


Stability of Impaired Humoral Immunity HIV-1 Models with Active and Latent Cellular Infections

Abstract
:1. Introduction
2. Model Incorporating Impaired Humoral Immunity and CI
2.1. Description of the System
2.2. Main Basic Properties
2.2.1. Maintaining Non-Negativity and Boundedness in the Solutions
2.2.2. Analysis of Reproductive Numbers and Equilibrium Points
- (i)
- the system always has an infection-free equilibrium point , and
- (ii)
- if , the system also has an infected equilibrium point .
- 1
- 2
- If , we have the equation . In this scenario, let us introduce a function defined on the interval as:Then
2.2.3. The Analysis of the Stability of the Equilibria and
3. Modeling Hiv-1 with Distributed Delays
3.1. Description of the System
- Healthy cells, which are contacted by HIV-1 particles or infected cells at time t, become (HIV-1)-latently infected cells, time units later. The recruitment of (HIV-1)-latently infected cells at time t is given by the number of cells that were newly contacted at time and are still alive at time t. Here, is assumed to be a constant death rate for contacted cells. Thus, the probability of surviving the time period from to t is .
- (HIV-1)-latently infected cells, become (HIV-1)-actively infected cells, time units later. The recruitment of (HIV-1)-actively infected cells at time t is given by the number of cells that were newly being (HIV-1)-latently infected cells at time and are still alive at time t. Here, is assumed to be a constant death rate for (HIV-1)-latently infected cells. Thus, the probability of surviving the time period from to t is .
- (HIV-1)-actively infected cells, produce new mature HIV-1 particles, time units later. The recruitment of HIV-1 particles at time t is given by the number of cells that were newly being (HIV-1)-actively infected cells at time and are still alive at time t. Here, is assumed to be a constant death rate for (HIV-1)-actively infected cells. Thus, the probability of surviving the time period from to t is .
3.2. Main Basic Properties
3.2.1. Maintaining Non-Negativity and Ultimate Boundedness in the Solutions
3.2.2. Analysis of Reproductive Numbers and Equilibrium Points
- (i)
- the system always has an infection-free equilibrium point , and
- (ii)
- if , the system also has an infected equilibrium point .
- 1
- 2
3.2.3. The Analysis of the Stability of the Equilibria and
4. Numerical Simulations
4.1. Numerical Simulation for Model (5)
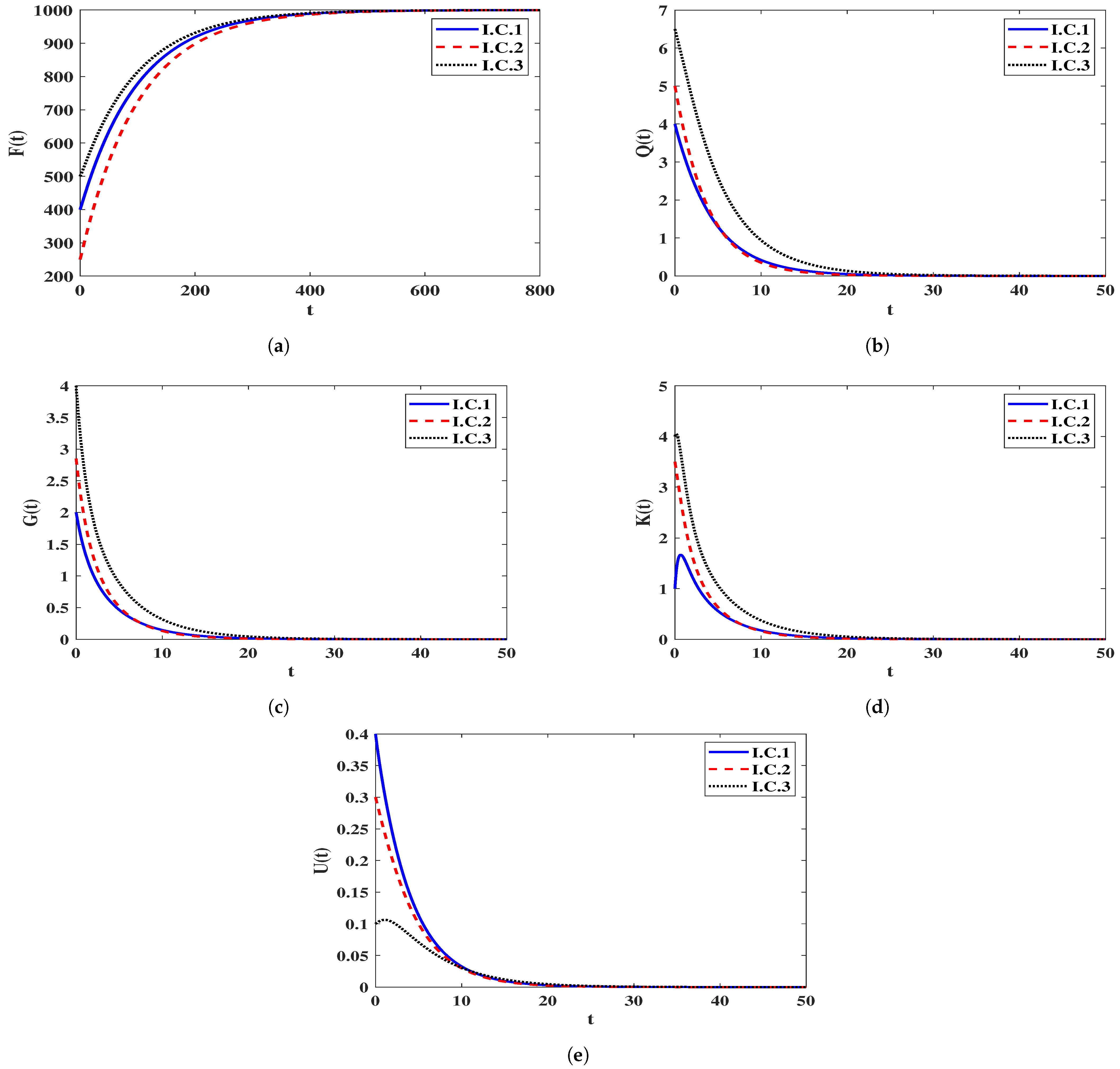
4.1.1. Stability of Equilibria
4.1.2. Effect of the Impaired Humoral Immunity
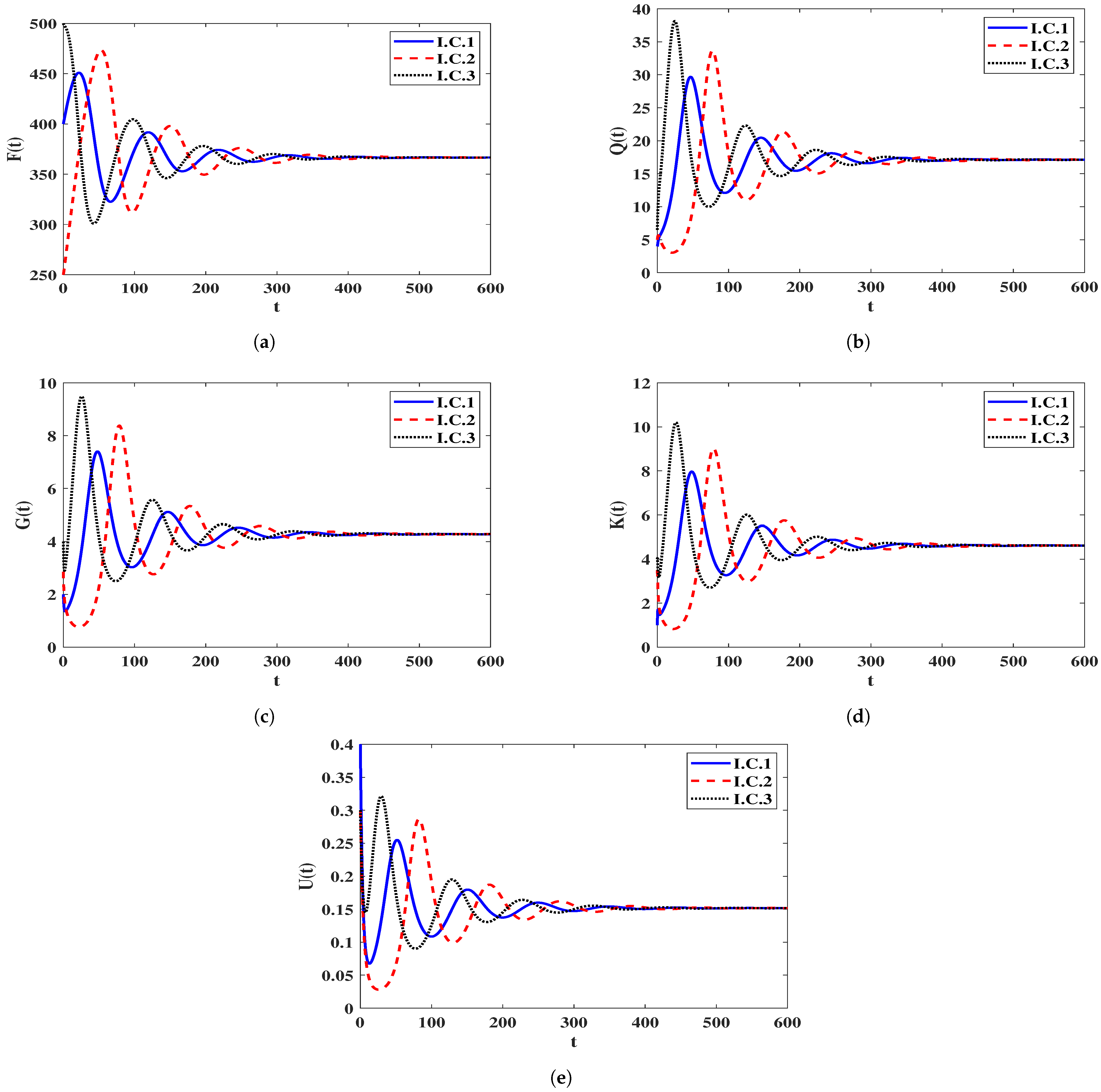
4.2. Numerical Simulation for Model (23)
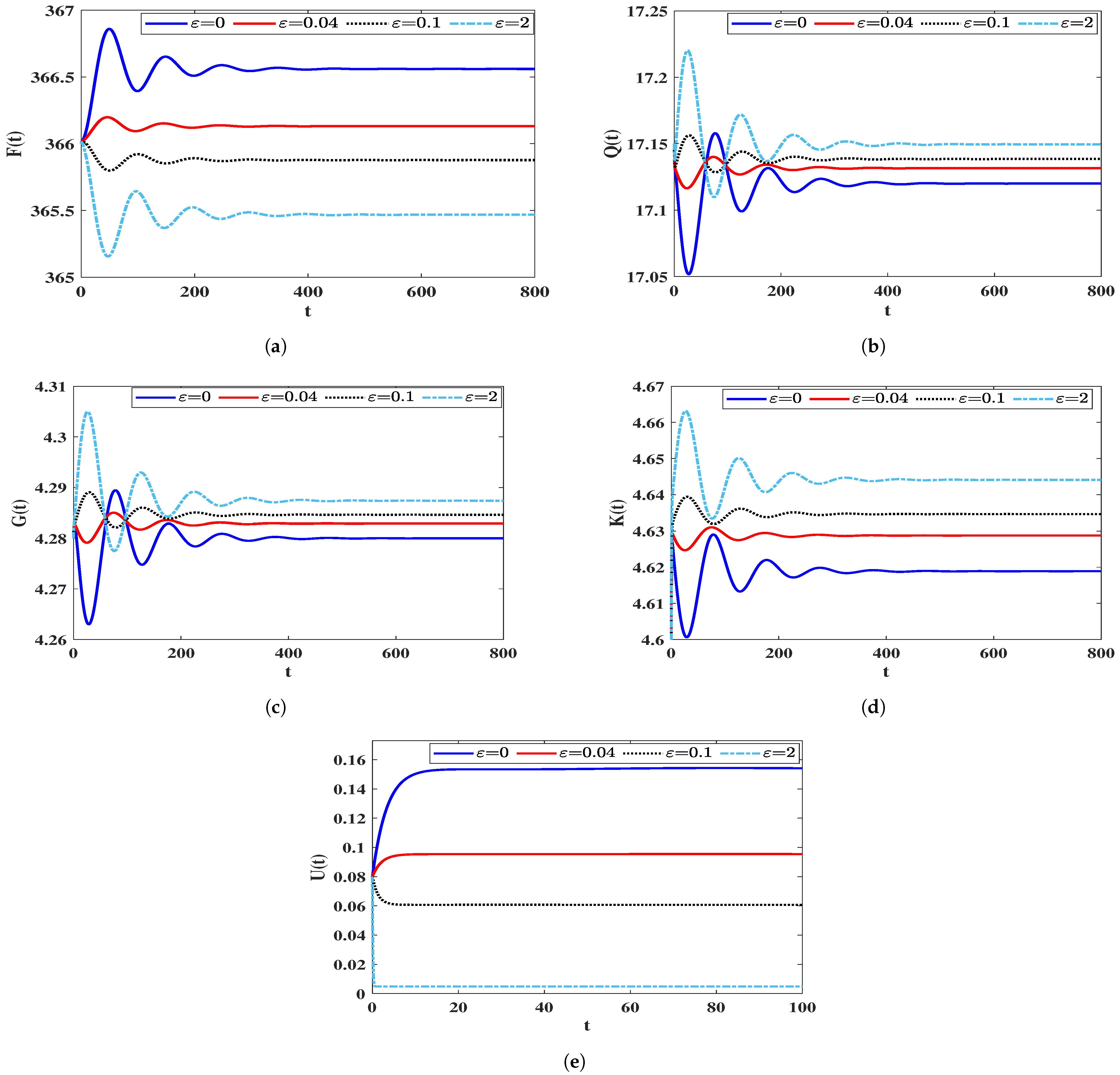
The Effect of the Time-Delays on the Stability of Equilibria
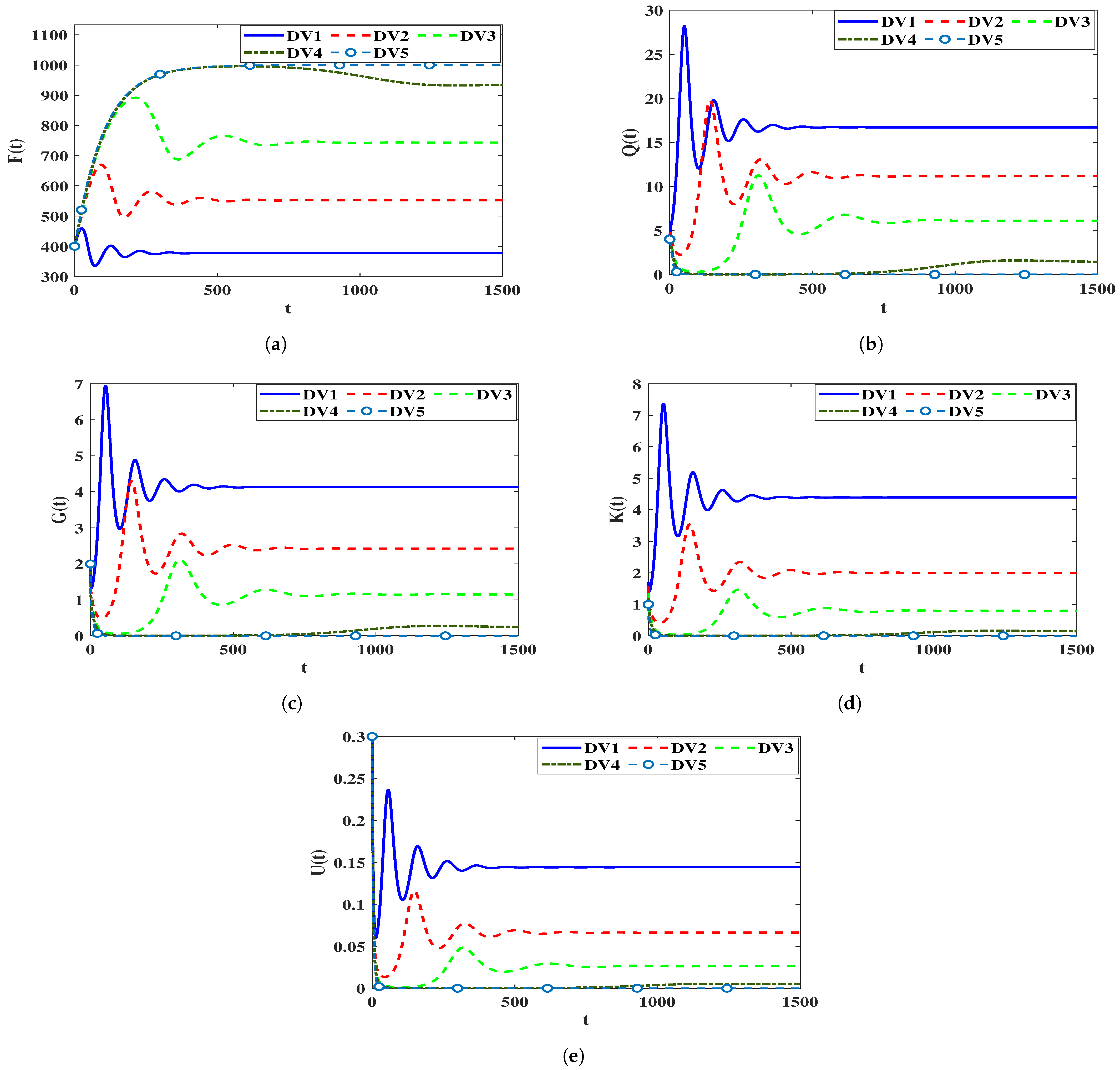
4.3. Sensitivity Analysis
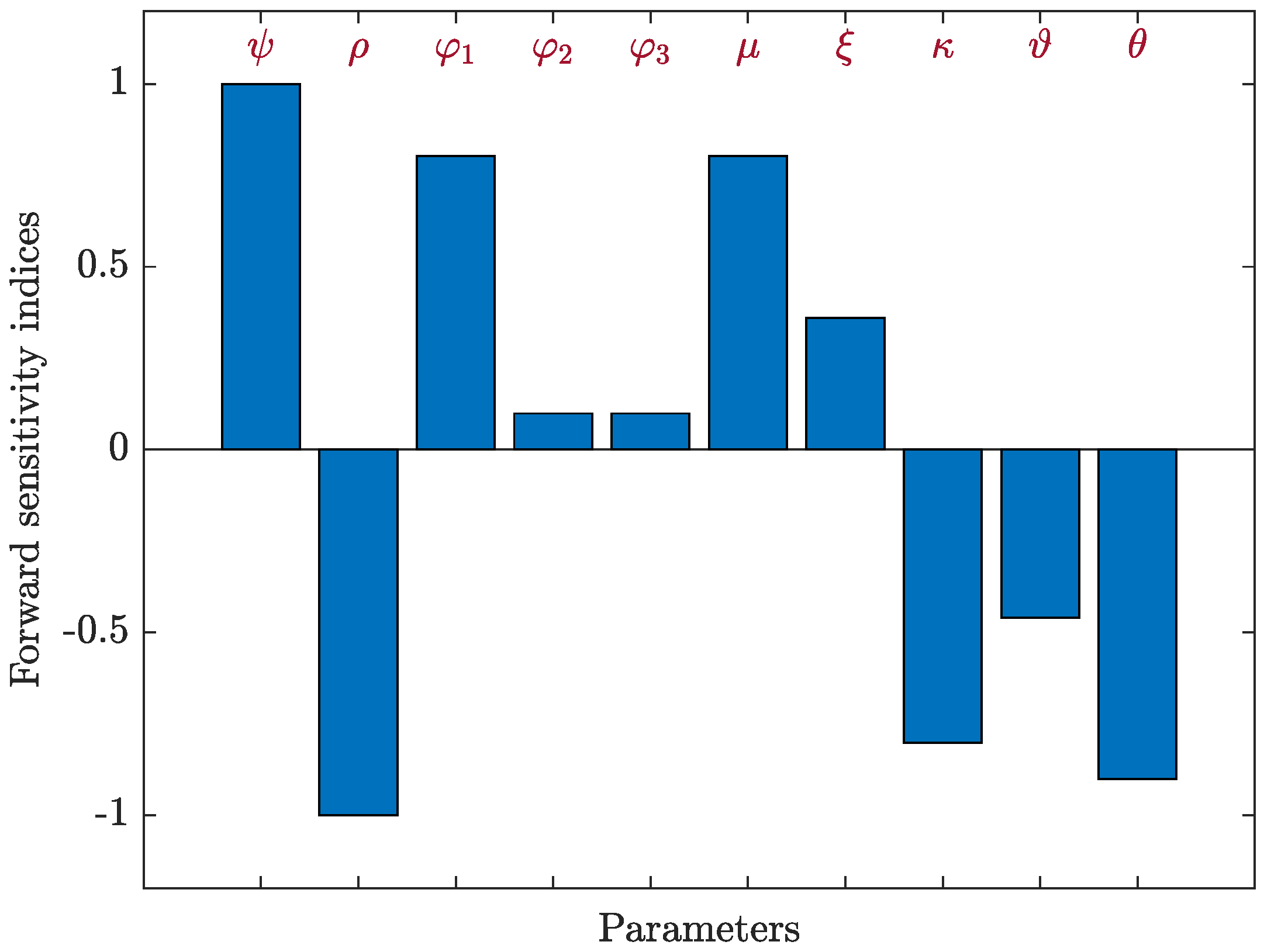
4.3.1. Sensitivity Analysis for Model (5)
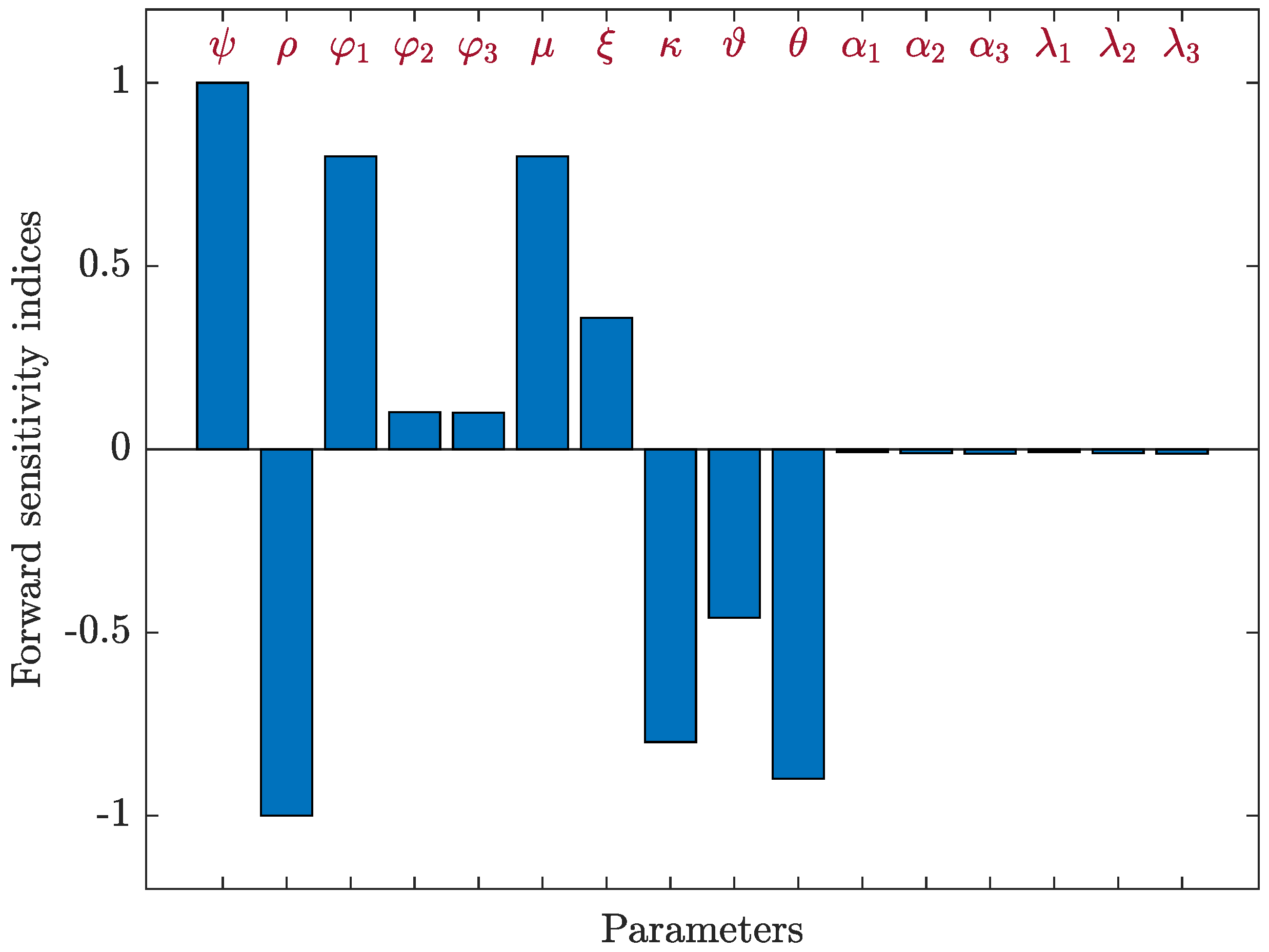
4.3.2. Sensitivity Analysis for Model (40)
5. Discussion
- 1.
