



Microbial Community Dynamics and Biogeochemical Cycling in Microplastic-Contaminated Sediment

Abstract

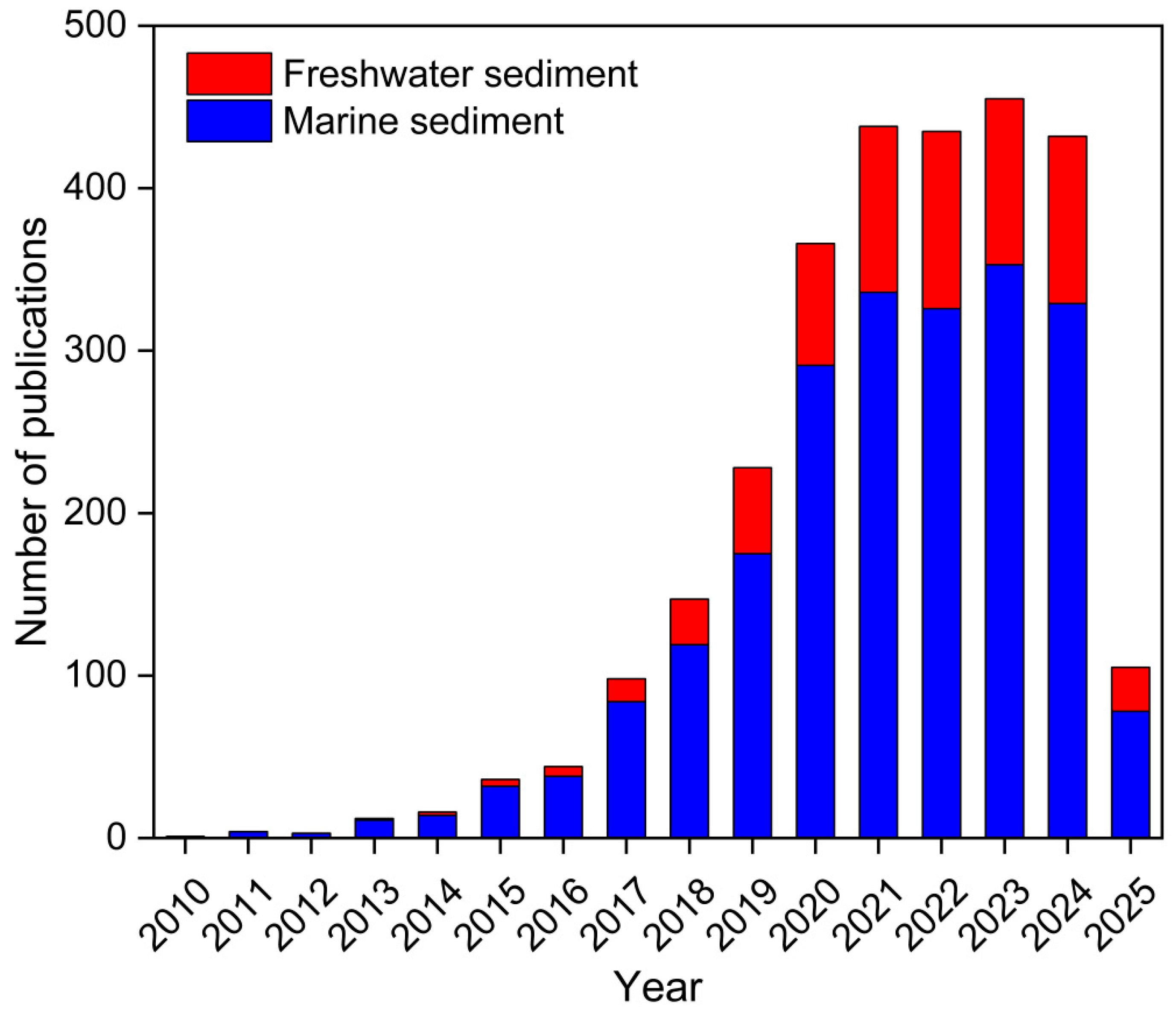
1. Introduction
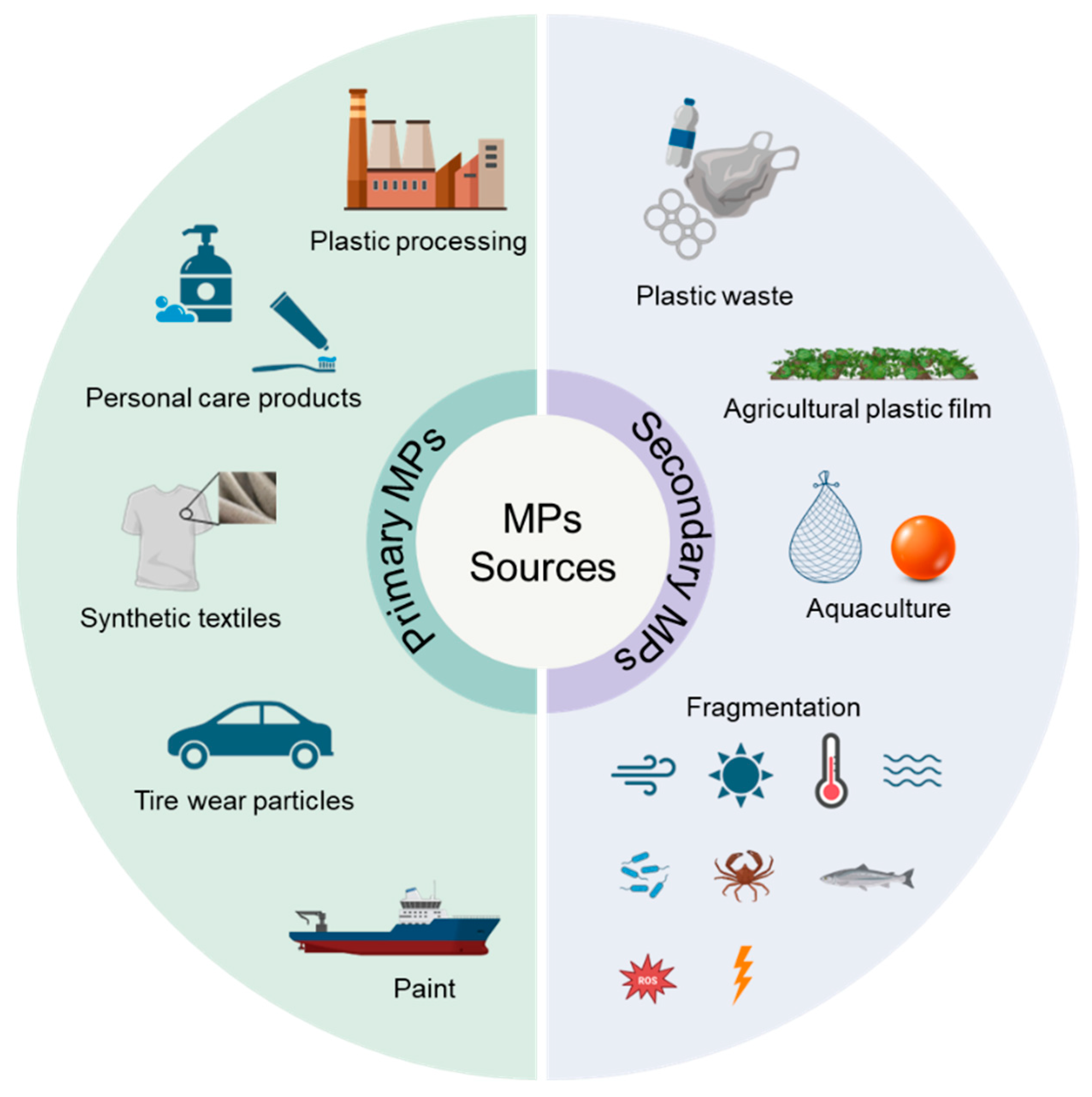
2. MPs in Sediments
3. Effects of MPs on Microbial Community in Sediments
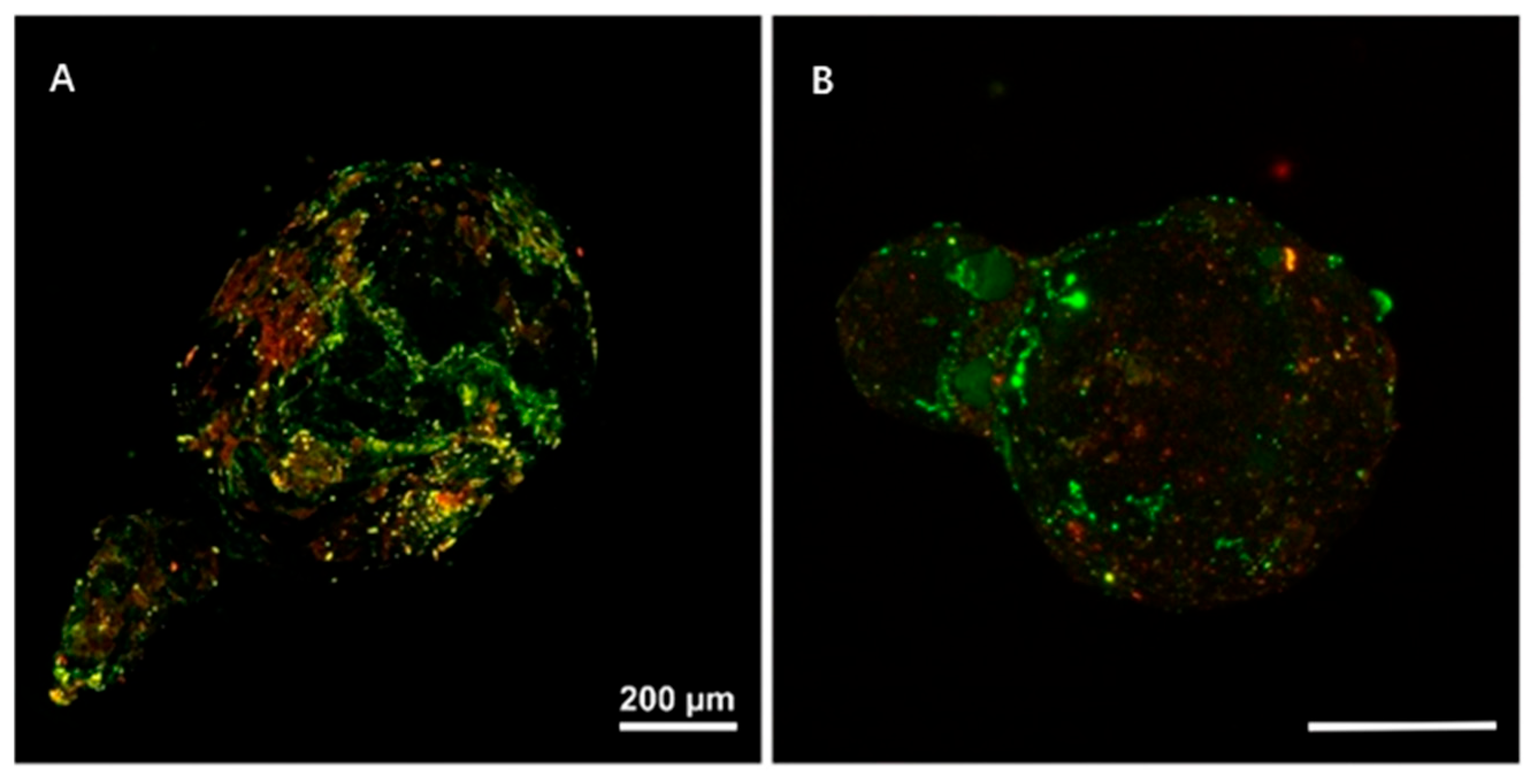
3.1. The Colonization of Microorganisms on the Surface of MPs
3.2. Microbial Diversity and Dominant Bacterial Phyla Affected by MPs
3.3. Effects of MPs on Microbial Functions
3.3.1. Nitrification and Denitrification
3.3.2. Nitrogen Fixation
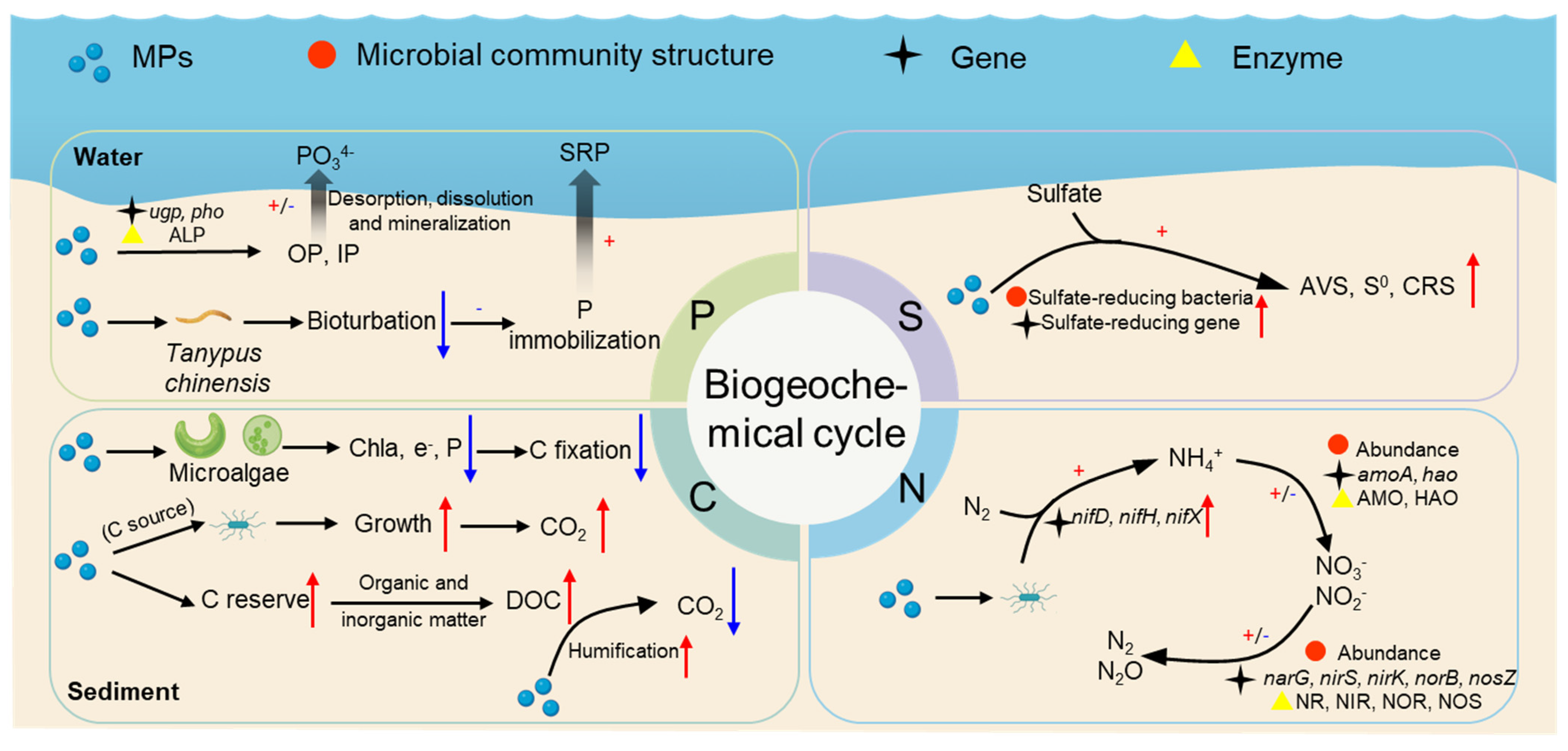
4. Effects of MPs on Biogeochemical Cycling in Sedimentary Environments
4.1. Nitrogen Cycle
4.2. Carbon Cycle
4.3. Phosphorus Cycling
4.4. Sulfur Cycling
5. Conclusions and Outlook
- (1)
- Because research content is trivial and the influencing factors are complex, it is challenging to establish a universal and common law. Discrepancies in the effects of environmental factors and different MP types on microbial communities still need to be further explored.
- (2)
- Current research is still focused on the short-term effect of MPs on microorganisms; however, the aging of MPs will lead to large changes in their properties. The attachment of biofilm can enhance their electronegativity. In addition, the photoaging of MPS increases numbers of oxygen-containing functional groups on the polymer molecular chain and also increases hydrophilicity. The physicochemical properties of MPs undergo significant changes over time, leading to alterations in their environmental behavior and biological toxicity in aquatic and sedimentary environments. These dynamic transformations are worth exploring further in future research. The following summarizes methods of research on MP aging:
- Laboratory-based aging simulations can be used to simulate natural environmental conditions over extended periods (e.g., UV radiation, temperature fluctuations and eco-corona).
- Long-term in situ aging of MPs is carried out in a real environment (e.g., marine and freshwater sediments).
- (3)
- There is a relative lack of research on the effects of MPs on biogeochemical cycles in sedimentary environments, especially phosphorus and sulfur cycles. Therefore, further studies are necessary to elucidate the response of elemental cycles after MP pollution in sedimentary environments, and to assess the broader implications for aquatic ecosystems and human health. Research gaps with regard to phosphorus and sulfur cycles might be attributed to an absence of standardized indicators. Future research can focus on the following crucial microbiota relevant to phosphorus and sulfur cycles: phosphate-solubilizing bacteria (PSB); phosphate-accumulating organisms (PAOs); sulfate-reducing bacteria (SRB); sulfur-oxidizing bacteria (SOB); and sulfur-disproportionating bacteria.
Funding
Acknowledgments
Conflicts of Interest
References
- Shen, M.; Song, B.; Zeng, G.; Zhang, Y.; Huang, W.; Wen, X.; Tang, W. Are biodegradable plastics a promising solution to solve the global plastic pollution? Environ. Pollut. 2020, 263, 114469. [Google Scholar] [CrossRef] [PubMed]
- Mao, R.; Lang, M.; Yu, X.; Wu, R.; Yang, X.; Guo, X. Aging mechanism of microplastics with UV irradiation and its effects on the adsorption of heavy metals. J. Hazard. Mater. 2020, 393, 122515. [Google Scholar] [CrossRef]
- Hodgson, D.J.; Bréchon, A.L.; Thompson, R.C. Ingestion and fragmentation of plastic carrier bags by the amphipod Orchestia gammarellus: Effects of plastic type and fouling load. Mar. Pollut. Bull. 2018, 127, 154–159. [Google Scholar] [CrossRef]
- Thompson, R.C.; Courtene-Jones, W.; Boucher, J.; Pahl, S.; Raubenheimer, K.; Koelmans, A.A. Twenty years of microplastic pollution research—What have we learned? Science 2024, 386, eadl2746. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Yang, Q.; Jiang, J.; Dalu, T.; Kadushkin, A.; Singh, J.; Fakhrullin, R.; Wang, F.; Cai, X.; Li, R. Coronas of micro/nano plastics: A key determinant in their risk assessments. Part. Fibre Toxicol. 2022, 19, 55. [Google Scholar] [CrossRef]
