
Human Keratoconus Cell Contractility is Mediated by Transforming Growth Factor-Beta Isoforms


Abstract
:1. Introduction
2. Results and Discussion
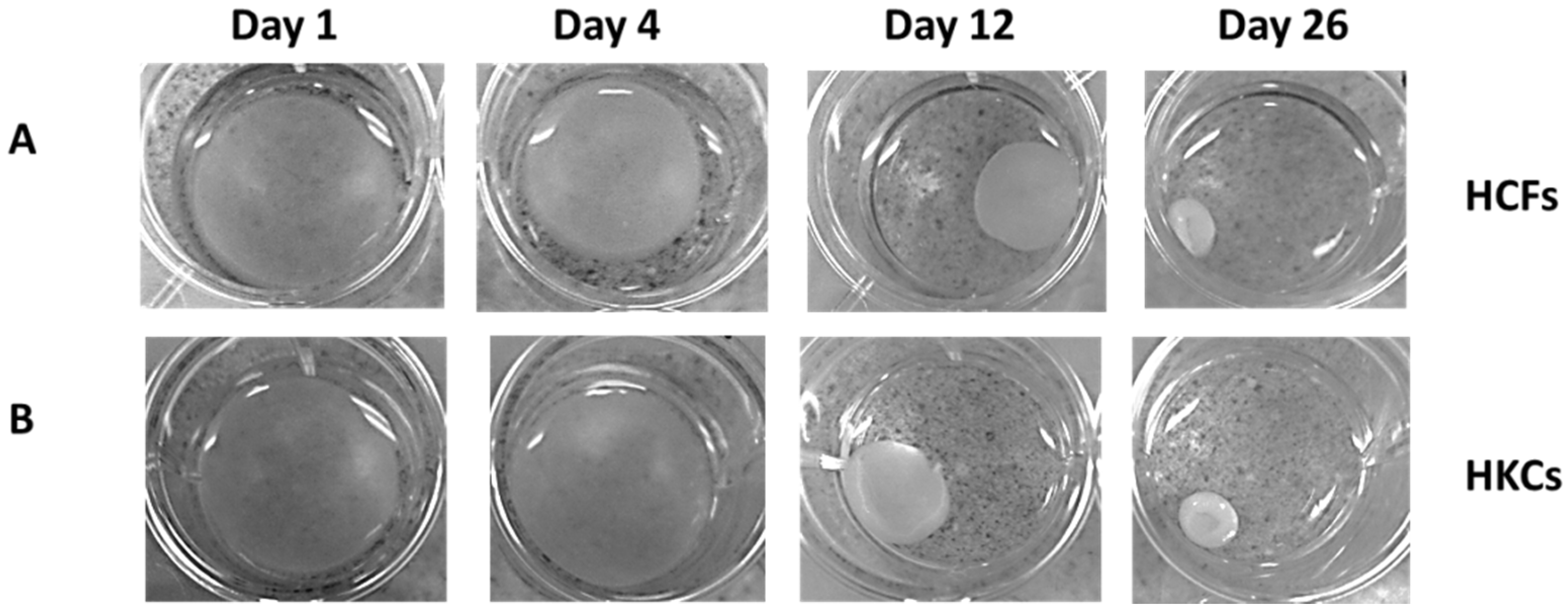
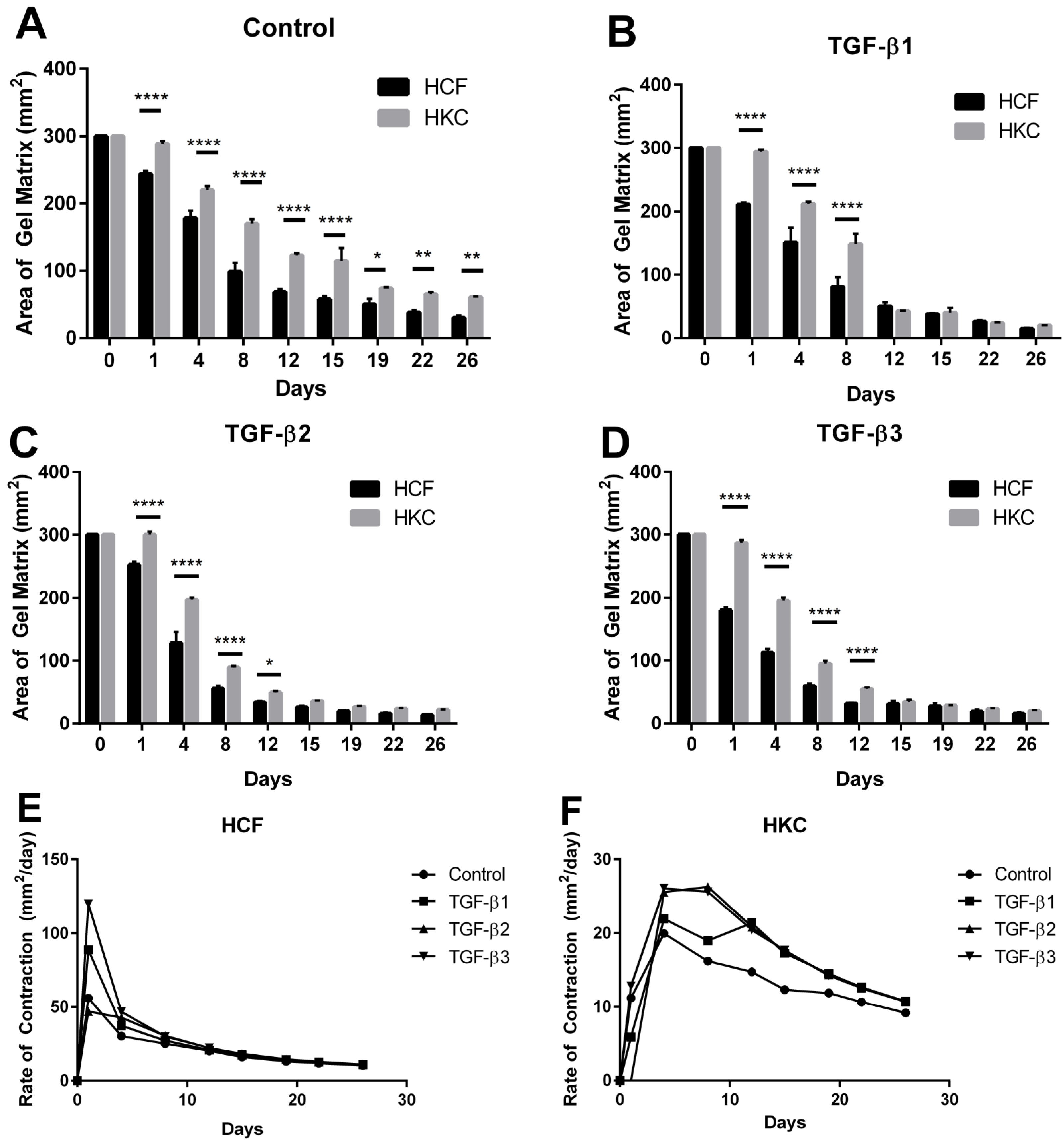
2.1. Contraction Profiles of HCFs and HKCs
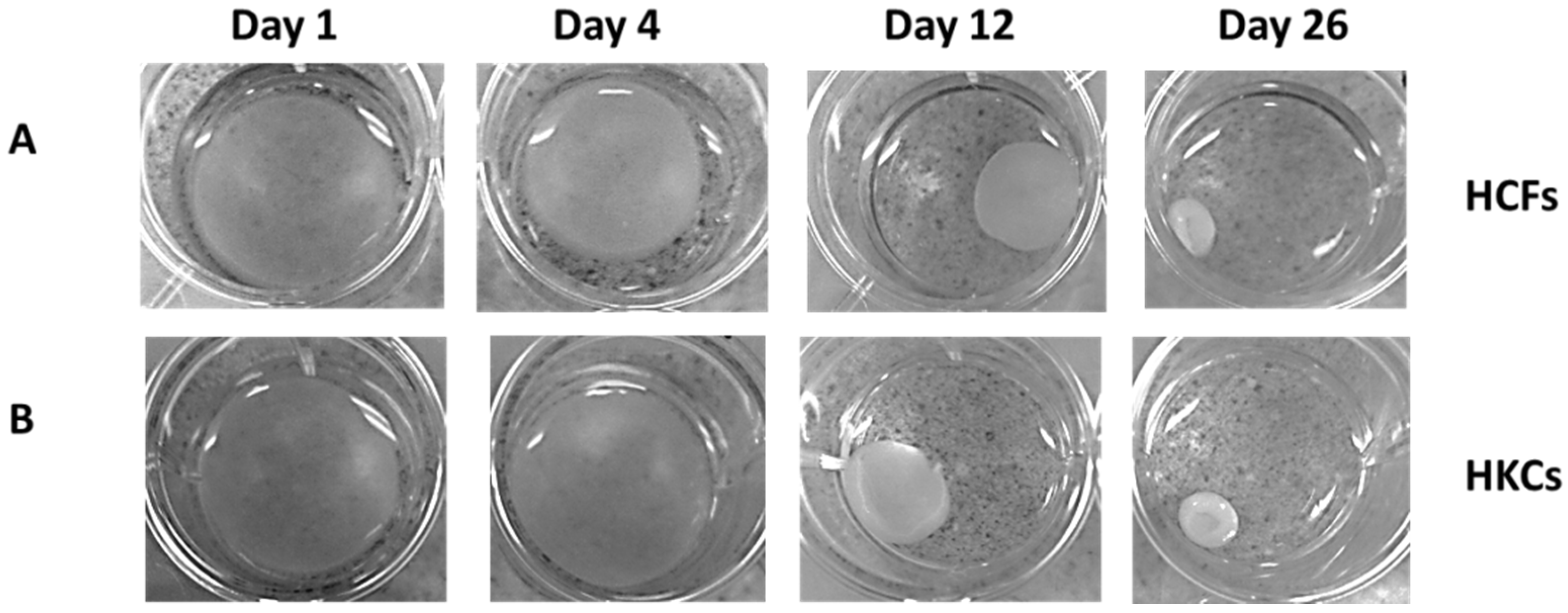
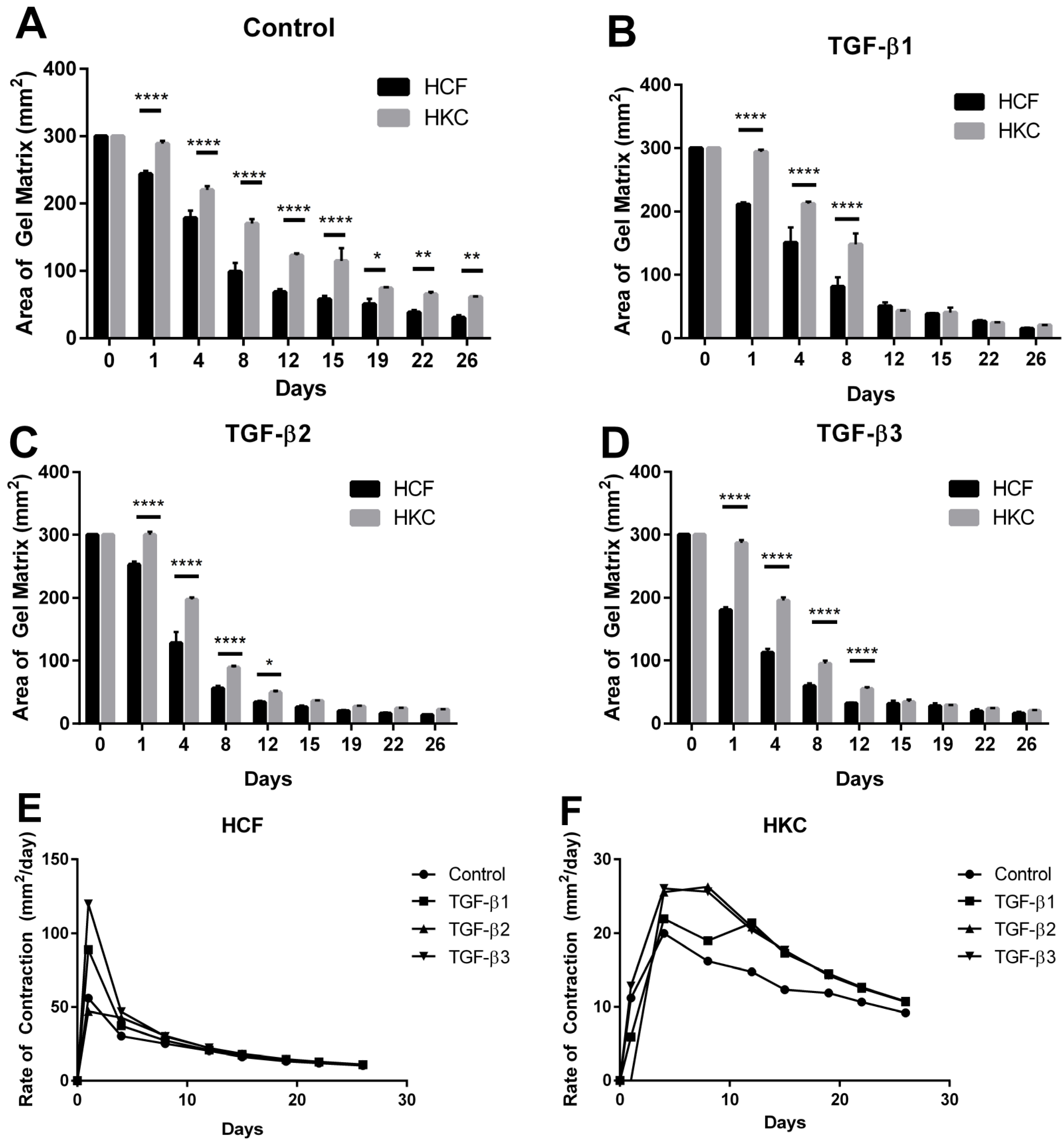
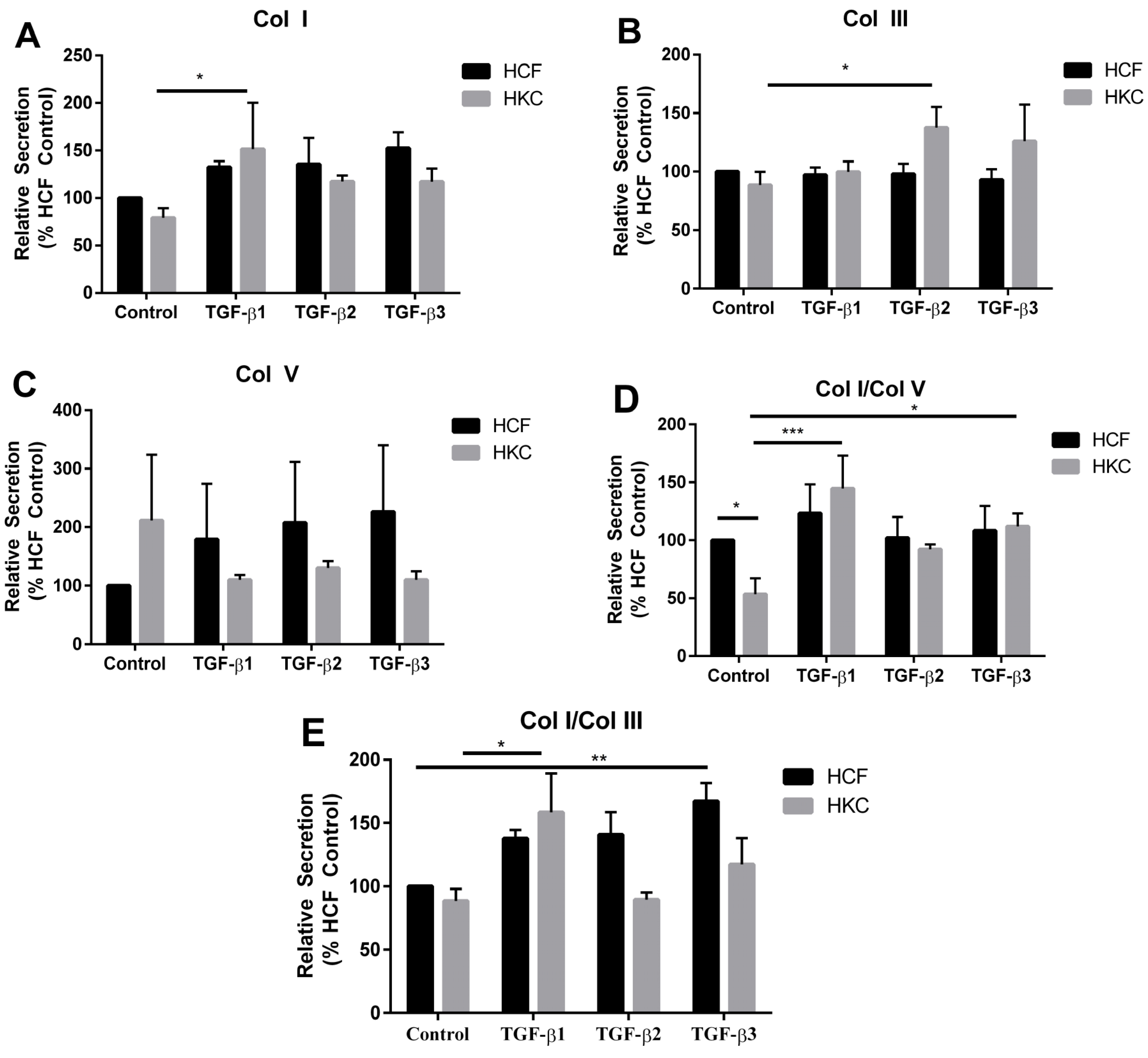
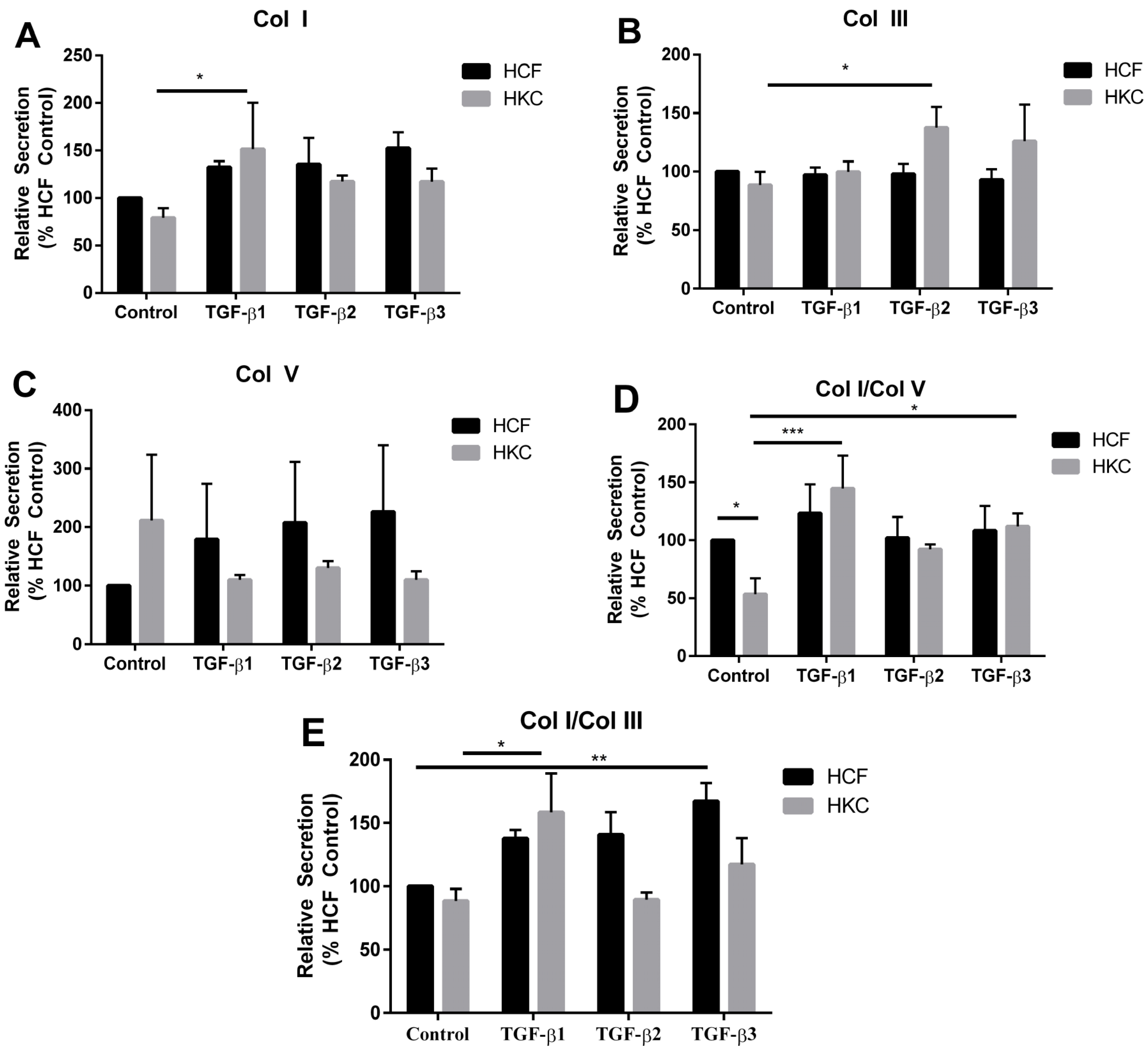
2.2. Collagen Secretion by HCFs and HKCs
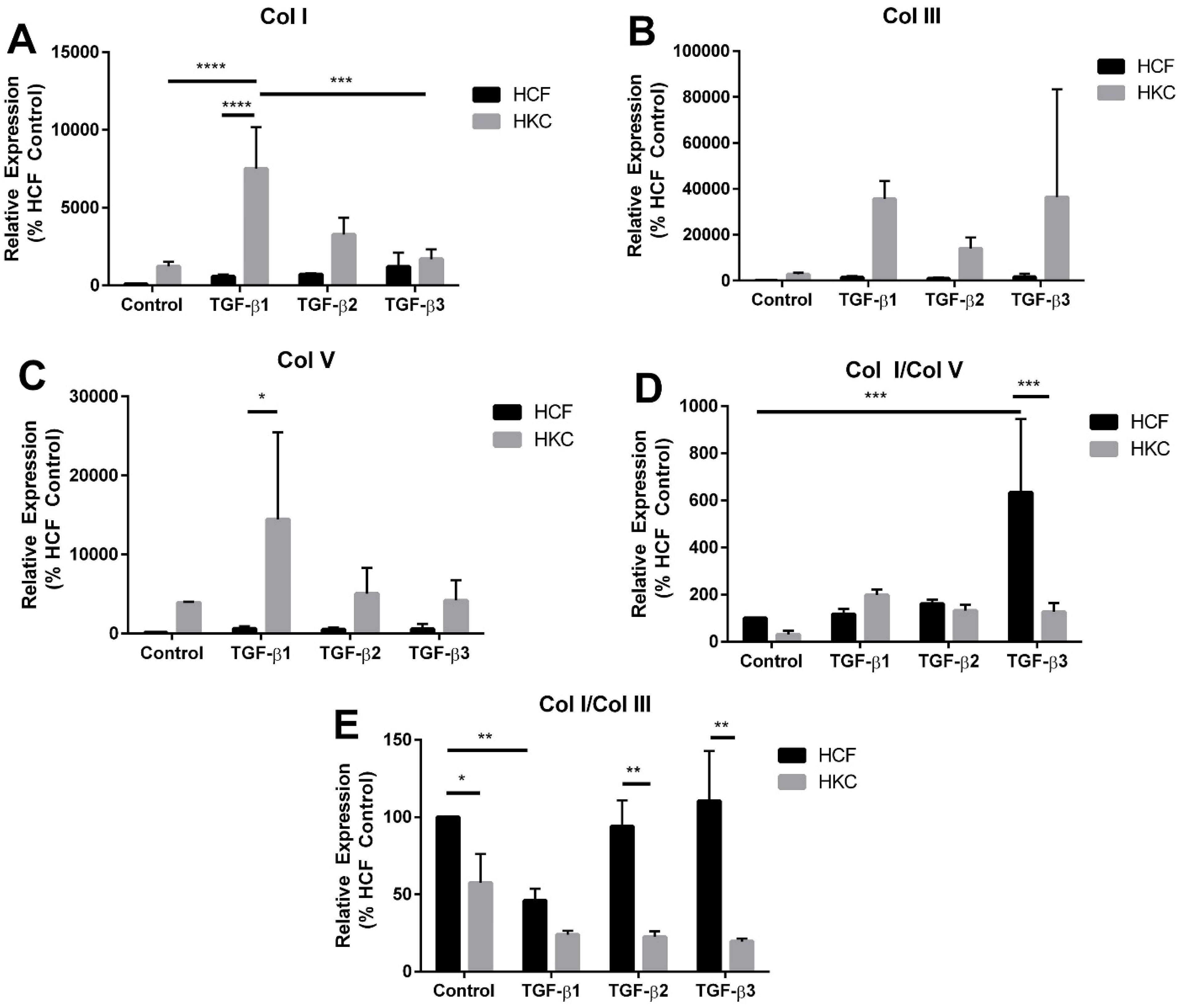
2.3. mRNA Expression of Collagen I, III, and V by HCFs and HKCs
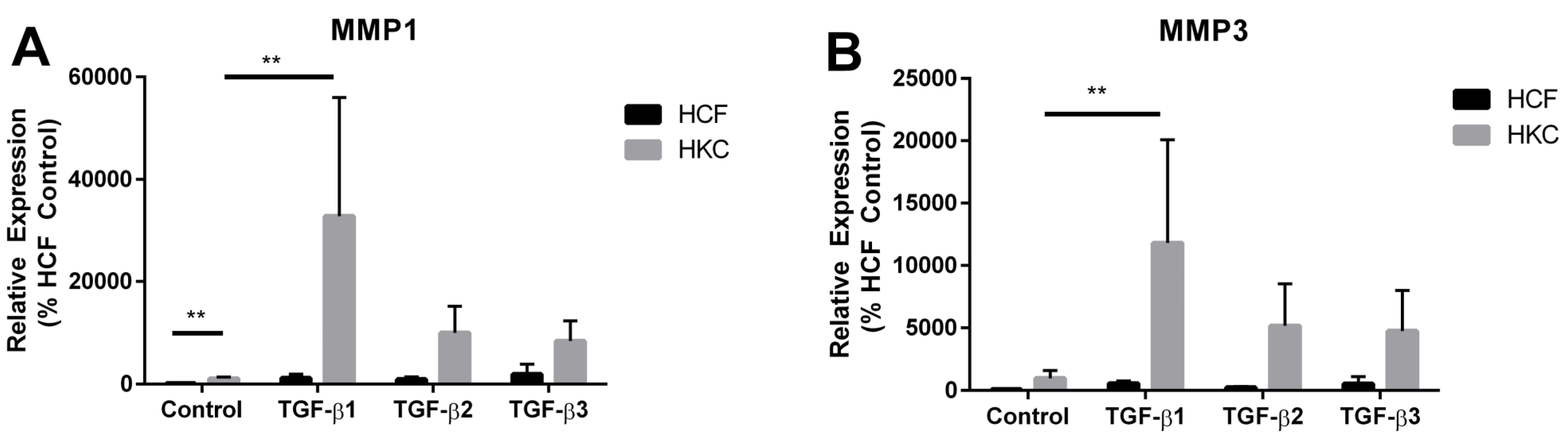
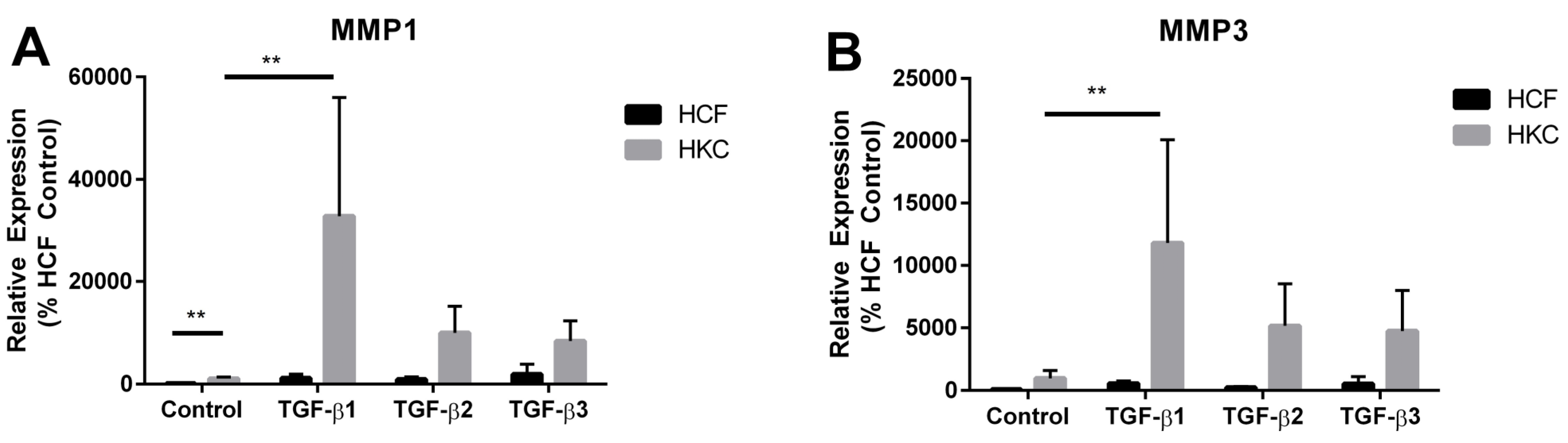
2.4. MMP1 and MMP3 Expression by HCFs and HKCs
3. Experimental Section
3.1. Cell Culture
3.2. Collagen Contraction Assay
3.3. RT-PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probe | Catalogue # | Company | Final Concentration |
|---|---|---|---|
| GAPDH | Hs99999905_m1 | Life Technologies | 1× |
| 18S | Hs99999901_s1 | Life Technologies | 1× |
| Col I | Hs00164004_m1 | Life Technologies | 1× |
| Col III | Hs00943809_m1 | Life Technologies | 1× |
| Col V | Hs00609133_m1 | Life Technologies | 1× |
| MMP 1 | Hs00899658_m1 | Life Technologies | 1× |
| MMP 3 | Hs00968305_m1 | Life Technologies | 1× |
3.4. Western Blot
| Antibody | Catalogue # | Company | Dilution |
|---|---|---|---|
| Col I | ab34710 | Abcam, Cambridge, MA, USA | 1/1000 |
| Col III | ab7778 | Abcam, Cambridge, MA, USA | 1/1000 |
| Col V | ab94673 | Abcam, Cambridge, MA,USA | 1/1000 |
3.5. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ambekar, R.; Toussaint, K.C., Jr.; Wagoner Johnson, A. The effect of keratoconus on the structural, mechanical, and optical properties of the cornea. J. Mech. Behav. Biomed. Mater. 2011, 4, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, R.H.; Bourne, W.M.; Dyer, J.A. A 48-year clinical and epidemiologic study of keratoconus. Am. J. Ophthalmol. 1986, 101, 267–273. [Google Scholar] [CrossRef]
- Ertan, A.; Muftuoglu, O. Keratoconus clinical findings according to different age and gender groups. Cornea 2008, 27, 1109–1113. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, J.L.O.; Jurado, J.C.G.; Rodriguez, F.J.B.; Laborda, D.S. Keratoconus: Age of onset and natural history. Optom. Vis. Sci. 1997, 74, 147–151. [Google Scholar] [CrossRef]