- Limited Availability of Real Data: There is a scarcity of real data from HIV-1 infected individuals, which hinders the accurate estimation of model parameters.
- 2.
- Precision Issues: Comparing our obtained results with the limited existing studies may lack precision due to the scarcity of data points.
- 3.
- Data Collection Challenges: Collecting real data from HIV-1 infected patients can be a challenging and resource-intensive task.
- 4.
- Experimental Scope: Conducting experiments to obtain real data falls outside the scope of this paper.
6. Conclusions
Future Works
- Utilizing real-world data to estimate model parameters accurately, which can enhance the model’s predictive capabilities and align it better with empirical observations.
- Broadening the scope of the model to incorporate the role of Cytotoxic T Lymphocytes (CTLs) alongside B-cells, allowing for a more comprehensive representation of the immune response.
- Investigating the integration of age structure into the infected cell population within the model, which can provide insights into how age-related factors impact disease dynamics.
- Exploring the effects of viral mutations on the dynamics of the model, considering how genetic changes in the virus may influence disease progression and response to interventions.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wodarz, D.; Levy, D.N. Human immunodeficiency virus evolution towards reduced replicative fitness in vivo and the development of AIDS. Proc. R. Soc. Biol. Sci. 2007, 274, 2481–2491. [Google Scholar] [CrossRef]
- Available online: https://www.who.int/data/gho/data/themes/hiv-aids (accessed on 1 July 2023).
- Wodarz, D.; May, R.M.; Nowak, M.A. The role of antigen-independent persistence of memory cytotoxic T lymphocytes. Int. Immunol. 2000, 12, 467–477. [Google Scholar] [CrossRef]
- Elaiw, A.M.; AlShamrani, N.H. Global stability of humoral immunity virus dynamics models with nonlinear infection rate and removal. Nonlinear Anal. Real World Appl. 2015, 26, 161–190. [Google Scholar] [CrossRef]
- Wang, S.; Zou, D. Global stability of in host viral models with humoral immunity and intracellular delays. Appl. Math. Model. 2012, 36, 1313–1322. [Google Scholar] [CrossRef]
- Xu, J.; Zhou, Y.; Li, Y.; Yang, Y. Global dynamics of a intracellular infection model with delays and humoral immunity. Math. Methods Appl. Sci. 2016, 39, 427–5435. [Google Scholar] [CrossRef]
- Miao, H.; Teng, Z.; Kang, C.; Muhammadhaji, A. Stability analysis of a virus infection model with humoral immunity response and two time delays. Math. Methods Appl. Sci. 2016, 39, 3434–3449. [Google Scholar] [CrossRef]
- Tang, S.; Teng, Z.; Miao, H. Global dynamics of a reaction–diffusion virus infection model with humoral immunity and nonlinear incidence. Comput. Math. Appl. 2019, 78, 786–806. [Google Scholar] [CrossRef]
- Zheng, T.; Luo, Y.; Teng, Z. Spatial dynamics of a viral infection model with immune response and nonlinear incidence. Z. Angew. Math. Phys. 2023, 74, 124. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Yuan, S. Global dynamics of an age-structured virus model with saturation effects. Math. Methods Appl. Sci. 2017, 40, 1851–1864. [Google Scholar] [CrossRef]
- Kajiwara, T.; Sasaki, T.; Otani, Y. Global stability for an age-structured multistrain virus dynamics model with humoral immunity. J. Appl. Math. Comput. 2020, 62, 239–279. [Google Scholar] [CrossRef]
- Avila-Vales, E.; Pérez, Á.G. Global properties of an age-structured virus model with saturated antibody-immune response, multi-target cells, and general incidence rate. Boletín Soc. Matemática Mex. 2021, 27, 26. [Google Scholar] [CrossRef]
- Nowak, M.A.; May, R.M. Virus Dynamics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Inoue, T.; Kajiwara, T.; Sasaki, T. Global stability of models of humoral immunity against multiple viral strains. J. Biol. Dyn. 2010, 4, 282–295. [Google Scholar] [CrossRef] [PubMed]
- Dhar, M.; Samaddar, S.; Bhattacharya, P. Modeling the effect of non-cytolytic immune response on viral infection dynamics in the presence of humoral immunity. Nonlinear Dyn. 2019, 98, 637–655. [Google Scholar] [CrossRef]
- Dhar, M.; Samaddar, S.; Bhattacharya, P. Modeling the cell-to-cell transmission dynamics of viral infection under the exposure of non-cytolytic cure. J. Appl. Math. Comput. 2021, 65, 885–911. [Google Scholar] [CrossRef]