- Eriksen, M.; Lebreton, L.C.; Carson, H.S.; Thiel, M.; Moore, C.J.; Borerro, J.C.; Galgani, F.; Ryan, P.G.; Reisser, J. Plastic pollution in the world’s oceans: More than 5 trillion plastic pieces weighing over 250,000 tons afloat at sea. PLoS ONE 2014, 9, e111913. [Google Scholar] [CrossRef]
- Boucher, C.; Morin, M.; Bendell, L.I. The influence of cosmetic microbeads on the sorptive behavior of cadmium and lead within intertidal sediments: A laboratory study. Reg. Stud. Mar. Sci. 2016, 3, 1–7. [Google Scholar] [CrossRef]
- Kuypers, M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef]
- Wu, H.; Cui, H.; Fu, C.; Li, R.; Qi, F.; Liu, Z.; Yang, G.; Xiao, K.; Qiao, M. Unveiling the crucial role of soil microorganisms in carbon cycling: A review. Sci. Total Environ. 2024, 909, 168627. [Google Scholar] [CrossRef]
- Wang, F.; Bai, Y.; Yang, F.; Zhu, Q.; Zhao, Q.; Zhang, X.; Wei, Y.; Liao, H. Degradation of nitrogen, phosphorus, and organic matter in urban river sediments by adding microorganisms. Sustainability 2021, 13, 2580. [Google Scholar] [CrossRef]
- Vijayanand, M.; Ramakrishnan, A.; Subramanian, R.; Issac, P.K.; Nasr, M.; Khoo, K.S.; Rajagopal, R.; Greff, B.; Azelee, N.I.W.; Jeon, B.; et al. Polyaromatic hydrocarbons (PAHs) in the water environment: A review on toxicity, microbial biodegradation, systematic biological advancements, and environmental fate. Environ. Res. 2023, 227, 115716. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Zhu, Y.; Zhang, Y.; Zeng, G.; Wen, X.; Yi, H.; Ye, S.; Ren, X.; Song, B. Micro (nano) plastics: Unignorable vectors for organisms. Mar. Pollut. Bull. 2019, 139, 328–331. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Nie, F.; Zhu, D.; Wei, Y.; Zhang, M.; Hu, Y.; Chen, M.; Kang, D.; Chen, Z.; Lin, H.; et al. Microplastic pollution and its impact on marine microbes in Zhanjiang, China. J. Coast. Conserv. 2022, 26, 66. [Google Scholar] [CrossRef]
- Li, W.; Wang, Z.; Li, W.; Li, Z. Impacts of microplastics addition on sediment environmental properties, enzymatic activities and bacterial diversity. Chemosphere 2022, 307, 135836. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Li, X.; Feng, Z.; Xiao, M.; Ge, T.; Li, Y.; Yao, H. Polyethylene microplastics alter the microbial functional gene abundances and increase nitrous oxide emissions from paddy soils. J. Hazard. Mater. 2022, 432, 128721. [Google Scholar] [CrossRef]
- Yin, M.; Yan, B.; Wang, H.; Wu, Y.; Wang, X.; Wang, J.; Zhu, Z.; Yan, X.; Liu, Y.; Liu, M.; et al. Effects of microplastics on nitrogen and phosphorus cycles and microbial communities in sediments. Environ. Pollut. 2023, 318, 120852. [Google Scholar] [CrossRef]
- Wang, X.; Li, J.; Wang, D.; Sun, C.; Zhang, X.; Zhao, J.; Teng, J.; Wang, Q. Unveiling microplastic’s role in nitrogen cycling: Metagenomic insights from estuarine sediment microcosms. Environ. Pollut. 2024, 359, 124591. [Google Scholar] [CrossRef]
- Seeley, M.E.; Song, B.; Passie, R.; Hale, R.C. Microplastics affect sedimentary microbial communities and nitrogen cycling. Nat. Commun. 2020, 11, 2372. [Google Scholar] [CrossRef]
- Wang, H.; Yang, Q.; Li, D.; Wu, J.; Yang, S.; Deng, Y.; Luo, C.; Jia, W.; Zhong, Y.; Peng, P. Stable isotopic and metagenomic analyses reveal microbial-mediated effects of microplastics on sulfur cycling in coastal sediments. Environ. Sci. Technol. 2023, 57, 1167–1176. [Google Scholar] [CrossRef]
- Li, J.; Liu, H.; Chen, J.P. Microplastics in freshwater systems: A review on occurrence, environmental effects, and methods for microplastics detection. Water Res. 2018, 137, 362–374. [Google Scholar] [CrossRef]
- Lin, L.; Zuo, L.; Peng, J.; Cai, L.; Fok, L.; Yan, Y.; Li, H.; Xu, X. Occurrence and distribution of microplastics in an urban river: A case study in the Pearl River along Guangzhou City, China. Sci. Total Environ. 2018, 644, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Lin, Y.; Booth, A.M.; Song, X.; Cui, Y.; Xia, B.; Gu, Z.; Li, Y.; Liu, F.; Cai, M. Fate, source and mass budget of sedimentary microplastics in the Bohai Sea and the Yellow Sea. Environ. Pollut. 2022, 294, 118640. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, B.; Gu, C.; Shen, C.; Yin, S.; Aamir, M.; Li, F. Are we underestimating the sources of microplastic pollution in terrestrial environment? J. Hazard. Mater. 2020, 400, 123228. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Sun, Y.; Zhou, J.; Sun, W.; Shah, K.J. Microplastics in wastewater plants: A review of sources, characteristics, distribution and removal technologies. J. Contam. Hydrol. 2024, 267, 104448. [Google Scholar] [CrossRef]
