Mapping the Memory Structure of High-Knowledge Students: A Longitudinal Semantic Network Analysis


Abstract
1. Introduction
The Present Research
2. Study 1
2.1. Materials and Methods
2.1.1. Participants
2.1.2. Materials
2.1.3. Group Construction
2.1.4. Semantic Memory Network Estimation
2.1.5. Procedure
2.2. Results
2.2.1. Fluency and Descriptives
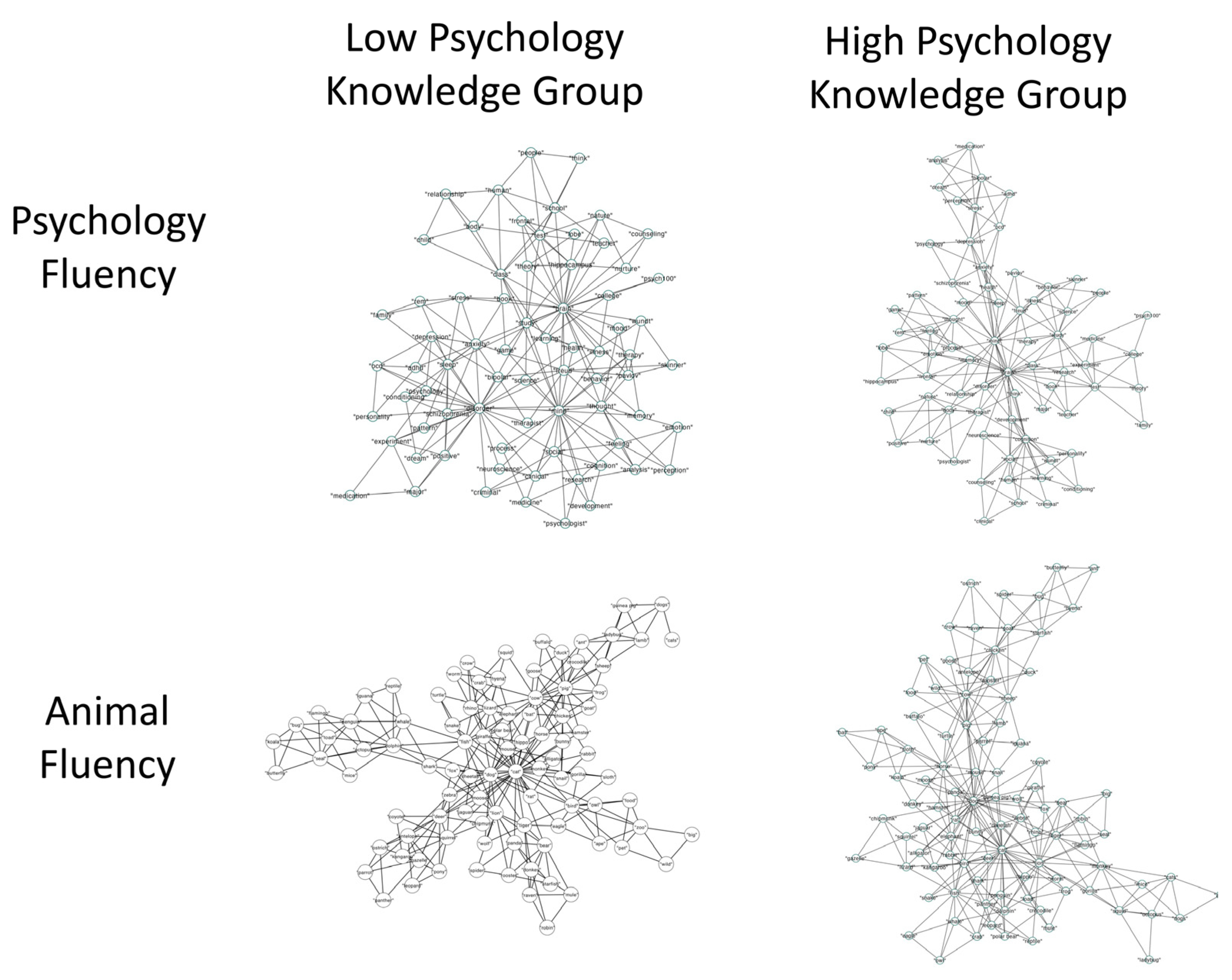
2.2.2. Semantic Memory Networks
2.3. Discussion
3. Study 2
3.1. Materials and Methods
3.1.1. Participants
3.1.2. Materials
3.1.3. Group Construction
3.1.4. Semantic Memory Network Estimation
3.1.5. Procedure
3.2. Results
3.2.1. Fluency and Descriptives
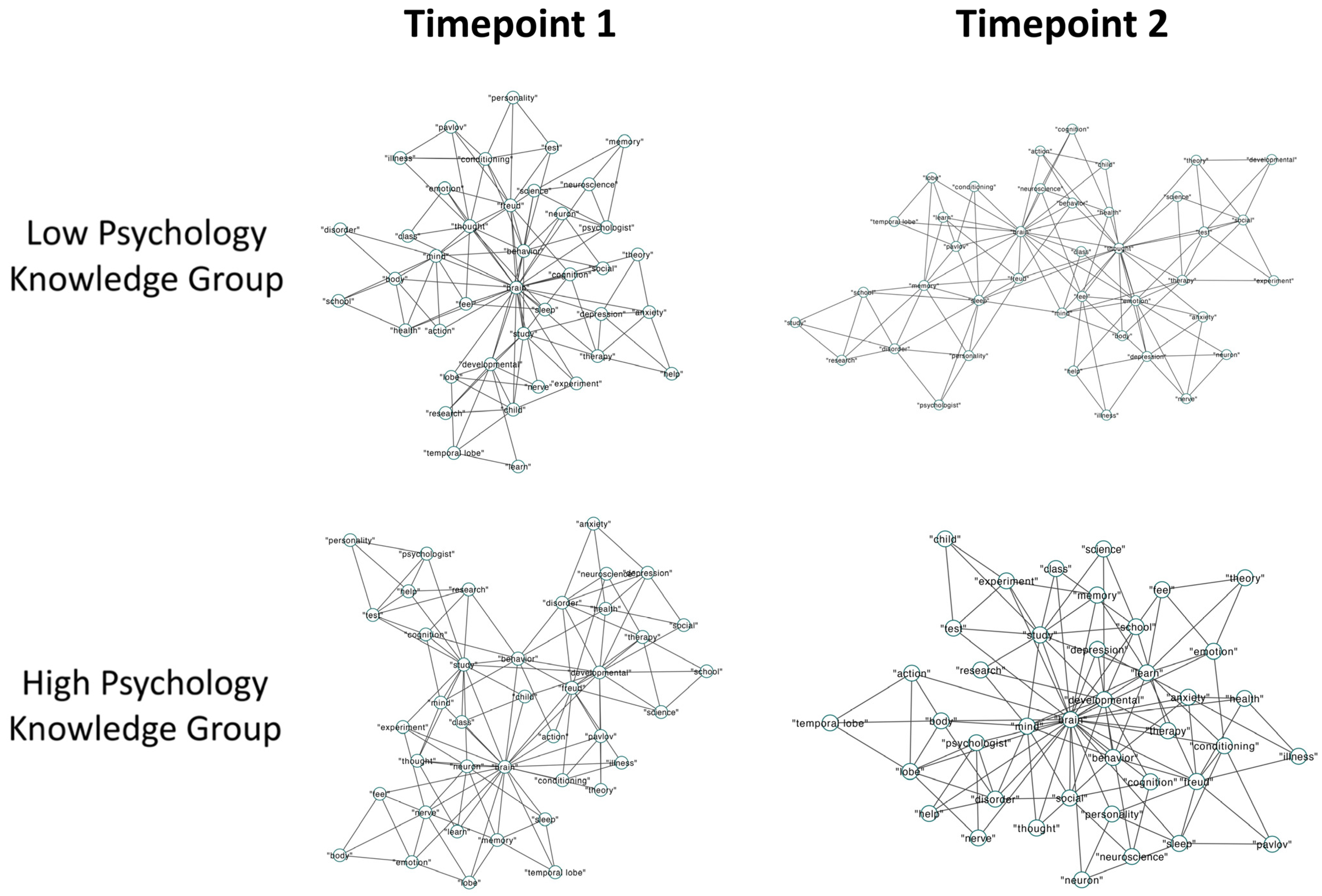
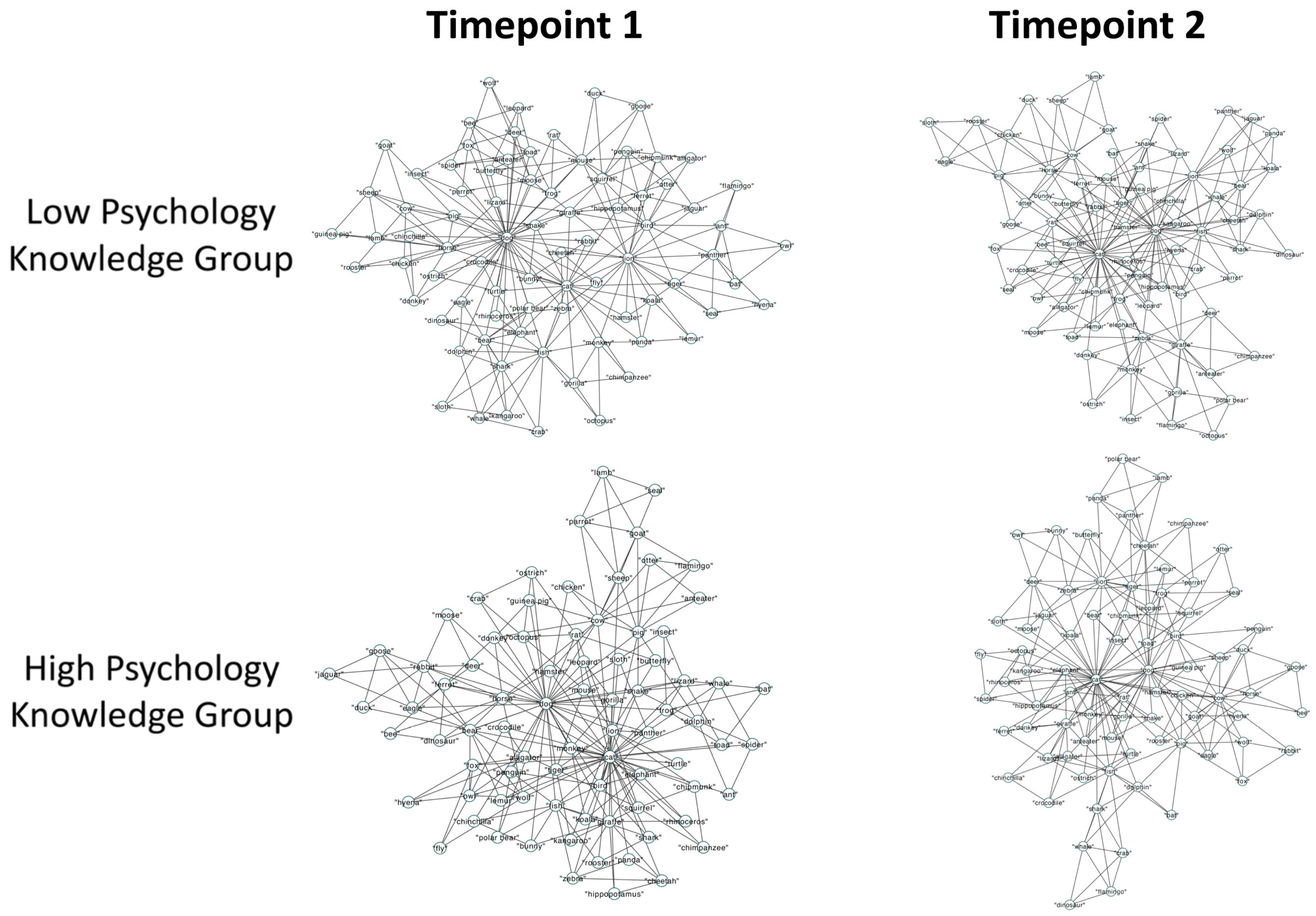
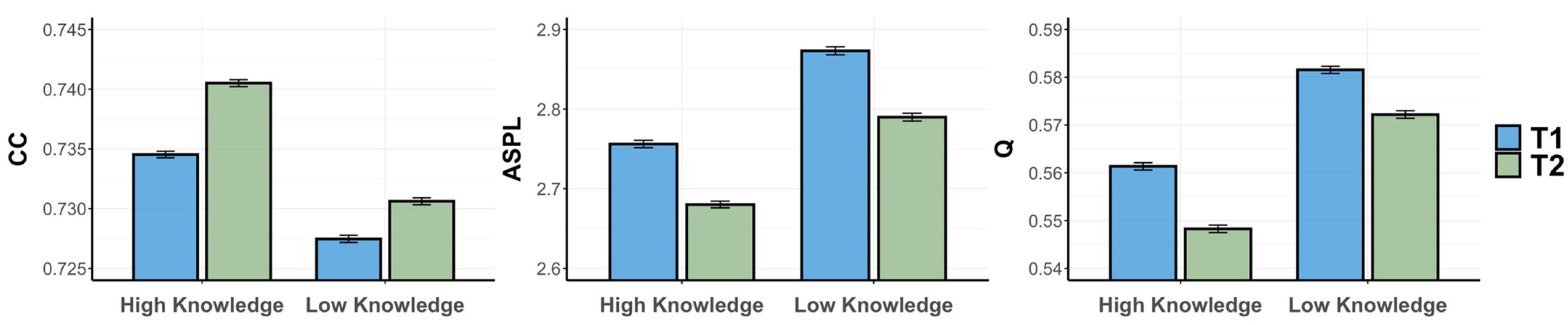
3.2.2. Semantic Memory Networks
3.3. Discussion
4. General Discussion
Limitations and Future Directions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
Intro Psych Test
- The cerebellum is primarily involved in directing
- The frontal lobes primary role is in supporting
- The hippocampus is a part of the
- Neuroplasticity refers to how the nervous system can
- The function of neurotransmitters in the nervous system is that of
- The process of inputting information into the memory system is called
- The memory store for personal life events is
- Recognition specifically refers to the ability of
- Retrieval specifically refers to the ability of
- Semantic memory primarily stores information about
- Altruism is a form of prosocial behavior that is motivated by
- Conformity refers to the
- Empathy refers to the ability to
- If group members modify their opinions to align with a perceived group consensus, this is an example of
- A set of group expectations for appropriate thoughts and behaviors of its members is called
- The emotional bond between an infant and parent that affects the infant’s sense of security is
- Cognitive development primarily concerns the strengthening of
- The idea that even if something is out of sight, it still exists is called
- An example of the sensitive period is the
- Temperament is thought of as
- Binocular vision requires
- A blind spot is understood to be
- An example of a gestalt principle is the
- Inattentional blindness is thought of as
- Perceptual constancy of shapes, brightness and size refers to the
- Associative learning occurs when an individual
- When a stimulus or experience occurs before a behavior that it gets paired with what occurs is
