
Dynamics of an Intraguild Predation Food Web Cooperation Model Under the Influence of Fear and Hunting



Abstract
1. Introduction
2. Materials and Methods
3. Model Analysis
3.1. The Existence of Equilibrium Points
- The trivial equilibria and always exist.
- The fourth point is , where
- and is a solution for the polynomial equation
- The fifth point is , where
- and is a positive root of the following seventh-order equation:
3.2. Stability Analysis
4. Global Stability
5. Bifurcation Analysis
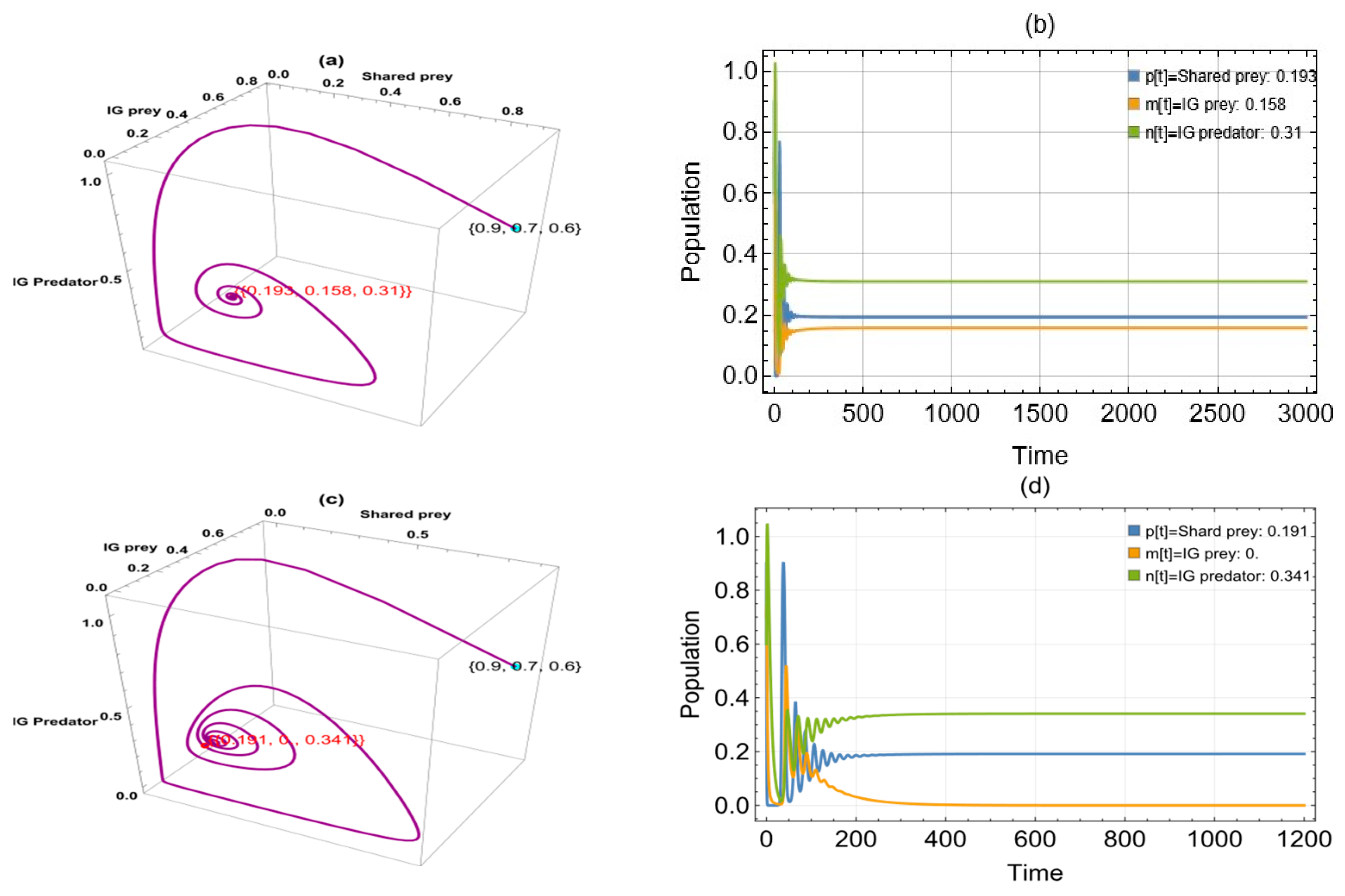
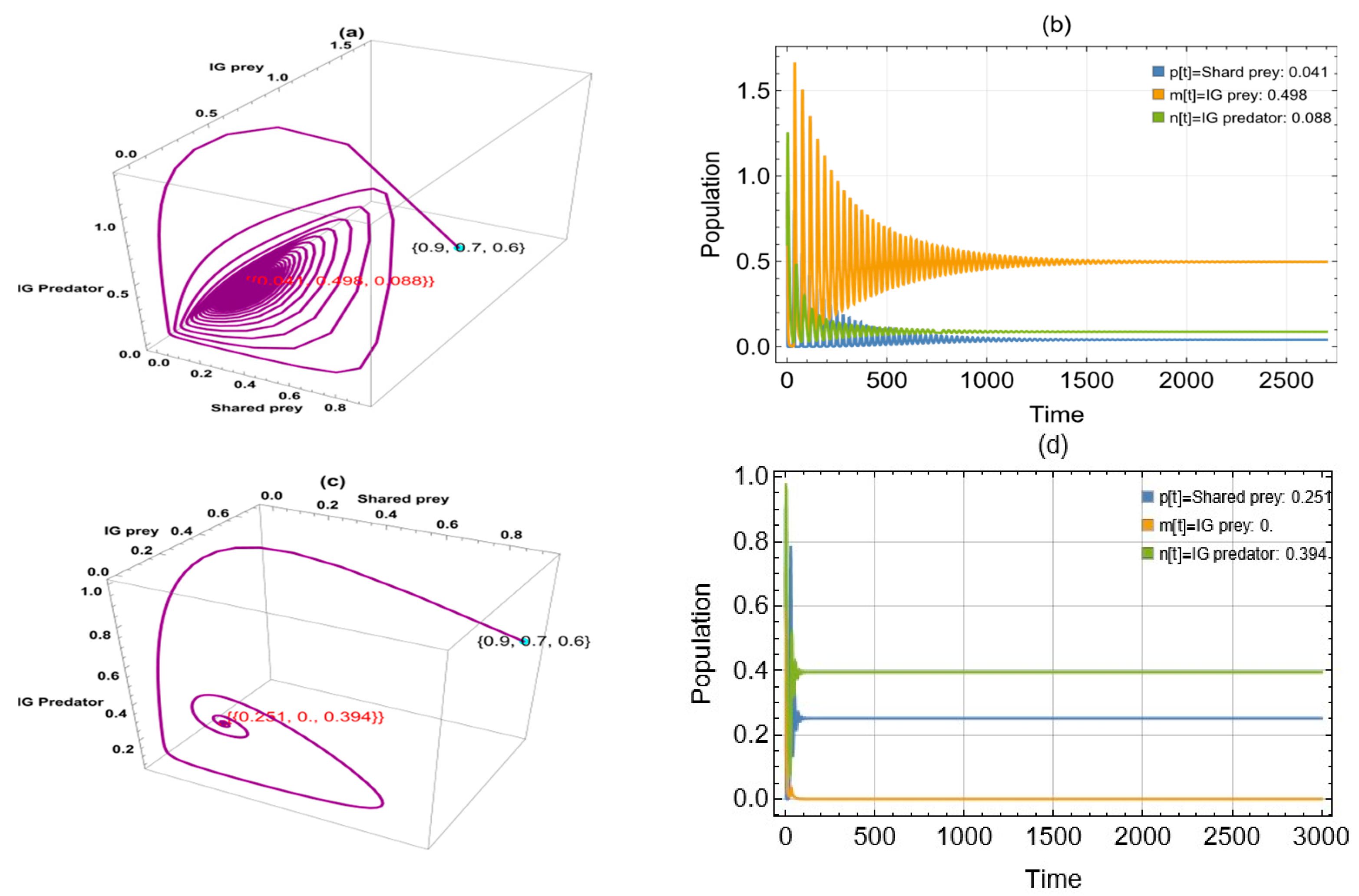
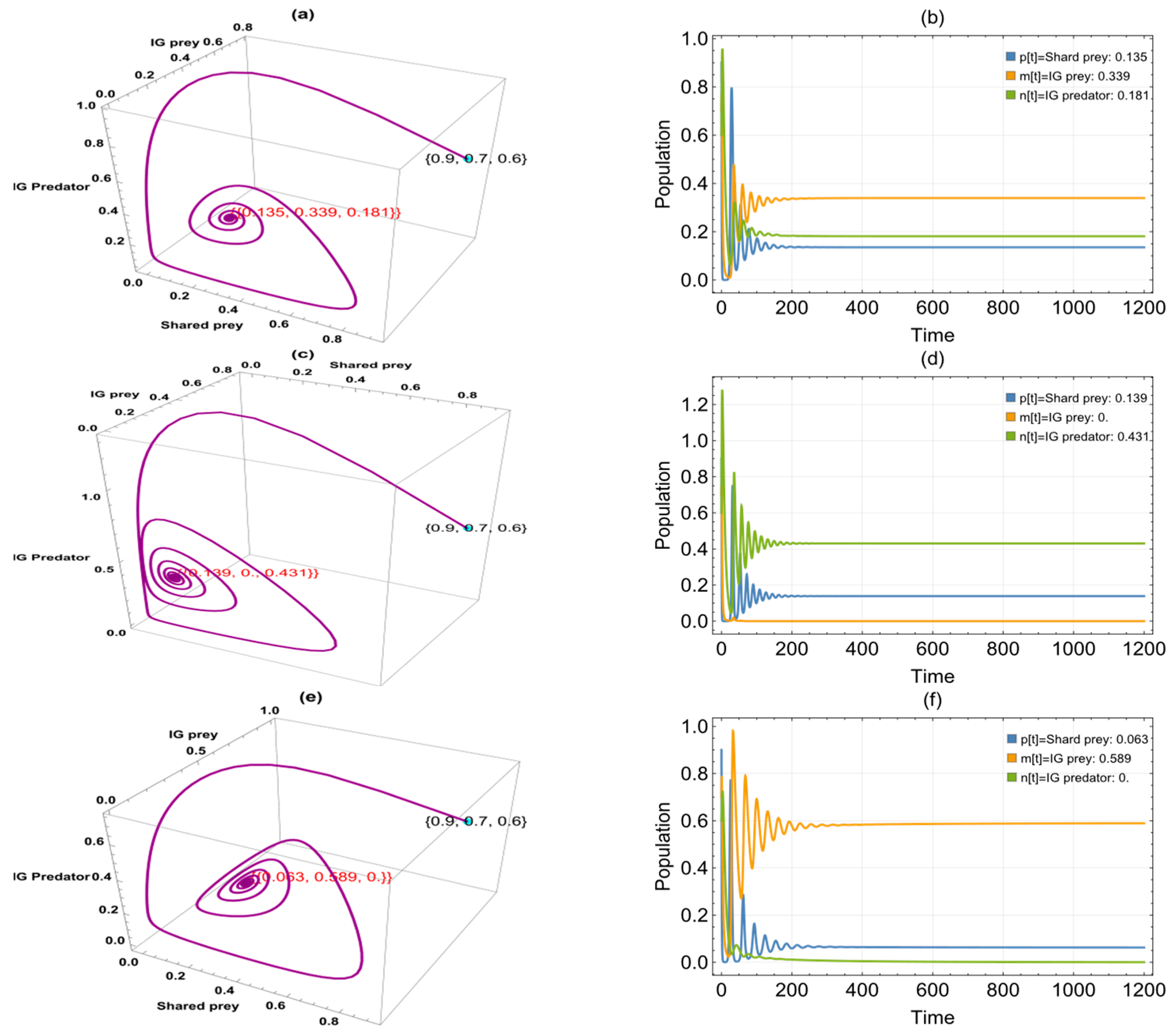
6. Numerical Simulation
7. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morin, P. Productivity, Intraguild predation, and population dynamics in experimental food webs. Ecology 1999, 80, 752–760. [Google Scholar] [CrossRef]
- Nakazawa, T.; Miki, T.; Namba, T. Influence of predator-specific defense adaptation on intraguild predation. Oikos 2010, 119, 418–427. [Google Scholar] [CrossRef]
- Sen, D.; Ghorai, S.; Banerjee, M. Complex dynamics of a three species prey-predator model with intraguild predation. Ecol. Complex. 2018, 34, 9–22. [Google Scholar] [CrossRef]
- Amarasekare, P. Spatial dynamics of communities with intraguild predation: The role of dispersal strategies. Am. Nat. 2007, 170, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Sk, N.; Mondal, B.; Sarkar, A.; Santra, S.S.; Baleanu, D.; Altanji, M. Chaos emergence and dissipation in a three-species food web model with intraguild predation and cooperative hunting. AIMS Math. 2024, 9, 1023–1045. [Google Scholar] [CrossRef]