- Ryberg, M.W.; Hauschild, M.Z.; Wang, F.; Averous-Monnery, S.; Laurent, A. Global environmental losses of plastics across their value chains. Resour. Conserv. Recycl. 2019, 151, 104459. [Google Scholar] [CrossRef]
- Knight, L.J.; Parker-Jurd, F.N.; Al-Sid-Cheikh, M.; Thompson, R.C. Tyre wear particles: An abundant yet widely unreported microplastic? Environ. Sci. Pollut. Res. 2020, 27, 18345–18354. [Google Scholar] [CrossRef]
- Belioka, M.; Achilias, D.S. The effect of weathering conditions in combination with natural phenomena/disasters on microplastics’ transport from aquatic environments to agricultural soils. Microplastics 2024, 3, 518–538. [Google Scholar] [CrossRef]
- Song, Y.K.; Kim, T.; Shim, W.J.; Hong, S.H.; Im, D. Microplastic emissions from fishing ropes: Quantification, characteristics, and implications for marine pollution. Mar. Pollut. Bull. 2025, 217, 118049. [Google Scholar] [CrossRef]
- Yu, F.; Pei, Y.; Zhang, X.; Wu, X.; Zhang, G.; Ma, J. Occurrence and distribution characteristics of aged microplastics in the surface water, sediment, and crabs of the aquaculture pond in the Yangtze River Delta of China. Sci. Total Environ. 2023, 871, 162039. [Google Scholar] [CrossRef]
- Shi, C.; Zhang, Y.; Shao, Y.; Ray, S.S.; Wang, B.; Zhao, Z.; Yu, B.; Zhang, X.; Li, W.; Ding, J.; et al. A review on the occurrence, detection methods, and ecotoxicity of biodegradable microplastics in the aquatic environment: New cause for concern. TrAC Trends Anal. Chem. 2024, 178, 117832. [Google Scholar] [CrossRef]
- Dhivert, E.; Pruvost, J.; Winiarski, T.; Gasperi, J.; Delor-Jestin, F.; Tassin, B.; Mourier, B. Time-varying microplastic contributions of a large urban and industrial area to river sediments. Environ. Pollut. 2024, 347, 123702. [Google Scholar] [CrossRef] [PubMed]
- Margenat, H.; Nel, H.A.; Stonedahl, S.H.; Krause, S.; Sabater, F.; Drummond, J.D. Hydrologic controls on the accumulation of different sized microplastics in the streambed sediments downstream of a wastewater treatment plant (Catalonia, Spain). Environ. Res. Lett. 2021, 16, 115012. [Google Scholar] [CrossRef]
- Mehra, S.; Sharma, K.; Sharma, G.; Singh, M.; Chadha, P. Sources, fate, and impact of microplastics in aquatic environment. In Emerging Contaminants; Nuro, A., Ed.; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef]
- Yuan, B.; Gan, W.; Sun, J.; Lin, B.; Chen, Z. Depth profiles of microplastics in sediments from inland water to coast and their influential factors. Sci. Total Environ. 2023, 903, 166151. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhang, Y.; Kang, S.; Wang, Z.; Wu, C. Microplastics in freshwater sediment: A review on methods, occurrence, and sources. Sci. Total Environ. 2021, 754, 141948. [Google Scholar] [CrossRef]
- Corcoran, P.L.; Norris, T.; Ceccanese, T.; Walzak, M.J.; Helm, P.A.; Marvin, C.H. Hidden plastics of Lake Ontario, Canada and their potential preservation in the sediment record. Environ. Pollut. 2015, 204, 17–25. [Google Scholar] [CrossRef]
- Harris, P.T. The fate of microplastic in marine sedimentary environments: A review and synthesis. Mar. Pollut. Bull. 2020, 158, 111398. [Google Scholar] [CrossRef]
- Bouzekry, A.; Mghili, B.; Mancuso, M.; Bouadil, O.; Bottari, T.; Aksissou, M. Anthropogenic Microparticles Abundance in Sandy Beach Sediments along the Tetouan Coast (Morocco Mediterranean). Environments 2024, 11, 83. [Google Scholar] [CrossRef]
- Guo, W.; Li, D.; Chen, B.; Li, J.; Li, Z.; Cao, X.; Qiu, H.; Zhao, L. Microbial colonization on four types of microplastics to form biofilm differentially affecting organic contaminant biodegradation. Chem. Eng. J. 2025, 503, 158060. [Google Scholar] [CrossRef]