- Romero-Jimenez, M.; Santodomingo-Rubido, J.; Wolffsohn, J.S. Keratoconus: A review. Cont. Lens. Anterior. Eye. 2010, 33, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Ghanem, R.C.; Santhiago, M.R.; Berti, T.; Netto, M.V.; Ghanem, V.C. Topographic, corneal wavefront, and refractive outcomes 2 years after collagen crosslinking for progressive keratoconus. Cornea 2014, 33, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Greenstein, S.A.; Fry, K.L.; Hersh, P.S. Corneal topography indices after corneal collagen crosslinking for keratoconus and corneal ectasia: One-year results. J. Cataract Refract. Surg. 2011, 37, 1282–1290. [Google Scholar] [CrossRef] [PubMed]
- Lesniak, S.P.; Hersh, P.S. Transepithelial corneal collagen crosslinking for keratoconus: Six-month results. J. Cataract Refract. Surg. 2014, 40, 1971–1979. [Google Scholar] [CrossRef] [PubMed]
- Karamichos, D.; Zareian, R.; Guo, X.; Hutcheon, A.E.; Ruberti, J.W.; Zieske, J.D. Novel model for keratoconus disease. J. Funct. Biomater. 2012, 3, 760–775. [Google Scholar] [CrossRef] [PubMed]
- Karamichos, D.; Hutcheon, A.E.; Rich, C.B.; Trinkaus-Randall, V.; Asara, J.M.; Zieske, J.D. In vitro model suggests oxidative stress involved in keratoconus disease. Sci. Rep. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Kropp, B.P.; Zhang, Y.; Tomasek, J.J.; Cowan, R.; Furness, P.D.; Vaughan, M.B.; Parizi, M.; Cheng, E.Y. Characterization of cultured bladder smooth muscle cells: Assessment of in vitro contractility. J. Urol. 1999, 162, 1779–1784. [Google Scholar] [CrossRef]
- Kimura, K.; Orita, T.; Fujitsu, Y.; Liu, Y.; Wakuta, M.; Morishige, N.; Suzuki, K.; Sonoda, K.H. Inhibition by female sex hormones of collagen gel contraction mediated by retinal pigment epithelial cells. Invest. Ophthalmol. Vis. Sci. 2014, 55, 2621–2630. [Google Scholar] [CrossRef] [PubMed]
- Pilcher, B.K.; Kim, D.W.; Carney, D.H.; Tomasek, J.J. Thrombin stimulates fibroblast-mediated collagen lattice contraction by its proteolytically activated receptor. Exp. Cell Res. 1994, 211, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Bell, E.; Ivarsson, B.; Merrill, C. Production of a tissue-like structure by contraction of collagen lattices by human fibroblasts of different proliferative potential in vitro. Proc. Natl. Acad. Sci. USA 1979, 76, 1274–1278. [Google Scholar] [CrossRef] [PubMed]
