
“Attractive” Treatment for Abdominal Aortic Aneurysm Repair: Magnetic Localization of Silk-Iron Packaged Extracellular Vesicles

 , , , , , , and
, , , , , , and 
Abstract
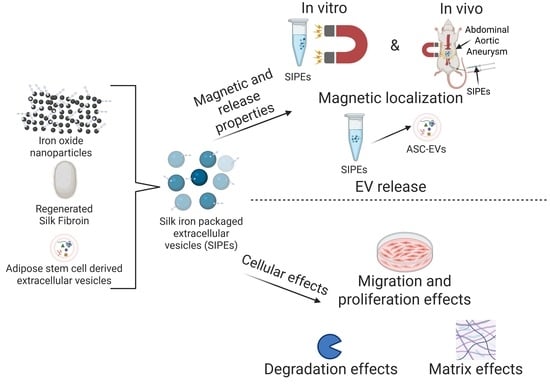
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Extracellular Vesicle Isolation or Preparation
2.3. Generation of Silk, Fabrication of IONPs, and Incorporation of Iron Oxide Nanoparticles (IONPs) into Regenerated Silk Fibroin (RSF) Solution
2.4. Fabrication of SIPEs and SIMPs
2.5. Extracellular Vesicle Characterization
2.6. Magnetic Moveability Analysis with a Ninhydrin Assay for Protein Detection
2.7. Flow Cytometry Releasate Experiments
2.8. EV Uptake
2.9. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR) of Smooth Muscle Cells in Fibrin Gels
2.10. Fibrin Gel System for Vascular Smooth Muscle Cells for Long-Term Matrix Deposition
2.11. Ninhydrin (Insoluble Elastin) and Hydroxyproline (Collagen) Assays
2.12. Development of Fibrin Gel Co-Culture System for Vascular Smooth Muscle Cells and U937 Cells
2.13. Zymogram to Monitor Matrix Metalloproteinase (MMP) Activity
2.14. SMC Migration Assay
2.15. SMC Proliferation Assay
2.16. Rat Model of AAA
2.17. Preparation and Injection of SIMPs
2.18. Aorta Explant Staining
2.19. Statistical Analysis
3. Results
3.1. EV and ASC Characterization
3.2. SEM of SIPEs
3.3. Magnetic Separation Assay of SIPEs, SIMPs, Silk EV MPs, and Silk MPs
3.4. Flow Cytometry of SIPE Releasates
3.5. SIPE Released EV Uptake at 6 H vs. 24 H
3.6. Real-Time Quantitative PCR of SMC Seeded Fibrin Gels
3.7. SIPE and ASC-EV Effects on Matrix Deposition
3.8. SIPE and EV Effect on Degradative Activity
3.9. Migration and Proliferation of SMCs
3.10. Magnetic Localization of SIMPs in an In Vivo AAA Rat Model
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AAA | Abdominal aortic aneurysm |
| AB | Alamar blue |
| ASC-EVs | ASC-derived extracellular vesicles |
| ASCs | Adipose-derived mesenchymal stem cells |
| ATCC | American Type Culture Collection |
| CM | Conditioned media |
| DPBS | Dulbecco’s phosphate-buffered saline |
| ECM | Extracellular matrix |
| FBS | Fetal bovine serum |
| IONPsLD | Iron oxide nanoparticles |
| MMP | Matrix metalloproteinase |
| MPs | Microparticles |
| nCM | Non-conditioned media |
| NTA | Nanoparticle tracking analysis |
| Pen/strep | Penicillin/streptomycin |
| RPMI | Roswell Park Memorial Institute |
| RSF | Regenerated silk fibroin |
| SBM | Supplemented basal media |
| SIMPs | Silk-iron microparticles |
| SIPEs | Silk-iron packaged extracellular vesicles |
| SMCs | Smooth muscle cells |
| TFF | Tangential flow filtration |
| UC | Ultracentrifugation |
References
- Murphy, S.L.; Xu, J.; Kochanek, K.D.; Arias, E.; Tejada-Vera, B. Deaths: Final Data for 2018. Natl. Vital. Stat. Rep. 2021, 69, 1–83. [Google Scholar]
- Patel, M.I.; Hardman, D.T.; Fisher, C.M.; Appleberg, M. Current views on the pathogenesis of abdominal aortic aneurysms. J. Am. Coll. Surg. 1995, 181, 371–382. [Google Scholar]
- Sen, I.; Franco-Mesa, C.; Erben, Y.; DeMartino, R.R. Abdominal Aortic and Visceral Artery Aneurysms. Cardiol. Clin. 2021, 39, 517–525. [Google Scholar] [CrossRef]
