Nanoscale Zero-Valent Iron Has Minimum Toxicological Risk on the Germination and Early Growth of Two Grass Species with Potential for Phytostabilization


 , ,
, , 

Abstract
1. Introduction
2. Materials and Methods
2.1. Sources of Soil and Seeds
2.2. Germination Experiment
2.3. Hydroponics Experiment
2.4. Analytical Procedures
2.5. Statistical Analysis
3. Results and Discussion
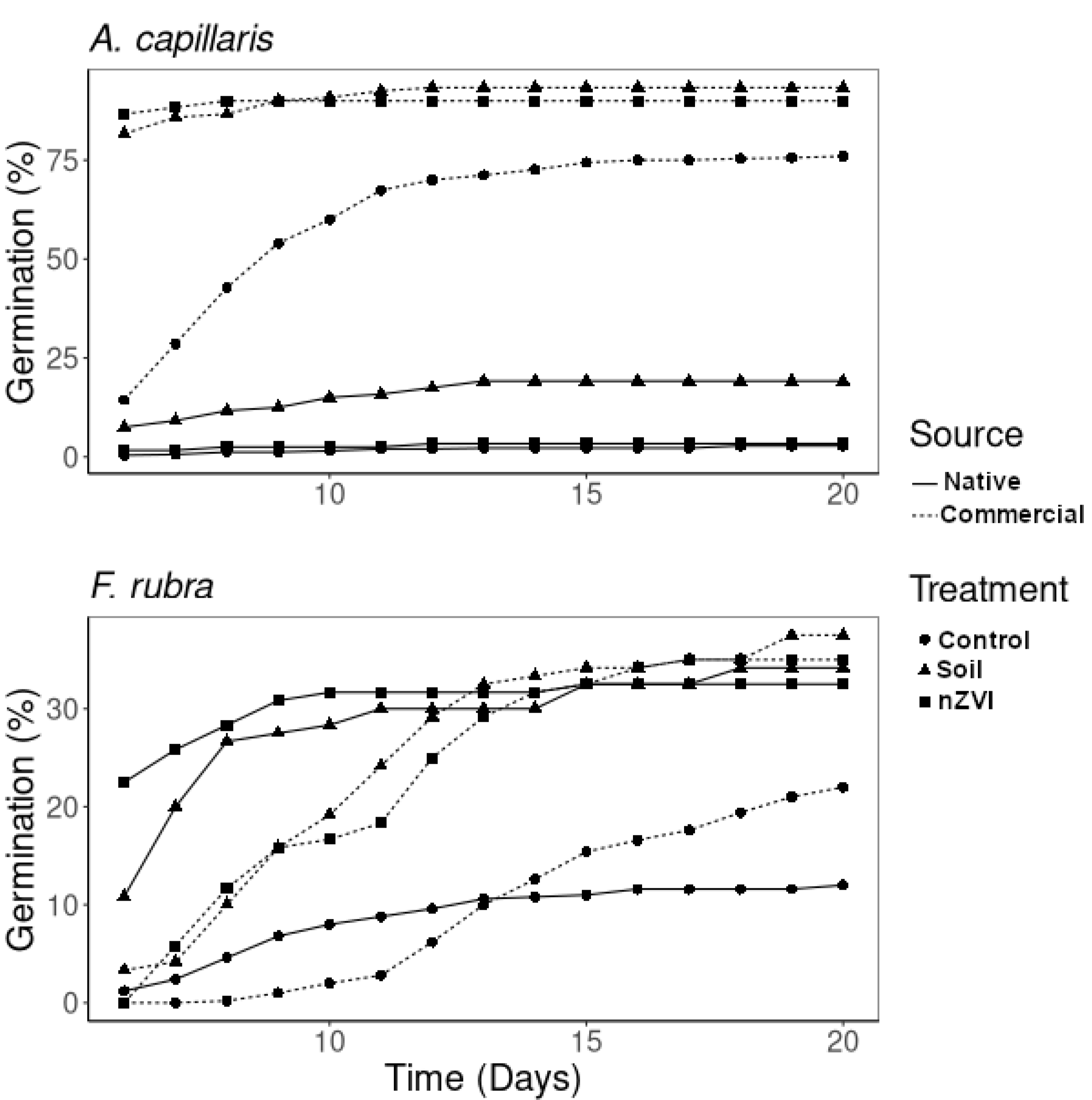
3.1. Germination Test
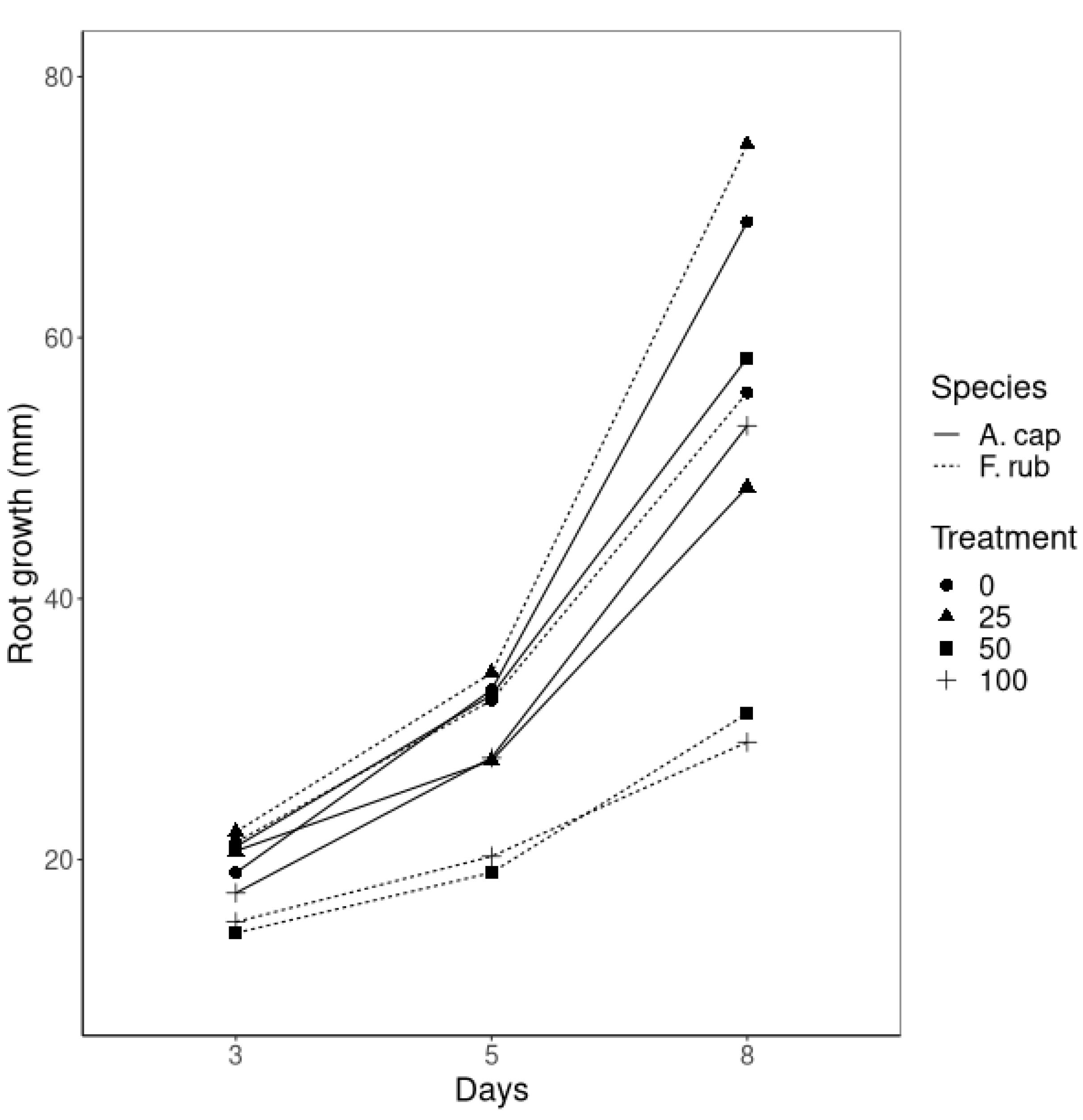
3.2. Hydroponics Experiment
3.3. Microscopic Root Observations
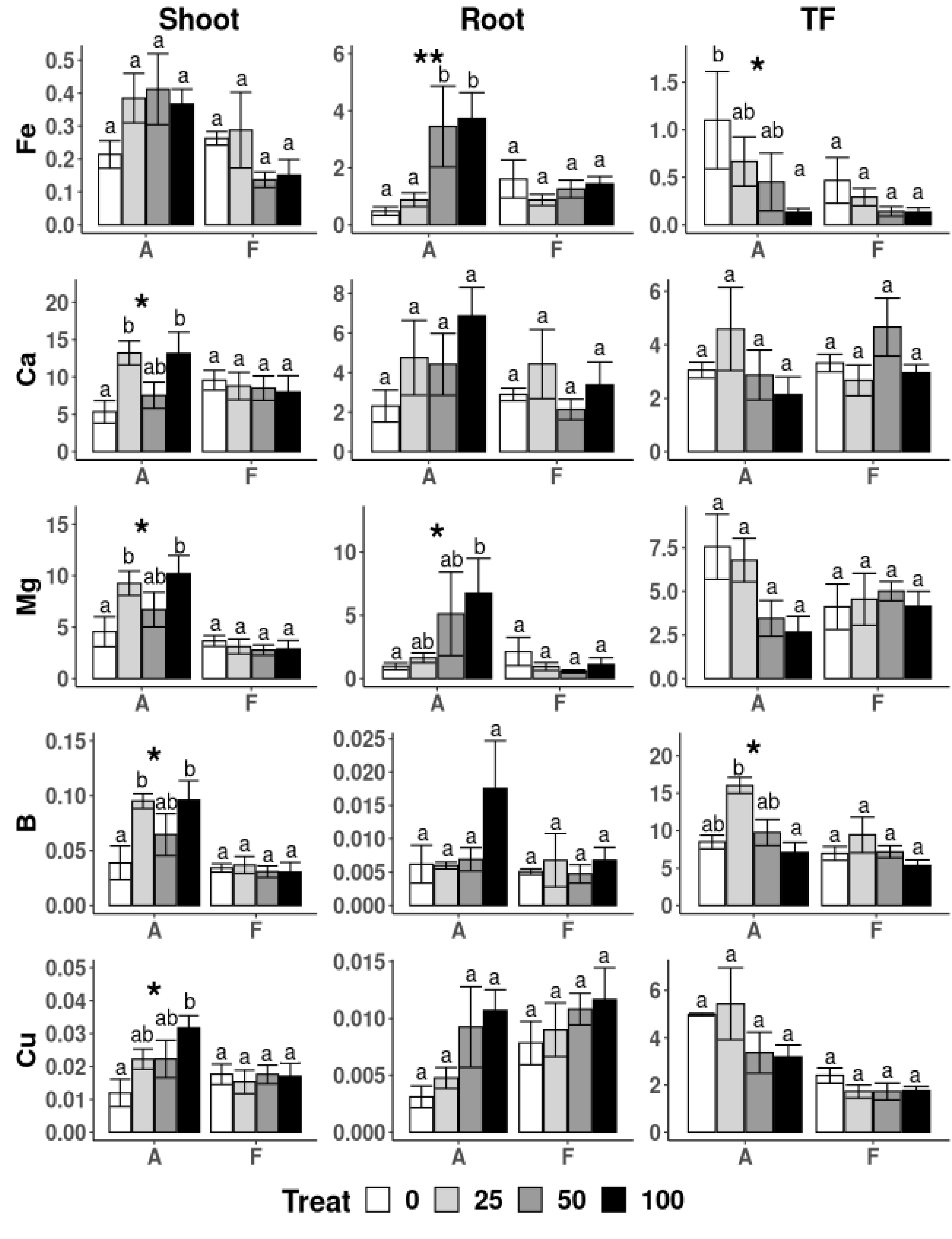
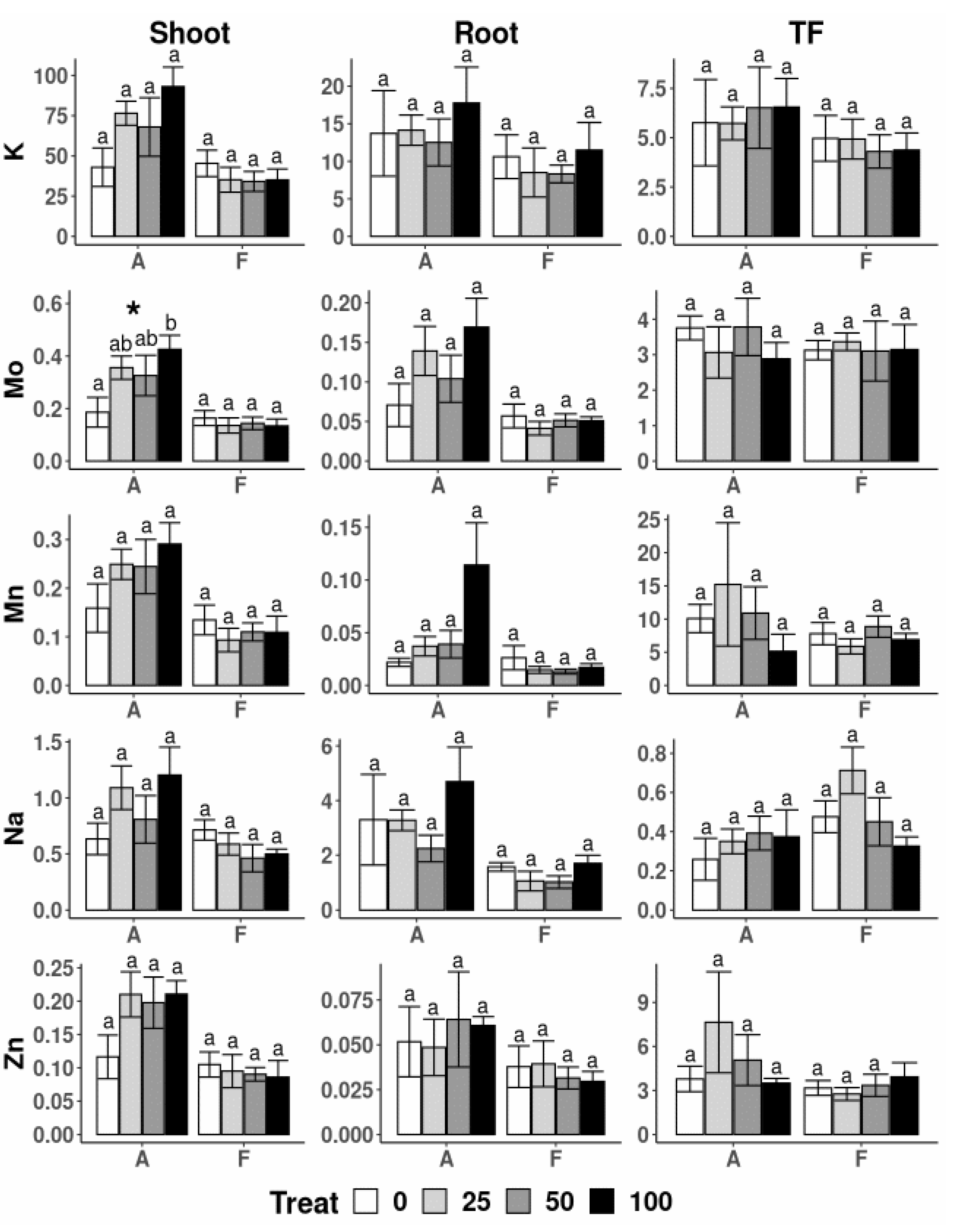
3.4. Physiological Parameters
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ali, A.; Guo, D.; Jeyasundar, P.G.S.A.; Li, Y.; Xiao, R.; Du, J.; Li, R.; Zhang, Z. Application of wood biochar in polluted soils stabilized the toxic metals and enhanced wheat (Triticum aestivum) growth and soil enzymatic activity. Ecotoxicol. Environ. Saf. 2019, 184, 109635. [Google Scholar] [CrossRef] [PubMed]
- Rinklebe, J.; Shaheen, S.M. Miscellaneous additives can enhance plant uptake and affect geochemical fractions of copper in a heavily polluted riparian grassland soil. Ecotoxicol. Environ. Saf. 2015, 119, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Grieger, K.; Fjordboge, A.; Hartmann, N.; Eriksson, E.; Bjerg, P.; Baun, A. Environmental benefits and risks of zero-valent iron nanoparticles (nZVI) for in situ remediation: Risk mitigation or trade-off? J. Contam. Hydrol. 2010, 118, 165–183. [Google Scholar] [CrossRef] [PubMed]
- Komárek, M.; Vanek, A.; Ettler, V. Chemical stabilization of metals and arsenic in contaminated soils using oxides—A review. Environ. Pollut. 2013, 172, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Stefaniuk, M.; Oleszczuk, P.; Ok, Y.S. Review on nano zerovalent iron (nZVI): From synthesis to environmental applications. Chem. Eng. J. 2016, 287, 618–632. [Google Scholar] [CrossRef]
- Lefevre, E.; Bossa, N.; Wiesner, M.R.; Gunsch, C.K. A review of the environmental implications of in situ remediation by nanoscale zero valent iron (nZVI): Behavior, transport and impacts on microbial communities. Sci. Total Environ. 2016, 565, 889–901. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Geiser-Lee, J.; Deng, Y.; Kolmakov, A. Interactions between engineered nanoparticles (ENPs) and plants: Phytotoxicity, uptake and accumulation. Sci. Total Environ. 2010, 408, 3053–3061. [Google Scholar] [CrossRef]
