
A Potential Effect of Circadian Rhythm in the Delivery/Therapeutic Performance of Paclitaxel–Dendrimer Nanosystems

 , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
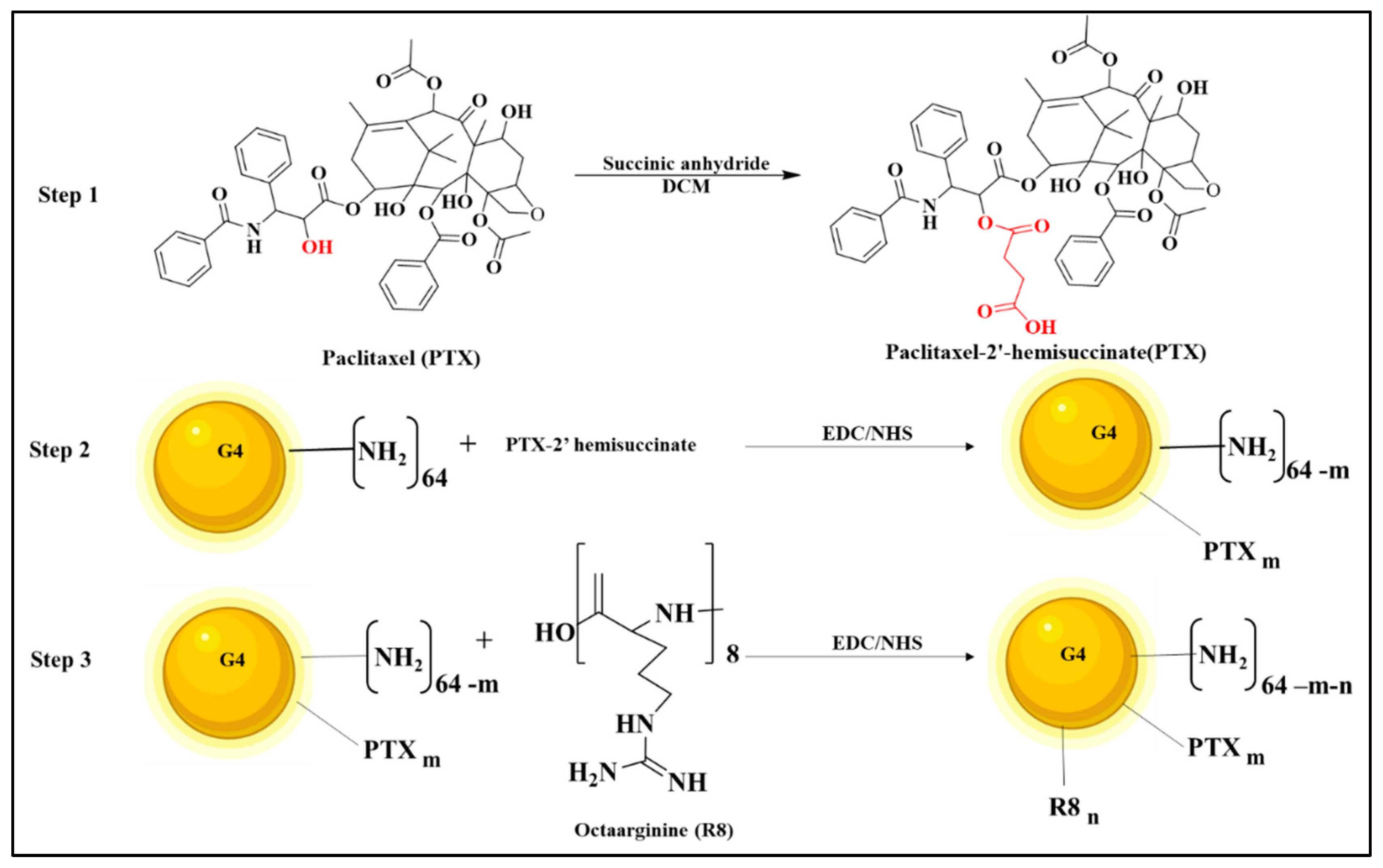
2.2. Preparation of PTX-2-Hemisuccinate (PTX-SA)
2.3. Preparation of PAMAM G4-PTX
2.4. Preparation of PAMAM G4-PTX-R8
2.5. Nuclear Magnetic Resonance (NMR) Spectroscopy of PAMAM G4-PTX-R8 Complexes
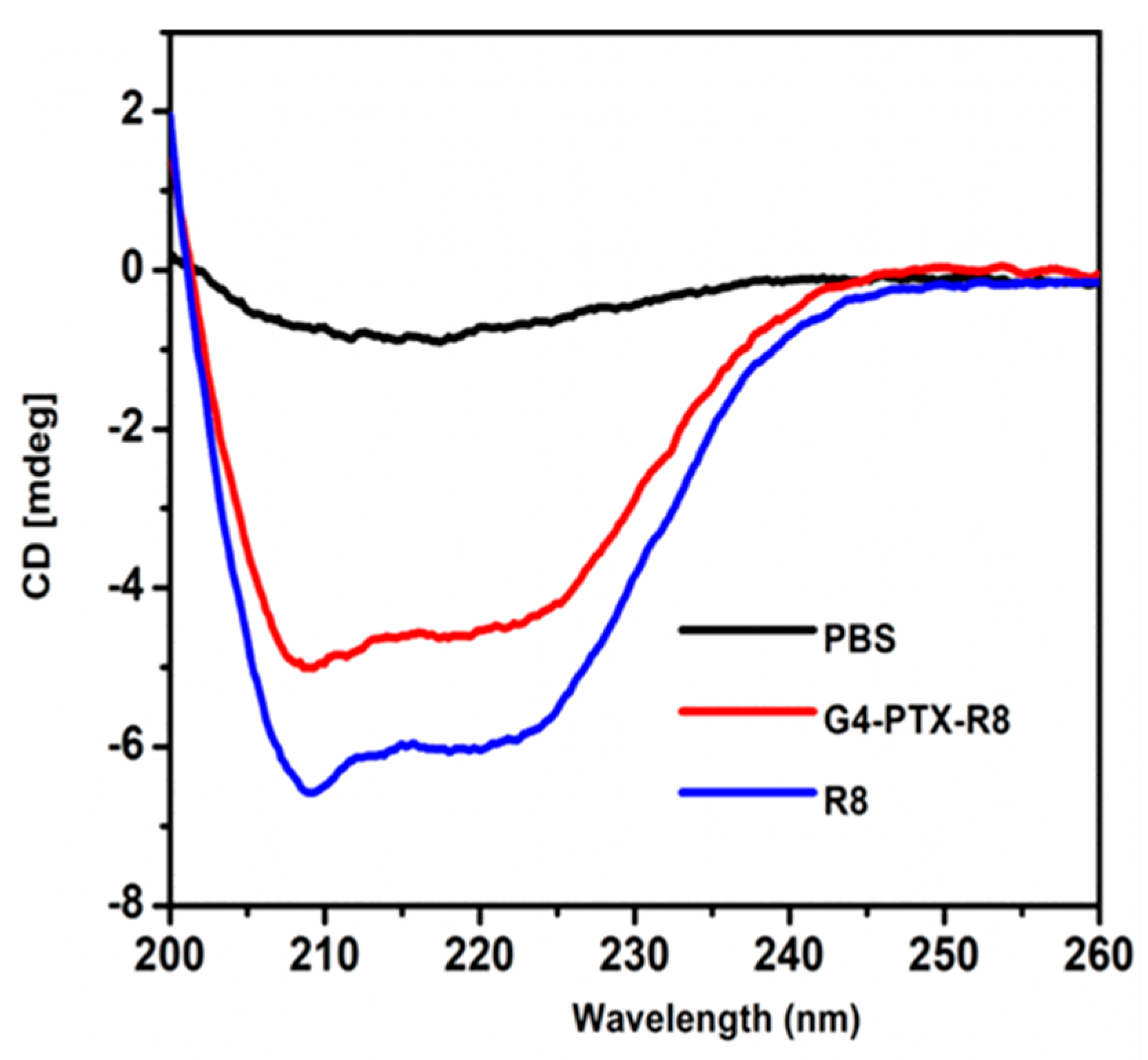
2.6. Circular Dichroism (CD) Spectroscopy
2.7. Zeta Potential and Particle Size
2.8. Morphological Analysis Using SEM
2.9. Cell Culture
2.10. Cytotoxicity Study
2.11. In Vitro Studies to Acess Circadian Rhythms Impact on Cellular Uptake of the Nanosystem
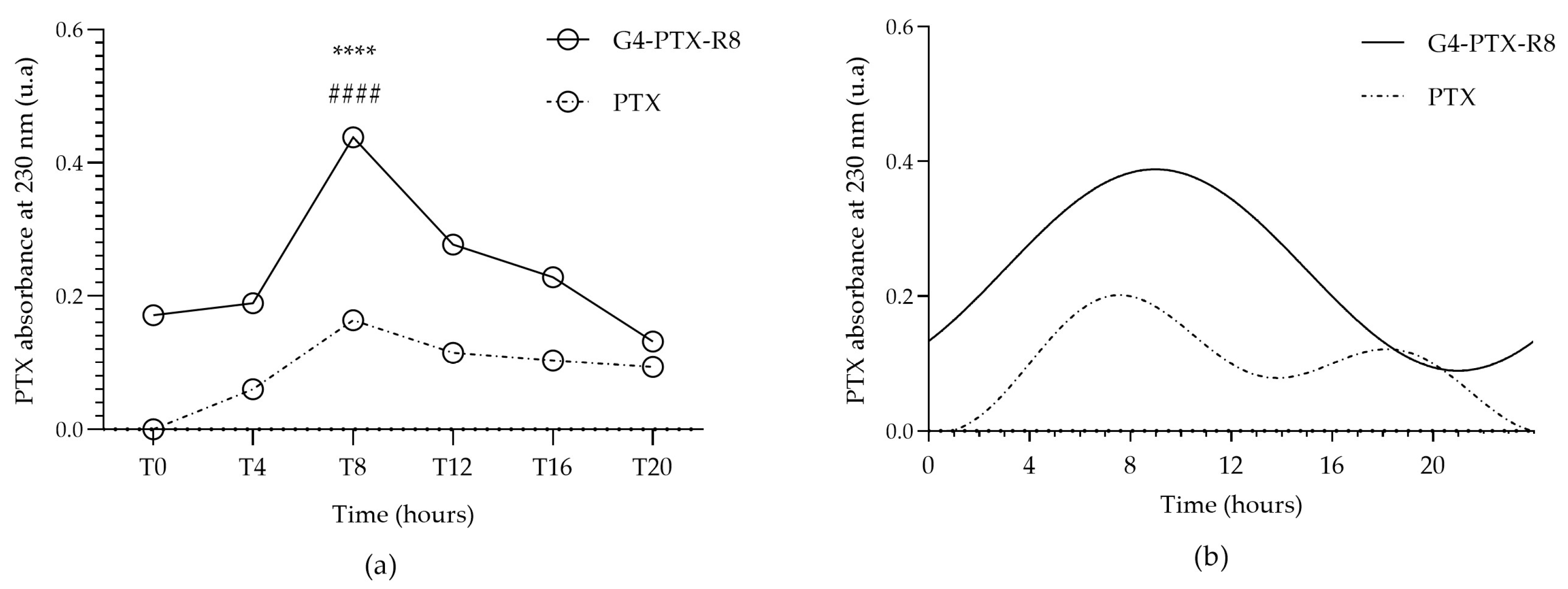
2.12. Cell-Associated PTX
2.13. Bmal1 Silencing on Hela Cell Line
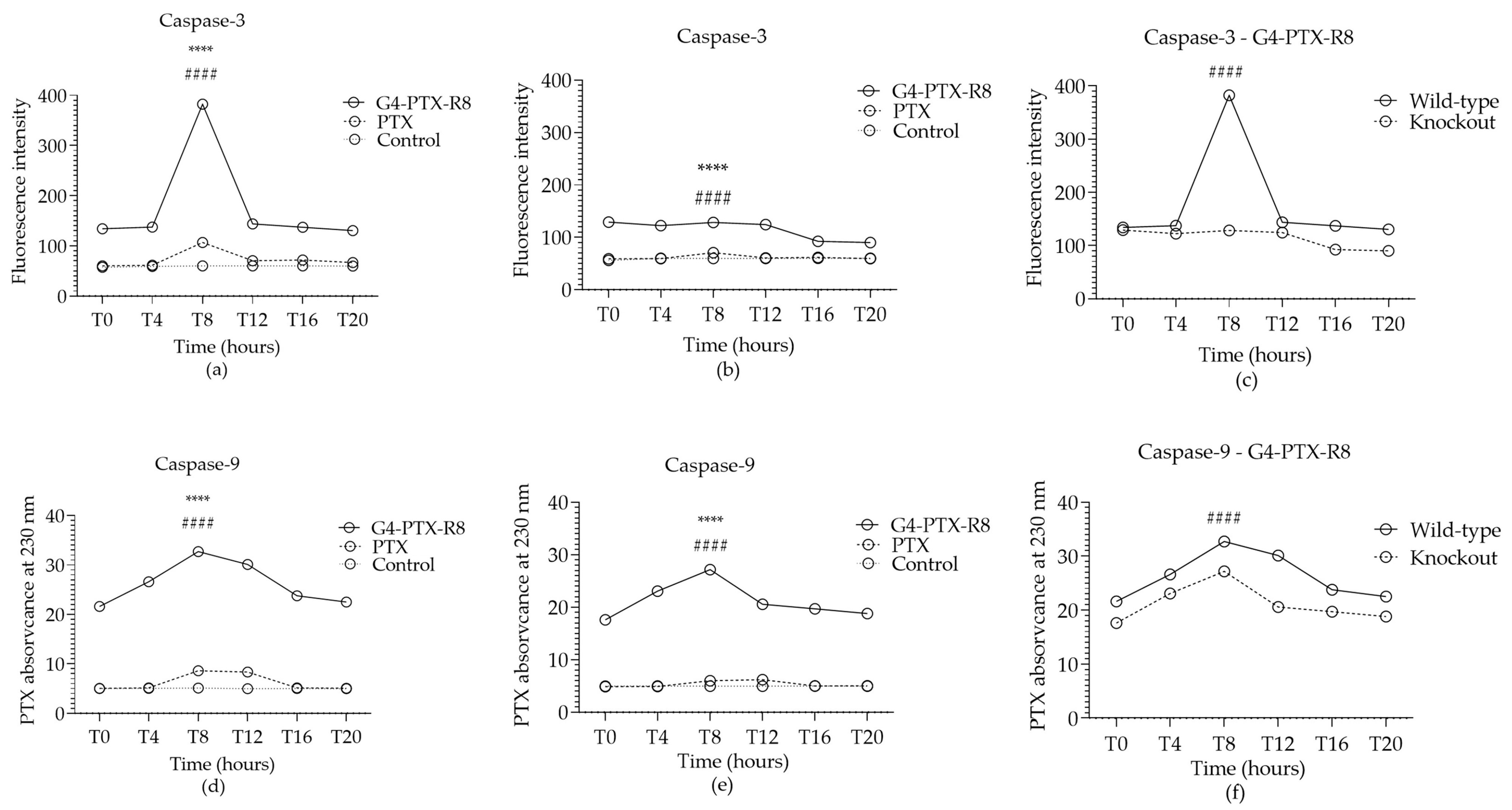
2.14. Caspase-3 and Caspase-9 Activity Assay
2.15. Statistical Analysis
3. Results
3.1. Preparation and Characterization of Multifunctional Dendrimer Conjugate
3.2. Cytotoxicity Studies
3.3. The Impact of Circadian Rhythm on Cellular Uptake/Internalization
3.3.1. Determination of Cell-Associated PTX
3.3.2. Generation of a Bmal1 Knockout Cell Line
3.3.3. Bmal1 Silencing Effect on Cellular Internalization of PTX
3.3.4. The Role of Bmal1 on Therapeutic Effect Mediated by G4-PTX-R8
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Pfeffer, C.M.; Singh, A.T.K. Apoptosis: A Target for Anticancer Therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef] [PubMed]
- Gavas, S.; Quazi, S.; Karpiński, T.M. Nanoparticles for Cancer Therapy: Current Progress and Challenges. Nanoscale Res. Lett. 2021, 16, 173. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Shen, J.; Wang, J.; Yang, X.; Dong, S.; Lu, S. Nanoparticle-Based Drug Delivery System: A Patient-Friendly Chemotherapy for Oncology. Dose-Response 2020, 18, 1559325820936161. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Wang, K.; Oppong-Gyebi, A.; Hu, J. Application of Nanotechnology in Cancer Diagnosis and Therapy—A Mini-Review. Int. J. Med. Sci. 2020, 17, 2964–2973. [Google Scholar] [CrossRef] [PubMed]
- Edis, Z.; Wang, J.; Waqas, M.K.; Ijaz, M.; Ijaz, M. Nanocarriers-Mediated Drug Delivery Systems for Anticancer Agents: An Overview and Perspectives. Int. J. Nanomed. 2021, 16, 1313–1330. [Google Scholar] [CrossRef]
- Blanco, E.; Shen, H.; Ferrari, M. Principles of nanoparticle design for overcoming biological barriers to drug delivery. Nat. Biotechnol. 2015, 33, 941–951. [Google Scholar] [CrossRef]
