
Cytotoxicity of Methacrylate Dental Resins to Human Gingival Fibroblasts

 ,
,
Abstract

1. Introduction
2. Materials and Methods
2.1. Preparation of Resins
2.2. Cell Culture
2.3. Cytotoxicity and Cell Cycle Assays
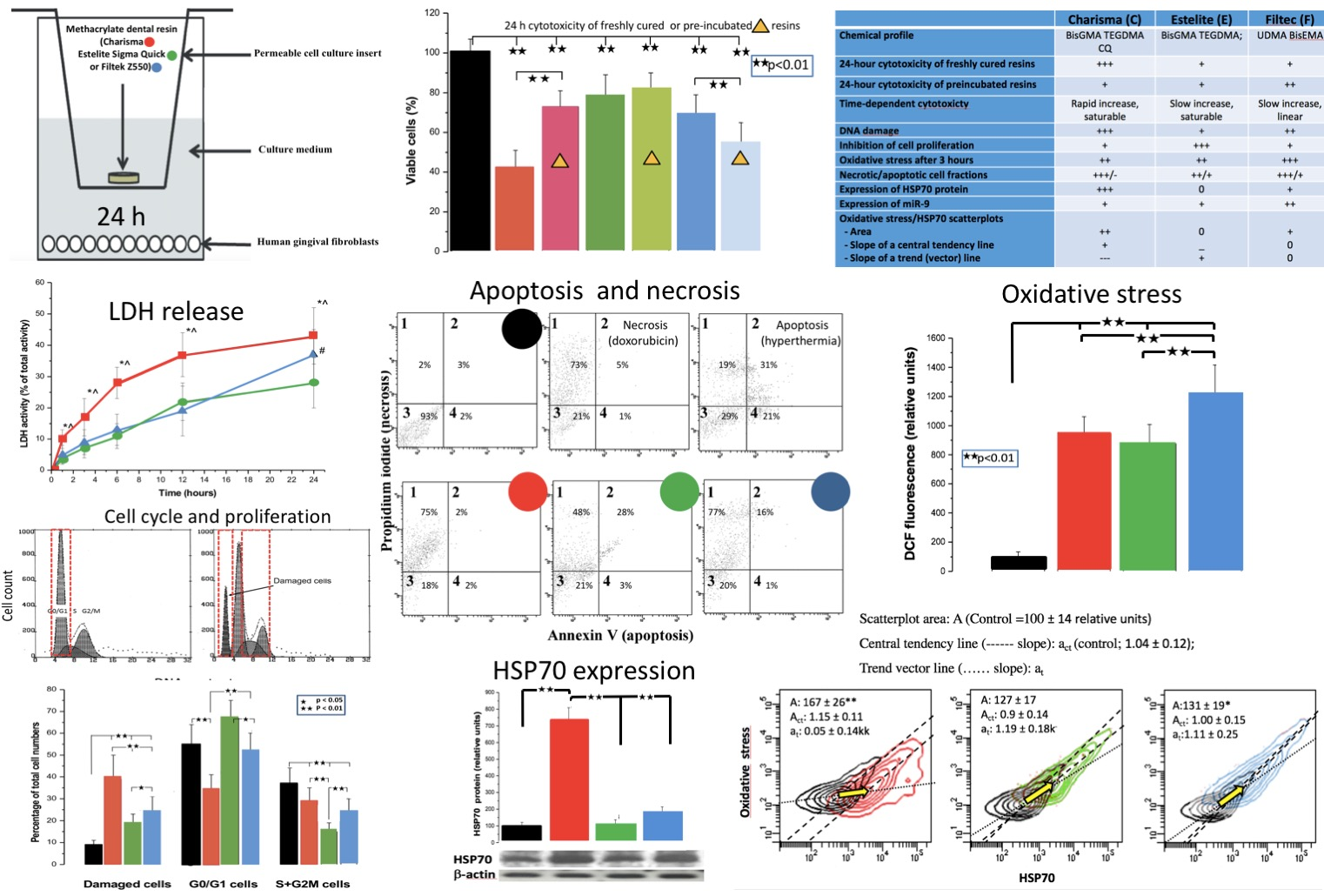
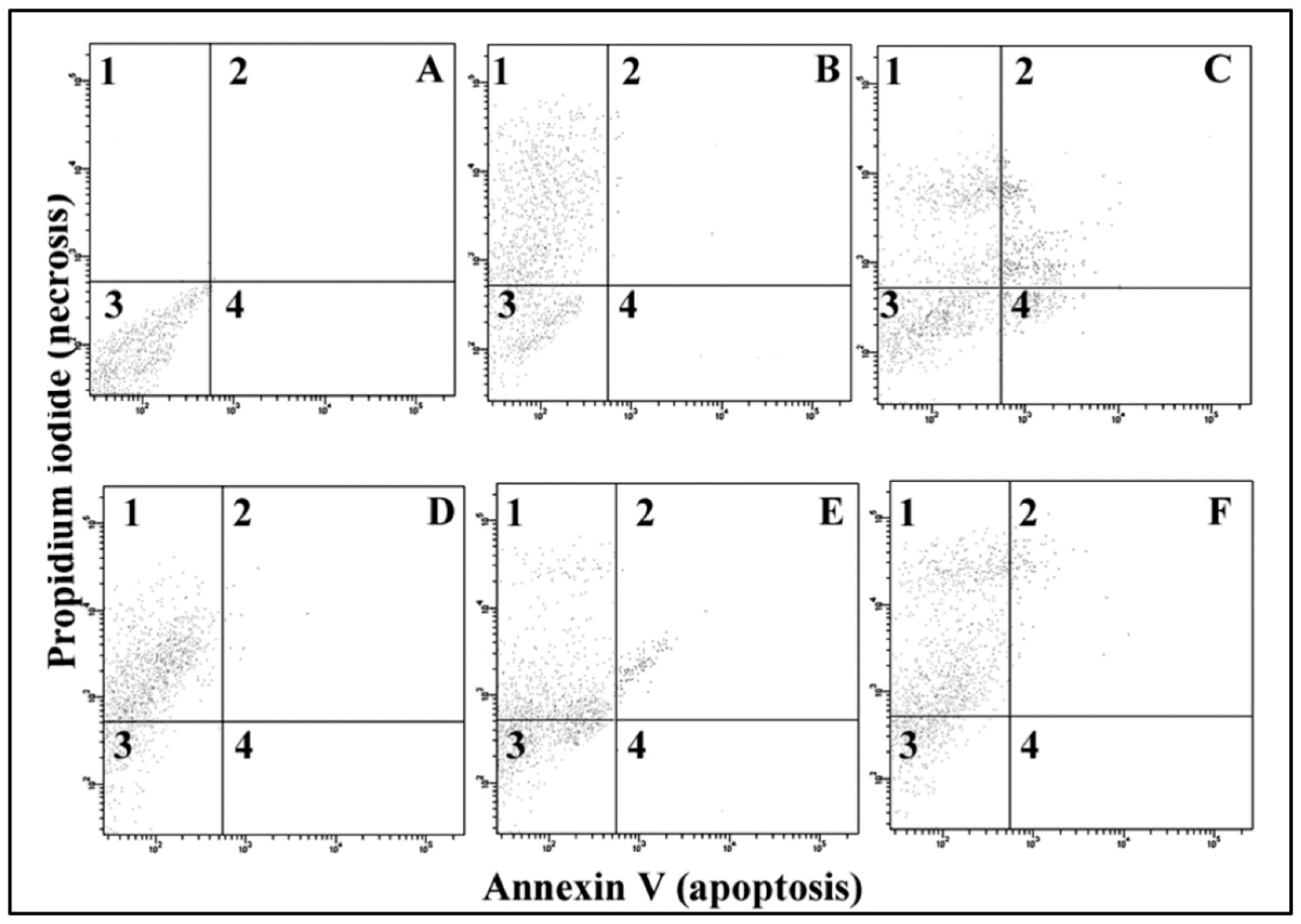
2.4. Apoptosis/Necrosis
2.5. Oxidative Stress
2.6. HSP70 Expression
2.7. MiR-9 Assay
2.8. Statistical Analysis
3. Results
3.1. MTT Test
3.2. Cell Cycle
3.3. Cell Membrane Damage
3.4. Apoptosis/Necrosis
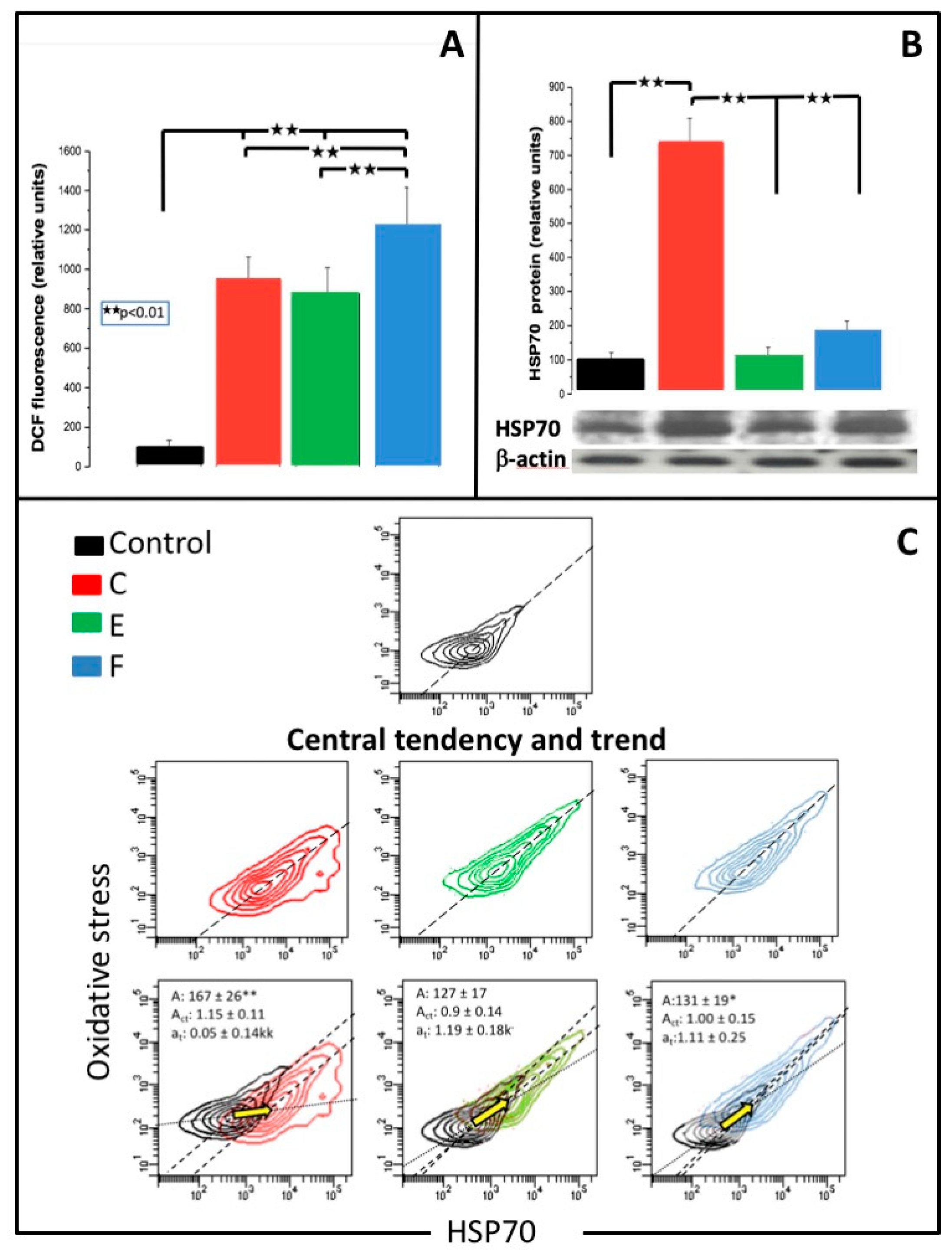
3.5. Oxidative Stress
3.6. MiR-9 and HSP70
3.7. Binary Scatter Plots of Oxidative Stress and HSP70
4. Discussion
4.1. Resin Cytotoxicity
Delayed Cytotoxicity and Cell Cycle
4.2. Oxidative Stress
4.3. Necrosis and Apoptosis
4.4. MiR-9 Expression
4.5. HSP70 Expression and HSP70/DCF Assay
4.6. Conclusion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mahadevan, G.; Valiyaveettil, S. Understanding the interactions of poly (methyl methacrylate) and poly(vinyl chloride) nanoparticles with BHK-21 cell line. Sci. Rep. 2021, 11, 2089. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-H.; Shih, W.-C.; Wang, Y.-L.; Tsai, Y.-L.; Chen, Y.-J.; Chang, M.-C.; Jeng, J.-H. Cytotoxicity and genotoxicity of DMABEE, a co-photoinitiator of resin polymerization, on CHO-K1 cells: Role of redox and carboxylesterase. J. Biomed. Mater. Res. Part B: Appl. Biomater. 2020, 108, 2088–2098. [Google Scholar] [CrossRef]
- Kraus, D.; Wolfgarten, M.; Enkling, N.; Helfgen, E.-H.; Frentzen, M.; Probstmeier, R.; Winter, J.; Stark, H. In-vitro cytocompatibility of dental resin monomers on osteoblast-like cells. J. Dent. 2017, 65, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Neves, S.O.; Magalhães, L.M.D.; Corrêa, J.D.; Dutra, W.O.; Gollob, K.J.; Silva, T.A.; Horta, M.C.R.; Souza, P.E.A. Composite-derived monomers affect cell viability and cytokine expression in human leukocytes stimulated with Porphyromonas gingivalis. J. Appl. Oral Sci. 2019, 27, e20180529. [Google Scholar] [CrossRef] [PubMed]