- Levi-Schaffer, F.; Garbuzenko, E.; Rubin, A.; Reich, R.; Pickholz, D.; Gillery, P.; Emonard, H.; Nagler, A.; Maquart, F.A. Human eosinophils regulate human lung- and skin-derived fibroblast properties in vitro: A role for transforming growth factor beta (TGF-beta). Proc. Natl. Acad. Sci. USA 1999, 96, 9660–9665. [Google Scholar] [CrossRef] [PubMed]
- Witte, M.B.; Barbul, A. General principles of wound healing. Surg. Clin. North Am. 1997, 77, 509–528. [Google Scholar] [CrossRef]
- Montesano, R.; Orci, L. Transforming growth factor beta stimulates collagen-matrix contraction by fibroblasts: Implications for wound healing. Proc. Natl. Acad. Sci. USA 1988, 85, 4894–4897. [Google Scholar] [CrossRef] [PubMed]
- Germain, L.; Jean, A.; Auger, F.A.; Garrel, D.R. Human wound healing fibroblasts have greater contractile properties than dermal fibroblasts. J. Surg. Res. 1994, 57, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Diegelmann, R.F.; Evans, M.C. Wound healing: An overview of acute, fibrotic and delayed healing. Front. Biosci. 2004, 9, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Werner, S.; Grose, R. Regulation of wound healing by growth factors and cytokines. Physiol. Rev. 2003, 83, 835–870. [Google Scholar] [PubMed]
- Cheung, I.M.; McGhee, C.N.; Sherwin, T. A new perspective on the pathobiology of keratoconus: Interplay of stromal wound healing and reactive species-associated processes. Clin. Exp. Optom. 2013, 96, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Cheung, I.M.; McGhee, C.; Sherwin, T. Deficient repair regulatory response to injury in keratoconic stromal cells. Clin. Exp. Optom. 2014, 97, 234–239. [Google Scholar] [CrossRef] [PubMed]
- McMonnies, C.W. Mechanisms of rubbing-related corneal trauma in keratoconus. Cornea 2009, 28, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Segev, F.; Heon, E.; Cole, W.G.; Wenstrup, R.J.; Young, F.; Slomovic, A.R.; Rootman, D.S.; Whitaker-Menezes, D.; Chervoneva, I.; Birk, D.E. Structural abnormalities of the cornea and lid resulting from collagen v mutations. Invest. Ophthalmol. Vis. Sci. 2006, 47, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M.K.; Foley, J.W.; Birk, D.E.; Fitch, J.M.; Linsenmayer, T.F. Type v collagen and bowman’s membrane. Quantitation of mrna in corneal epithelium and stroma. J. Biol. Chem. 1994, 269, 24959–24966. [Google Scholar] [PubMed]