- Blose, K.J.; Ennis, T.L.; Arif, B.; Weinbaum, J.S.; Curci, J.A.; Vorp, D.A. Periadventitial adipose-derived stem cell treatment halts elastase-induced abdominal aortic aneurysm progression. Regen. Med. 2014, 9, 733–741. [Google Scholar] [CrossRef]
- Sharma, A.K.; Lu, G.; Jester, A.; Johnston, W.F.; Zhao, Y.; Hajzus, V.A.; Saadatzadeh, M.R.; Su, G.; Bhamidipati, C.M.; Mehta, G.S.; et al. Experimental abdominal aortic aneurysm formation is mediated by IL-17 and attenuated by mesenchymal stem cell treatment. Circulation 2012, 126, S38–S45. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wen, H.; Lv, J.; Luan, B.; Meng, J.; Gong, S.; Wen, J.; Xin, S. Therapeutic efficacy of mesenchymal stem cells for abdominal aortic aneurysm: A meta-analysis of preclinical studies. Stem Cell Res. Ther. 2022, 13, 81. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.K.; Green, L.A.; Gutwein, A.R.; Drucker, N.A.; Motaganahalli, R.L.; Fajardo, A.; Babbey, C.M.; Murphy, M.P. Rationale and Design of the ARREST Trial Investigating Mesenchymal Stem Cells in the Treatment of Small Abdominal Aortic Aneurysm. Ann. Vasc. Surg. 2018, 47, 230–237. [Google Scholar] [CrossRef]
- Krawiec, J.T.; Weinbaum, J.S.; Liao, H.T.; Ramaswamy, A.K.; Pezzone, D.J.; Josowitz, A.D.; D’Amore, A.; Rubin, J.P.; Wagner, W.R.; Vorp, D.A. In Vivo Functional Evaluation of Tissue-Engineered Vascular Grafts Fabricated Using Human Adipose-Derived Stem Cells from High Cardiovascular Risk Populations. Tissue Eng. Part A 2016, 22, 765–775. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhang, H.; Liang, X.; Hong, Y.; Mao, M.; Han, Q.; He, H.; Tao, W.; Jiang, G.; Zhang, Y.; et al. Adipose-Derived Mesenchymal Stem Cells Isolated from Patients with Abdominal Aortic Aneurysm Exhibit Senescence Phenomena. Oxidative Med. Cell. Longev. 2019, 2019, 1305049. [Google Scholar] [CrossRef]
- Krawiec, J.T.; Liao, H.T.; Kwan, L.L.; D’Amore, A.; Weinbaum, J.S.; Rubin, J.P.; Wagner, W.R.; Vorp, D.A. Evaluation of the stromal vascular fraction of adipose tissue as the basis for a stem cell-based tissue-engineered vascular graft. J. Vasc. Surg. 2017, 66, 883–890.e1. [Google Scholar] [CrossRef]
- Horwitz, E.M.; Gordon, P.L.; Koo, W.K.; Marx, J.C.; Neel, M.D.; McNall, R.Y.; Muul, L.; Hofmann, T. Isolated allogeneic bone marrow-derived mesenchymal cells engraft and stimulate growth in children with osteogenesis imperfecta: Implications for cell therapy of bone. Proc. Natl. Acad. Sci. USA 2002, 99, 8932–8937. [Google Scholar] [CrossRef]
- James, A.W. Review of Signaling Pathways Governing MSC Osteogenic and Adipogenic Differentiation. Scientifica 2013, 2013, 1–17. [Google Scholar] [CrossRef]
- Doorn, J.; Moll, G.; Le Blanc, K.; van Blitterswijk, C.; de Boer, J. Therapeutic applications of mesenchymal stromal cells: Paracrine effects and potential improvements. Tissue Eng. Part B Rev. 2012, 18, 101–115. [Google Scholar] [CrossRef]