- Observational learning is thought to largely derive from
- Operant conditioning is a form of learning where
- Taking away a pleasant stimulus to stop a behavior is an example of
- The ability to self-monitor in social situations will especially depend on the
- A lesion to the hippocampus would render an individual entirely unable to
- Associative learning mostly relies on
- What is likely to play the largest role in determining recognition performance on a cognitive task?
- What kind of memory is impacted by infantile amnesia?
- Social norms will typically be stored in
- The development of secure attachment will crucially depend on the caregiver demonstrating high levels of
References
- Anderson, John Robert. 2000. Learning and Memory: An Integrated Approach. Hoboken: John Wiley & Sons Inc. [Google Scholar]
- Arbesman, Samuel, Steven H. Strogatz, and Michael S. Vitevitch. 2010. The structure of phonological networks across multiple languages. International Journal of Bifurcation and Chaos 20: 679–85. [Google Scholar] [CrossRef]
- Ardila, Alfredo, Feggy Ostrosky-Solís, and Byron Bernal. 2006. Cognitive testing toward the future: The example of Semantic Verbal Fluency (ANIMALS). International Journal of Psychology 41: 324–32. [Google Scholar] [CrossRef]
- Baronchelli, Andrea, Ramon Ferrer-i-Cancho, Romualdo Pastor-Satorras, Nick Chater, and Morten H. Christiansen. 2013. Networks in cognitive science. Trends in Cognitive Sciences 17: 348–60. [Google Scholar] [CrossRef]
- Beaty, Roger E., and Dan R. Johnson. 2021. Automating creativity assessment with SemDis: An open platform for computing semantic distance. Behavior Research Methods 53: 757–80. [Google Scholar] [CrossRef] [PubMed]
- Becker, William E., and Michael Watts. 2001. Teaching methods in U.S. undergraduate economics courses. Journal of Economic Education 32: 269–79. [Google Scholar] [CrossRef]
- Benedek, Mathias, Yoed N. Kenett, Konstantin Umdasch, David Anaki, Miriam Faust, and Aljoscha C. Neubauer. 2017. How semantic memory structure and intelligence contribute to creative thought: A network science approach. Thinking & Reasoning 23: 158–83. [Google Scholar]
- Bertail, Patrice. 1997. Second-order properties of an extrapolated bootstrap without replacement under weak assumptions. Bernoulli 3: 149–79. [Google Scholar] [CrossRef]
- Biggs, John B. 1973. Study behaviour and performance in objective and essay formats. Australian Journal of Education 17: 157–67. [Google Scholar] [CrossRef]