- Michálková, Z.; Martínez-Fernández, D.; Komárek, M. Interactions of two novel stabilizing amendments with sunflower plants grown in a contaminated soil. Chemosphere 2017, 186, 374–380. [Google Scholar] [CrossRef]
- Wu, S.; Vosatka, M.; Vogel-Mikus, K.; Kavcic, A.; Kelemen, M.; Sepec, L.; Pelicon, P.; Skala, R.; Valero Powter, A.R.; Teodoro, M.; et al. Nano zero-valent iron mediated metal(loid) uptake and translocation by arbuscular mycorrhizal symbioses. Environ. Sci. Technol. 2018, 14, 7640–7651. [Google Scholar] [CrossRef]
- Zuverza-Mena, N.; Martínez-Fernández, D.; Du, W.; Hernandez-Viezcas, J.A.; Bonilla-Bird, N.; Lopez-Moreno, M.L.; Komárek, M.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Exposure of engineered nanomaterials to plants: Insights into the physiological and biochemical responses—A review. Plant Physiol. Biochem. 2017, 110, 236–264. [Google Scholar] [CrossRef]
- Martínez-Fernández, D.; Vítková, M.; Bernal, M.P.; Komárek, M. Effects of nano-maghemite on trace element accumulation and drought response of Helianthus annuus L. in a Contaminated Mine soil. Water Air Soil Pollut. 2015, 226, 1–9. [Google Scholar] [CrossRef]
- Kumpiene, J.; Lagerkvist, A.; Maurice, C. Stabilization of As, Cr, Cu, Pb and Zn in soil using amendments? A review. Waste Manag. 2008, 28, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Lebrun, M.; Miard, F.; Nandillon, R.; Scippa, G.S.; Bourgerie, S.; Morabito, D. Biochar effect associated with compost and iron to promote Pb and As soil stabilization and Salix viminalis L. growth. Chemosphere 2019, 222, 810–822. [Google Scholar] [CrossRef]
- Sigua, G.C.; Novak, J.M.; Watts, D.W.; Ippolito, J.A.; Ducey, T.F.; Johnson, M.G.; Spokas, K.A. Phytostabilization of Zn and Cd in mine soil using corn in combination with biochars and manure-based compost. Environments 2019, 6, 69. [Google Scholar] [CrossRef]
- Doubkova, P.; Sudova, R. Limited impact of arbuscular mycorrhizal fungi on clones of Agrostis capillaris with different heavy metal tolerance. Appl. Soil Ecol. 2016, 99, 78–88. [Google Scholar] [CrossRef]