- Zettler, E.R.; Mincer, T.J.; Amaral-Zettler, L.A. Life in the “plastisphere”: Microbial communities on plastic marine debris. Environ. Sci. Technol. 2013, 47, 7137–7146. [Google Scholar] [CrossRef]
- Nauendorf, A.; Krause, S.; Bigalke, N.K.; Gorb, E.V.; Gorb, S.N.; Haeckel, M.; Wahl, M.; Treude, T. Microbial colonization and degradation of polyethylene and biodegradable plastic bags in temperate fine-grained organic-rich marine sediments. Mar. Pollut. Bull. 2016, 103, 168–178. [Google Scholar] [CrossRef]
- Li, Y.; Yang, R.; Guo, L.; Gao, W.; Su, P.; Xu, Z.; Xiao, H.; Ma, Z.; Liu, X.; Gao, P.; et al. The composition, biotic network, and assembly of plastisphere protistan taxonomic and functional communities in plastic-mulching croplands. J. Hazard. Mater. 2022, 430, 128390. [Google Scholar] [CrossRef] [PubMed]
- Sturm, M.T.; Schuhen, K.; Horn, H. Method for rapid biofilm cultivation on microplastics and investigation of its effect on the agglomeration and removal of microplastics using organosilanes. Sci. Total Environ. 2022, 806, 151388. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Zhang, Y.; Zhao, Z.; He, J.; Li, W.; Li, J.; Xu, W.; Ma, Y.; Niu, Z. Colonization characteristics of bacterial communities on microplastics compared with ambient environments (water and sediment) in Haihe Estuary. Sci. Total Environ. 2020, 708, 134876. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Guo, X.; Xue, J. Biofilm-developed microplastics as vectors of pollutants in aquatic environments. Environ. Sci. Technol. 2021, 55, 12780–12790. [Google Scholar] [CrossRef]
- Neto, J.A.B.; Gaylarde, C.; Beech, I.; Bastos, A.C.; Da Silva Quaresma, V.; de Carvalho, D.G. Microplastics and attached microorganisms in sediments of the Vitória bay estuarine system in SE Brazil. Ocean Coast. Manag. 2019, 169, 247–253. [Google Scholar] [CrossRef]
- Su, X.; Yang, L.; Yang, K.; Tang, Y.; Wen, T.; Wang, Y.; Rillig, M.C.; Rohe, L.; Pan, J.; Li, H.; et al. Estuarine plastisphere as an overlooked source of N2O production. Nat. Commun. 2022, 13, 3884. [Google Scholar] [CrossRef]
- Deng, H.; Fu, Q.; Zhang, Y.; Li, D.; He, J.; Feng, D.; Zhao, Y.; Yu, H.; Ge, C. Bacterial communities on polyethylene microplastics in mangrove ecosystems as a function of exposure sites: Compositions and ecological functions. J. Environ. Chem. Eng. 2022, 10, 107924. [Google Scholar] [CrossRef]
- Ashar, M.; Fraser, M.A.; Li, J.; Wang, C.; Huang, W.; Zhang, D.; Zhang, C. Interaction between microbial communities and various plastic types under different aquatic systems. Mar. Environ. Res. 2020, 162, 105151. [Google Scholar] [CrossRef]
- Carson, H.S.; Nerheim, M.S.; Carroll, K.A.; Eriksen, M. The plastic-associated microorganisms of the North Pacific Gyre. Mar. Pollut. Bull. 2013, 75, 126–132. [Google Scholar] [CrossRef]
- Qiang, L.; Cheng, J.; Mirzoyan, S.; Kerkhof, L.J.; Häggblom, M.M. Characterization of microplastic-associated biofilm development along a freshwater-estuarine gradient. Environ. Sci. Technol. 2021, 55, 16402–16412. [Google Scholar] [CrossRef]
- Xie, H.; Chen, J.; Feng, L.; He, L.; Zhou, C.; Hong, P.; Sun, S.; Zhao, H.; Liang, Y.; Ren, L.; et al. Chemotaxis-selective colonization of mangrove rhizosphere microbes on nine different microplastics. Sci. Total Environ. 2021, 752, 142223. [Google Scholar] [CrossRef] [PubMed]
- Rosato, A.; Barone, M.; Negroni, A.; Brigidi, P.; Fava, F.; Xu, P.; Candela, M.; Zanaroli, G. Microbial colonization of different microplastic types and biotransformation of sorbed PCBs by a marine anaerobic bacterial community. Sci. Total Environ. 2020, 705, 135790. [Google Scholar] [CrossRef] [PubMed]
- Rosato, A.; Barone, M.; Negroni, A.; Brigidi, P.; Fava, F.; Biagi, E.; Candela, M.; Zanaroli, G. Bacterial colonization dynamics of different microplastic types in an anoxic salt marsh sediment and impact of adsorbed polychlorinated biphenyls on the plastisphere. Environ. Pollut. 2022, 315, 120411. [Google Scholar] [CrossRef]
- Li, W.; Zhang, Y.; Wu, N.; Zhao, Z.; Xu, W.A.; Ma, Y.; Niu, Z. Colonization characteristics of bacterial communities on plastic debris influenced by environmental factors and polymer types in the Haihe Estuary of Bohai Bay, China. Environ. Sci. Technol. 2019, 53, 10763–10773. [Google Scholar] [CrossRef]