- Guillén, M.I.; Platas, J.; Pérez Del Caz, M.D.; Mirabet, V.; Alcaraz, M.J. Paracrine Anti-inflammatory Effects of Adipose Tissue-Derived Mesenchymal Stem Cells in Human Monocytes. Front. Physiol. 2018, 9, 661. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.R.; Teixeira, G.Q.; Santos, S.G.; Barbosa, M.A.; Almeida-Porada, G.; Goncalves, R.M. Mesenchymal Stromal Cell Secretome: Influencing Therapeutic Potential by Cellular Pre-conditioning. Front. Immunol. 2018, 9, 2837. [Google Scholar] [CrossRef]
- Foo, J.B.; Looi, Q.H.; Chong, P.P.; Hassan, N.H.; Yeo, G.E.C.; Ng, C.Y.; Koh, B.; How, C.W.; Lee, S.H.; Law, J.X. Comparing the Therapeutic Potential of Stem Cells and their Secretory Products in Regenerative Medicine. Stem Cells Int. 2021, 2021, 2616807. [Google Scholar] [CrossRef] [PubMed]
- Gallina, C.; Turinetto, V.; Giachino, C. A New Paradigm in Cardiac Regeneration: The Mesenchymal Stem Cell Secretome. Stem Cells Int. 2015, 2015, 765846. [Google Scholar] [CrossRef]
- Spinosa, M.; Lu, G.; Su, G.; Bontha, S.V.; Gehrau, R.; Salmon, M.D.; Smith, J.R.; Weiss, M.L.; Mas, V.R.; Upchurch, G.R., Jr.; et al. Human mesenchymal stromal cell-derived extracellular vesicles attenuate aortic aneurysm formation and macrophage activation via microRNA-147. FASEB J. 2018, 32, fj201701138RR. [Google Scholar] [CrossRef]
- Cunnane, E.M.; Lorentz, K.L.; Ramaswamy, A.K.; Gupta, P.; Mandal, B.B.; O’Brien, F.J.; Weinbaum, J.S.; Vorp, D.A. Extracellular Vesicles Enhance the Remodeling of Cell-Free Silk Vascular Scaffolds in Rat Aortae. ACS Appl. Mater. Interfaces 2020, 12, 26955–26965. [Google Scholar] [CrossRef]
- Cunnane, E.M.; Ramaswamy, A.K.; Lorentz, K.L.; Vorp, D.A.; Weinbaum, J.S. Extracellular Vesicles Derived from Primary Adipose Stromal Cells Induce Elastin and Collagen Deposition by Smooth Muscle Cells within 3D Fibrin Gel Culture. Bioengineering 2021, 8, 51. [Google Scholar] [CrossRef] [PubMed]
- Alcaraz, M.J.; Compan, A.; Guillen, M.I. Extracellular Vesicles from Mesenchymal Stem Cells as Novel Treatments for Musculoskeletal Diseases. Cells 2019, 9, 98. [Google Scholar] [CrossRef]
- Almeria, C.; Weiss, R.; Roy, M.; Tripisciano, C.; Kasper, C.; Weber, V.; Egger, D. Hypoxia Conditioned Mesenchymal Stem Cell-Derived Extracellular Vesicles Induce Increased Vascular Tube Formation in vitro. Front. Bioeng. Biotechnol. 2019, 7, 292. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Y.; Wang, Z.; Zhang, L.; Xu, Y.; Li, Y.; Zhang, L.; Wang, G.; Yang, S.; Xue, G. Mesenchymal stem cell-derived extracellular vesicles protect against abdominal aortic aneurysm formation by inhibiting NET-induced ferroptosis. Exp. Mol. Med. 2023, 55, 939–951. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.S.; Cho, W.L.; Choi, Y.J.; Kim, J.D.; Park, H.A.; Kim, S.Y.; Park, J.H.; Jo, D.G.; Cho, Y.W. Functional recovery in photo-damaged human dermal fibroblasts by human adipose-derived stem cell extracellular vesicles. J. Extracell. Vesicles 2019, 8, 1565885. [Google Scholar] [CrossRef] [PubMed]
- Eleuteri, S.; Fierabracci, A. Insights into the Secretome of Mesenchymal Stem Cells and Its Potential Applications. Int. J. Mol. Sci. 2019, 20, 4597. [Google Scholar] [CrossRef] [PubMed]