- Pal, S.; Pal, N.; Samanta, S.; Chattopadhyay, J. Effect of hunting cooperation and fear in a predator-prey model. Ecol. Complex. 2019, 39, 100770. [Google Scholar] [CrossRef]
- AL-Momen, S.; Naji, R.K. Effect of hunting cooperation and fear in a food chain model with intraspecific competition. Commun. Math. Biol. Neurosci. 2023, 2023, 119. [Google Scholar] [CrossRef]
- Meng, X.Y.; Feng, Y. Dynamical behaviour of an intraguild predator–prey model with prey refuge and hunting cooperation. J. Biol. Dyn. 2023, 17, 2222142. [Google Scholar] [CrossRef]
- Mendonça, J.P.; Gleria, I.; Lyra, M.L. Prey refuge and morphological defense mechanisms as nonlinear triggers in an intraguild predation food web. Commun. Nonlinear Sci. Numer. Simul. 2020, 90, 105373. [Google Scholar] [CrossRef]
- Hadi, Z.M.; Bahlool, D.K. The effect of alternative resource and refuge on the dynamical behavior of food chain model. Malays. J. Math. Sci. 2023, 17, 731–754. [Google Scholar] [CrossRef]
- Bahlool, D.K. The dynamics of a stage-structure prey-predator model with hunting cooperation and anti-predator behavior. Commun. Math. Biol. Neurosci. 2023, 2023, 59. [Google Scholar] [CrossRef]
- Xing, Z.; Cui, H.; Zhang, J. Dynamics of a stochastic intraguild predation model. Appl. Sci. 2016, 6, 118. [Google Scholar] [CrossRef]