- Börner, Katy, Soma Sanyal, and Alessandro Vespignani. 2008. Network Science. Annual Review of Information Science and Technology 41: 537–607. [Google Scholar] [CrossRef]
- Chen, Bodong, and Oleksandra Poquet. 2022. Networks in learning analytics: Where theory, methodology, and practice intersect. Journal of Learning Analytics 9: 1–12. [Google Scholar] [CrossRef]
- Chi, Michelene T. H., Paul J. Feltovich, and Robert Glaser. 1981. Categorization and representation of physics problems by experts and novices. Cognitive Science 5: 121–52. [Google Scholar] [CrossRef]
- Christensen, Alexander P., and Yoed N. Kenett. 2023. Semantic network analysis (SemNA): A tutorial on preprocessing, estimating, and analyzing semantic networks. Psychological Methods 28: 860–79. [Google Scholar] [CrossRef] [PubMed]
- Collins, Allan M., and Elizabeth F. Loftus. 1975. A spreading-activation theory of semantic processing. Psychological Review 82: 407. [Google Scholar] [CrossRef]
- Conway, Andrew R. A., Michael J. Kane, Michael F. Bunting, D. Zach Hambrick, Oliver Wilhelm, and Randall W. Engle. 2005. Working memory span tasks: A methodological review and user’s guide. Psychonomic Bulletin & Review 12: 769–86. [Google Scholar]
- Cosgrove, Abigail L., Roger E. Beaty, Michele T. Diaz, and Yoed N. Kenett. 2023. Age differences in semantic network structure: Acquiring knowledge shapes semantic memory. Psychology and Aging 38: 87–102. [Google Scholar] [CrossRef] [PubMed]
- Deary, Ian J., Steve Strand, Pauline Smith, and Cres Fernandes. 2007. Intelligence and educational achievement. Intelligence 35: 13–21. [Google Scholar] [CrossRef]
- Denervaud, Solange, Alexander P. Christensen, Yoed N. Kenett, and Roger E. Beaty. 2021. Education shapes the structure of semantic memory and impacts creative thinking. npj Science of Learning 6: 1–7. [Google Scholar] [CrossRef] [PubMed]
- DiSessa, Andrea A., and Bruce L. Sherin. 1998. What changes in conceptual change? International Journal of Science Education 20: 1155–91. [Google Scholar] [CrossRef]
- Efron, Bradley. 1979. Bootstrap method: Another look at the jackknife. The Annals of Statistics 7: 1–26. [Google Scholar] [CrossRef]
- Entwistle, Abigail, and Noel Entwistle. 1992. Experiences of understanding in revising for degree examinations. Learning and Instruction 2: 1–22. [Google Scholar] [CrossRef]
- Fortunato, Santo. 2010. Community detection in graphs. Physics Reports 486: 75–174. [Google Scholar] [CrossRef]