- Sun, M.; Chen, S.; Adams, S.M.; Florer, J.B.; Liu, H.; Kao, W.W.; Wenstrup, R.J.; Birk, D.E. Collagen v is a dominant regulator of collagen fibrillogenesis: Dysfunctional regulation of structure and function in a corneal-stroma-specific col5a1-null mouse model. J. Cell Sci. 2011, 124, 4096–4105. [Google Scholar] [CrossRef] [PubMed]
- Ruberti, J.W.; Roy, A.S.; Roberts, C.J. Corneal biomechanics and biomaterials. Annu. Rev. Biomed. Eng. 2011, 13, 269–295. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, S.; Bron, A.J.; Salvi, S.M.; Hawksworth, N.R.; Tuft, S.J.; Meek, K.M. Ultrastructural analysis of collagen fibrils and proteoglycans in keratoconus. Acta Ophthalmol. (Copenh.) 2008, 86, 764–772. [Google Scholar] [CrossRef] [PubMed]
- Meek, K.M.; Tuft, S.J.; Huang, Y.; Gill, P.S.; Hayes, S.; Newton, R.H.; Bron, A.J. Changes in collagen orientation and distribution in keratoconus corneas. Invest. Ophthalmol. Vis. Sci. 2005, 46, 1948–1956. [Google Scholar] [CrossRef] [PubMed]
- Chaerkady, R.; Shao, H.; Scott, S.-G.; Pandey, A.; Jun, A.S.; Chakravarti, S. The keratoconus corneal proteome: Loss of epithelial integrity and stromal degeneration. J. Proteomics 2013, 87, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Delaigue, O.; Arbeille, B.; Lemesle, M.; Roingeard, P.; Rossazza, C. Quantitative analysis of immunogold labellings of collagen types I, III, IV and Vi in healthy and pathological human corneas. Graefe’s Arch. Clin. Exp. Ophthalmol. 1995, 233, 331–338. [Google Scholar] [CrossRef]
- McKay, T.B.; Lyon, D.; Sarker-Nag, A.; Priyadarsini, S.; Asara, J.M.; Karamichos, D. Quercetin attenuates lactate production and extracellular matrix secretion in keratoconus. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Bykhovskaya, Y.; Canedo, A.L.; Haritunians, T.; Siscovick, D.; Aldave, A.J.; Szczotka-Flynn, L.; Iyengar, S.K.; Rotter, J.I.; Taylor, K.D.; et al. Genetic association of COL5A1 variants in keratoconus patients suggests a complex connection between corneal thinning and keratoconus. Invest. Ophthalmol. Vis. Sci. 2013, 54, 2696–2704. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.A.; McCoy, G.A.; Folkvord, J.M.; McPherson, J.M. TGF-beta 1 stimulates cultured human fibroblasts to proliferate and produce tissue-like fibroplasia: A fibronectin matrix-dependent event. J. Cell. Physiol. 1997, 170, 69–80. [Google Scholar] [CrossRef]
- Roberts, C.J.; Birkenmeier, T.M.; McQuillan, J.J.; Akiyama, S.K.; Yamada, S.S.; Chen, W.T.; Yamada, K.M.; McDonald, J.A. Transforming growth factor beta stimulates the expression of fibronectin and of both subunits of the human fibronectin receptor by cultured human lung fibroblasts. J. Biol. Chem. 1988, 263, 4586–4592. [Google Scholar] [PubMed]