- Miao, L.; Yu, Y.; Adyel, T.M.; Wang, C.; Liu, Z.; Liu, S.; Huang, L.; You, G.; Meng, M.; Qu, H.; et al. Distinct microbial metabolic activities of biofilms colonizing microplastics in three freshwater ecosystems. J. Hazard. Mater. 2021, 403, 123577. [Google Scholar] [CrossRef]
- Xu, X.; Wang, S.; Gao, F.; Li, J.; Zheng, L.; Sun, C.; He, C.; Wang, Z.; Qu, L. Marine microplastic-associated bacterial community succession in response to geography, exposure time, and plastic type in China’s coastal seawaters. Mar. Pollut. Bull. 2019, 145, 278–286. [Google Scholar] [CrossRef]
- Fang, C.; Yang, Y.; Zhang, S.; He, Y.; Pan, S.; Zhou, L.; Wang, J.; Yang, H. Unveiling the impact of microplastics with distinct polymer types and concentrations on tidal sediment microbiome and nitrogen cycling. J. Hazard. Mater. 2024, 472, 134387. [Google Scholar] [CrossRef]
- Hao, Y.; Sun, Y.; Li, M.; Fang, X.; Wang, Z.; Zuo, J.; Zhang, C. Adverse effects of polystyrene microplastics in the freshwater commercial fish, grass carp (Ctenopharyngodon idella): Emphasis on physiological response and intestinal microbiome. Sci. Total Environ. 2023, 856, 159270. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; He, Y.; Yang, Y.; Fu, B.; Pan, S.; Jiao, F.; Wang, J.; Yang, H. Laboratory tidal microcosm deciphers responses of sediment archaeal and bacterial communities to microplastic exposure. J. Hazard. Mater. 2023, 458, 131813. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, G.; Zhang, F.; Wang, H. Diagnostic strategy for the combined effects of microplastics and potentially toxic elements on microbial communities in catchment scale. Sci. Total Environ. 2023, 860, 160499. [Google Scholar] [CrossRef]
- Li, C.; Gan, Y.; Dong, J.; Fang, J.; Chen, H.; Quan, Q.; Liu, J. Impact of microplastics on microbial community in sediments of the Huangjinxia Reservoir-water source of a water diversion project in western China. Chemosphere 2020, 253, 126740. [Google Scholar] [CrossRef] [PubMed]
- Chi, J.; Zhang, H.; Zhao, D. Impact of microplastic addition on degradation of dibutyl phthalate in offshore sediments. Mar. Pollut. Bull. 2021, 162, 111881. [Google Scholar] [CrossRef] [PubMed]
- Kleinteich, J.; Seidensticker, S.; Marggrander, N.; Zarfl, C. Microplastics reduce short-term effects of environmental contaminants. Part II: Polyethylene particles decrease the effect of polycyclic aromatic hydrocarbons on microorganisms. Int. J. Environ. Res. Public Health 2018, 15, 287. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Zhang, X.; Liu, W.; Wang, X.; Wang, Z.; Liu, J.; Zhai, W.; Wang, J.; Zhao, Z. Deciphering the effects of long-term exposure to conventional and biodegradable microplastics on the soil microbiome. J. Hazard. Mater. 2025, 491, 137890. [Google Scholar] [CrossRef]
- Wang, Z.; Xie, J.; Wang, G.; Yang, H.; Li, Z.; Zhang, K.; Shu, R.; Xie, W.; Tian, J.; Li, H.; et al. Enhanced gut damage and microbial imbalance in bullfrog tadpoles (Lithobates catesbeiana) exposed to polystyrene microplastics under high-temperature conditions. Environ. Pollut. 2025, 375, 126339. [Google Scholar] [CrossRef]
- Shen, H.; Sun, Y.; Duan, H.; Ye, J.; Zhou, A.; Meng, H.; Zhu, F.; He, H.; Gu, C. Effect of PVC microplastics on soil microbial community and nitrogen availability under laboratory-controlled and field-relevant temperatures. Appl. Soil Ecol. 2023, 184, 104794. [Google Scholar] [CrossRef]
- Zeng, Q.; Xiang, J.; Yang, C.; Wu, J.; Li, Y.; Sun, Y.; Liu, Q.; Shi, S.; Gong, Z. Microplastics affect nitrogen cycling and antibiotic resistance genes transfer of sediment. Chem. Eng. J. 2023, 454, 140193. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, H.; Yan, M.; Liu, Y.; Ni, X.; Song, J.; Yi, X. Toxicity of tire wear particles and the leachates to microorganisms in marine sediments. Environ. Pollut. 2022, 309, 119744. [Google Scholar] [CrossRef]
- Zhang, X.; Xia, X.; Dai, M.; Cen, J.; Zhou, L.; Xie, J. Microplastic pollution and its relationship with the bacterial community in coastal sediments near Guangdong Province, South China. Sci. Total Environ. 2021, 760, 144091. [Google Scholar] [CrossRef]