- Kucharski, R.; Sas-Nowosielska, A.; Malkowski, E.; Japenga, J.; Kuperberg, J.; Pogrzeba, M.; Krzyzak, J. The use of indigenous plant species and calcium phosphate for the stabilization of highly metal-polluted sites in southern Poland. Plant Soil 2005, 273, 291–305. [Google Scholar] [CrossRef]
- Kumpiene, J.; Antelo, J.; Brännvall, E.; Carabante, I.; Ek, K.; Komárek, M.; Söderberg, C.; Warell, L. In situ chemical stabilization of trace elementcontaminated soil—Field demonstrations and barriers to transition from laboratory to the field—A review. Appl. Geochem. 2019, 100, 335–351. [Google Scholar] [CrossRef]
- Clemente, R.; Arco-Lázaro, E.; Pardo, T.; Martıén, I.; Sánchez-Guerrero, A.; Sevilla, F.; Bernal, M. Combination of soil organic and inorganic amendments helps plants overcome trace element induced oxidative stress and allows phytostabilisation. Chemosphere 2019, 223, 223–231. [Google Scholar] [CrossRef]
- Kim, M.S.; Min, H.G.; Lee, S.H.; Kim, J.G. A comparative study on poaceae and leguminosae forage crops for aided phytostabilization in trace-element-contaminated soil. Agronomy 2018, 8, 105. [Google Scholar] [CrossRef]
- Radziemska, M.; Koda, E.; Vaverkovà, M.D.; Gusiatin, Z.M.; Cerdà, A.; Brtnický, M.; Mazur, Z. Soils from an iron and steel scrap storage yard remediated with aided phytostabilization. Land Degrad. Dev. 2018, 30, 202–211. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, X.; Shi, Z.; Tong, R.; Adams, C.A.; Shi, X. Arbuscular mycorrhizae alleviate negative effects of zinc oxide nanoparticle and zinc accumulation in maize plants e A soil microcosm experiment. Chemosphere 2016, 147, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.; Zhao, H.M.; Li, Y.W.; Huang, X.P.; Wu, X.L.; Zhai, T.; Yuan, Y.; Cai, Q.Y.; Mo, C.H. Effects of the size and morphology of zinc oxide nanoparticles on the germination of Chinese cabbage seeds. Environ. Sci. Pollut. Res. 2015, 22, 10452–10462. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Fernández, D.; Komárek, M. Comparative effects of nanoscale zero-valent iron (nzvi) and Fe2O3 nanoparticles on root hydraulic conductivity of Solanum lycopersicum L. Environ. Exp. Bot. 2016, 131, 128–136. [Google Scholar] [CrossRef]