- Zhou, L.; Lopes, J.E.; Chong, M.M.; Ivanov, I.I.; Min, R.; Victora, G.D.; Shen, Y.; Du, J.; Rubtsov, Y.P.; Rudensky, A.Y.; et al. TGF-beta-induced Foxp3 inhibits T(H)17 cell differentiation by antagonizing rorgammat function. Nature 2008, 453, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Massague, J.; Xi, Q. TGF-beta control of stem cell differentiation genes. FEBS Lett. 2012, 586, 1953–1958. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.S.; Huang, J.S. TGF-beta control of cell proliferation. J. Cell. Biochem. 2005, 96, 447–462. [Google Scholar] [CrossRef] [PubMed]
- Connor, T.B., Jr.; Roberts, A.B.; Sporn, M.B.; Danielpour, D.; Dart, L.L.; Michels, R.G.; de Bustros, S.; Enger, C.; Kato, H.; Lansing, M.; et al. Correlation of fibrosis and transforming growth factor-beta type 2 levels in the eye. J. Clin. Invest. 1989, 83, 1661–1666. [Google Scholar] [CrossRef] [PubMed]
- Nakatsukasa, H.; Nagy, P.; Evarts, R.P.; Hsia, C.C.; Marsden, E.; Thorgeirsson, S.S. Cellular distribution of transforming growth factor-beta 1 and procollagen types I, III, and IV transcripts in carbon tetrachloride-induced rat liver fibrosis. J. Clin. Invest. 1990, 85, 1833–1843. [Google Scholar] [CrossRef] [PubMed]
- Chang, Z.; Kishimoto, Y.; Hasan, A.; Welham, N.V. TGF-beta 3 modulates the inflammatory environment and reduces scar formation following vocal fold mucosal injury in rats. Dis. Model. Mech. 2014, 7, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Priyadarsini, S.; Hjortdal, J.; Sarker-Nag, A.; Sejersen, H.; Asara, J.M.; Karamichos, D. Gross cystic disease fluid protein-15/prolactin-inducible protein as a biomarker for keratoconus disease. PLoS One 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Karamichos, D.; Hutcheon, A.E.K.; Zieske, J.D. Transforming growth factor-β3 regulates assembly of a non-fibrotic matrix in a 3D corneal model. J. Tissue Eng. Regen. Med. 2011, 5, e228–e238. [Google Scholar] [CrossRef] [PubMed]
- Bystrom, B.; Carracedo, S.; Behndig, A.; Gullberg, D.; Pedrosa-Domellof, F. Alpha11 integrin in the human cornea: Importance in development and disease. Invest. Ophthalmol. Vis. Sci. 2009, 50, 5044–5053. [Google Scholar] [CrossRef] [PubMed]