- Jolly, C.; Sattentau, Q. Retroviral spread by induction of virological synapses. Traffic 2004, 5, 643–650. [Google Scholar] [CrossRef]
- Sato, H.; Orenstein, J.; Dimitrov, D.; Martin, M. Cell-to-cell spread of HIV-1 occurs within minutes and may not involve the participation of virus particles. Virology 1992, 186, 712–724. [Google Scholar] [CrossRef]
- Iwami, S.; Takeuchi, J.S.; Nakaoka, S.; Mammano, F.; Clavel, F.; Inaba, H.; Kobayashi, T.; Misawa, N.; Aihara, K.; Koyanagi, Y.; et al. Cell-to-cell infection by HIV contributes over half of virus infection. eLife 2015, 4, e08150. [Google Scholar] [CrossRef]
- Komarova, N.L.; Wodarz, D. Virus dynamics in the presence of synaptic transmission. Math. Biosci. 2013, 242, 161–171. [Google Scholar] [CrossRef]
- Sourisseau, M.; Sol-Foulon, N.; Porrot, F.; Blanchet, F.; Schwartz, O. Inefficient human immunodeficiency virus replication in mobile lymphocytes. J. Virol. 2007, 81, 1000–1012. [Google Scholar] [CrossRef]
- Sigal, A.; Kim, J.T.; Balazs, A.B.; Dekel, E.; Mayo, A.; Milo, R.; Baltimore, D. Cell-to-cell spread of HIV permits ongoing replication despite antiretroviral therapy. Nature 2011, 477, 95–98. [Google Scholar] [CrossRef]
- Martin, N.; Sattentau, Q. Cell-to-cell HIV-1 spread and its implications for immune evasion. Curr. Opin. HIV AIDS 2009, 4, 143–149. [Google Scholar] [CrossRef]
- Lin, J.; Xu, R.; Tian, X. Threshold dynamics of an HIV-1 virus model with both virus-to-cell and cell-to-cell transmissions, intracellular delay, and humoral immunity. Appl. Math. Comput. 2017, 315, 516–530. [Google Scholar] [CrossRef]
- Luo, Y.; Zhang, L.; Zheng, T.; Teng, Z. Analysis of a diffusive virus infection model with humoral immunity, cell-to-cell transmission and nonlinear incidence. Phys. Stat. Mech. Its Appl. 2019, 535, 122415. [Google Scholar] [CrossRef]
- Lydyard, P.; Whelan, A.; Fanger, M. BIOS Instant Notes in Immunology; Taylor & Francis e-Library: Abingdon, UK, 2005. [Google Scholar]
- Miao, H.; Abdurahman, X.; Teng, Z.; Zhang, L. Dynamical analysis of a delayed reaction-diffusion virus infection model with logistic growth and humoral immune impairment. Chaos Solitons Fractals 2018, 110, 280–291. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Alshehaiween, S.F.; Hobiny, A.D. Global properties of delay-distributed HIV dynamics model including impairment of B-cell functions. Mathematics 2019, 7, 837. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Alshehaiween, S.F. Global stability of delay-distributed viral infection model with two modes of viral transmission and B-cell impairment. Math. Methods Appl. Sci. 2020, 43, 6677–6701. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Alshehaiween, S.F.; Hobiny, A.D. Impact of B-cell impairment on virus dynamics with time delay and two modes of transmission. Chaos Solitons Fractals 2020, 130, 109455. [Google Scholar] [CrossRef]
- Miao, H.; Liu, R.; Jiao, M. Global dynamics of a delayed latent virus model with both virus-to-cell and cell-to-cell transmissions and humoral immunity. J. Inequalities Appl. 2021, 2021, 156. [Google Scholar] [CrossRef]
- Elaiw, A.M.; AlShamrani, N.H. Stability of a general adaptive immunity virus dynamics model with multi-stages of infected cells and two routes of infection. Math. Methods Appl. Sci. 2020, 43, 1145–1175. [Google Scholar] [CrossRef]
- Elaiw, A.M.; AlShamrani, N.H. Global stability of a delayed adaptive immunity viral infection with two routes of infection and multi-stages of infected cells. Commun. Nonlinear Sci. Numer. Simul. 2020, 86, 105259. [Google Scholar] [CrossRef]
- Agosto, L.; Herring, M.; Mothes, W.; Henderson, A. HIV-1-infected CD4+ T cells facilitate latent infection of resting CD4+ T cells through cell-cell contact. Cell Rep. 2018, 24, 2088–2100. [Google Scholar] [CrossRef]
- Wang, W.; Wang, X.; Guo, K.; Ma, W. Global analysis of a diffusive viral model with cell-to-cell infection and incubation period. Math. Methods Appl. Sci. 2020, 43, 5963–5978. [Google Scholar] [CrossRef]
- Alshamrani, N.H. Stability of a general adaptive immunity HIV infection model with silent infected cell-to-cell spread. Chaos Solitons Fractals 2021, 150, 110422. [Google Scholar] [CrossRef]
- Elaiw, A.M.; AlShamrani, N.H. Stability of a delayed adaptive immunity hiv infection model with silent infected cells and cellular infection. J. Appl. Anal. Comput. 2021, 11, 964–1005. [Google Scholar] [CrossRef] [PubMed]