- Wang, J.; Fang, Z.; Cheng, W.; Tsang, P.E.; Zhao, D. Ageing decreases the phytotoxicity of zero-valent iron nanoparticles in soil cultivated with Oryza sativa. Ecotoxicology 2016, 25, 1202–1210. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Gurung, A.; Deng, Y. Phytotoxicity and uptake of nanoscale zero-valent iron (nZVI) by two plant species. Sci. Total Environ. 2013, 443, 844–849. [Google Scholar] [CrossRef] [PubMed]
- Visioli, G.; Conti, F.D.; Gardi, C.; Menta, C. Germination and root elongation bioassays in six different plant species for testing Ni contamination in soil. Bull. Environ. Contam. Toxicol. 2014, 92, 490–496. [Google Scholar] [CrossRef]
- Yasur, J.; Rani, P.U. Environmental effects of nanosilver: Impact on castor seed germination, seedling growth, and plant physiology. Environ. Sci. Pollut. Res. 2013, 20, 8636–8648. [Google Scholar] [CrossRef]
- Libralato, G.; Devoti, A.C.; Zanella, M.; Sabbioni, E.; Miceti, I.; Manodori, L.; Pigozzo, A.; Manenti, S.; Groppi , F.; Ghirardini, A.V. Phytotoxicity of ionic, micro- and nano-sized iron in three plant species. Ecotoxicol. Environ. Saf. 2016, 123, 81–88. [Google Scholar] [CrossRef]
- Nguyen, N.T.; McInturf, S.A.; Mendoza-Cozat, D.G. Hydroponics: A versatile system to study nutrient allocation and plant responses to nutrient availability and exposure to toxic elements. J. Vis. Exp. 2016, 113, e54317. [Google Scholar] [CrossRef]
- Trovato, M.; Mattioli, R.; Costantino, P. Multiple roles of proline in plant stress tolerance and development. Rend. Lincei 2008, 19, 325–346. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B. Proline: A key player in plant abiotic stress tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Banerjee, R.; Goswami, P.; Lavania, S.; Mukherjee, A.; Lavania, U.C. Vetiver grass is a potential candidate for phytoremediation of iron ore mine spoil dumps. Ecol. Eng. 2019, 132, 120–136. [Google Scholar] [CrossRef]
- Gajić, G.; Mitrović, M.; Pavlović, P. Feasibility of Festuca rubra L. native grass in phytoremediation. In Phytoremediation Potential of Perennial Grasses; Elsevier: Amsterdam, The Netherlands, 2020; pp. 115–164. [Google Scholar]
- Teodoro, M.; Hejcman, M.; Vítková, M.; Wu, S.; Komárek, M. Seasonal fluctuations of Zn, Pb, As and Cd contents in the biomass of selected grass species growing on contaminated soils: Implications for in situ phytostabilization. Sci. Total Environ. 2020, 703, 134710. [Google Scholar] [CrossRef] [PubMed]