- Huang, Y.; Li, W.; Gao, J.; Wang, F.; Yang, W.; Han, L.; Lin, D.; Min, B.; Zhi, Y.; Grieger, K.; et al. Effect of microplastics on ecosystem functioning: Microbial nitrogen removal mediated by benthic invertebrates. Sci. Total Environ. 2021, 754, 142133. [Google Scholar] [CrossRef]
- Yu, H.; Liu, M.; Gang, D.; Peng, J.; Hu, C.; Qu, J. Polyethylene microplastics interfere with the nutrient cycle in water-plant-sediment systems. Water Res. 2022, 214, 118191. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Jiang, X.; Liu, X. Response process and adaptation mechanism of estuarine benthic microbiota to polyvinyl chloride microplastics with and without phthalates. Sci. Total Environ. 2022, 806, 150693. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Yin, H.; Yuan, Y.; Liu, H.; Qi, X.; Ren, Y.; Dang, Z. Discrepancy strategies of sediment abundant and rare microbial communities in response to floating microplastic disturbances: Study using a microcosmic experiment. Sci. Total Environ. 2022, 835, 155346. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, X.; Liu, L.; Lu, H.; Wang, L.; Tang, J. Effects of microplastics on greenhouse gas emissions and microbial communities in sediment of freshwater systems. J. Hazard. Mater. 2022, 435, 129030. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Pan, J.; Xiao, S.; Wang, J.; Gong, X.; Yin, G.; Hou, L.; Liu, M.; Zheng, Y. Microplastics alter nitrous oxide production and pathways through affecting microbiome in estuarine sediments. Water Res. 2022, 221, 118733. [Google Scholar] [CrossRef]
- Qin, R.; Su, C.; Liu, W.; Tang, L.; Li, X.; Deng, X.; Wang, A.; Chen, Z. Effects of exposure to polyether sulfone microplastic on the nitrifying process and microbial community structure in aerobic granular sludge. Bioresour. Technol. 2020, 302, 122827. [Google Scholar] [CrossRef]
- Chen, H.; Wang, Y.; Sun, X.; Peng, Y.; Xiao, L. Mixing effect of polylactic acid microplastic and straw residue on soil property and ecological function. Chemosphere 2020, 243, 125271. [Google Scholar] [CrossRef]
- Dai, H.; Gao, J.; Wang, Z.; Zhao, Y.; Zhang, D. Behavior of nitrogen, phosphorus and antibiotic resistance genes under polyvinyl chloride microplastics pressures in an aerobic granular sludge system. J. Clean. Prod. 2020, 256, 120402. [Google Scholar] [CrossRef]
- Azizi, S.M.M.; Hai, F.I.; Lu, W.; Al-Mamun, A.; Dhar, B.R. A review of mechanisms underlying the impacts of (nano) microplastics on anaerobic digestion. Bioresour. Technol. 2021, 329, 124894. [Google Scholar] [CrossRef]
- Shen, M.; Zhang, Y.; Zhu, Y.; Song, B.; Zeng, G.; Hu, D.; Wen, X.; Ren, X. Recent advances in toxicological research of nanoplastics in the environment: A review. Environ. Pollut. 2019, 252, 511–521. [Google Scholar] [CrossRef]
- Huang, J.; Wen, B.; Miao, L.; Liu, X.; Li, Z.; Ma, T.; Xu, L.; Gao, J.; Chen, Z. Microplastics drive nitrification by enriching functional microorganisms in aquaculture pond waters. Chemosphere 2022, 309, 136646. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Meng, F.; Chen, H.; Chen, Q.; Hu, A.; Yu, C.; Chen, L.; Lv, M. Leachable additives of tire particles explain the shift in microbial community composition and function in coastal sediments. Environ. Sci. Technol. 2022, 56, 12257–12266. [Google Scholar] [CrossRef] [PubMed]
- Nie, Z.; Wang, L.; Lin, Y.; Xiao, N.; Zhao, J.; Wan, X.; Hu, J. Effects of polylactic acid (PLA) and polybutylene adipate-co-terephthalate (PBAT) biodegradable microplastics on the abundance and diversity of denitrifying and anammox bacteria in freshwater sediment. Environ. Pollut. 2022, 315, 120343. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; He, S.; Wang, Q.; Luo, Y.; Tu, C.; Wu, W.; Jiang, H. Nanoplastics enhance the denitrification process and microbial interaction network in wetland soils. Water Res. 2024, 259, 121796. [Google Scholar] [CrossRef]
- Sun, Y.; Duan, C.; Cao, N.; Ding, C.; Huang, Y.; Wang, J. Biodegradable and conventional microplastics exhibit distinct microbiome, functionality, and metabolome changes in soil. J. Hazard. Mater. 2022, 424, 127282. [Google Scholar] [CrossRef]