- Figliolini, F.; Ranghino, A.; Grange, C.; Cedrino, M.; Tapparo, M.; Cavallari, C.; Rossi, A.; Togliatto, G.; Femminò, S.; Gugliuzza, M.V.; et al. Extracellular Vesicles From Adipose Stem Cells Prevent Muscle Damage and Inflammation in a Mouse Model of Hind Limb Ischemia. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 239–254. [Google Scholar] [CrossRef]
- Sajeesh, S.; Camardo, A.; Dahal, S.; Ramamurthi, A. Surface-Functionalized Stem Cell-Derived Extracellular Vesicles for Vascular Elastic Matrix Regenerative Repair. Mol. Pharm. 2023, 20, 2801–2813. [Google Scholar]
- Sajeesh, S.; Broekelman, T.; Mecham, R.P.; Ramamurthi, A. Stem cell derived extracellular vesicles for vascular elastic matrix regenerative repair. Acta Biomater. 2020, 113, 267–278. [Google Scholar] [CrossRef]
- Adamiak, M.; Sahoo, S. Exosomes in Myocardial Repair: Advances and Challenges in the Development of Next-Generation Therapeutics. Mol. Ther. 2018, 26, 1635–1643. [Google Scholar] [CrossRef]
- Bari, E.; Ferrarotti, I.; Di Silvestre, D.; Grisoli, P.; Barzon, V.; Balderacchi, A.; Torre, M.L.; Rossi, R.; Mauri, P.; Corsico, A.G.; et al. Adipose Mesenchymal Extracellular Vesicles as Alpha-1-Antitrypsin Physiological Delivery Systems for Lung Regeneration. Cells 2019, 8, 965. [Google Scholar] [CrossRef]
- Heo, J.S.; Choi, Y.; Kim, H.O. Adipose-Derived Mesenchymal Stem Cells Promote M2 Macrophage Phenotype through Exosomes. Stem Cells Int. 2019, 2019, 7921760. [Google Scholar] [CrossRef] [PubMed]
- Chabria, Y.; Duffy, G.; Lowery, A.; Dwyer, R. Hydrogels: 3D Drug Delivery Systems for Nanoparticles and Extracellular Vesicles. Biomedicines 2021, 9, 1694. [Google Scholar] [CrossRef] [PubMed]
- Wiklander, O.P.B.; Nordin, J.Z.; O’Loughlin, A.; Gustafsson, Y.; Corso, G.; Mäger, I.; Vader, P.; Lee, Y.; Sork, H.; Seow, Y.; et al. Extracellular vesicle in vivo biodistribution is determined by cell source, route of administration and targeting. J. Extracell. Vesicles 2015, 4, 26316. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Jordan, V.; Blenkiron, C.; Chamley, L.W. Biodistribution of extracellular vesicles following administration into animals: A systematic review. J. Extracell. Vesicles 2021, 10, e12085. [Google Scholar] [CrossRef]
- Kulaj, K.; Harger, A.; Bauer, M.; Caliskan, Ö.S.; Gupta, T.K.; Chiang, D.M.; Milbank, E.; Reber, J.; Karlas, A.; Kotzbeck, P.; et al. Adipocyte-derived extracellular vesicles increase insulin secretion through transport of insulinotropic protein cargo. Nat. Commun. 2023, 14, 709. [Google Scholar] [CrossRef]
- Marini, A.X.; Tomaraei, G.N.; Weinbaum, J.S.; Bedewy, M.; Vorp, D.A. Chemical Conjugation of Iron Oxide Nanoparticles for the Development of Magnetically Directable Silk Particles. ACS Appl. Mater. Interfaces 2025, 17, 8901–8913. [Google Scholar] [CrossRef]
- Kucharczyk, K.; Rybka, J.D.; Hilgendorff, M.; Krupinski, M.; Slachcinski, M.; Mackiewicz, A.; Giersig, M.; Dams-Kozlowska, H. Composite spheres made of bioengineered spider silk and iron oxide nanoparticles for theranostics applications. PLoS ONE 2019, 14, e0219790. [Google Scholar] [CrossRef]
- Kucharczyk, K.; Weiss, M.; Jastrzebska, K.; Luczak, M.; Ptak, A.; Kozak, M.; Mackiewicz, A.; Dams-Kozlowska, H. Bioengineering the spider silk sequence to modify its affinity for drugs. Int. J. Nanomed. 2018, 13, 4247–4261. [Google Scholar] [CrossRef]