- Ettler, V.; Vaněk, A.; Mihaljevič, M.; Bezdička, P. Contrasting lead speciation in forest and tilled soils heavily polluted by lead metallurgy. Chemosphere 2005, 58, 1449–1459. [Google Scholar] [CrossRef]
- Vítková, M.; Rakosova, S.; Michálková, Z.; Komárek, M. Metal(loid)s behaviour in soils amended with nano zero-valent iron as a function of pH and time. J. Environ. Manag. 2016, 186, 268–276. [Google Scholar] [CrossRef]
- Bareke, T. Biology of seed development and germination physiology. Adv. Plants Agric. Res. 2018, 8, 336–346. [Google Scholar] [CrossRef]
- Martínez-Fernández, D.; Barroso, D.; Komárek, M. Root water transport of Helianthus annus L. under iron oxide nanoparticle exposure. Environ. Sci. Pollut. Res. 2015, 23, 1732–1741. [Google Scholar] [CrossRef]
- Bates, L.; Waldern, R.; Teare, I. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Lee, Y.; Takahashi, T. An improved clorimetric determination of aminoacids with the use of ninhydrin. Anal. Biochem. 1966, 14, 71–77. [Google Scholar] [CrossRef]
- Teodoro, M. Rompimiento de Latencia en Semillas de Mimosa Lacerata Rose. Master’s Thesis, Universidad Autonoma Chapingo, Texcoco, Mexico, 2011. [Google Scholar]
- Munzuroglu, O.; Geckil, H. Effects of metals on seed germination, root elongation, and coleoptile and hypocotyl growth in Triticum aestivum and Cucumis sativus. Arch. Environ. Contam. Toxicol. 2002, 43, 203–213. [Google Scholar] [CrossRef]
- Savithramma, N.; Ankanna, S.; Bhumi, G. Effect of nanoparticles on seed germination and seedling growth of Boswellia ovalifoliolata—An endemic and endangered medicinal tree taxon. Nano Vis. 2012, 2, 61–68. [Google Scholar]
- Vítková, M.; Puschenreiter, M.; Komárek, M. Effect of nano zero-valent iron application on As, Cd, Pb, and Zn availability in the rhizosphere of metal(loid) contaminated soils. Chemosphere 2018, 200, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Mitzia, A.; Vítková, M.; Komárek, M. Assessment of biochar and/or nano zero-valent iron for the stabilisation of Zn, Pb and Cd: A temporal study of solid phase geochemistry under changing soil conditions. Chemosphere 2020, 242, 125248. [Google Scholar] [CrossRef]