- Parapuram, S.K.; Huh, K.; Liu, S.; Leask, A. Integrin beta1 is necessary for the maintenance of corneal structural integrity. Invest. Ophthalmol. Vis. Sci. 2011, 52, 7799–7806. [Google Scholar] [CrossRef] [PubMed]
- Weston, B.S.; Wahab, N.A.; Mason, R.M. CTGF mediates TGF-beta-induced fibronectin matrix deposition by upregulating active alpha5beta1 integrin in human mesangial cells. J. Am. Soc. Nephrol. 2003, 14, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Zambruno, G.; Marchisio, P.C.; Marconi, A.; Vaschieri, C.; Melchiori, A.; Giannetti, A.; De Luca, M. Transforming growth factor-beta 1 modulates beta 1 and beta 5 integrin receptors and induces the de novo expression of the alpha v beta 6 heterodimer in normal human keratinocytes: Implications for wound healing. J. Cell Biol. 1995, 129, 853–865. [Google Scholar] [CrossRef] [PubMed]
- Kagami, S.; Kuhara, T.; Yasutomo, K.; Okada, K.; Loster, K.; Reutter, W.; Kuroda, Y. Transforming growth factor-beta (TGF-beta) stimulates the expression of beta1 integrins and adhesion by rat mesangial cells. Exp. Cell Res. 1996, 229, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhou, G.H.; Birkenmeier, T.M.; Gong, J.; Sun, L.; Brattain, M.G. Autocrine transforming growth factor beta 1 modulates the expression of integrin alpha 5 beta 1 in human colon carcinoma FET cells. J. Biol. Chem. 1995, 270, 14154–14159. [Google Scholar] [CrossRef] [PubMed]
- Maier, P.; Broszinski, A.; Heizmann, U.; Bohringer, D.; Reinhardau, T. Active transforming growth factor-beta2 is increased in the aqueous humor of keratoconus patients. Mol. Vis. 2007, 13, 1198–1202. [Google Scholar] [PubMed]
- Saee-Rad, S.; Raoofian, R.; Mahbod, M.; Miraftab, M.; Mojarrad, M.; Asgari, S.; Rezvan, F.; Hashemi, H. Analysis of superoxide dismutase 1, dual-specificity phosphatase 1, and transforming growth factor, beta 1 genes expression in keratoconic and non-keratoconic corneas. Mol. Vis. 2013, 19, 2501–2507. [Google Scholar] [PubMed]
- Nakayasu, K.; Tanaka, M.; Konomi, H.; Hayashi, T. Distribution of types I, II, III, IV and V collagen in normal and keratoconus corneas. Ophthalmic Res. 1986, 18, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wenstrup, R.J.; Florer, J.B.; Brunskill, E.W.; Bell, S.M.; Chervoneva, I.; Birk, D.E. Type v collagen controls the initiation of collagen fibril assembly. J. Biol. Chem. 2004, 279, 53331–53337. [Google Scholar] [CrossRef] [PubMed]