- Mwangi, T.K.; Bowles, R.D.; Tainter, D.M.; Bell, R.D.; Kaplan, D.L.; Setton, L.A. Synthesis and characterization of silk fibroin microparticles for intra-articular drug delivery. Int. J. Pharm. 2015, 485, 7–14. [Google Scholar] [CrossRef]
- Wu, J.; Wang, J.; Zhang, J.; Zheng, Z.; Kaplan, D.L.; Li, G.; Wang, X. Oral Delivery of Curcumin Using Silk Nano- and Microparticles. ACS Biomater. Sci. Eng. 2018, 4, 3885–3894. [Google Scholar] [CrossRef]
- Wang, X.; Wenk, E.; Matsumoto, A.; Meinel, L.; Li, C.; Kaplan, D.L. Silk microspheres for encapsulation and controlled release. J. Control Release 2007, 117, 360–370. [Google Scholar] [CrossRef]
- Bessa, P.C.; Balmayor, E.R.; Azevedo, H.S.; Nurnberger, S.; Casal, M.; van Griensven, M.; Reis, R.L.; Redl, H. Silk fibroin microparticles as carriers for delivery of human recombinant BMPs. Physical characterization and drug release. J. Tissue Eng. Regen. Med. 2010, 4, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Elia, R.; Guo, J.; Budijono, S.; Normand, V.; Benczedi, D.; Omenetto, F.; Kaplan, D.L. Encapsulation of Volatile Compounds in Silk Microparticles. J. Coat. Technol. Res. 2015, 12, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chen, X.; Ding, F.; Zhang, P.; Liu, J.; Gu, X. Biocompatibility evaluation of silk fibroin with peripheral nerve tissues and cells in vitro. Biomaterials 2007, 28, 1643–1652. [Google Scholar] [CrossRef] [PubMed]
- Karahaliloğlu, Z.; Yalçın, E.; Demirbilek, M.; Denkbaş, E.B. Magnetic silk fibroin e-gel scaffolds for bone tissue engineering applications. J. Bioact. Compat. Polym. 2017, 32, 596–614. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, B. Biodegradation of silk biomaterials. Int. J. Mol. Sci. 2009, 10, 1514–1524. [Google Scholar] [CrossRef]
- Shi, P.; Goh, J.C. Release and cellular acceptance of multiple drugs loaded silk fibroin particles. Int. J. Pharm. 2011, 420, 282–289. [Google Scholar] [CrossRef]
- Rockwood, D.N.; Preda, R.C.; Yucel, T.; Wang, X.; Lovett, M.L.; Kaplan, D.L. Materials fabrication from Bombyx mori silk fibroin. Nat. Protoc. 2011, 6, 1612–1631. [Google Scholar] [CrossRef]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Welsh, J.A.; Goberdhan, D.C.I.; O’Driscoll, L.; Buzas, E.I.; Blenkiron, C.; Bussolati, B.; Cai, H.; Di Vizio, D.; Driedonks, T.A.P.; Erdbrugger, U.; et al. Minimal information for studies of extracellular vesicles (MISEV2023): From basic to advanced approaches. J. Extracell. Vesicles 2024, 13, e12404. [Google Scholar] [CrossRef]
- Ramaswamy, A.K.; Sides, R.E.; Cunnane, E.M.; Lorentz, K.L.; Reines, L.M.; Vorp, D.A.; Weinbaum, J.S. Adipose-derived stromal cell secreted factors induce the elastogenesis cascade within 3D aortic smooth muscle cell constructs. Matrix Biol. Plus 2019, 4, 100014. [Google Scholar] [CrossRef]
- Xing, Y.; Yerneni, S.S.; Wang, W.; Taylor, R.E.; Campbell, P.G.; Ren, X. Engineering pro-angiogenic biomaterials via chemoselective extracellular vesicle immobilization. Biomaterials 2022, 281, 121357. [Google Scholar] [CrossRef]