- Wang, X.; Qiu, Y.; Wang, M.; Zhang, C.; Zhang, T.; Zhou, H.; Zhao, W.; Zhao, W.; Xia, G.; Shao, R. Endocytosis and Organelle Targeting of Nanomedicines in Cancer Therapy. Int. J. Nanomed. 2020, 15, 9447–9467. [Google Scholar] [CrossRef]
- Singh, V.; Sahebkar, A.; Kesharwani, P. Poly (propylene imine) dendrimer as an emerging polymeric nanocarrier for anticancer drug and gene delivery. Eur. Polym. J. 2021, 158, 110683. [Google Scholar] [CrossRef]
- Nimesh, S. 13—Dendrimers. In Gene Therapy; Nimesh, S., Ed.; Woodhead Publishing: Sawston, UK, 2013; pp. 259–285. [Google Scholar] [CrossRef]
- Karandikar, S.; Mirani, A.; Waybhase, V.; Patravale, V.B.; Patankar, S. Chapter 10—Nanovaccines for oral delivery-formulation strategies and challenges. In Nanostructures for Oral Medicine; Andronescu, E., Grumezescu, A.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 263–293. [Google Scholar] [CrossRef]
- Rompicharla, S.V.K.; Kumari, P.; Ghosh, B.; Biswas, S. Octa-arginine modified poly(amidoamine) dendrimers for improved delivery and cytotoxic effect of paclitaxel in cancer. Artif. Cells Nanomed. Biotechnol. 2018, 46, 847–859. [Google Scholar] [CrossRef]
- Khatri, S.; Das, N.G.; Das, S.K. Effect of methotrexate conjugated PAMAM dendrimers on the viability of MES-SA uterine cancer cells. J. Pharm. Bioallied Sci. 2014, 6, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhu, J.; Zheng, Y.; Guo, R.; Wang, S.; Mignani, S.; Caminade, A.M.; Majoral, J.P.; Shi, X. Doxorubicin-Conjugated PAMAM Dendrimers for pH-Responsive Drug Release and Folic Acid-Targeted Cancer Therapy. Pharmaceutics 2018, 10, 162. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zuo, A.; Huang, X.; Ying, Y.; Shu, X.; Chen, X.; Yang, Y.; Ma, J.; Lin, G.; Wang, X.; et al. Docetaxel-loaded PAMAM-based poly (γ-benzyl-l-glutamate)-b-d-α-tocopheryl polyethylene glycol 1000 succinate nanoparticles in human breast cancer and human cervical cancer therapy. J. Microencapsul. 2019, 36, 552–565. [Google Scholar] [CrossRef]
- Boisguérin, P.; Konate, K.; Josse, E.; Vivès, E.; Deshayes, S. Peptide-Based Nanoparticles for Therapeutic Nucleic Acid Delivery. Biomedicines 2021, 9, 583. [Google Scholar] [CrossRef] [PubMed]
- Neves, A.R.; Albuquerque, T.; Faria, R.; Gonçalves, A.M.; Santos, C.; Vivès, E.; Boisguérin, P.; Passarinha, L.A.; Sousa, Â.; Costa, D. Development of WRAP5 Peptide Complexes for Targeted Drug/Gene Co-Delivery toward Glioblastoma Therapy. Pharmaceutics 2022, 14, 2213. [Google Scholar] [CrossRef]