- He, Li, Yoed N. Kenett, Kaixiang Zhuang, Cheng Liu, Rongcan Zeng, Tingrui Yan, Tengbin Huo, and Jiang Qiu. 2021. The relation between semantic memory structure, associative abilities, and verbal and figural creativity. Thinking & Reasoning 27: 268–93. [Google Scholar]
- Irwin, Julie R., and Gary H. McClelland. 2003. Negative consequences of dichotomizing continuous predictor variables. Journal of Marketing Research 40: 366–71. [Google Scholar] [CrossRef]
- Kenett, Yoed N. 2024. The Role of Knowledge in Creative Thinking. Creativity Research Journal, 1–8. [Google Scholar] [CrossRef]
- Kenett, Yoed N., Effi Levi, David Anaki, and Miriam Faust. 2017. The semantic distance task: Quantifying semantic distance with semantic network path length. Journal of Experimental Psychology: Learning, Memory, & Cognition 43: 1470–89. [Google Scholar]
- Kenett, Yoed N., Orr Levy, Dror Y. Kenett, H. Eugene Stanley, Miriam Faust, and Shlomo Havlin. 2018. Flexibility of thought in high creative individuals represented by percolation analysis. Proceedings of the National Academy of Sciences 115: 867–72. [Google Scholar] [CrossRef] [PubMed]
- Kinchin, Ian M., David B. Hay, and Alan Adams. 2000. How a qualitative approach to concept map analysis can be used to aid learning by illustrating patterns of conceptual development. Educational Research 42: 43–57. [Google Scholar] [CrossRef]
- Koponen, Ismo T., and Maija Nousiainen. 2014. Concept networks in learning: Finding key concepts in learners’ representations of the interlinked structure of scientific knowledge. Journal of Complex Networks 2: 187–202. [Google Scholar] [CrossRef]
- Koponen, Ismo T., and Maija Pehkonen. 2010. Coherent knowledge structures of physics represented as concept networks in teacher education. Science & Education 19: 259–82. [Google Scholar]
- Kumar, Abhilasha A., David A. Balota, and Mark Steyvers. 2020. Distant connectivity and multiple-step priming in large-scale semantic networks. Journal of Experimental Psychology: Learning, Memory, and Cognition 46: 2261. [Google Scholar] [CrossRef]
- Kunina, Olga, Oliver Wilhelm, Maren Formazin, Kathrin Jonkmann, and Ulrich Schroeders. 2007. Extended criteria and predictors in college admission: Exploring the structure of study success and investigating the validity of domain knowledge. Psychology Science 49: 88–114. [Google Scholar]
- Landauer, Thomas K., and Susan T. Dumais. 1997. A solution to Plato’s problem: The latent semantic analysis theory of acquisition, induction, and representation of knowledge. Psychological Review 104: 211–40. [Google Scholar] [CrossRef]
