
Combined Effect of NZVI and H2O2 on the Cyanobacterium Microcystis aeruginosa: Performance and Mechanism

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cyanobacterium and Chemical Reagents
2.2. Experimental Procedures
2.3. Determination of Chlorophyll and Photosynthetic Pigments
2.4. Analytical Methods
2.4.1. Determination of Zeta Potential
2.4.2. Determination of Antioxidant Ability
2.4.3. Total Organic Carbon Analysis
2.4.4. Excitation–Emission Matrix (EEM) Fluorescence Spectroscopy Analysis
3. Results and Discussion
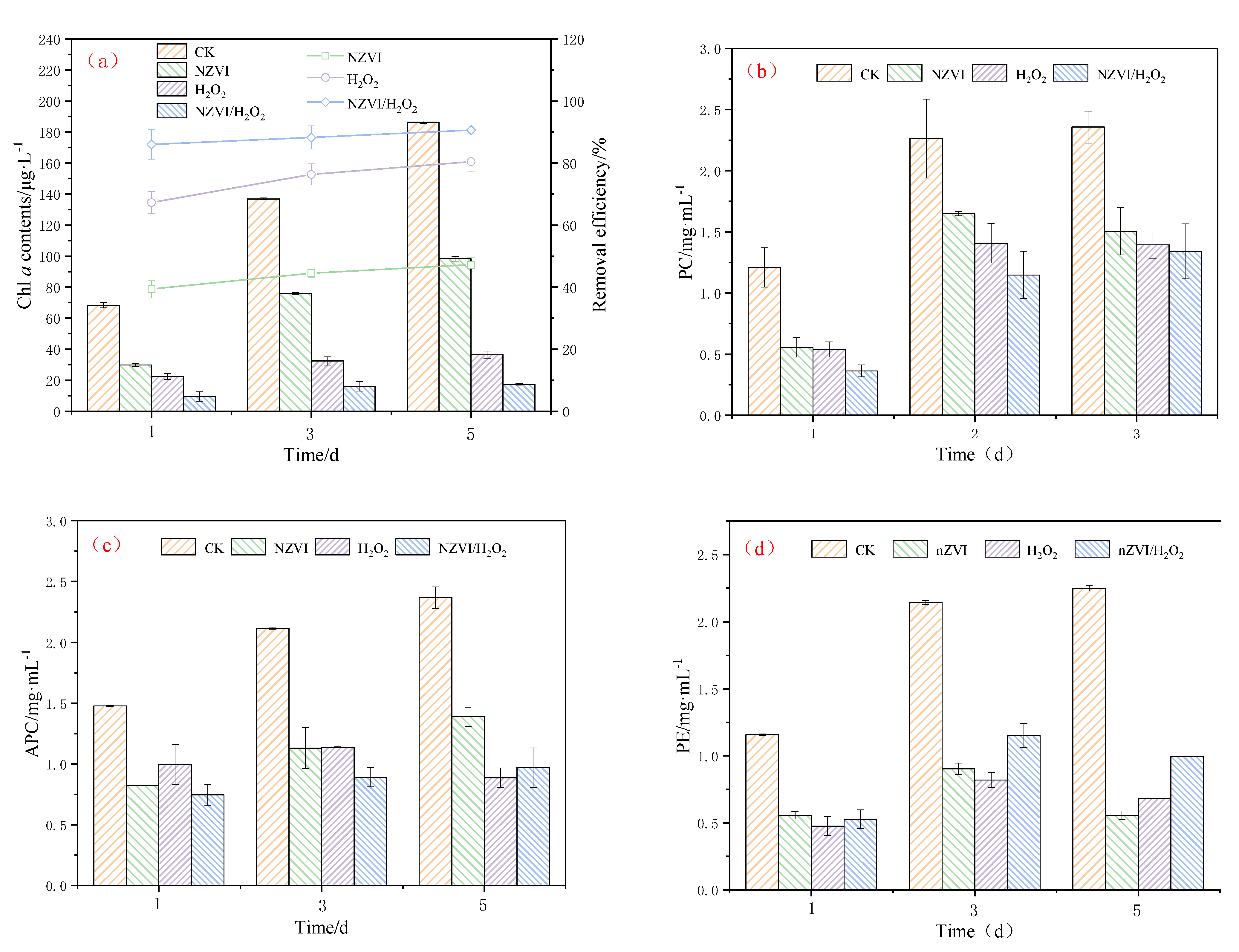
3.1. Growth Inhibition of NZVI and H2O2 on M. aeruginosa
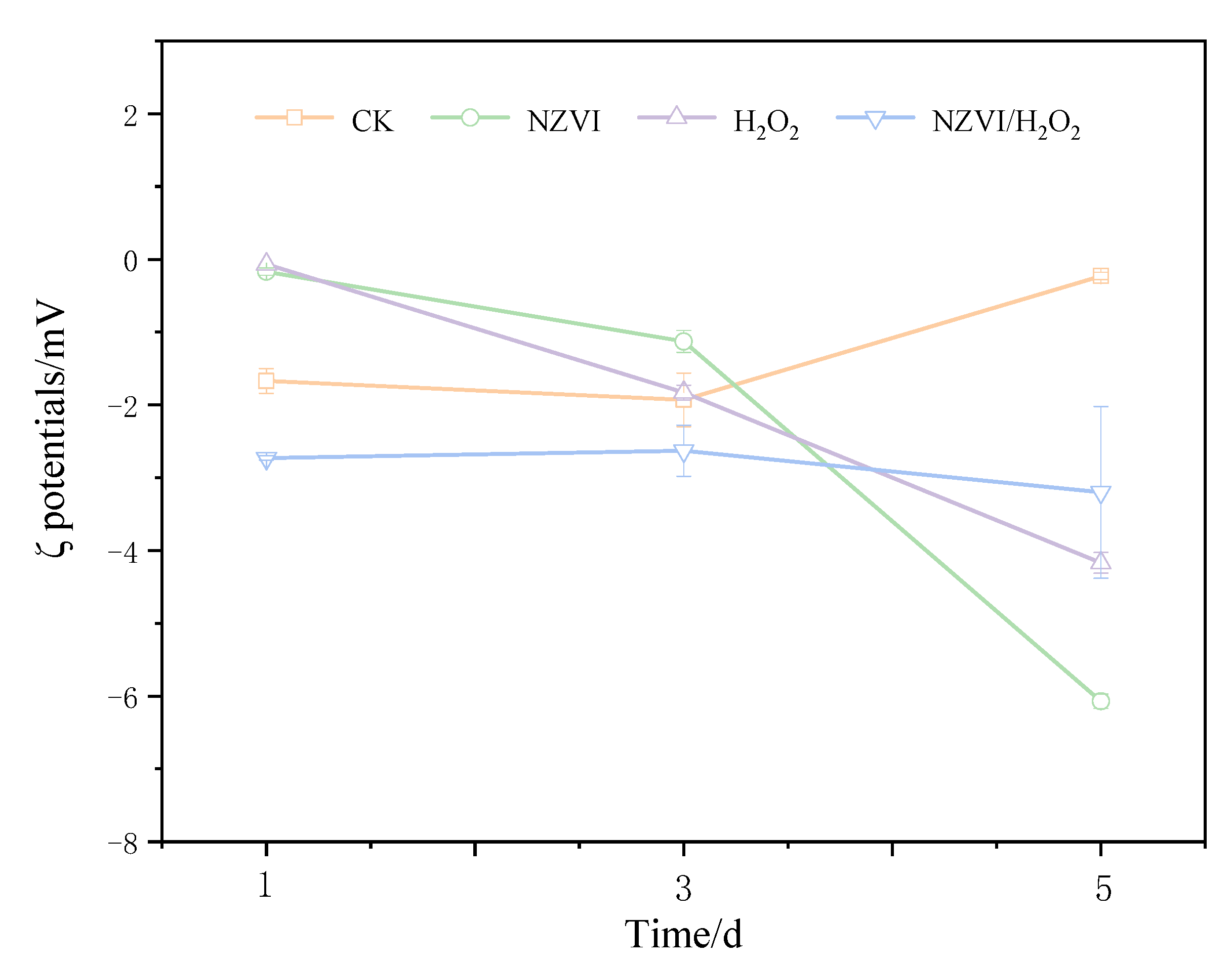
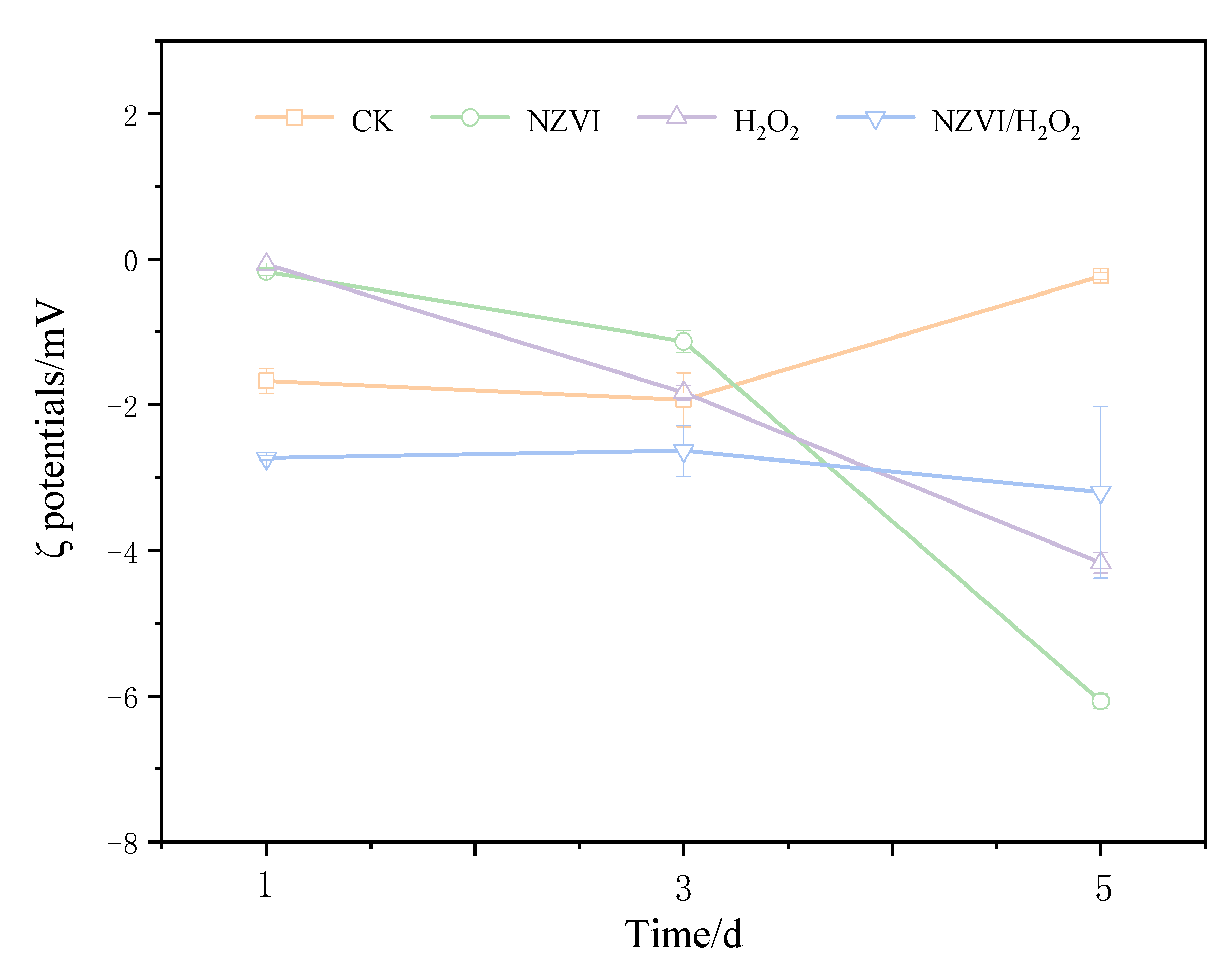
3.2. Effects of NZVI and H2O2 on the Zeta Potential
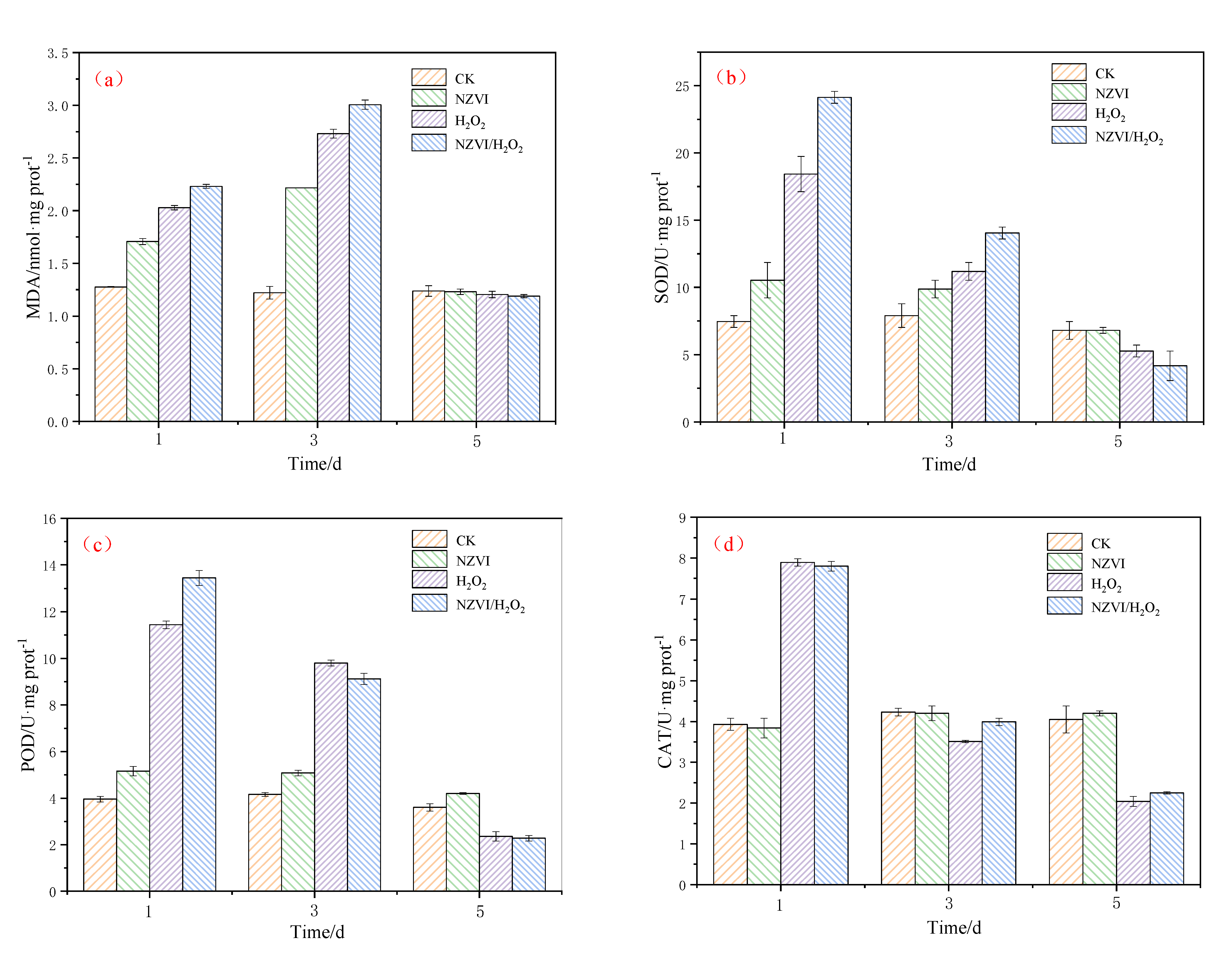
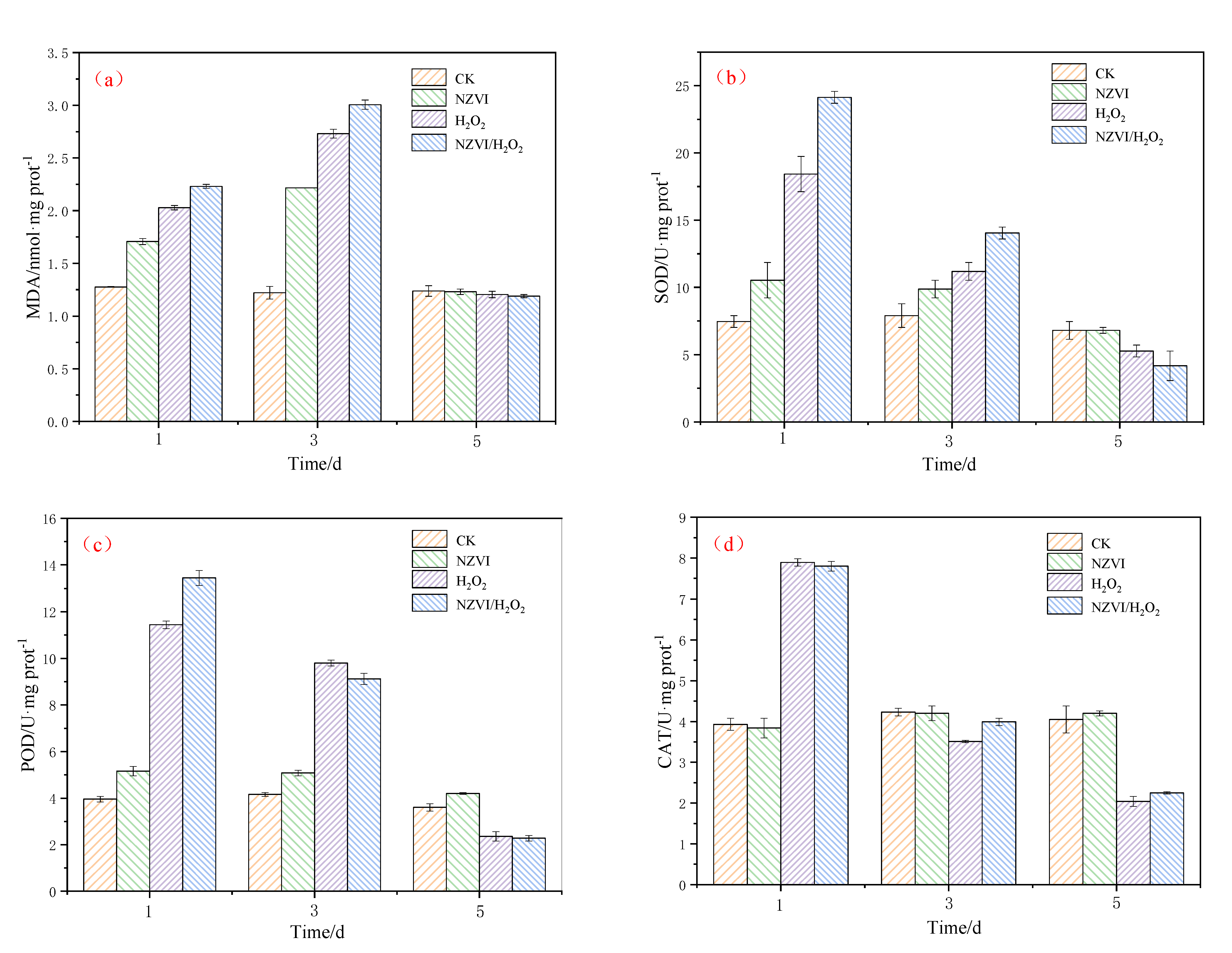
3.3. Effects of NZVI and H2O2 on the Antioxidant System
3.4. Effects of NZVI and H2O2 on AOMs
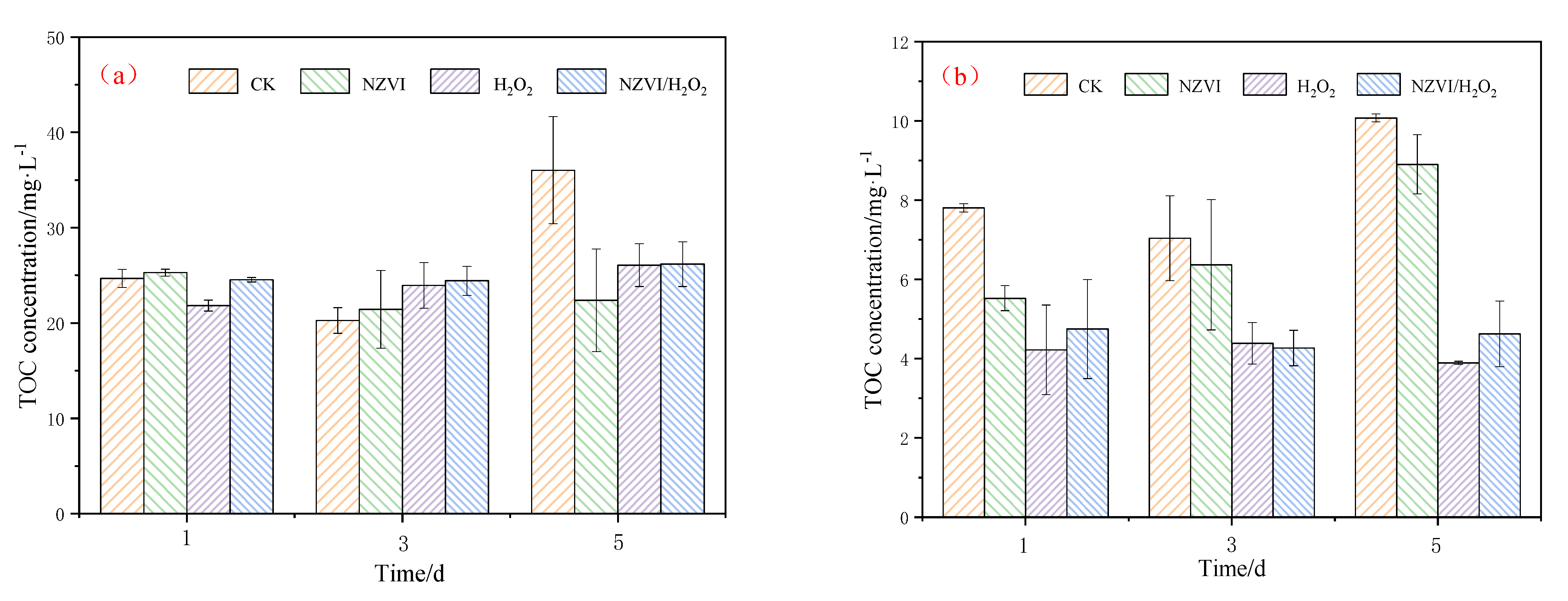
3.4.1. TOC Variations of AOMs
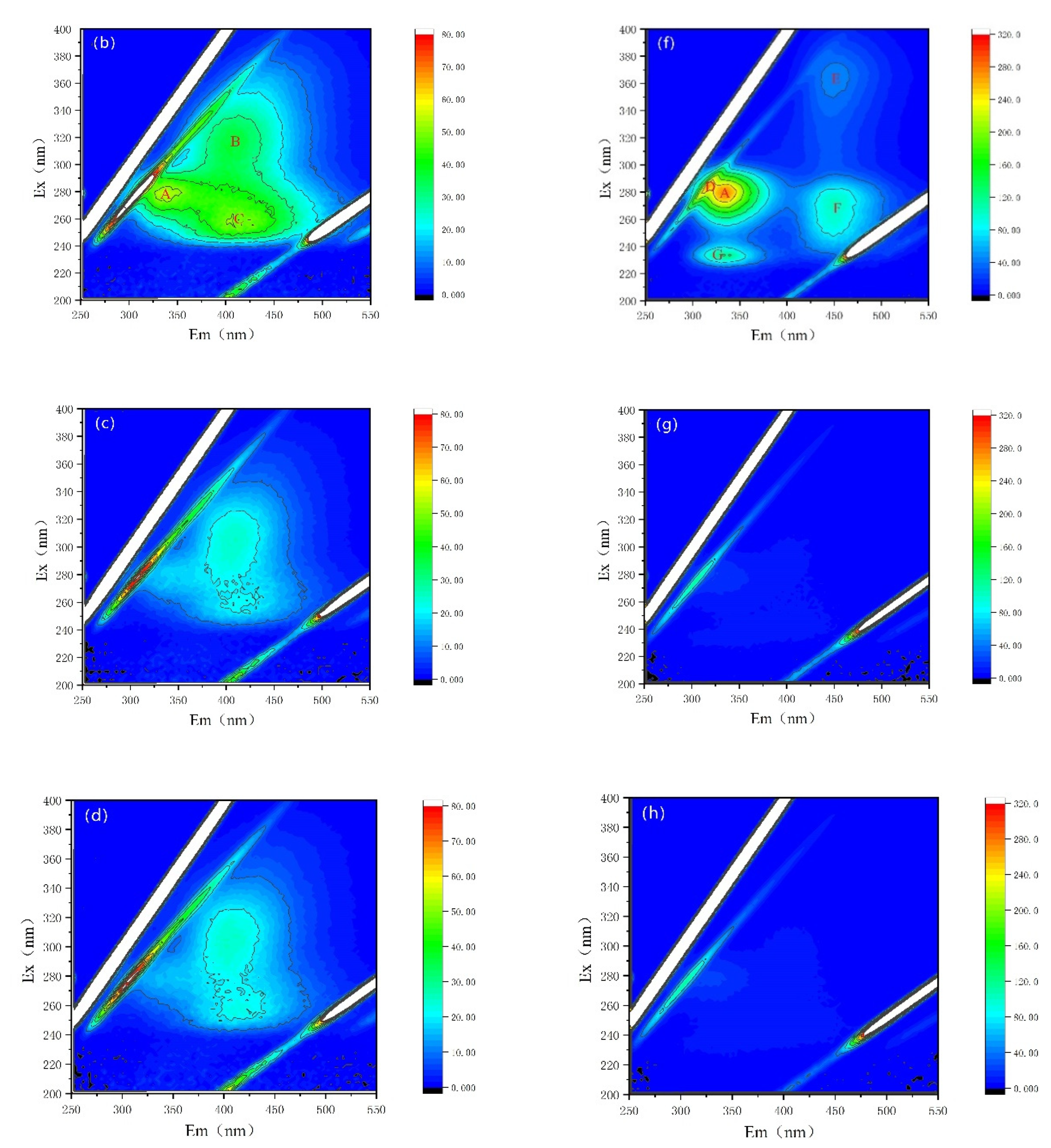
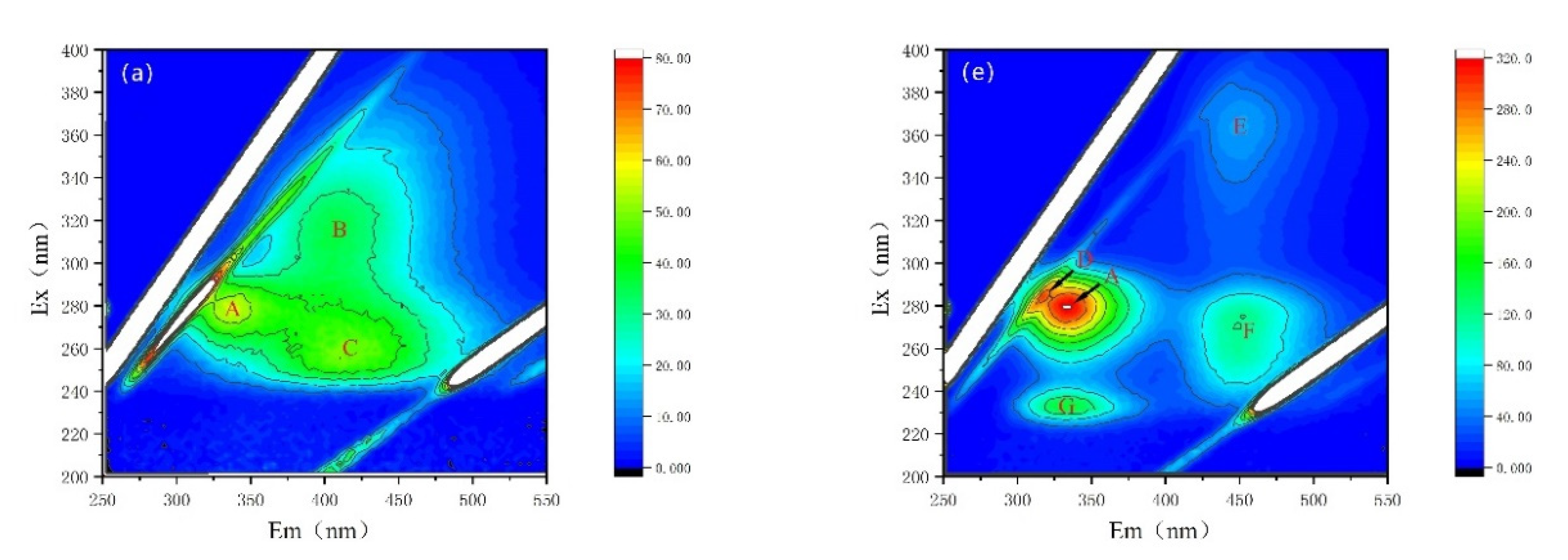
3.4.2. Fluorescence Properties of AOMs
3.5. Discussion of the Mechanism
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kibuye, F.A.; Zamyadi, A.; Wert, E.C. A critical review on operation and performance of source water control strategies for cyanobacterial blooms: Part II-mechanical and biological control methods. Harmful Algae 2021, 109, 102119. [Google Scholar] [CrossRef]
- Zheng, X.; Niu, X.; Zhang, D.; Ye, X.; Ma, J.; Lv, M.; Lin, Z. Removal of Microcystis aeruginosa by natural pyrite-activated persulfate: Performance and the significance of iron species. Chem. Eng. J. 2021, 428, 132565. [Google Scholar] [CrossRef]