- Liu, R.; Liu, G. Complex dynamics and optimal harvesting for a stochastic food-web model with intraguild predation and time delays. Int. J. Biomath. 2022, 15, 2250050. [Google Scholar] [CrossRef]
- Liu, R.; Liu, G. Analysis on stochastic food-web model with intraguild predation and mixed functional responses. Phys. A Stat. Mech. Its Appl. 2019, 531, 121782. [Google Scholar] [CrossRef]
- Qi, H.; Meng, X. Dynamics of a stochastic predator-prey model with fear effect and hunting cooperation. J. Appl. Math. Comput. 2022, 69, 2077–2103. [Google Scholar] [CrossRef]
- Ch-Chaoui, M.; Mokni, K. A multi-parameter bifurcation analysis of a prey–predator model incorporating the prey Allee effect and predator-induced fear. Nonlinear Dyn. 2025, 2023, 1–23. [Google Scholar] [CrossRef]
- Feng, Y.; Meng, X. Dynamic behaviors of a diffusion intraguild predation model with hunting cooperation. Adv. Appl. Math. 2022, 11, 4323–4334. [Google Scholar] [CrossRef]
- Ji, J.; Lin, G.; Wang, L.; Mai, A. Spatiotemporal dynamics induced by intraguild predator diffusion in an intraguild predation model. J. Math. Biol. 2022, 85, 1. [Google Scholar] [CrossRef] [PubMed]
- Friederike Prowe, A.E.; Su, B.; Nejstgaard, J.C.; Schartau, M. Food web structure and intraguild predation affect ecosystem functioning in an established plankton model. Limnol. Oceanogr. 2022, 67, 843–855. [Google Scholar] [CrossRef]
- Giricheva, E. Stability and bifurcation analysis of a tri-trophic food chain model with intraguild predation. Int. J. Biomath. 2022, 16, 2250073. [Google Scholar] [CrossRef]