- Zhan, Y.; Wu, X.; Lin, J.; Zhang, Z.; Zhao, Y.; Yu, Y.; Wang, Y. Combined use of calcium nitrate addition and anion exchange resin capping to control sedimentary phosphorus release and its nitrate-nitrogen releasing risk. Sci. Total Environ. 2019, 689, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Sharmila, P.; Kumari, P.K.; Singh, K.; Prasad, N.V.S.R.K.; Pardha-Saradhi, P. Cadmium toxicity-induced proline accumulation is coupled to iron depletion. Protoplasma 2017, 254, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zeng, B.; Sun, Z.; Zhu, C. Relationship between proline and Hg2+ -induced oxidative stress in a tolerant rice mutant. Arch. Environ. Contam. Toxicol. 2008, 56, 723–731. [Google Scholar] [CrossRef]
- Ullah, R.; Hadi, F.; Ahmad, S.; Jan, A.U.; Rongliang, Q. Phytoremediation of lead and chromium contaminated soil improves with the endogenous phenolics and proline production in parthenium, cannabis, euphorbia, and rumex species. Water Air Soil Pollut. 2019, 230, 40. [Google Scholar] [CrossRef]
- Teodoro, M.; Trakal, L.; Gallagher, B.N.; Šimek, P.; Soudek, P.; Pohořelý, M.; Beesley, L.; Jačka, L.; Kovář, M.; Seyedsadr, S.; et al. Application of co-composted biochar significantly improved plant-growth relevant physical/chemical properties of a metal contaminated soil. Chemosphere 2020, 242, 125255. [Google Scholar] [CrossRef]
- Kobayashi, T.; Nozoye, T.; Nishizawa, N.K. Iron transport and its regulation in plants. Free Radic. Biol. Med. 2019, 133, 11–20. [Google Scholar] [CrossRef]
- Zandi, P.; Yang, J.; Xia, X.; Tian, Y.; Li, Q.; Mozdzen, K.; Barabasz-Krasny, B.; Wang, Y. Do sulfur addition and rhizoplane iron plaque affect chromium uptake by rice (Oryza sativa L.) seedlings in solution culture? J. Hazard. Mater. 2020, 388, 121803. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, F.; Mao, D. Effect of iron plaque outside roots on nutrient uptake by rice (Oryza sativa L.): Phosphorus uptake. Plant Soil 1999, 209, 187–192. [Google Scholar] [CrossRef]
- De Araujo, T.O.; Isaure, M.P.; Alchoubassi, G.; Bierla, K.; Szpunar, J.; Trcera, N.; Chay, S.; Alcon, C.; da Silva, L.C.; Curie, C.; et al. Paspalum urvillei and Setaria parviflora, two grasses naturally adapted to extreme iron-rich environments. Plant Physiol. Biochem. 2020, 151, 144–156. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Details | 1 Bulk Soil | 2 Rhizosphere |