- Lavigne, Nancy C. 2005. Mutually informative measures of knowledge: Concept maps plus problem sorts in statistics. Educational Assessment 10: 39–71. [Google Scholar] [CrossRef]
- Li, Yangping, Roger E. Beaty, Simone Luchini, Weiping Hu, and Yoed N. Kenett. 2024. The role of semantic memory networks in crystallized intelligence and creative thinking ability. Learning and Individual Differences 111: 102426. [Google Scholar] [CrossRef]
- Luchini, Simone A., Yoed N. Kenett, Daniel C. Zeitlen, Alexander P. Christensen, Derek M. Ellis, Gene A. Brewer, and Roger E. Beaty. 2023. Convergent thinking and insight problem solving relate to semantic memory network structure. Thinking Skills and Creativity 48: 101277. [Google Scholar] [CrossRef]
- MacCallum, Robert C., Shaobo Zhang, Kristopher J. Preacher, and Derek D. Rucker. 2002. On the practice of dichotomization of quantitative variables. Psychological Methods 7: 19–40. [Google Scholar] [CrossRef]
- Maker, C. June, and Robert H. Zimmerman. 2020. Concept maps as assessments of expertise: Understanding of the complexity and interrelationships of concepts in science. Journal of Advanced Academics 31: 254–97. [Google Scholar] [CrossRef]
- Martinez, Michael E. 1999. Cognition and the question of test item format. Educational Psychologist 34: 207–18. [Google Scholar] [CrossRef]
- Massara, Guido Previde, Tiziana Di Matteo, and Tomaso Aste. 2016. Network filtering for big data: Triangulated maximally filtered graph. Journal of Complex Networks 5: 161–78. [Google Scholar] [CrossRef]
- Morais, Ana Sofia, Henrik Olsson, and Lael J. Schooler. 2013. Mapping the structure of semantic memory. Cognitive Science 37: 125–45. [Google Scholar] [CrossRef]
- Nesbit, John C., and Olusola O. Adesope. 2006. Learning with concept and knowledge maps: A meta-analysis. Review of Educational Research 76: 413–48. [Google Scholar] [CrossRef]
- Newman, Mark, Albert-László Barabási, and Duncan J. Watts. 2006. The Structure and Dynamics of Networks. Princeton: Princeton University Press. [Google Scholar]
- Novak, Joseph D. 2010. Learning, Creating, and Using Knowledge: Concept Maps as Facilitative Tools in Schools and Corporations. New York: Routledge. [Google Scholar]
- Novak, Joseph D., and Alberto J. Cañas. 2007. Theoretical origins of concept maps, how to construct them, and uses in education. Reflecting Education 3: 29–42. [Google Scholar]