- Krifka, S.; Spagnuolo, G.; Schmalz, G.; Schweikl, H. A review of adaptive mechanisms in cell responses towards oxidative stress caused by dental resin monomers. Biomaterials 2013, 34, 4555–4563. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Saxena, P.; Pant, V.A.; Pant, A.B. Release and toxicity of dental resin composite. Toxicol. Int. 2012, 19, 225–234. [Google Scholar] [CrossRef]
- Szczesio-Wlodarczyk, A.; Sokolowski, J.; Kleczewska, J.; Bociong, K. Ageing of Dental Composites Based on Methacrylate Resins—A Critical Review of the Causes and Method of Assessment. Polymers 2020, 12, 882. [Google Scholar] [CrossRef]
- Ponce-Bravo, S.; Ledesma-Montes, C.; Martínez-Rivera, J.-L.; Garcés-Ortíz, M. Toxicity test of a dental commercial composite. J. Clin. Exp. Dent. 2015, 7, e289–e292. [Google Scholar] [CrossRef]
- Schneider, T.R.; Hakami-Tafreshi, R.; Tomasino-Perez, A.; Tayebi, L.; Lobner, D. Effects of dental composite resin monomers on dental pulp cells. Dent. Mater. J. 2019, 38, 579–583. [Google Scholar] [CrossRef]
- Eteti, G.; Orsini, G.; Salvatore, V.; Efocaroli, S.; Emazzotti, M.C.; Eruggeri, A.; Mattioli-Belmonte, M.; Efalconi, M. HEMA but not TEGDMA induces autophagy in human gingival fibroblasts. Front. Physiol. 2015, 6, 275. [Google Scholar] [CrossRef]
- Gallorini, M.; Petzel, C.; Bolay, C.; Hiller, K.-A.; Cataldi, A.; Buchalla, W.; Krifka, S.; Schweikl, H. Activation of the Nrf2-regulated antioxidant cell response inhibits HEMA-induced oxidative stress and supports cell viability. Biomaterials 2015, 56, 114–128. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.-L. The effects of cytotoxicity and genotoxicity induced by 2,2-bis[4-(acryloxypropoxy)phenyl]propane via caspases in human gingival fibroblasts. Toxicol. Ind. Health 2014, 30, 755–764. [Google Scholar] [CrossRef]
- Bakopoulou, A.; Leyhausen, G.; Volk, J.; Papachristou, E.; Koidis, P.; Geurtsen, W. Effects of resinous monomers on the odontogenic differentiation and mineralization potential of highly proliferative and clonogenic cultured apical papilla stem cells. Dent. Mater. 2012, 28, 327–339. [Google Scholar] [CrossRef]