|---|---|---|
| Clay | 5% | - |
| Silt | 20% | - |
| Sand | 75% | - |
| pH-H2O | 5.95 ± 0.01 | 5.62 ± 0.27 |
| pH-KCl | 5.14 ± 0.03 | 5.37 ± 0.37 |
| K | 6583 ± 293 | 5534 ± 716 |
| Ca | 1778 ± 104 | 2463 ± 655 |
| S | 490 ± 6 | 410 ± 154 |
| Cu | 71.9 ± 3.0 | 84 ± 12 |
| Mn | 4276 ± 28 | 1857 ± 36,728 |
| Fe | 37,408 ± 159 | 23,930 ± 2684 |
| Zn | 4002 ± 55 | 2910 ± 493 |
| Pb | 3539 ± 30 | 2796 ± 425 |
| As | 296 ± 5 | 237 ± 48 |
| Cd | 39 ± 0.1 | 36 ± 6 |
| Source | Treat | Germ | Root | Shoot | RGR | EIR | GI |
|---|---|---|---|---|---|---|---|
| (%) | (cm) | (cm) | |||||
| Agrostis capillaris | |||||||
| Native | C | a 2.8 ± 2.1 | c 2.5 ± 1.7 | b 1.6 ± 0.5 | 1.00 | 0.00 | 100 |
| Soil | b 19.2 ± 3.8 | a 0.3 ± 0.2 | a 1 ± 0.8 | 6.85 | 90.7 | 63.4 | |
| Soil + nZVI | a 3.3 ± 4.1 | b 0.5 ± 0.38 | b 1.8 ± 0.7 | 1.19 | 80.1 | 23.6 | |
| nZVI effect | 0.17 | −114 | 37.3 | ||||
| Commercial | C | c 76 ± 6.7 | c 3.7 ± 1.1 | b 2 ± 0.5 | 1.00 | 0.00 | 100 |
| Soil | d 91.7 ± 7.5 | b 0.8 ± 0.3 | c 3.5 ± 0.9 | 1.21 | 78.3 | 26.1 | |
| Soil + nZVI | d 90 ± 10 | b 0.8 ± 0.3 | c 3.2 ± 0.7 | 1.18 | 79.0 | 24.9 | |
| nZVI effect | 0.98 | 2.89 | 95.3 | ||||
| Festuca rubra | |||||||
| Native | C | a 12.0 ± 2.8 | bc 3.0 ± 1.7 | a 4.2 ± 4.2 | 1.00 | 0.00 | 100 |
| Soil | b 34.2 ± 15.3 | abc 2.4 ± 1.4 | c 6.9 ± 2.3 | 2.85 | 19.4 | 229.6 | |
| Soil + nZVI | b 32.5 ± 12.9 | ab 2.0 ± 0.9 | c 7.6 ± 2.3 | 2.71 | 32.2 | 183.6 | |
| nZVI effect | 0.95 | 15.9 | 79.9 | ||||
| Commercial | C | a 22.0 ± 8.5 | c 3.3 ± 1.3 | a 3.5 ± 1.2 | 1.00 | 0.00 | 100 |
| Soil | b 37.5 ± 19.2 | a 1.7 ± 1.0 | ab 4.6 ± 2.1 | 1.70 | 48.8 | 87.3 | |
| Soil + nZVI | b 35 ± 11.4 | abc 2.4 ± 0.4 | bc 6.9 ± 1.3 | 1.59 | 27.8 | 114.8 | |
| nZVI effect | 0.93 | −40.9 | 131.4 | ||||
| T | Part | Fresh Weight | Dry Weight | Aminoacids | Proline |
|---|---|---|---|---|---|
| (g) | (g) | μM g−1 | μM g−1 | ||
| A. capillaris | |||||
| 0 | Shoot | a 8.72 ± 1.7 | a 2.17 ± 1.56 | a 147 ± 44 | a 5.21 ± 2.48 |
| 25 | bc 21.6 ± 4.4 | a 4.13 ± 0.85 | a 102 ± 64 | a 4.02 ± 2.36 | |
| 50 | ab 12.9 ± 5.0 | a 3.48 ± 2.12 | a 167 ± 58 | a 5.42 ± 1.57 | |
| 100 | c 26.3 ± 9.4 | a 5.08 ± 1.58 | a 155 ± 36 | a 3.4 ± 0.88 | |
| 0 | Root | a 5.11 ± 1.1 | a 0.43 ± 0.09 | a 72 ± 16 | a 2.71 ± 2.68 |