- Preacher, Kristopher J., Derek D. Rucker, Robert C. MacCallum, and W. Alan Nicewander. 2005. Use of the extreme groups approach: A critical reexamination and new recommendations. Psychological Methods 10: 178. [Google Scholar] [CrossRef] [PubMed]
- Rastelli, Clara, Antonino Greco, and Chiara Finocchiaro. 2020. Revealing the role of divergent thinking and fluid intelligence in children’s semantic memory organization. Journal of Intelligence 8: 43. [Google Scholar] [CrossRef] [PubMed]
- Rittle-Johnson, Bethany, and Michael Schneider. 2015. Developing Conceptual and Procedural Knowledge of Mathematics. In Oxford Handbook of Numerical Cognition. Oxford: Oxford University Press. [Google Scholar]
- Ruiz-Primo, Maria Araceli, and Richard J. Shavelson. 1996. Problems and issues in the use of concept maps in science assessment. Journal of Research in Science Teaching: The Official Journal of the National Association for Research in Science Teaching 33: 569–600. [Google Scholar] [CrossRef]
- Shannon, Paul, Andrew Markiel, Owen Ozier, Nitin S. Baliga, Jonathan T. Wang, Daniel Ramage, Nada Amin, Benno Schwikowski, and Trey Ideker. 2003. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Research 13: 2498–504. [Google Scholar] [CrossRef] [PubMed]
- Siew, Cynthia S. Q. 2019. Using network science to analyze concept maps of psychology undergraduates. Applied Cognitive Psychology 33: 662–68. [Google Scholar] [CrossRef]
- Siew, Cynthia S. Q. 2020. Applications of network science to education research: Quantifying knowledge and the development of expertise through network analysis. Education Sciences 10: 101. [Google Scholar] [CrossRef]
- Siew, Cynthia S. Q., and Anutra Guru. 2022. Investigating the network structure of domain-specific knowledge using the semantic fluency task. Memory & Cognition 51: 623–46. [Google Scholar]
- Siew, Cynthia S. Q., Dirk U. Wulff, Nicole M. Beckage, and Yoed N. Kenett. 2019. Cognitive network science: A review of research on cognition through the lens of network representations, processes, and dynamics. Complexity 2019: 2108423. [Google Scholar] [CrossRef]
- Simkin, Mark G., and William L. Kuechler. 2005. Multiple-choice tests and student understanding: What is the connection? Decision Sciences Journal of Innovative Education 3: 73–98. [Google Scholar] [CrossRef]
- Smith, Edward E., Edward J. Shoben, and Lance J. Rips. 1974. Structure and process in semantic memory: A featural model for semantic decisions. Psychological Review 81: 214. [Google Scholar] [CrossRef]