- Bhatt, H.; Ghosh, B.; Biswas, S. Cell-Penetrating Peptide and α-Tocopherol-Conjugated Poly(amidoamine) Dendrimers for Improved Delivery and Anticancer Activity of Loaded Paclitaxel. ACS Appl. Bio Mater. 2020, 3, 3157–3169. [Google Scholar] [CrossRef]
- Herman, J.H. Chapter 9—Circadian Rhythm Disorders: Diagnosis and Treatment. In Principles and Practice of Pediatric Sleep Medicine; Sheldon, S.H., Ferber, R., Kryger, M.H., Eds.; W.B. Saunders: Philadelphia, PA, USA, 2005; pp. 101–111. [Google Scholar] [CrossRef]
- Innominato, P.F.; Ballesta, A.; Huang, Q.; Focan, C.; Chollet, P.; Karaboué, A.; Giacchetti, S.; Bouchahda, M.; Adam, R.; Garufi, C.; et al. Sex-dependent least toxic timing of irinotecan combined with chronomodulated chemotherapy for metastatic colorectal cancer: Randomized multicenter EORTC 05011 trial. Cancer Med. 2020, 9, 4148–4159. [Google Scholar] [CrossRef]
- Zhang, P.X.; Jin, F.; Li, Z.L.; Wu, W.L.; Li, Y.Y.; Long, J.H.; Chen, G.Y.; Chen, X.X.; Gan, J.Y.; Gong, X.Y.; et al. A randomized phase II trial of induction chemotherapy followed by cisplatin chronotherapy versus constant rate delivery combined with radiotherapy. Chronobiol. Int. 2018, 35, 240–248. [Google Scholar] [CrossRef]
- Kettner, N.M.; Voicu, H.; Finegold, M.J.; Coarfa, C.; Sreekumar, A.; Putluri, N.; Katchy, C.A.; Lee, C.; Moore, D.D.; Fu, L. Circadian Homeostasis of Liver Metabolism Suppresses Hepatocarcinogenesis. Cancer Cell 2016, 30, 909–924. [Google Scholar] [CrossRef]
- Hanley, K.L.; Liang, Y.; Wang, G.; Lin, X.; Yang, M.; Karin, M.; Fu, W.; Feng, G.-S. Concurrent Disruption of the Ras/MAPK and NF-κB Pathways Induces Circadian Deregulation and Hepatocarcinogenesis. Mol. Cancer Res. 2022, 20, 337–349. [Google Scholar] [CrossRef]
- Pett, J.P.; Kondoff, M.; Bordyugov, G.; Kramer, A.; Herzel, H. Co-existing feedback loops generate tissue-specific circadian rhythms. Life Sci. Alliance 2018, 1, e201800078. [Google Scholar] [CrossRef] [PubMed]
- Slat, E.A.; Sponagel, J.; Marpegan, L.; Simon, T.; Kfoury, N.; Kim, A.; Binz, A.; Herzog, E.D.; Rubin, J.B. Cell-intrinsic, Bmal1-dependent Circadian Regulation of Temozolomide Sensitivity in Glioblastoma. J. Biol. Rhythm. 2017, 32, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Dulong, S.; Ballesta, A.; Okyar, A.; Lévi, F. Identification of Circadian Determinants of Cancer Chronotherapy through In Vitro Chronopharmacology and Mathematical Modeling. Mol. Cancer Ther. 2015, 14, 2154–2164. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, T.; Neves, A.R.; Quintela, T.; Costa, D. The Influence of Circadian Rhythm on Cancer Cells Targeting and Transfection Efficiency of a Polycation-Drug/Gene Delivery Vector. Polymers 2022, 14, 681. [Google Scholar] [CrossRef]