| 25 | b 16.8 ± 3.4 | ab 1.42 ± 0.2 | a 76 ± 10 | a 4.53 ± 2.11 | |
| 50 | ab 15.9 ± 10.2 | ab 1.33 ± 1 | a 74 ± 13 | a 4.44 ± 2.94 | |
| 100 | b 19.4 ± 6.4 | b 1.67 ± 0.48 | a 78 ± 23 | a 3.86 ± 2.53 | |
| F. rubra | |||||
| 0 | Shoot | a 12.21 ± 5.1 | a 2.16 ± 0.76 | a 161 ± 36 | a 5.93 ± 2.77 |
| 25 | a 9.63 ± 4.7 | a 1.87 ± 0.8 | a 159 ± 34 | a 4.31 ± 1.47 | |
| 50 | a 8.88 ± 3.3 | a 1.78 ± 0.61 | a 158 ± 58 | a 3.21 ± 1.08 | |
| 100 | a 10.05 ± 5.1 | a 2 ± 1.04 | a 142 ± 30 | a 4.42 ± 0.85 | |
| 0 | Root | a 9.81 ± 3.7 | a 0.75 ± 0.26 | a 49 ± 6 | a 3.48 ± 2.73 |
| 25 | a 8.43 ± 4.6 | a 0.7 ± 0.32 | a 76 ± 47 | a 3.46 ± 2.62 | |
| 50 | a 8.40 ± 2.9 | a 0.72 ± 0.27 | a 46 ± 13 | a 2.77 ± 2.86 | |
| 100 | a 8.71 ± 5.6 | a 0.77 ± 0.48 | a 66 ± 34 | a 3.63 ± 2.8 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teodoro, M.; Clemente, R.; Ferrer-Bustins, E.; Martínez-Fernández, D.; Pilar Bernal, M.; Vítková, M.; Vítek, P.; Komárek, M. Nanoscale Zero-Valent Iron Has Minimum Toxicological Risk on the Germination and Early Growth of Two Grass Species with Potential for Phytostabilization. Nanomaterials 2020, 10, 1537. https://doi.org/10.3390/nano10081537
Teodoro M, Clemente R, Ferrer-Bustins E, Martínez-Fernández D, Pilar Bernal M, Vítková M, Vítek P, Komárek M. Nanoscale Zero-Valent Iron Has Minimum Toxicological Risk on the Germination and Early Growth of Two Grass Species with Potential for Phytostabilization. Nanomaterials. 2020; 10(8):1537. https://doi.org/10.3390/nano10081537
Chicago/Turabian StyleTeodoro, Manuel, Rafael Clemente, Ermengol Ferrer-Bustins, Domingo Martínez-Fernández, Maria Pilar Bernal, Martina Vítková, Petr Vítek, and Michael Komárek. 2020. "Nanoscale Zero-Valent Iron Has Minimum Toxicological Risk on the Germination and Early Growth of Two Grass Species with Potential for Phytostabilization" Nanomaterials 10, no. 8: 1537. https://doi.org/10.3390/nano10081537
APA StyleTeodoro, M., Clemente, R., Ferrer-Bustins, E., Martínez-Fernández, D., Pilar Bernal, M., Vítková, M., Vítek, P., & Komárek, M. (2020). Nanoscale Zero-Valent Iron Has Minimum Toxicological Risk on the Germination and Early Growth of Two Grass Species with Potential for Phytostabilization. Nanomaterials, 10(8), 1537. https://doi.org/10.3390/nano10081537