- Ahmann, K.A.; Weinbaum, J.S.; Johnson, S.L.; Tranquillo, R.T. Fibrin degradation enhances vascular smooth muscle cell proliferation and matrix deposition in fibrin-based tissue constructs fabricated in vitro. Tissue Eng. Part A 2010, 16, 3261–3270. [Google Scholar] [CrossRef] [PubMed]
- Gueldner, P.H.; Marini, A.X.; Li, B.; Darvish, C.J.; Chung, T.K.; Weinbaum, J.S.; Curci, J.A.; Vorp, D.A. Mechanical and matrix effects of short and long-duration exposure to beta-aminopropionitrile in elastase-induced model abdominal aortic aneurysm in mice. JVS-Vasc. Sci. 2023, 4, 100098. [Google Scholar] [CrossRef]
- Airhart, N.; Brownstein, B.H.; Cobb, J.P.; Schierding, W.; Arif, B.; Ennis, T.L.; Thompson, R.W.; Curci, J.A. Smooth muscle cells from abdominal aortic aneurysms are unique and can independently and synergistically degrade insoluble elastin. J. Vasc. Surg. 2014, 60, 1033–1041. [Google Scholar] [CrossRef]
- Sastriques-Dunlop, S.; Elizondo-Benedetto, S.; Arif, B.; Meade, R.; Zaghloul, M.S.; Luehmann, H.; Heo, G.S.; English, S.J.; Liu, Y.; Zayed, M.A. Ketosis prevents abdominal aortic aneurysm rupture through C-C chemokine receptor type 2 downregulation and enhanced extracellular matrix balance. Sci. Rep. 2024, 14, 1438. [Google Scholar] [CrossRef]
- English, S.J.; Diaz, J.A.; Shao, X.; Gordon, D.; Bevard, M.; Su, G.; Henke, P.K.; Rogers, V.E.; Upchurch, G.R.; Piert, M. Utility of 18 F-FDG and 11C-PBR28 microPET for the assessment of rat aortic aneurysm inflammation. EJNMMI Res. 2014, 4, 20. [Google Scholar] [CrossRef]
- Bari, E.; Di Gravina, G.M.; Scocozza, F.; Perteghella, S.; Frongia, B.; Tengattini, S.; Segale, L.; Torre, M.L.; Conti, M. Silk Fibroin Bioink for 3D Printing in Tissue Regeneration: Controlled Release of MSC extracellular Vesicles. Pharmaceutics 2023, 15, 383. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.; Wang, X.; Zhao, M.; Cai, T.; Liu, P.; Li, J.; Willard, B.; Zu, L.; Zhou, E.; Li, Y.; et al. Macrophage Foam Cell-Derived Extracellular Vesicles Promote Vascular Smooth Muscle Cell Migration and Adhesion. J. Am. Heart Assoc. 2016, 5, e004099. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.P.; Salmon, M.; Pope, N.H.; Lu, G.; Su, G.; Sharma, A.K.; Ailawadi, G.; Upchurch, G.R. Attenuation of aortic aneurysms with stem cells from different genders. J. Surg. Res. 2015, 199, 249–258. [Google Scholar] [CrossRef]
- Visan, K.S.; Lobb, R.J.; Ham, S.; Lima, L.G.; Palma, C.; Edna, C.P.Z.; Wu, L.Y.; Gowda, H.; Datta, K.K.; Hartel, G.; et al. Comparative analysis of tangential flow filtration and ultracentrifugation, both combined with subsequent size exclusion chromatography, for the isolation of small extracellular vesicles. J. Extracell. Vesicles 2022, 11, 12266. [Google Scholar] [CrossRef] [PubMed]
- Bordanaba-Florit, G.; Royo, F.; Kruglik, S.G.; Falcón-Pérez, J.M. Using single-vesicle technologies to unravel the heterogeneity of extracellular vesicles. Nat. Protoc. 2021, 16, 3163–3185. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.S.; Kim, D.K.; Kim, Y.K.; Gho, Y.S. Proteomics, transcriptomics and lipidomics of exosomes and ectosomes. Proteomics 2013, 13, 1554–1571. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.W.; Seo, M.K.; Woo, E.Y.; Kim, S.H.; Park, E.J.; Kim, S. Exosomes from human adipose-derived stem cells promote proliferation and migration of skin fibroblasts. Exp. Dermatol. 2018, 27, 1170–1172. [Google Scholar] [CrossRef]