- Majoros, I.J.; Myc, A.; Thomas, T.; Mehta, C.B.; Baker, J.R. PAMAM Dendrimer-Based Multifunctional Conjugate for Cancer Therapy: Synthesis, Characterization, and Functionality. Biomacromolecules 2006, 7, 572–579. [Google Scholar] [CrossRef]
- Sousa-Herves, A.; Würfel, P.; Wegner, N.; Khandare, J.; Licha, K.; Haag, R.; Welker, P.; Calderón, M. Dendritic polyglycerol sulfate as a novel platform for paclitaxel delivery: Pitfalls of ester linkage. Nanoscale 2015, 7, 3923–3932. [Google Scholar] [CrossRef]
- Paul, M.; Itoo, A.M.; Ghosh, B.; Biswas, S. Hypoxia alleviating platinum(IV)/chlorin e6-based combination chemotherapeutic-photodynamic nanomedicine for oropharyngeal carcinoma. J. Photochem. Photobiol. B Biol. 2023, 238, 112627. [Google Scholar] [CrossRef]
- Lin, R.; Zhang, P.; Cheetham, A.G.; Walston, J.; Abadir, P.; Cui, H. Dual peptide conjugation strategy for improved cellular uptake and mitochondria targeting. Bioconjugate Chem. 2015, 26, 71–77. [Google Scholar] [CrossRef]
- Vysyaraju, N.R.; Paul, M.; Ch, S.; Ghosh, B.; Biswas, S. Olaparib@human serum albumin nanoparticles as sustained drug-releasing tumour-targeting nanomedicine to inhibit growth and metastasis in the mouse model of triple-negative breast cancer. J. Drug Target. 2022, 30, 1088–1105. [Google Scholar] [CrossRef]
- Kumbham, S.; Paul, M.; Itoo, A.; Ghosh, B.; Biswas, S. Oleanolic acid-conjugated human serum albumin nanoparticles encapsulating doxorubicin as synergistic combination chemotherapy in oropharyngeal carcinoma and melanoma. Int. J. Pharm. 2022, 614, 121479. [Google Scholar] [CrossRef]
- Neves, A.R.; Albuquerque, T.; Faria, R.; Paul, M.; Biswas, S.; Sousa, Â.; Costa, D. Development of Tailor-Made Dendrimer Ternary Complexes for Drug/Gene Co-Delivery in Cancer. Pharmaceutics 2021, 13, 1256. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.; Gomes, D.; Neto, M.; Passarinha, L.A.; Costa, D.; Sousa, Â. Development and Characterization of Quercetin-Loaded Delivery Systems for Increasing Its Bioavailability in Cervical Cancer Cells. Pharmaceutics 2023, 15, 936. [Google Scholar] [CrossRef] [PubMed]