- Soares, Diana Lopes, Gina C. Lemos, Ricardo Primi, and Leandro S. Almeida. 2015. The relationship between intelligence and academic achievement throughout middle school: The role of students’ prior academic performance. Learning and Individual Differences 41: 73–78. [Google Scholar] [CrossRef]
- Steyvers, Mark, and Joshua B. Tenenbaum. 2005. The large-scale structure of semantic networks: Statistical analyses and a model of semantic growth. Cognitive Science 29: 41–78. [Google Scholar] [CrossRef] [PubMed]
- Stiggins, Richard J. 1991. Facing challenges of a new era of educational assessment. Applied Measurement in Education 4: 263–73. [Google Scholar] [CrossRef]
- Suskie, Linda. 2018. Assessing Student Learning: A common Sense Guide. San Francisco: John Wiley & Sons. [Google Scholar]
- Van Wijk, Bernadette C. M., Cornelis J. Stam, and Andreas Daffertshofer. 2010. Comparing brain networks of different size and connectivity density using graph theory. PLoS ONE 5: e13701. [Google Scholar] [CrossRef]
- Vitevitch, Michael S., Kit Ying Chan, and Steven Roodenrys. 2012. Complex network structure influences processing in long-term and short-term memory. Journal of Memory and Language 67: 30–44. [Google Scholar] [CrossRef]
- Wagenmakers, Eric-Jan. 2007. A practical solution to the pervasive problems of p values. Psychonomic Bulletin & Review 14: 779–804. [Google Scholar]
- Watts, Duncan J., and Steven H. Strogatz. 1998. Collective dynamics of ‘small-world’networks. Nature 393: 440–42. [Google Scholar] [CrossRef]
- Wetzels, Ruud, Dora Matzke, Michael D. Lee, Jeffrey N. Rouder, Geoffrey J. Iverson, and Eric-Jan Wagenmakers. 2011. Statistical evidence in experimental psychology: An empirical comparison using 855 t tests. Perspectives on Psychological Science 6: 291–98. [Google Scholar] [CrossRef]
- Wulff, Dirk U., Simon De Deyne, Samuel Aeschbach, and Rui Mata. 2022. Using network science to understand the aging lexicon: Linking individuals’ experience, semantic networks, and cognitive performance. Topics in Cognitive Science 14: 93–110. [Google Scholar] [CrossRef] [PubMed]
- Zemla, Jeffrey C., and Joseph L. Austerweil. 2018. Estimating semantic networks of groups and individuals from fluency data. Computational Brain Behavior 1: 36–58. [Google Scholar] [CrossRef] [PubMed]
- Zemla, Jeffrey C., Kesong Cao, Kimberly D. Mueller, and Joseph L. Austerweil. 2020. SNAFU: The semantic network and fluency utility. Behavior Research Methods 52: 1681–99. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Psychology Fluency Task | Animal Fluency Task | |||||||