- Von der Mark, K.; von der Mark, H.; Timpl, R.; Trelstad, R.L. Immunofluorescent localization of collagen types i, ii, and iii in the embryonic chick eye. Dev. Biol. 1977, 59, 75–85. [Google Scholar] [CrossRef]
- Malley, D.S.; Steinert, R.F.; Puliafito, C.A.; Dobi, E.T. Immunofluorescence study of corneal wound healing after excimer laser anterior keratectomy in the monkey eye. Arch. Ophthalmol. 1990, 108, 1316–1322. [Google Scholar] [CrossRef] [PubMed]
- Robert, L.; Legeais, J.M.; Robert, A.M.; Renard, G. Corneal collagens. Path. Biol. 2001, 49, 353–363. [Google Scholar] [CrossRef]
- Birk, D.E.; Fitch, J.M.; Babiarz, J.P.; Doane, K.J.; Linsenmayer, T.F. Collagen fibrillogenesis in vitro: Interaction of types I and V collagen regulates fibril diameter. J. Cell Sci. 1990, 95, 649–657. [Google Scholar] [PubMed]
- Page-McCaw, A.; Ewald, A.J.; Werb, Z. Matrix metalloproteinases and the regulation of tissue remodelling. Nat. Rev. Mol. Cell Biol. 2007, 8, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Collier, S.A. Is the corneal degradation in keratoconus caused by matrix-metalloproteinases? Clin. Exp. Ophthalmol. 2001, 29, 340–344. [Google Scholar] [CrossRef]
- Smith, V.A.; Hoh, H.B.; Littleton, M.; Easty, D.L. Over-expression of a gelatinase a activity in keratoconus. Eye 1995, 9, 429–433. [Google Scholar] [CrossRef] [PubMed]
- Seppala, H.P.; Maatta, M.; Rautia, M.; Mackiewicz, Z.; Tuisku, I.; Tervo, T.; Konttinen, Y.T. EMMPRIN and MMP-1 in keratoconus. Cornea 2006, 25, 325–330. [Google Scholar] [PubMed]
- Mackiewicz, Z.; Maatta, M.; Stenman, M.; Konttinen, L.; Tervo, T.; Konttinen, Y.T. Collagenolytic proteinases in keratoconus. Cornea 2006, 25, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Moeen Rezakhanlou, A.; Chavez-Munoz, C.; Lai, A.; Ghahary, A. Keratinocyte-releasable factors increased the expression of mmp1 and mmp3 in co-cultured fibroblasts under both 2D and 3D culture conditions. Mol. Cell. Biochem. 2009, 332, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gomes, L.R.; Terra, L.F.; Wailemann, R.A.; Labriola, L.; Sogayar, M.C. TGF-beta1 modulates the homeostasis between MMPs and MMP inhibitors through p38 MAPK and ERK1/2 in highly invasive breast cancer cells. BMC Cancer 2012, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Kang, L.; Wei, Q.; Cui, X.; Wang, S.; Chen, Y.; Jiang, Y. Expression and regulation of MMP1, MMP3, and MMP9 in the chicken ovary in response to gonadotropins, sex hormones, and TGFB1. Biol. Reprod. 2014, 90. [Google Scholar] [CrossRef] [PubMed]