- Vuelta, E.; Ordonez, J.; Alonso Pérez, V.; Mendez, L.; Hernández-Carabias, P.; Saldana, R.; Sevilla, J.; Sebastian, E.; Muntion, S.; Sanchez-Guijo, F.; et al. CRISPR/Cas9 technology abolishes the BCR/ABL1 oncogene effect in chronic myeloid leukemia and restores normal hematopoiesis. bioRxiv 2020, 103–112. [Google Scholar] [CrossRef]
- Cai, H.; Xiang, Y.; Zeng, Y.; Li, Z.; Zheng, X.; Luo, Q.; Zhu, H.; Gong, Q.; Gu, Z.; Liu, Y.; et al. Cathepsin B-responsive and gadolinium-labeled branched glycopolymer-PTX conjugate-derived nanotheranostics for cancer treatment. Acta Pharm. Sin. B 2021, 11, 544–559. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Murata, T.; Kawaguchi, K.; Kiyuna, T.; Igarashi, K.; Hwang, H.K.; Hiroshima, Y.; Hozumi, C.; Komatsu, S.; Kikuchi, T.; et al. Cervical Cancer Patient-Derived Orthotopic Xenograft (PDOX) Is Sensitive to Cisplatinum and Resistant to Nab-paclitaxel. Anticancer Res. 2017, 37, 61–65. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, K.; Zhu, X.; Zheng, X.; Wang, C.; Niu, X.; Jiang, T.; Ji, X.; Zhao, W.; Pang, L.; et al. Synergistic Inhibition of Drug-Resistant Colon Cancer Growth with PI3K/mTOR Dual Inhibitor BEZ235 and Nano-Emulsioned Paclitaxel via Reducing Multidrug Resistance and Promoting Apoptosis. Int. J. Nanomed. 2021, 16, 2173–2186. [Google Scholar] [CrossRef]
- Nawara, H.M.; Afify, S.M.; Hassan, G.; Zahra, M.H.; Seno, A.; Seno, M. Paclitaxel-Based Chemotherapy Targeting Cancer Stem Cells from Mono- to Combination Therapy. Biomedicines 2021, 9, 500. [Google Scholar] [CrossRef]
- Mudalige, T.; Qu, H.; Haute, D.; Ansar, S.; Paredes, A.; Ingle, T. Characterization of Nanomaterials. In Nanomaterials for Food Applications; Elsevier: Amsterdam, The Netherlands, 2019; pp. 313–353. [Google Scholar] [CrossRef]
- Valsalakumari, R.; Yadava, S.K.; Szwed, M.; Pandya, A.D.; Mælandsmo, G.M.; Torgersen, M.L.; Iversen, T.-G.; Skotland, T.; Sandvig, K.; Giri, J. Mechanism of cellular uptake and cytotoxicity of paclitaxel loaded lipid nanocapsules in breast cancer cells. Int. J. Pharm. 2021, 597, 120217. [Google Scholar] [CrossRef]
- Kreuter, J. Influence of chronobiology on the nanoparticle-mediated drug uptake into the brain. Pharmaceutics 2015, 7, 3–9. [Google Scholar] [CrossRef]
- Hu, J.; Fu, S.; Peng, Q.; Han, Y.; Xie, J.; Zan, N.; Chen, Y.; Fan, J. Paclitaxel-loaded polymeric nanoparticles combined with chronomodulated chemotherapy on lung cancer: In vitro and in vivo evaluation. Int. J. Pharm. 2017, 516, 313–322. [Google Scholar] [CrossRef]