- Zhao, Z.; Sun, W.; Ray, A.K.; Mao, T.; Ray, M.B. Coagulation and disinfection by-products formation potential of extracellular and intracellular matter of algae and cyanobacteria. Chemosphere 2019, 245, 125669. [Google Scholar] [CrossRef]
- Qi, J.; Lan, H.; Liu, R.; Liu, H.; Qu, J. Fe(II)-regulated moderate pre-oxidation of Microcystis aeruginosa and formation of size-controlled algae flocs for efficient flotation of algae cell and organic matter. Water Res. 2018, 137, 57–63. [Google Scholar] [CrossRef]
- Wang, C.; Wang, X.; Wang, P.; Chen, B.; Hou, J.; Qian, J.; Yang, Y. Effects of iron on growth, antioxidant enzyme activity, bound extracellular polymeric substances and microcystin production of Microcystis aeruginosa FACHB-905. Ecotoxicol. Environ. Saf. 2016, 132, 231–239. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, Y.; Qin, Y.; Li, H. Removal of Microcystis aeruginosa and control of algal organic matter by Fe(II)/peroxymonosulfate pre-oxidation enhanced coagulation. Chem. Eng. J. 2021, 403, 126381. [Google Scholar] [CrossRef]
- Zhou, S.; Shao, Y.; Gao, N.; Deng, Y.; Li, L.; Deng, J.; Tan, C. Characterization of algal organic matters of Microcystis aeruginosa: Biodegradability, DBP formation and membrane fouling potential. Water Res. 2014, 52, 199–207. [Google Scholar] [CrossRef]
- Kong, Y.; Zou, P.; Yang, Q.; Xu, X.; Miao, L.; Zhu, L. Physiological responses of Microcystis aeruginosa under the stress of antialgal actinomycetes. J. Hazard. Mater. 2013, 262, 274–280. [Google Scholar] [CrossRef]
- Li, L.; Zhang, H.; Pan, G. Influence of zeta potential on the flocculation of cyanobacteria cells using chitosan modified soil. J. Environ. Sci. 2015, 28, 47–53. [Google Scholar] [CrossRef]