|---|---|---|---|---|---|---|---|---|
| N (Average) | N (within) | N (between) | N (Average) | N (within) | N (between) | |||
| Group | M (SD) | Range | M (SD) | Range | ||||
| Low knowledge | 9.7 (4.1) | 3–22 | 307 | 156 | 16.4 (5.6) | 3–29 | 362 | 177 |
| High knowledge | 10.2 (3.9) | 3–24 | 359 | 208 | 17.4 (4.6) | 3–31 | 354 | 185 |
| M | SD | NA | Min, Max | 1 | 2 | 3 | 4 | |
|---|---|---|---|---|---|---|---|---|
| Intro Psych Test | 17.99 | 5.9 | 0 | 5, 32 | 1 | |||
| Self-Reported Grades | 6.86 | 2.53 | 12 | 1, 10 | 0.30 | 1 | ||
| Psychology Fluency | 9.98 | 3.98 | 0 | 3, 24 | 0.06 | 0.15 | 1 | |
| Animal Fluency | 16.98 | 5.05 | 0 | 3, 31 | 0.13 | 0.14 | 0.43 | 1 |
| M | SD | NA | Min, Max | 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Intro Psych Test T1 | 19.32 | 4.54 | 0 | 9, 30 | |||||||
| Intro Psych Test T2 | 22.26 | 5.02 | 0 | 11, 34 | 0.61 | ||||||
| PsyKT | 18.9 | 4.47 | 0 | 9, 29 | 0.4 | 0.55 | |||||
| Self-Reported Grades | 8.71 | 1.73 | 5 | 2, 10 | 0.27 | 0.37 | 0.21 | ||||
| Psychology Fluency T1 | 12.14 | 3.83 | 0 | 4, 26 | 0.05 | 0.02 | 0.006 | 0.07 | |||
| Psychology Fluency T2 | 13.45 | 4.45 | 2 | 3, 29 | −0.005 | 0.07 | 0.04 | 0.06 | 0.56 | ||
| Animal Fluency T1 | 18.89 | 4.32 | 0 | 6, 31 | 0.18 | 0.24 | 0.1 | 0.19 | 0.43 | 0.23 | |
| Animal Fluency T2 | 19.16 | 4.51 | 1 | 6, 29 | 0.15 | 0.17 | 0.05 | 15 | 0.47 | 0.49 | 0.54 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luchini, S.A.; Wang, S.; Kenett, Y.N.; Beaty, R.E. Mapping the Memory Structure of High-Knowledge Students: A Longitudinal Semantic Network Analysis. J. Intell. 2024, 12, 56. https://doi.org/10.3390/jintelligence12060056
Luchini SA, Wang S, Kenett YN, Beaty RE. Mapping the Memory Structure of High-Knowledge Students: A Longitudinal Semantic Network Analysis. Journal of Intelligence. 2024; 12(6):56. https://doi.org/10.3390/jintelligence12060056
Chicago/Turabian StyleLuchini, Simone A., Shuyao Wang, Yoed N. Kenett, and Roger E. Beaty. 2024. "Mapping the Memory Structure of High-Knowledge Students: A Longitudinal Semantic Network Analysis" Journal of Intelligence 12, no. 6: 56. https://doi.org/10.3390/jintelligence12060056
APA StyleLuchini, S. A., Wang, S., Kenett, Y. N., & Beaty, R. E. (2024). Mapping the Memory Structure of High-Knowledge Students: A Longitudinal Semantic Network Analysis. Journal of Intelligence, 12(6), 56. https://doi.org/10.3390/jintelligence12060056