- García-Tuñón, I.; Vuelta, E.; Pérez-Ramos, S.; Hernández, J.; Méndez, L.; Herrero, M.; Sanchez-Martin, M. CRISPR-ERA for Switching Off (Onco) Genes. In Modulating Gene Expression-Abridging the RNAi and CRISPR-Cas9 Technologies; Intechopen: London, UK, 2018. [Google Scholar] [CrossRef]
- Golombek, D.A.; Casiraghi, L.P.; Agostino, P.V.; Paladino, N.; Duhart, J.M.; Plano, S.A.; Chiesa, J.J. The times they’re a-changing: Effects of circadian desynchronization on physiology and disease. J. Physiol. 2013, 107, 310–322. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Zhang, T.; Zhou, C.; Xu, H.; Guo, L.; Chen, M.; Wu, B. The Circadian Clock Gene Bmal1 Controls Intestinal Exporter MRP2 and Drug Disposition. Theranostics 2019, 9, 2754–2767. [Google Scholar] [CrossRef] [PubMed]
- Andrews, J.L.; Zhang, X.; McCarthy, J.J.; McDearmon, E.L.; Hornberger, T.A.; Russell, B.; Campbell, K.S.; Arbogast, S.; Reid, M.B.; Walker, J.R.; et al. CLOCK and BMAL1 regulate MyoD and are necessary for maintenance of skeletal muscle phenotype and function. Proc. Natl. Acad. Sci. USA 2010, 107, 19090–19095. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.-H.; Joo, H.; Lee, K.; Kim, H.; Didier, R.; Yang, Y.; Shin, H.; Lee, C. Streamlined procedure for gene knockouts using all-in-one adenoviral CRISPR-Cas9. Sci. Rep. 2019, 9, 277. [Google Scholar] [CrossRef]
- Fox, L.J.; Richardson, R.M.; Briscoe, W.H. PAMAM dendrimer—Cell membrane interactions. Adv. Colloid. Interface Sci. 2018, 257, 1–18. [Google Scholar] [CrossRef]
- Green, D.R.; Llambi, F. Cell Death Signaling. Cold Spring Harb. Perspect. Biol. 2015, 7, a006080. [Google Scholar] [CrossRef]
- Li, P.; Zhou, L.; Zhao, T.; Liu, X.; Zhang, P.; Liu, Y.; Zheng, X.; Li, Q. Caspase—9: Structure, mechanisms and clinical application. Oncotarget 2017, 8, 23996–24008. [Google Scholar] [CrossRef]
- Lossi, L.; Castagna, C.; Merighi, A. Caspase-3 Mediated Cell Death in the Normal Development of the Mammalian Cerebellum. Int. J. Mol. Sci. 2018, 19, 3999. [Google Scholar] [CrossRef]
- Fulda, S.; Debatin, K.M. Extrinsic versus intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene 2006, 25, 4798–4811. [Google Scholar] [CrossRef]
- Morales-Cano, D.; Calviño, E.; Rubio, V.; Herráez, A.; Sancho, P.; Tejedor, M.C.; Diez, J.C. Apoptosis induced by paclitaxel via Bcl-2, Bax and caspases 3 and 9 activation in NB4 human leukaemia cells is not modulated by ERK inhibition. Exp. Toxicol. Pathol. Off. J. Ges. Fur Toxikol. Pathol. 2013, 65, 1101–1108. [Google Scholar] [CrossRef]