- Ma, J.; Xia, W.; Fu, X.; Ding, L.; Kong, Y.; Zhang, H.; Fu, K. Magnetic flocculation of algae-laden raw water and removal of extracellular organic matter by using composite flocculant of Fe3O4/cationic polyacrylamide. J. Clean. Prod. 2020, 248, 119276. [Google Scholar] [CrossRef]
- Ishag, A.; Li, Y.; Zhang, N.; Wang, H.; Guo, H.; Mei, P.; Sun, Y. Environmental application of emerging zero-valent iron-based materials on removal of radionuclides from the wastewater: A review. Environ. Res. 2020, 188, 109855. [Google Scholar] [CrossRef]
- Lei, C.; Zhang, L.; Yang, K.; Zhu, L.; Lin, D. Toxicity of iron-based nanoparticles to green algae: Effects of particle size, crystal phase, oxidation state and environmental aging. Environ. Pollut. 2016, 218, 505–512. [Google Scholar] [CrossRef]
- Marsalek, B.; Jancula, D.; Marsalkova, E.; Mashlan, M.; Safarova, K.; Tucek, J.; Zboril, R. Multimodal Action and Selective Toxicity of Zerovalent Iron Nanoparticles against Cyanobacteria. Environ. Sci. Technol. 2012, 46, 2316–2323. [Google Scholar] [CrossRef]
- Fujii, M.; Dang, T.C.; Rose, A.L.; Omura, T.; Waite, T.D. effect of light on iron uptake by the freshwater Cyanobacterium Microcystis aeruginosa. Environ. Sci. Technol. 2011, 45, 1391–1398. [Google Scholar] [CrossRef]
- Qiu, Y.; Wang, Z.; Liu, F.; Liu, J.; Zhou, T. Effect of different kinds of complex iron on the growth of Anabaena flos-aquae. Environ. Technol. 2018, 40, 2889–2896. [Google Scholar] [CrossRef]
- Fan, J.; Hu, Y.-B.; Li, X.-Y. Nanoscale zero-valent iron coated with magnesium hydroxide for effective removal of cyanobacteria from water. ACS Sustain. Chem. Eng. 2018, 6, 15135–15142. [Google Scholar] [CrossRef]