- Riveros, G.; Urrutia, H.; Araya, J.; Zagal, E.; Schoebitz, M. Microplastic pollution on the soil and its consequences on the nitrogen cycle: A review. Environ. Sci. Pollut. Res. 2022, 29, 7997–8011. [Google Scholar] [CrossRef]
- Rogers, K.L.; Carreres-Calabuig, J.A.; Gorokhova, E.; Posth, N.R. Micro-by-micro interactions: How microorganisms influence the fate of marine microplastics. Limnol. Oceanogr. Lett. 2020, 5, 18–36. [Google Scholar] [CrossRef]
- Hirai, H.; Takada, H.; Ogata, Y.; Yamashita, R.; Mizukawa, K.; Saha, M.; Kwan, C.; Moore, C.; Gray, H.; Laursen, D.; et al. Organic micropollutants in marine plastics debris from the open ocean and remote and urban beaches. Mar. Pollut. Bull. 2011, 62, 1683–1692. [Google Scholar] [CrossRef]
- Shimao, M. Biodegradation of plastics. Curr. Opin. Biotechnol. 2001, 12, 242–247. [Google Scholar] [CrossRef]
- Galgani, L.; Tsapakis, M.; Pitta, P.; Tsiola, A.; Tzempelikou, E.; Kalantzi, I.; Esposito, C.; Loiselle, A.; Tsotskou, A.; Zivanovic, S.; et al. Microplastics increase the marine production of particulate forms of organic matter. Environ. Res. Lett. 2019, 14, 124085. [Google Scholar] [CrossRef]
- Liu, J.; Xu, G.; Zhao, S.; He, J. Microbiomes of coastal sediments and plastispheres shaped by microplastics and decabrominated diphenyl ether. Water Res. 2025, 280, 123417. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Liu, S.; Bi, M.; Yang, X.; Deng, R.; Chen, Y. Aging behavior of microplastics affected DOM in riparian sediments: From the characteristics to bioavailability. J. Hazard. Mater. 2022, 431, 128522. [Google Scholar] [CrossRef]
- Shen, M.; Ye, S.; Zeng, G.; Zhang, Y.; Xing, L.; Tang, W.; Wen, X.; Liu, S. Can microplastics pose a threat to ocean carbon sequestration? Mar. Pollut. Bull. 2020, 150, 110712. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wangjin, X.; Zhang, Y.; Wang, N.; Wang, Y.; Meng, G.; Chen, Y. The toxicity of virgin and UV-aged PVC microplastics on the growth of freshwater algae Chlamydomonas reinhardtii. Sci. Total. Environ. 2020, 749, 141603. [Google Scholar] [CrossRef] [PubMed]
- You, X.; You, M.; Lyu, Y.; Peng, G.; Sun, W. Single and combined exposure to micro (nano) plastics and azithromycin disturbing the photosynthetic carbon fixation of Synechocystis sp. Environ. Sci. Nano 2022, 9, 4354–4366. [Google Scholar] [CrossRef]
- Pinnell, L.J.; Turner, J.W. Shotgun metagenomics reveals the benthic microbial community response to plastic and bioplastic in a coastal marine environment. Front. Microbiol. 2019, 10, 1252. [Google Scholar] [CrossRef]
- Hayden, H.L.; Drake, J.; Imhof, M.; Oxley, A.P.; Norng, S.; Mele, P.M. The abundance of nitrogen cycle genes amoA and nifH depends on land-uses and soil types in South-Eastern Australia. Soil Biol. Biochem. 2010, 42, 1774–1783. [Google Scholar] [CrossRef]
- Luo, G.; Jin, T.; Zhang, H.; Peng, J.; Zuo, N.; Huang, Y.; Han, Y.; Tian, C.; Yang, Y.; Peng, K.; et al. Deciphering the diversity and functions of plastisphere bacterial communities in plastic-mulching croplands of subtropical China. J. Hazard. Mater. 2022, 422, 126865. [Google Scholar] [CrossRef]
- Luo, G.; Xue, C.; Jiang, Q.; Xiao, Y.; Zhang, F.; Guo, S.; Shen, Q.; Ling, N. Soil carbon, nitrogen, and phosphorus cycling microbial populations and their resistance to global change depend on soil C:N:P stoichiometry. Msystems 2020, 5, 10–1128. [Google Scholar] [CrossRef]
- Kim, T.; Lee, J.; Kim, J.; Oh, J. Behavioral characteristics of phosphorus in sediments according to the forms of phosphorus. J. Ecol. Environ. 2015, 38, 319–326. [Google Scholar] [CrossRef]
- Song, X.; Ding, J.; Zhang, Y.; Zhu, M.; Peng, Y.; Wang, Z.; Pan, G.; Zou, H. New insights into changes in phosphorus profile at sediment-water interface by microplastics: Role of benthic bioturbation. J. Hazard. Mater. 2024, 469, 134047. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Tran, P.Q.; Cowley, E.S.; Trembath-Reichert, E.; Anantharaman, K. Diversity and ecology of microbial sulfur metabolism. Nat. Rev. Microbiol. 2025, 23, 122–140. [Google Scholar] [CrossRef] [PubMed]