- Styllou, M.; Reichl, F.-X.; Styllou, P.; Urcan, E.; Rothmund, L.; Hickel, R.; Högg, C.; Scherthan, H. Dental composite components induce DNA-damage and altered nuclear morphology in gingiva fibroblasts. Dent. Mater. 2015, 31, 1335–1344. [Google Scholar] [CrossRef] [PubMed]
- Ginzkey, C.; Zinnitsch, S.; Steussloff, G.; Koehler, C.; Hackenberg, S.; Hagen, R.; Kleinsasser, N.H.; Froelich, K. Assessment of HEMA and TEGDMA induced DNA damage by multiple genotoxicological endpoints in human lymphocytes. Dent. Mater. 2015, 31, 865–876. [Google Scholar] [CrossRef] [PubMed]
- Durner, J.; Wellner, P.; Hickel, R.; Reichl, F. Synergistic interaction caused to human gingival fibroblasts from dental monomers. Dent. Mater. 2012, 28, 818–823. [Google Scholar] [CrossRef] [PubMed]
- Al-Hiyasat, A.S.; Darmani, H.; Milhem, M.M. Cytotoxicity evaluation of dental resin composites and their flowable derivatives. Clin. Oral Investig. 2005, 9, 21–25. [Google Scholar] [CrossRef]
- Ilie, N.; Kreppel, I.; Durner, J. Effect of radical amplified photopolymerization (RAP) in resin-based composites. Clin. Oral Investig. 2014, 18, 1081–1088. [Google Scholar] [CrossRef]
- Luczaj-Cepowicz, E.; Marczuk-Kolada, G.; Pawinska, M.; Obidzinska, M.; Holownia, A. Evaluation of cytotoxicity and pH changes generated by various dental pulp capping materials—An in vitro study. Folia Histochem. et Cytobiol. 2017, 55, 86–93. [Google Scholar] [CrossRef][Green Version]
- Ipek, S.; Üstündağ, A.; Eke, B.C. Three-dimensional (3D) cell culture studies: A review of the field of toxicology. Drug Chem. Toxicol. 2022, 1–11. [Google Scholar] [CrossRef]
- Khafaei, M.; Rezaie, E.; Mohammadi, A.; Gerdehsang, P.S.; Ghavidel, S.; Kadkhoda, S.; Zahra, A.Z.; Forouzanfar, N.; Arabameri, H.; Tavallaie, M. miR-9: From function to therapeutic potential in cancer. J. Cell. Physiol. 2019, 234, 14651–14665. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Fernández, M.R.; Valpuesta, J.M. Hsp70 chaperone: A master player in protein homeostasis. F1000Research 2018, 7, 1497. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, M.; Fornasiero, M.C.; Isetta, A.M. MTT colorimetric assay for testing macrophage cytotoxic activity in vitro. J. Immunol. Methods 1990, 131, 165–172. [Google Scholar] [CrossRef]