- Garg, S.; Wang, K.; Waite, T.D. Impact of Microcystis aeruginosa exudate on the formation and reactivity of iron oxide particles following Fe(II) and Fe(III) addition. Environ. Sci. Technol. 2017, 51, 5500–5510. [Google Scholar] [CrossRef]
- Le, K.T.N.; Goitom, E.; Trigui, H.; Sauve, S.; Prevost, M.; Dorner, S. The effects of ferric sulfate (Fe2(SO4)3) on the removal of cyanobacteria and cyanotoxins: A mesocosm experiment. Toxins 2021, 13, 753. [Google Scholar] [CrossRef]
- Wang, J.; Wan, Y.; Yue, S.; Ding, J.; Xie, P.; Wang, Z. Simultaneous Removal of Microcystis aeruginosa and 2,4,6-Trichlorophenol by UV/Persulfate Process. Front. Chem. 2020, 8, 591641. [Google Scholar] [CrossRef]
- Zhang, X.; Ma, Y.; Tang, T.; Xiong, Y.; Dai, R. Removal of cyanobacteria and control of algal organic matter by simultaneous oxidation and coagulation—Comparing the H2O2/Fe(II) and H2O2/Fe(III) processes. Sci. Total Environ. 2020, 720, 137653. [Google Scholar] [CrossRef]
- Barešová, M.; Načeradská, J.; Novotná, K.; Čermáková, L.; Pivokonský, M. The impact of preozonation on the coagulation of cellular organic matter produced by Microcystis aeruginosa and its toxin degradation. J. Environ. Sci. 2020, 98, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Kwon, M.; Ahn, Y.; Jung, Y.; Nam, S.-N.; Choi, I.-H.; Kang, J.-W. Characteristics of intracellular algogenic organic matter and its reactivity with hydroxyl radicals. Water Res. 2018, 144, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Li, J.; Chen, M.; Koh, K.Y.; Du, Z.; Gin, K.Y.