- Karnatak, R.; Wollrab, S. Mixotrophy and intraguild predation—Dynamic consequences of shifts between food web motifs. Eur. Phys. J. Spec. Top. 2017, 226, 2135–2144. [Google Scholar] [CrossRef]
- Chen, M.; Zhang, J.; Ejegul, S. Dynamics of autotroph–mixotroph interactions with the intraguild predation structure. J. Biol. Dyn. 2022, 16, 186–206. [Google Scholar] [CrossRef]
- Woodward, G.; Hildrew, A.G. Body-size determinants of niche overlap and intraguild predation within a complex food web. J. Anim. Ecol. 2002, 71, 1063–1074. [Google Scholar] [CrossRef]
- Costa, M.I.d.S.; dos Anjos, L. Intraguild predation does not necessarily have negative effects on pest biological control: Insights from a multiple consumer-multiple resource food web population model. BioControl 2022, 67, 263–274. [Google Scholar] [CrossRef]
- Amiri-Jami, A. Effect of ant-attendance on the occurrence of intraguild predation. Food Webs 2022, 32, e00240. [Google Scholar] [CrossRef]
- Faria, L.D.B.; Costa, M.I.d.S.; Godoy, W.A.C. A model for intraguild predation dynamics between immature stages. Ecol. Model. 2011, 222, 3295–3299. [Google Scholar] [CrossRef]
- Meng, X.-Y.; Xiao, L. Hopf bifurcation and Turing instability of a delayed diffusive Zooplankton–Phytoplankton model with hunting cooperation. Int. J. Bifurc. Chaos 2024, 34, 2450090. [Google Scholar] [CrossRef]