- Rousselle, C.; Robert-Nicoud, M.; Ronot, X. Flow cytometric analysis of DNA content of living and fixed cells: A comparative study using various fixatives. Histochem. J. 1998, 30, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Gougeon, M.-L.; Lecoeur, H.; Prévost, M. Oncosis is associated with exposure of phosphatidylserine residues on the outside layer of the plasma membrane: A reconsideration of the specificity of the annexin V/propidium iodide assay. Cytometry 2001, 44, 65–72. [Google Scholar] [CrossRef]
- Ubezio, P.; Civoli, F. Flow cytometric detection of hydrogen peroxide production induced by doxorubicin in cancer cells. Free Radic. Biol. Med. 1994, 16, 509–516. [Google Scholar] [CrossRef]
- Aranha, A.M.F.; Giro, E.M.A.; Hebling, J.; Lessa, F.; Costa, C.A.D.S. Effects of light-curing time on the cytotoxicity of a restorative composite resin on odontoblast-like cells. J. Appl. Oral Sci. 2010, 18, 461–466. [Google Scholar] [CrossRef]
- Nalçaci, A.; Ulusoy, N.; Atakol, O.; Nalçacı, A. Time-based Elution of TEGDMA and BisGMA from Resin Composite Cured with LED, QTH and High-intensity QTH Lights. Oper. Dent. 2006, 31, 197–203. [Google Scholar] [CrossRef]
- Reichl, F.-X.; Esters, M.; Simon, S.; Seiss, M.; Kehe, K.; Kleinsasser, N.; Folwaczny, M.; Glas, J.; Hickel, R. Cell death effects of resin-based dental material compounds and mercurials in human gingival fibroblasts. Arch. Toxicol. 2005, 80, 370–377. [Google Scholar] [CrossRef]
- Bakopoulou, A.; Papadopoulos, T.; Garefis, P. Molecular Toxicology of Substances Released from Resin–Based Dental Restorative Materials. Int. J. Mol. Sci. 2009, 10, 3861–3899. [Google Scholar] [CrossRef]