- Hattaf, K.; Dutta, H. Modeling the dynamics of viral infections in presence of latently infected cells. Chaos Solitons Fractals 2020, 136, 109916. [Google Scholar] [CrossRef]
- van den Driessche, P.; Watmough, J. Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission. Math. Biosci. 2002, 180, 29–48. [Google Scholar] [CrossRef]
- Willems, J.L. Stability Theory of Dynamical Systems; Wiley: New York, NY, USA, 1970. [Google Scholar]
- Korobeinikov, A. Global properties of basic virus dynamics models. Bull. Math. Biol. 2004, 66, 879–883. [Google Scholar] [CrossRef] [PubMed]
- Hale, J.K.; Lunel, S.M.V. Introduction to Functional Differential Equations; Springer: New York, NY, USA, 1993. [Google Scholar]
- Kuang, Y. Delay Differential Equations with Applications in Population Dynamics; Academic Press: San Diego, CA, USA, 1993. [Google Scholar]
- Hetmaniok, E.; Pleszczyński, M. Comparison of the selected methods used for solving the ordinary differential equations and their systems. Mathematics 2022, 10, 306. [Google Scholar] [CrossRef]
- Hetmaniok, E.; Pleszczyński, M.; Khan, Y. Solving the integral differential equations with delayed argument by using the DTM method. Sensors 2022, 22, 4124. [Google Scholar] [CrossRef]
- Sahani, S.K.; YASHI. Effects of eclipse phase and delay on the dynamics of HIV infection. J. Biol. Syst. 2018, 26, 421–454. [Google Scholar] [CrossRef]
- Wang, S.; Song, X.; Ge, Z. Dynamics analysis of a delayed viral infection model with immune impairment. Appl. Math. Model. 2011, 35, 4877–4885. [Google Scholar] [CrossRef]
- Allali, K.; Danane, J. Global analysis for an HIV infection model with CTL immune response and infected cells in eclipse phase. Appl. Sci. 2017, 7, 861. [Google Scholar] [CrossRef]
- Sun, C.; Li, L.; Jia, J. Hopf bifurcation of an HIV-1 virus model with two delays and logistic growth. Math. Model. Nat. Phenom. 2020, 15, 16. [Google Scholar] [CrossRef]
- Wang, X.; Rong, L. HIV low viral load persistence under treatment: Insights from a model of cell-to-cell viral transmission. Appl. Math. Lett. 2019, 94, 44–51. [Google Scholar] [CrossRef]
- Bellomo, N.; Painter, K.J.; Tao, Y.; Winkler, M. Occurrence vs. Absence of taxis-driven instabilities in a May-Nowak model for virus infection. SIAM J. Appl. Math. 2019, 79, 1990–2010. [Google Scholar] [CrossRef]
- Ren, X.; Tian, Y.; Liu, L.; Liu, X. A reaction-diffusion within-host HIV model with cell-to-cell transmission. J. Math. Biol. 2018, 76, 831–1872. [Google Scholar] [CrossRef]
- Bellomo, N.; Outada, N.; Soler, J.; Tao, Y.; Winkler, M. Chemotaxis and cross-diffusion models in complex environments: Models and analytic problems toward a multiscale vision. Math. Model. Methods Appl. Sci. 2022, 32, 713–792. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value | Reference | Parameter | Value | Reference |
|---|---|---|---|---|---|
| 10 | [46] | [47] | |||
| [46] | [48] | ||||
| varied | - | [48] | |||
| varied | - | [49] | |||
| varied | - | [46] | |||
| [46] | [46] | ||||
| [46] | varied | - |
| Equilibria | |
|---|---|
| Delay Parameters | Equilibria | |
|---|---|---|
| Parameter S | Value of | Parameter S | Value of |
|---|---|---|---|
| 1 | |||
| Parameter S | Value of | Parameter S | Value of |
|---|---|---|---|
| 1 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
AlShamrani, N.H.; Halawani, R.H.; Shammakh, W.; Elaiw, A.M. Stability of Impaired Humoral Immunity HIV-1 Models with Active and Latent Cellular Infections. Computation 2023, 11, 207. https://doi.org/10.3390/computation11100207
AlShamrani NH, Halawani RH, Shammakh W, Elaiw AM. Stability of Impaired Humoral Immunity HIV-1 Models with Active and Latent Cellular Infections. Computation. 2023; 11(10):207. https://doi.org/10.3390/computation11100207
Chicago/Turabian StyleAlShamrani, Noura H., Reham H. Halawani, Wafa Shammakh, and Ahmed M. Elaiw. 2023. "Stability of Impaired Humoral Immunity HIV-1 Models with Active and Latent Cellular Infections" Computation 11, no. 10: 207. https://doi.org/10.3390/computation11100207
APA StyleAlShamrani, N. H., Halawani, R. H., Shammakh, W., & Elaiw, A. M. (2023). Stability of Impaired Humoral Immunity HIV-1 Models with Active and Latent Cellular Infections. Computation, 11(10), 207. https://doi.org/10.3390/computation11100207