-H.; He, Y.; Ong, C.N.; Chen, J.P. Microcystis aeruginosa removal by peroxides of hydrogen peroxide, peroxymonosulfate and peroxydisulfate without additional activators. Water Res. 2021, 201, 117263. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Liu, J.; Zhao, Z.; Peng, W.; Cui, F.; Liang, Z. Microcystis aeruginosa-laden water treatment using peroxymonosulfate enhanced Fe(II) coagulation: Performance and the role of in situ formed Fe3O4. Chem. Eng. J. 2019, 382, 123012. [Google Scholar] [CrossRef]
- Liu, M.; Shi, X.; Chen, C.; Yu, L.; Sun, C. Responses of Microcystis colonies of different sizes to hydrogen peroxide stress. Toxins 2017, 9, 306. [Google Scholar] [CrossRef] [PubMed]
- Shahi, N.K.; Maeng, M.; Choi, I.; Dockko, S. Degradation effect of ultraviolet-induced advanced oxidation of chlorine, chlorine dioxide, and hydrogen peroxide and its impact on coagulation of extracellular organic matter produced by Microcystis aeruginosa. Chemosphere 2021, 281, 130765. [Google Scholar] [CrossRef]
- Jia, P.; Zhou, Y.; Zhang, X.; Zhang, Y.; Dai, R. Cyanobacterium removal and control of algal organic matter (AOM) release by UV/H2O2 pre-oxidation enhanced Fe(II) coagulation. Water Res. 2018, 131, 122–130. [Google Scholar] [CrossRef]
- Xin, H.; Yang, S.; Tang, Y.; Wu, M.; Deng, Y.; Xu, B.; Gao, N. Mechanisms and performance of calcium peroxide-enhanced Fe(II) coagulation for treatment of Microcystis aeruginosa-laden water. Environ. Sci. Water Res. Technol. 2020, 6, 1272–1285. [Google Scholar] [CrossRef]
- Kong, Y.; Zhu, L.; Zou, P.; Qi, J.; Yang, Q.; Song, L.; Xu, X. Isolation and characterization of dissolved organic matter fractions from antialgal products of Microcystis aeruginosa. Environ. Sci. Pollut. Res. 2013, 21, 3946–3954. [Google Scholar] [CrossRef]