- Miller, A.V.; Hicks, M.A.; Nakajima, W.; Richardson, A.C.; Windle, J.J.; Harada, H. Paclitaxel-Induced Apoptosis Is BAK-Dependent, but BAX and BIM-Independent in Breast Tumor. PLoS ONE 2013, 8, e60685. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, Y.; Subban, K.; Chelliah, J. A novel synergistic anticancer effect of fungal cholestanol glucoside and paclitaxel: Apoptosis induced by an intrinsic pathway through ROS generation in cervical cancer cell line (HeLa). Toxicol. In Vitro 2021, 72, 105079. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Tang, Y.; Wang, R.; Najafi, M. Mechanisms of cancer cell death induction by paclitaxel: An updated review. Apoptosis 2022, 27, 647–667. [Google Scholar] [CrossRef] [PubMed]
- Kheraldine, H.; Gupta, I.; Alhussain, H.; Jabeen, A.; Cyprian, F.S.; Akhtar, S.; Al Moustafa, A.E.; Rachid, O. Substantial cell apoptosis provoked by naked PAMAM dendrimers in HER2-positive human breast cancer via JNK and ERK1/ERK2 signalling pathways. Comput. Struct. Biotechnol. J. 2021, 19, 2881–2890. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Fong, S.Y.; Shon, J.; Zhang, S.L.; Brooks, R.; Lahens, N.F.; Chen, D.; Dang, C.V.; Field, J.M.; Sehgal, A. Time-of-day specificity of anticancer drugs may be mediated by circadian regulation of the cell cycle. Sci. Adv. 2021, 7, eabd2645. [Google Scholar] [CrossRef]
- Wang, F.; Li, C.; Yongluo; Chen, L. The Circadian Gene Clock Plays an Important Role in Cell Apoptosis and the DNA Damage Response In Vitro. Technol. Cancer Res. Treat. 2016, 15, 480–486. [Google Scholar] [CrossRef]
- Sadiq, Z.; Varghese, E.; Büsselberg, D. Cisplatin’s dual-effect on the circadian clock triggers proliferation and apoptosis. Neurobiol. Sleep Circadian Rhythm. 2020, 9, 100054. [Google Scholar] [CrossRef]
- Gwon, D.H.; Lee, W.Y.; Shin, N.; Kim, S.I.; Jeong, K.; Lee, W.H.; Kim, D.W.; Hong, J.; Lee, S.Y. BMAL1 Suppresses Proliferation, Migration, and Invasion of U87MG Cells by Downregulating Cyclin B1, Phospho-AKT, and Metalloproteinase-9. Int. J. Mol. Sci. 2020, 21, 2352. [Google Scholar] [CrossRef]
- Tang, Q.; Cheng, B.; Xie, M.; Chen, Y.; Zhao, J.; Zhou, X.; Chen, L. Circadian Clock Gene Bmal1 Inhibits Tumorigenesis and Increases Paclitaxel Sensitivity in Tongue Squamous Cell Carcinoma. Cancer Res. 2017, 77, 532–544. [Google Scholar] [CrossRef]
- Zhao, C.; Meng, X.; Li, Y.; Liu, L.; He, Q.; Jiang, J.; Chen, Y.; Li, X.; Li, Y.; Tang, Y.; et al. Circadian clock gene BMAL1 inhibits the proliferation and tumor-formation ability of nasopharyngeal carcinoma cells and increases the sensitivity of radiotherapy. Chronobiol. Int. 2022, 39, 1340–1351. [Google Scholar] [CrossRef]