- Atsumi, T.; Ishihara, M.; Kadoma, Y.; Tonosaki, K.; Fujisawa, S. Comparative radical production and cytotoxicity induced by camphorquinone and 9-fluorenone against human pulp fibroblasts. J. Oral Rehabil. 2004, 31, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Di Nisio, C.; D’Aurora, M.; Di Giacomo, V.; Stuppia, L.; Cataldi, A.; Gatta, V. Transcriptome modifications in human gingival fibroblasts exposed to 2-hydroxyethyl methacrylate. Gene 2016, 582, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Nilsen, B.W.; Simon-Santamaria, J.; Örtengren, U.; Jensen, E.; Bruun, J.-A.; Michelsen, V.B.; Sørensen, K.K. Dose- and time-dependent effects of triethylene glycol dimethacrylate on the proteome of human THP-1 monocytes. Eur. J. Oral Sci. 2018, 126, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Fujioka-Kobayashi, M.; Miron, R.J.; Lussi, A.; Gruber, R.; Ilie, N.; Price, R.B.; Schmalz, G. Effect of the degree of conversion of resin-based composites on cytotoxicity, cell attachment, and gene expression. Dent. Mater. 2019, 35, 1173–1193. [Google Scholar] [CrossRef] [PubMed]
- Issa, Y.; Watts, D.C.; Brunton, P.A.; Waters, C.M.; Duxbury, A.J. Resin composite monomers alter MTT and LDH activity of human gingival fibroblasts in vitro. Dent. Mater. 2004, 20, 12–20. [Google Scholar] [CrossRef]
- Nomura, Y.; Teshima, W.; Kawahara, T.; Tanaka, N.; Ishibashi, H.; Okazaki, M.; Arizono, K. Genotoxicity of dental resin polymerization initiators in vitro. J. Mater. Sci. Mater. Med. 2006, 17, 29–32. [Google Scholar] [CrossRef]
- Randolph, L.D.; Steinhaus, J.; Möginger, B.; Gallez, B.; Stansbury, J.; Palin, W.M.; Leloup, G.; Leprince, J.G. Photopolymerization of highly filled dimethacrylate-based composites using Type I or Type II photoinitiators and varying co-monomer ratios. Dent. Mater. 2016, 32, 136–148. [Google Scholar] [CrossRef]
- D’Adamo, S.; Cetrullo, S.; Guidotti, S.; Borzì, R.; Flamigni, F. Hydroxytyrosol modulates the levels of microRNA-9 and its target sirtuin-1 thereby counteracting oxidative stress-induced chondrocyte death. Osteoarthr. Cartil. 2017, 25, 600–610. [Google Scholar] [CrossRef]
- Wang, H.; Wu, Q.; Zhang, Y.; Zhang, H.-N.; Wang, Y.-B.; Wang, W. TGF-β1-induced epithelial–mesenchymal transition in lung cancer cells involves upregulation of miR-9 and downregulation of its target, E-cadherin. Cell. Mol. Biol. Lett. 2017, 22, 1–10. [Google Scholar] [CrossRef]