- Li, Y.; Ding, M. Impact of hunting cooperation in predator and anti-predator behaviors in prey in a predator–prey model. Math. Methods Appl. Sci. 2024, 48, 237–252. [Google Scholar] [CrossRef]
- Tao, W.; Wang, Z.-A. Global well-posedness and Turing–Hopf bifurcation of prey-taxis systems with hunting cooperation. Eur. J. Appl. Math. 2025, 1–27. [Google Scholar] [CrossRef]
- Peng, M.; Lin, R.; Zhang, Z.; Huang, L. The dynamics of a delayed predator-prey model with square root functional response and stage structure. Electron. Res. Arch. 2024, 32, 3275–3298. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 0.15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naser, A.H.; Bahlool, D.K. Dynamics of an Intraguild Predation Food Web Cooperation Model Under the Influence of Fear and Hunting. Computation 2025, 13, 128. https://doi.org/10.3390/computation13060128
Naser AH, Bahlool DK. Dynamics of an Intraguild Predation Food Web Cooperation Model Under the Influence of Fear and Hunting. Computation. 2025; 13(6):128. https://doi.org/10.3390/computation13060128
Chicago/Turabian StyleNaser, Alyaa Hussain, and Dahlia Khaled Bahlool. 2025. "Dynamics of an Intraguild Predation Food Web Cooperation Model Under the Influence of Fear and Hunting" Computation 13, no. 6: 128. https://doi.org/10.3390/computation13060128
APA StyleNaser, A. H., & Bahlool, D. K. (2025). Dynamics of an Intraguild Predation Food Web Cooperation Model Under the Influence of Fear and Hunting. Computation, 13(6), 128. https://doi.org/10.3390/computation13060128