- Dombrowski, N.; Teske, A.P.; Baker, B.J. Expansive microbial metabolic versatility and biodiversity in dynamic Guaymas Basin hydrothermal sediments. Nat. Commun. 2018, 9, 4999. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MPs | Dominant Species | Abundance | Reference | |
|---|---|---|---|---|
| Decreasing | Increasing | |||
| PE, PVC, PUF, PLA | Bacteriodes, Proteobacteria (classes Deltaproteobacteria and Gammaproteobacteria) |
|
| [18] |
| PVC, PP, PLA | Proteobacteria, Acidobacteriota, Chloroflexi, Bacteroidota, Desulfobacterota |
|
| [16] |
| PE, PET, PVC | Proteobacteria, Actinobacteriota |
|
| [68] |
| TWPs | Proteobacteria, Desulfobacteriota | Defluvimonas, Defluviitaleaceae, Fusibacter, Lutibacter, Desulfopila, Citrobacter, Stenotrophomonas | [69] | |
| Proteobacteria, Bacteroidetes |
| [70] | ||
| PE | Proteobacteria, Firmicutes, Chloroflexi |
| [71] | |
| PE | Actinobacteria, Firmicutes | [72] | ||
| PVC | Bacteria (Actinobacteria, Bacteroidetes, Chloroflexi, Firmicutes, Gemmatimonadetes, and Planctomycetes) and eukaryotes (Ascomycota, Bacillariophyta, Chordata, and Streptophyta) | [73] | ||
| PE, PVC | Chloroflexi, Proteobacteria, Bacteroidota, Firmicutes, Actinobacteriota | Proteobacteria, Bacteroidota | [14] | |
| PE, PS, PVC, PLA | Proteobacteria (Betaproteobacteria and Gammaproteobacteria), Bacteroides, Nitrospirae |
|
| [74] |
| PET | Proteobacteria, Chloroflexi, Actinbacteriota, Bacterioidota, Desulfobacterota, Acidobacteriota | Acidiferrobacteraceae, Cellulomonadaceae, norank_o_EPR3968-O8a-Bc78, norank_o__Oligoflexales, Vulgatibacteraceae | FTLpost3, MAT_CR_H6_H10, MWH-UniP_aquatic_group, norank_o_Chthonomonadal, norank_o__norank_c__Microgenomatia, Pedosphaeraceae, Saprospiraceae | [75] |
| PE, PLA, PVC | Bacteroidota, Proteobacteria, Firmicutes, Actinobacteria, Chloroflexi, Acidobacteriota |
| [76] | |
| PE, PLA, PET, PBS, PC | Proteobacteria, Chloroflexi, Actinobacteriota |
|
| [58] |
| Location | Microplastic | Nitrification | Denitrification | Reference |
|---|---|---|---|---|
| Blue Sea Aquaculture Farm, Shanghai, China | PA | (+) | [82] | |
| The Bohai Sea | PE, PET, PVC | PE and PET (+); PVC (−) | PVC (+) | [68] |
| The York River estuary in Gloucester Point, VA | PVC, PUF, PLA | PVC (−); PUF and PLA (+) | PVC (−); PUF and PLA (+) | [18] |
| Lake Mati | PET | (+) | (+) | [75] |
| Gaoyang Lake | PLA, PVC, PP | PLA (−); PVC, PP (+) | PLA (+) | [16] |
| Low tide of an intertidal marsh in Yantai, China | TP | (−) | [83] | |
| Chongming eastern intertidal flat | PLA | PLA (+) | [76] | |
| The Daihai Lake in Ulaanchab, Inner Mongolia Autonomous Region | PE, PVC | (−) | [14] | |
| Lake Jinhu in Chongqing, China | PE | (+) | [71] | |
| Nearshore area of Liaohe Estuary National Nature Reserve, China | TWP | (+) | [69] | |
| 29°04–29°22 N, 112°17–112°42 E | PE | (+) | [72] | |
| Yangchun Lake, Wuhan, China | PLA, PBAT | (−) | [84] | |
| Suishi Ferry of the Pearl River, Guangzhou, China | PE, PS, PLA, PVC | (−) | (−) | [74] |
| Core mature mangrove area in Shantou City, southeastern China | PE, PLA, PET, PBS, PC | PC (−); PE (+); PLA (+) | PLA (+); PC (−); PE (+) | [58] |
| Poyang Lake wetland in China | (+) | [85] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Ding, D.; Liu, Y.; Yao, Z.; Duan, P.; Yuan, H.; Fan, H.; Dai, Y. Microbial Community Dynamics and Biogeochemical Cycling in Microplastic-Contaminated Sediment. Nanomaterials 2025, 15, 902. https://doi.org/10.3390/nano15120902
Zhang X, Ding D, Liu Y, Yao Z, Duan P, Yuan H, Fan H, Dai Y. Microbial Community Dynamics and Biogeochemical Cycling in Microplastic-Contaminated Sediment. Nanomaterials. 2025; 15(12):902. https://doi.org/10.3390/nano15120902
Chicago/Turabian StyleZhang, Xuanxuan, Dina Ding, Yinglin Liu, Zhiming Yao, Pingping Duan, Hanyu Yuan, Hanzhong Fan, and Yanhui Dai. 2025. "Microbial Community Dynamics and Biogeochemical Cycling in Microplastic-Contaminated Sediment" Nanomaterials 15, no. 12: 902. https://doi.org/10.3390/nano15120902
APA StyleZhang, X., Ding, D., Liu, Y., Yao, Z., Duan, P., Yuan, H., Fan, H., & Dai, Y. (2025). Microbial Community Dynamics and Biogeochemical Cycling in Microplastic-Contaminated Sediment. Nanomaterials, 15(12), 902. https://doi.org/10.3390/nano15120902