- Hou, J.; Yang, Y.; Wang, P.; Wang, C.; Miao, L.; Wang, X.; Lv, B.; You, G.; Liu, Z. Effects of CeO2, CuO, and ZnO nanoparticles on physiological features of Microcystis aeruginosa and the production and composition of extracellular polymeric substances. Environ. Sci. Pollut. Res. 2016, 24, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, Z.; Chen, H.; Wen, Y. Effect of hydrogen peroxide on Microcystic aeruginosa: Role of cytochromes P450. Sci. Total Environ. 2018, 626, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Tzubari, Y.; Magnezi, L.; Be’Er, A.; Berman-Frank, I. Iron and phosphorus deprivation induce sociality in the marine bloom-forming cyanobacterium Trichodesmium. ISME J. 2018, 12, 1682–1693. [Google Scholar] [CrossRef] [PubMed]
- Henderson, R.; Parsons, S.A.; Jefferson, B. Successful Removal of Algae through the Control of Zeta Potential. Sep. Sci. Technol. 2008, 43, 1653–1666. [Google Scholar] [CrossRef]
- Liu, J.; Yang, C.; Chi, Y.; Wu, D.; Dai, X.; Zhang, X.; Igarashi, Y.; Luo, F. Algicidal characterization and mechanism of Bacillus licheniformis Sp34 against Microcystis aeruginosa in Dianchi Lake. J. Basic Microbiol. 2019, 59, 1112–1124. [Google Scholar] [CrossRef]
- Qian, H.; Zhu, K.; Lu, H.; Lavoie, M.; Chen, S.; Zhou, Z.; Deng, Z.; Chen, J.; Fu, Z. Contrasting silver nanoparticle toxicity and detoxification strategies in Microcystis aeruginosa and Chlorella vulgaris: New insights from proteomic and physiological analyses. Sci. Total Environ. 2016, 572, 1213–1221. [Google Scholar] [CrossRef]