- Detassis, S.; Grasso, M.; Del Vescovo, V.; Denti, M.A. microRNAs Make the Call in Cancer Personalized Medicine. Front. Cell Dev. Biol. 2017, 5, 86. [Google Scholar] [CrossRef]
- Qu, B.; Jia, Y.; Liu, Y.; Wang, H.; Ren, G.; Wang, H. The detection and role of heat shock protein 70 in various nondisease conditions and disease conditions: A literature review. Cell Stress Chaperon. 2015, 20, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Noda, M.; Wataha, J.C.; Kaga, M.; Lockwood, P.E.; Volkmann, K.R.; Sano, H. Components of dentinal adhesives modulate heat shock protein 72 expression in heat-stressed THP-1 human monocytes at sublethal concentrations. J. Dent. Res. 2002, 81, 265–269. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Resin | Chemical Constituents |
|---|---|
| C | Bisphenol-A-glycidyl methacrylate (BisGMA); Triethylene glycol dimethacrylate (TEGDMA); Camphorquinone (CQ) |
| E | Bisphenol-A-glycidyl methacrylate (BisGMA); Triethylene glycol dimethacrylate (TEGDMA) |
| F | Urethane dimethacrylate (UDMA); Bisphenol-A- polyethethylene glycol diether dimethacrylate (BisEMA) |
| Biochemical Indices of Resin Toxicity | Control | C | E | F | Significance |
|---|---|---|---|---|---|
| MTT test (% of control) | |||||
| Freshly-cured resin | 100 ± 7 | 42 ± 9 ** | 78 ± 11 **## | 69 ± 10 **## | Toxicity C >> E = F |
| Preincubated resin | 73 ± 8 **++ | 81 ± 9 **## | 54 ± 11 **##^^ | F > E > E | |
| LDH (% of total activity) | |||||
| 1 h | 10 ± 3 * | 4 ± 2 | 5 ± 3 * | C—Rapid increase, saturable | |
| 3 h | 17 ± 6 ** | 7 ± 3 ** | 9 ± 4 ** | E—Slow increase, saturable | |
| 6 h | 28 ± 5 ** | 11 ± 4 ** | 13 ± 5 ** | F—Slow increase, linear | |
| 12 h | 37 ± 7 ** | 22 ± 6 ** | 19 ± 8 ** | ||
| 24 h | 43 ± 9 ** | 28 ± 8 ** | 37 ± 8 ** | ||
| Cell cycle (%) | |||||
| Cells with damaged DNA | 8 ± 3 | 39 ± 11 ** | 18 ± 5 **## | 24 ± 7 **##^ | Increased C > F > E |
| Pre-DNA synthesis/ resting | 55 ± 9 | 34 ± 7 ** | 67 ± 8 ## | 52 ± 9 ##^ | Variable E > C = F |