- Karamichos, D.; Guo, X.Q.; Hutcheon, A.E.; Zieske, J.D. Human corneal fibrosis: An in vitro model. Invest. Ophthalmol. Vis. Sci. 2010, 51, 1382–1388. [Google Scholar] [CrossRef] [PubMed]
- Midwood, K.S.; Williams, L.V.; Schwarzbauer, J.E. Tissue repair and the dynamics of the extracellular matrix. Int. J. Biochem. Cell Biol. 2004, 36, 1031–1037. [Google Scholar] [CrossRef] [PubMed]
- Netto, M.V.; Mohan, R.R.; Ambrósio, R.J.; Hutcheon, A.E.K.; Zieske, J.D.; Wilson, S.E. Wound healing in the cornea: A review of refractive surgery complications and new prospects for therapy. Cornea 2005, 24, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.E.; Netto, M.; Ambrósio, R. Corneal cells: Chatty in development, homeostasis, wound healing, and disease. Am. J. Ophthalmol. 2003, 136, 530–536. [Google Scholar] [CrossRef]
- Wilson, S.E.; Kim, W.-J. Keratocyte apoptosis: Implications on corneal wound healing, tissue organization, and disease. Investig. Ophthalmol. Vis. Sci. 1998, 39, 220–226. [Google Scholar]
- Gupta, A.; Monroy, D.; Ji, Z.; Yoshino, K.; Huang, A.; Pflugfelder, S.C. Transforming growth factor beta-1 and beta-2 in human tear fluid. Curr. Eye Res. 1996, 15, 605–614. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyon, D.; McKay, T.B.; Sarkar-Nag, A.; Priyadarsini, S.; Karamichos, D. Human Keratoconus Cell Contractility is Mediated by Transforming Growth Factor-Beta Isoforms. J. Funct. Biomater. 2015, 6, 422-438. https://doi.org/10.3390/jfb6020422
Lyon D, McKay TB, Sarkar-Nag A, Priyadarsini S, Karamichos D. Human Keratoconus Cell Contractility is Mediated by Transforming Growth Factor-Beta Isoforms. Journal of Functional Biomaterials. 2015; 6(2):422-438. https://doi.org/10.3390/jfb6020422
Chicago/Turabian StyleLyon, Desiree', Tina B. McKay, Akhee Sarkar-Nag, Shrestha Priyadarsini, and Dimitrios Karamichos. 2015. "Human Keratoconus Cell Contractility is Mediated by Transforming Growth Factor-Beta Isoforms" Journal of Functional Biomaterials 6, no. 2: 422-438. https://doi.org/10.3390/jfb6020422
APA StyleLyon, D., McKay, T. B., Sarkar-Nag, A., Priyadarsini, S., & Karamichos, D. (2015). Human Keratoconus Cell Contractility is Mediated by Transforming Growth Factor-Beta Isoforms. Journal of Functional Biomaterials, 6(2), 422-438. https://doi.org/10.3390/jfb6020422