- Wang, J.; Shi, Y.; Zhang, M.; Li, Y.; Liu, Q.; Xie, S. Allelopathically inhibitory effects of 5,4′-dihydroxyflavone on the growth, antioxidant system and photosynthesis of the bloom-forming cyanobacterium Microcystis aeruginosa. J. Appl. Phycol. 2021, 33, 3843–3852. [Google Scholar] [CrossRef]
- Pádrová, K.; Lukavský, J.; Nedbalová, L.; Čejková, A.; Cajthaml, T.; Sigler, K.; Vítová, M.; Řezanka, T. Trace concentrations of iron nanoparticles cause overproduction of biomass and lipids during cultivation of cyanobacteria and microalgae. J. Appl. Phycol. 2014, 27, 1443–1451. [Google Scholar] [CrossRef]
- Wang, C.; Yang, Y.; Hou, J.; Wang, P.; Miao, L.; Wang, X.; Guo, L. Optimization of cyanobacterial harvesting and extracellular organic matter removal utilizing magnetic nanoparticles and response surface methodology: A comparative study. Algal Res. 2019, 45, 101756. [Google Scholar] [CrossRef]
- Pivokonsky, M.; Safarikova, J.; Baresova, M.; Pivokonska, L.; Kopecka, I. A comparison of the character of algal extracellular versus cellular organic matter produced by cyanobacterium, diatom and green alga. Water Res. 2014, 51, 37–46. [Google Scholar] [CrossRef]
- Pivokonský, M.; Načeradská, J.; Kopecká, I.; Baresova, M.; Jefferson, B.; Li, X.; Henderson, R.K. The impact of algogenic organic matter on water treatment plant operation and water quality: A review. Crit. Rev. Environ. Sci. Technol. 2015, 46, 291–335. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
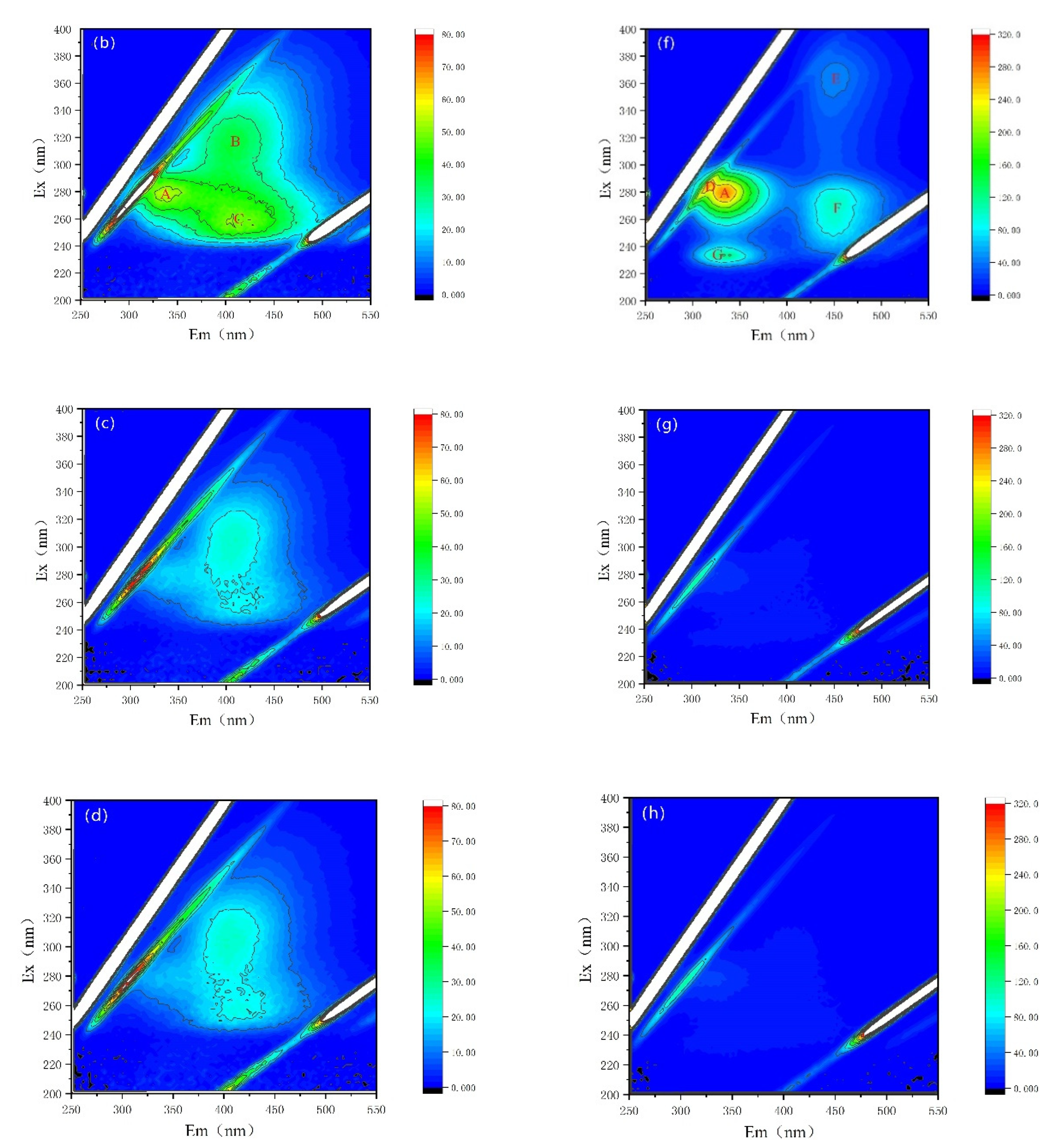{kind=link}
| Peak A | Peak B | Peak C | Peak D | Peak E | Peak F | Peak G | ||
|---|---|---|---|---|---|---|---|---|
| Ex/Em(nm/nm) | 278~280/334~335 | 314/400 | 255/406 | 282/313 | 364/451 | 272/451 | 232/331 | |