| Proliferation | 37 ± 7 | 27 ± 8 ** | 15 ± 4 **## | 24 ± 6 **^^ | Decreased E > F > C |
| Oxidative stress (relative units) | 100 ± 34 | 951 ± 111 ** | 877 ± 132 ** | 1215 ± 201 **##^^ | F > E = C |
| Apoptosis/Necrosis (%) | |||||
| Naive cells | 93 ± 7 | 18 ± 6 ** | 21 ± 11 ** | 20 ± 5 ** | C-predominantly necrosis |
| Necrotic cells | 2 ± 3 | 75 ± 10 ** | 48 ± 14 **# | 73 ± 11 ** | |
| Apoptotic cells | 2 ± 2 | 2 ± 2 | 3 ± 2 | 1 ± 2 | D - some apoptosis |
| Necrotic/apoptotic cells | 3 ± 2 | 5 ± 4 | 28 ± 13 **# | 16 ± 7 **##^^ | F-mostly necrosis |
| miR-9 (fold change) | 1 | 1.76 * | 1.92 * | 3.39 * | Increased F > C = E |
| HSP70 (relative units) | 100 ± 17 | 733 ± 76 ** | 112 ± 23 ## | 177 ± 28 ## | Increased C >> (E = F normal) |
| DCF/HSP70 (relative units) | |||||
| Scatterplot area | 100 ± 14 | 167 ± 26 ** | 127 ± 17 | 131 ± 19 * | Increased C > E = F |
| Central tendency line (slope act) | 1.04 ± 0.12 | 1.15 ± 0.11 | 0.9 ± 0.14 | 1.00 ± 0.15 | |
| Trend (vector) line (slope at) | - | 0.05 ± 0.14 κκ | 1.19 ± 0.18 κ− | 1.11 ± 0.25 | C ≠ E = F |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sulek, J.; Luczaj-Cepowicz, E.; Marczuk-Kolada, G.; Rosłan, M.; Holownia, A. Cytotoxicity of Methacrylate Dental Resins to Human Gingival Fibroblasts. J. Funct. Biomater. 2022, 13, 56. https://doi.org/10.3390/jfb13020056
Sulek J, Luczaj-Cepowicz E, Marczuk-Kolada G, Rosłan M, Holownia A. Cytotoxicity of Methacrylate Dental Resins to Human Gingival Fibroblasts. Journal of Functional Biomaterials. 2022; 13(2):56. https://doi.org/10.3390/jfb13020056
Chicago/Turabian StyleSulek, Jolanta, Elzbieta Luczaj-Cepowicz, Grazyna Marczuk-Kolada, Maciej Rosłan, and Adam Holownia. 2022. "Cytotoxicity of Methacrylate Dental Resins to Human Gingival Fibroblasts" Journal of Functional Biomaterials 13, no. 2: 56. https://doi.org/10.3390/jfb13020056
APA StyleSulek, J., Luczaj-Cepowicz, E., Marczuk-Kolada, G., Rosłan, M., & Holownia, A. (2022). Cytotoxicity of Methacrylate Dental Resins to Human Gingival Fibroblasts. Journal of Functional Biomaterials, 13(2), 56. https://doi.org/10.3390/jfb13020056