- Chen, W.D.; Wen, M.S.; Shie, S.S.; Lo, Y.L.; Wo, H.T.; Wang, C.C.; Hsieh, I.C.; Lee, T.H.; Wang, C.Y. The circadian rhythm controls telomeres and telomerase activity. Biochem. Biophys. Res. Commun. 2014, 451, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Qu, M.; Zhang, G.; Qu, H.; Vu, A.; Wu, R.; Tsukamoto, H.; Jia, Z.; Huang, W.; Lenz, H.-J.; Rich, J.N.; et al. Circadian regulator BMAL1::CLOCK promotes cell proliferation in hepatocellular carcinoma by controlling apoptosis and cell cycle. Proc. Natl. Acad. Sci. USA 2023, 120, e2214829120. [Google Scholar] [CrossRef] [PubMed]
- Elshazley, M.; Sato, M.; Hase, T.; Yamashita, R.; Yoshida, K.; Toyokuni, S.; Ishiguro, F.; Osada, H.; Sekido, Y.; Yokoi, K.; et al. The circadian clock gene BMAL1 is a novel therapeutic target for malignant pleural mesothelioma. Int. J. Cancer 2012, 131, 2820–2831. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Zeta Potential (mV) | PdI | Size (nm) | |
|---|---|---|---|
| PAMAM G4-PTX-R8 | +9.325 | 0.287 | 23.74 ± 0.454 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albuquerque, T.; Neves, A.R.; Paul, M.; Biswas, S.; Vuelta, E.; García-Tuñón, I.; Sánchez-Martin, M.; Quintela, T.; Costa, D. A Potential Effect of Circadian Rhythm in the Delivery/Therapeutic Performance of Paclitaxel–Dendrimer Nanosystems. J. Funct. Biomater. 2023, 14, 362. https://doi.org/10.3390/jfb14070362
Albuquerque T, Neves AR, Paul M, Biswas S, Vuelta E, García-Tuñón I, Sánchez-Martin M, Quintela T, Costa D. A Potential Effect of Circadian Rhythm in the Delivery/Therapeutic Performance of Paclitaxel–Dendrimer Nanosystems. Journal of Functional Biomaterials. 2023; 14(7):362. https://doi.org/10.3390/jfb14070362
Chicago/Turabian StyleAlbuquerque, Tânia, Ana Raquel Neves, Milan Paul, Swati Biswas, Elena Vuelta, Ignacio García-Tuñón, Manuel Sánchez-Martin, Telma Quintela, and Diana Costa. 2023. "A Potential Effect of Circadian Rhythm in the Delivery/Therapeutic Performance of Paclitaxel–Dendrimer Nanosystems" Journal of Functional Biomaterials 14, no. 7: 362. https://doi.org/10.3390/jfb14070362
APA StyleAlbuquerque, T., Neves, A. R., Paul, M., Biswas, S., Vuelta, E., García-Tuñón, I., Sánchez-Martin, M., Quintela, T., & Costa, D. (2023). A Potential Effect of Circadian Rhythm in the Delivery/Therapeutic Performance of Paclitaxel–Dendrimer Nanosystems. Journal of Functional Biomaterials, 14(7), 362. https://doi.org/10.3390/jfb14070362