- Fontaine, M.; Herkenne, S.; Ek, O.; Paquot, A.; Boeckx, A.; Paques, C.; Nivelles, O.; Thiry, M.; Struman, I. Extracellular Vesicles Mediate Communication between Endothelial and Vascular Smooth Muscle Cells. Int. J. Mol. Sci. 2021, 23, 331. [Google Scholar] [CrossRef]
- Costa Verdera, H.; Gitz-Francois, J.J.; Schiffelers, R.M.; Vader, P. Cellular uptake of extracellular vesicles is mediated by clathrin-independent endocytosis and macropinocytosis. J. Control Release 2017, 266, 100–108. [Google Scholar] [CrossRef]
- Kwok, Z.H.; Wang, C.; Jin, Y. Extracellular Vesicle Transportation and Uptake by Recipient Cells: A Critical Process to Regulate Human Diseases. Processes 2021, 9, 273. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| ELN (Tropoelastin) | AGTTGGCATTTCCCCCGAAG | TAACCCAAACTGGGCGGCTT |
| COL1A1 | AGTGTGGCCCAGAAGAACTG | CCGCCATACTCGAACTGGAA |
| FBN1 | CGTCAACACTGATGGCTCCT | CTCCGCATGTGTGTGTCAAC |
| LOX | CGACCCTTACAACCCCTACA | CAGGTCTGGGAGACCGTACT |
| EFEMP2 | GCCCGAGTGTGTGGACATAG | ACACAGGAGCGGTTGTTAGG |
| FBLN5 | TTCTTCTCGCCTTCGCATCT | ATTCGTGCACTGTGCCTGT |
| LOXL1 | CAGACTTCCTCCCCAACCG | ATGCTGTGGTAATGCTGGT |
| RPS9 | CTGAAGCTGATCGGCGAGTA | GGGTCCTTCTCATCAAGCGT |
| DPBS | ASC-EVs | SIMPs | SIPEs | |
|---|---|---|---|---|
| Migration | 1.00 ± 0.85 | 77.31 ± 19.30 | 0.69 ± 0.35 | 1.12 ± 0.44 |
| Proliferation 48 h | 1.00 ± 0.03 | 1.22 ± 0.01 | 1.08 ± 0.07 | 1.20 ± 0.09 |
| Proliferation 96 h | 1.00 ± 0.19 | 1.29 ± 0.11 | 0.97 ± 0.03 | 1.12 ± 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marini, A.X.; McLoughlin, K.J.; Pellegrino, A.R.; Tomaraei, G.N.; Li, B.; Curci, J.A.; Bedewy, M.; Weinbaum, J.S.; Vorp, D.A. “Attractive” Treatment for Abdominal Aortic Aneurysm Repair: Magnetic Localization of Silk-Iron Packaged Extracellular Vesicles. J. Funct. Biomater. 2025, 16, 395. https://doi.org/10.3390/jfb16110395
Marini AX, McLoughlin KJ, Pellegrino AR, Tomaraei GN, Li B, Curci JA, Bedewy M, Weinbaum JS, Vorp DA. “Attractive” Treatment for Abdominal Aortic Aneurysm Repair: Magnetic Localization of Silk-Iron Packaged Extracellular Vesicles. Journal of Functional Biomaterials. 2025; 16(11):395. https://doi.org/10.3390/jfb16110395
Chicago/Turabian StyleMarini, Ande X., Kiran J. McLoughlin, Amanda R. Pellegrino, Golnaz N. Tomaraei, Bo Li, John A. Curci, Mostafa Bedewy, Justin S. Weinbaum, and David A. Vorp. 2025. "“Attractive” Treatment for Abdominal Aortic Aneurysm Repair: Magnetic Localization of Silk-Iron Packaged Extracellular Vesicles" Journal of Functional Biomaterials 16, no. 11: 395. https://doi.org/10.3390/jfb16110395
APA StyleMarini, A. X., McLoughlin, K. J., Pellegrino, A. R., Tomaraei, G. N., Li, B., Curci, J. A., Bedewy, M., Weinbaum, J. S., & Vorp, D. A. (2025). “Attractive” Treatment for Abdominal Aortic Aneurysm Repair: Magnetic Localization of Silk-Iron Packaged Extracellular Vesicles. Journal of Functional Biomaterials, 16(11), 395. https://doi.org/10.3390/jfb16110395