ms |
| R-iGluSnFR [30] | Glutamate | cpmApple | 4.9 | 11 µM | ND |
| iGABASnFR [31] | GABA | cpSFGFP | 2.5 | 9 µM | ND |
| iArchSnFR [32] | Acetylcholine | cpSFGFP | 12 | 1.3 µM | τon = ~80 ms|τoff = 1.9 s |
| iSeroSnFR [33] | Serotonin | cpSFGFP | 8 | 310 µM | τon = ~0.5-10 ms (fast), ~5-18 s (slow)|τoff = ~4 ms (fast), ~150 ms (slow) |
| GPCR-based neurotransmitter indicators | |||||
| α2AAR-cam [34] | Norepinephrine | CFP/YFP | −0.05 | 17 nM | τon = 40 ms |
| dLight 1.1 [35] | Dopamine | cpGFP | 2.3 | 330 nM | τon = 10 ms|τoff = 100 ms |
| dLight 1.2 [35] | Dopamine | cpGFP | 3.4 | 770 nM | τon = 10 ms|τoff = 100 ms |
| dLight 1.3a [35] | Dopamine | cpGFP | 6.6 | 2.3 µM | τon = 10 ms|τoff = 100 ms |
| dLight 1.3b [35] | Dopamine | cpGFP | 9.3 | 1.7 µM | τon = 10 ms|τoff = 100 ms |
| dLight 1.4 [35] | Dopamine | cpGFP | 1.7 | 4 nM | τon = 10 ms|τoff = 100 ms |
| YdLight 1.1 [36] | Dopamine | cpGFP V203Y/S72A | 3.06 | 1.63 µM | ND/ND |
| RdLight 1 [36] | Dopamine | cpmApple | 2.48 | 859 nM | τon = 14.1 ms|τoff = 0.398 s |
| GRABDA1m [37] | Dopamine | cpEGFP | 0.9 | 130 nM | τon = 60 ms|τoff = 920 ms |
| GRABDA1h [37] | Dopamine | cpEGFP | 0.9 | 10 nM | τon = 130 ms|τoff = 1.9 s |
| GRABNE1m [38] | Norepinephrine | cpEGFP | 2.3 | 930 nM | τon = 72 ms|τoff = 680 ms |
| GRABNE1h [38] | Norepinephrine | cpEGFP | 1.3 | 83 nM | τon = 36 ms|τoff = 1890 ms |
| GACH 2.0 [23] | Acetylcholine | cpEGFP | 0.76 | 2 µM | τon = 280 ms|τoff = 762 ms |
| GACH 3.0 [22] | Acetylcholine | cpEGFP | 2.8 | 2 µM | τon = 105 ms|τoff = 3.7 s |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bi, X.; Beck, C.; Gong, Y. Genetically Encoded Fluorescent Indicators for Imaging Brain Chemistry. Biosensors 2021, 11, 116. https://doi.org/10.3390/bios11040116
Bi X, Beck C, Gong Y. Genetically Encoded Fluorescent Indicators for Imaging Brain Chemistry. Biosensors. 2021; 11(4):116. https://doi.org/10.3390/bios11040116
Chicago/Turabian StyleBi, Xiaoke, Connor Beck, and Yiyang Gong. 2021. "Genetically Encoded Fluorescent Indicators for Imaging Brain Chemistry" Biosensors 11, no. 4: 116. https://doi.org/10.3390/bios11040116
APA StyleBi, X., Beck, C., & Gong, Y. (2021). Genetically Encoded Fluorescent Indicators for Imaging Brain Chemistry. Biosensors, 11(4), 116. https://doi.org/10.3390/bios11040116