| Substance | Soluble cyanobacterial metabolic byproducts | Fulvic-like acids | Fulvic-like acids | Soluble cyanobacterial metabolic byproducts | Fulvic-like acids | Fulvic-like acids | Aromatic proteins II | |
| EOM | CK | 56.70 | 36.43 | 50.17 | / | / | / | / |
| NZVI | 55.17 | 37.96 | 51.08 | / | / | / | / | |
| H2O2 | / | 25.59 | 22.46 | / | / | / | / | |
| NZVI/H2O2 | / | 25.56 | 22.46 | / | / | / | / | |
| IOM | CK | 321.7 | / | / | 296.5 | 56.19 | 124.0 | 152.6 |
| NZVI | 274.1 | / | / | 268.5 | 45.83 | 103.2 | 126.4 | |
| H2O2 | 20.35 | / | / | / | / | / | / | |
| NZVI/H2O2 | 22.82 | / | / | / | / | / | / | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, Y.; Ji, L.; Wang, Y.; Li, J.; Lu, H.; Mo, S.; Wang, X.; Zhu, L.; Xu, X.; Zheng, X. Combined Effect of NZVI and H2O2 on the Cyanobacterium Microcystis aeruginosa: Performance and Mechanism. Nanomaterials 2022, 12, 3017. https://doi.org/10.3390/nano12173017
Kong Y, Ji L, Wang Y, Li J, Lu H, Mo S, Wang X, Zhu L, Xu X, Zheng X. Combined Effect of NZVI and H2O2 on the Cyanobacterium Microcystis aeruginosa: Performance and Mechanism. Nanomaterials. 2022; 12(17):3017. https://doi.org/10.3390/nano12173017
Chicago/Turabian StyleKong, Yun, Lipeng Ji, Yue Wang, Jiake Li, Hao Lu, Shuhong Mo, Xianxun Wang, Liang Zhu, Xiangyang Xu, and Xing Zheng. 2022. "Combined Effect of NZVI and H2O2 on the Cyanobacterium Microcystis aeruginosa: Performance and Mechanism" Nanomaterials 12, no. 17: 3017. https://doi.org/10.3390/nano12173017
APA StyleKong, Y., Ji, L., Wang, Y., Li, J., Lu, H., Mo, S., Wang, X., Zhu, L., Xu, X., & Zheng, X. (2022). Combined Effect of NZVI and H2O2 on the Cyanobacterium Microcystis aeruginosa: Performance and Mechanism. Nanomaterials, 12(17), 3017. https://doi.org/10.3390/nano12173017