Psychophysiological Measures of Reactance to Persuasive Messages Advocating Limited Meat Consumption

 ,
,  and
and
Abstract
:1. Introduction
1.1. Psychological Reactance Is Situation Specific
1.2. Measuring the Psychophysiology of Reactant Responses
1.3. Study Aim and Hypotheses
2. Method
2.1. Participants
2.2. Manipulation
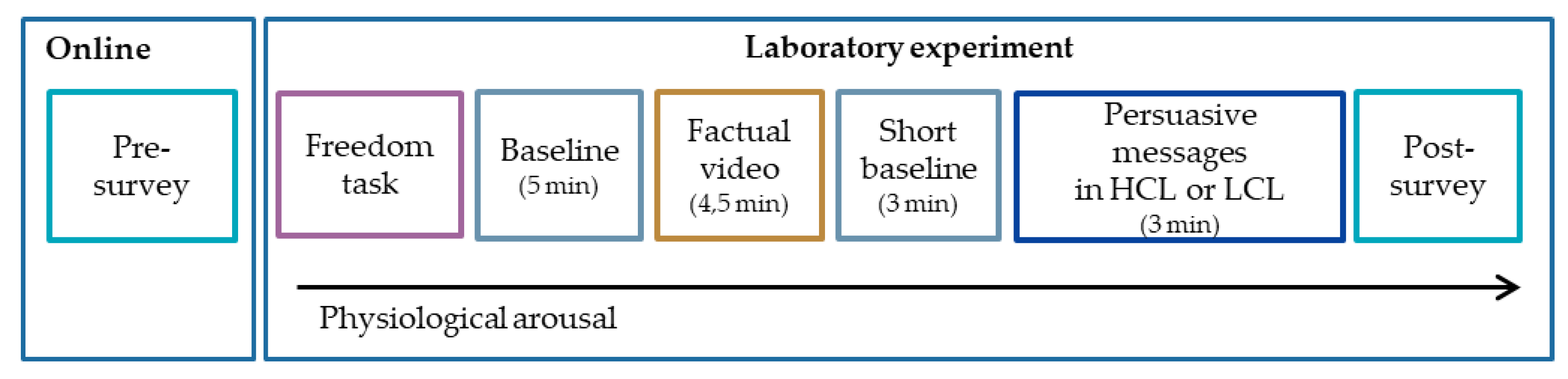
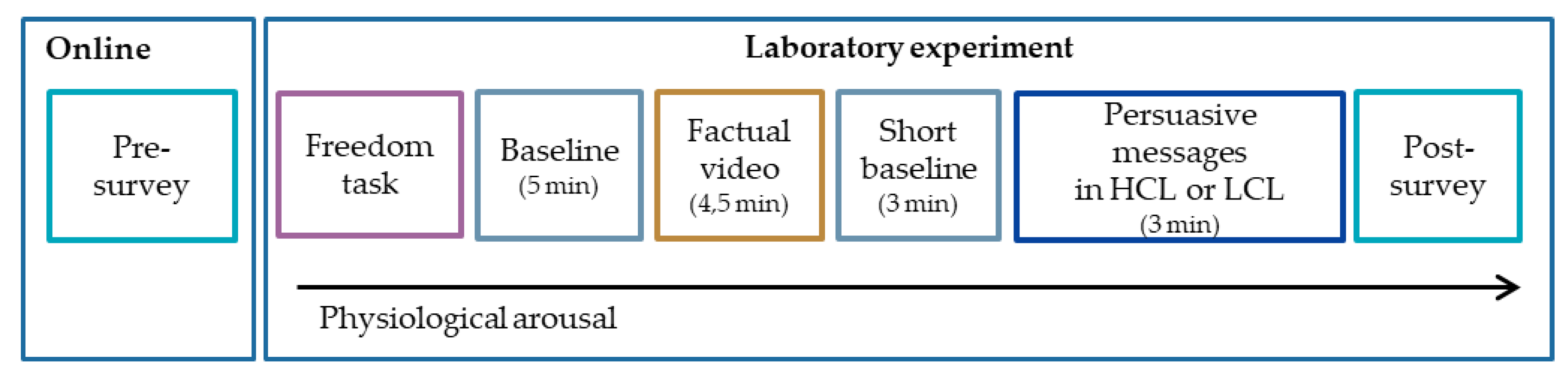
2.3. Procedure
2.4. Materials
2.4.1. Subjective Data
2.4.2. Physiological Data
2.5. Analyses
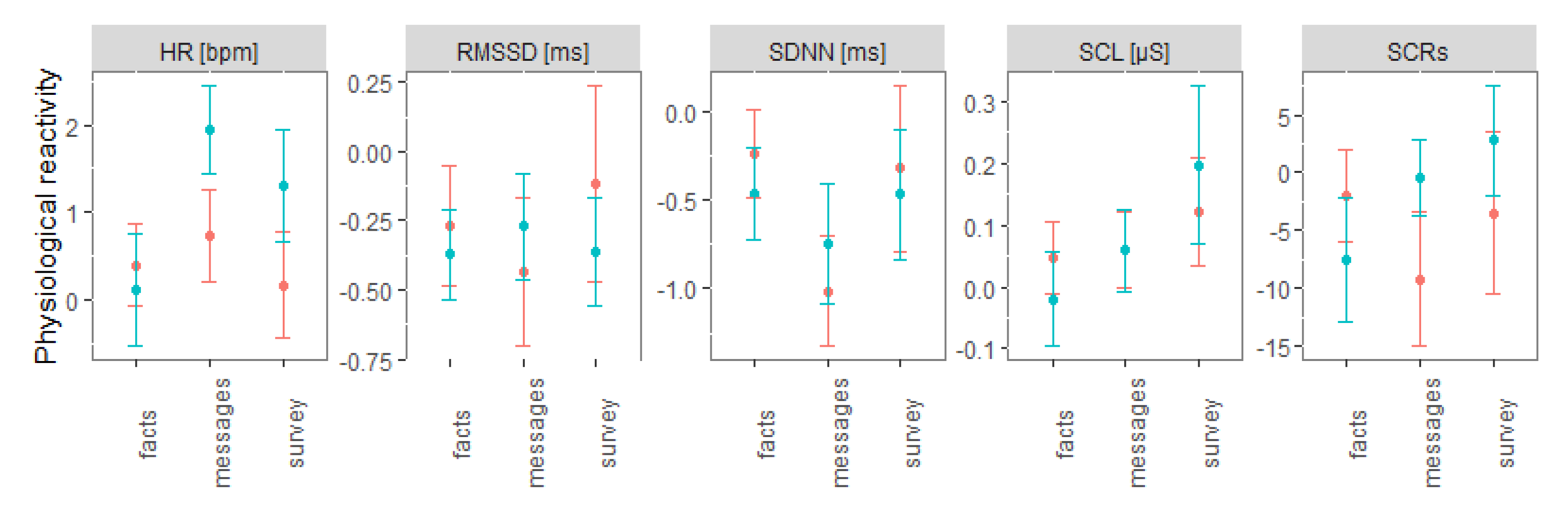
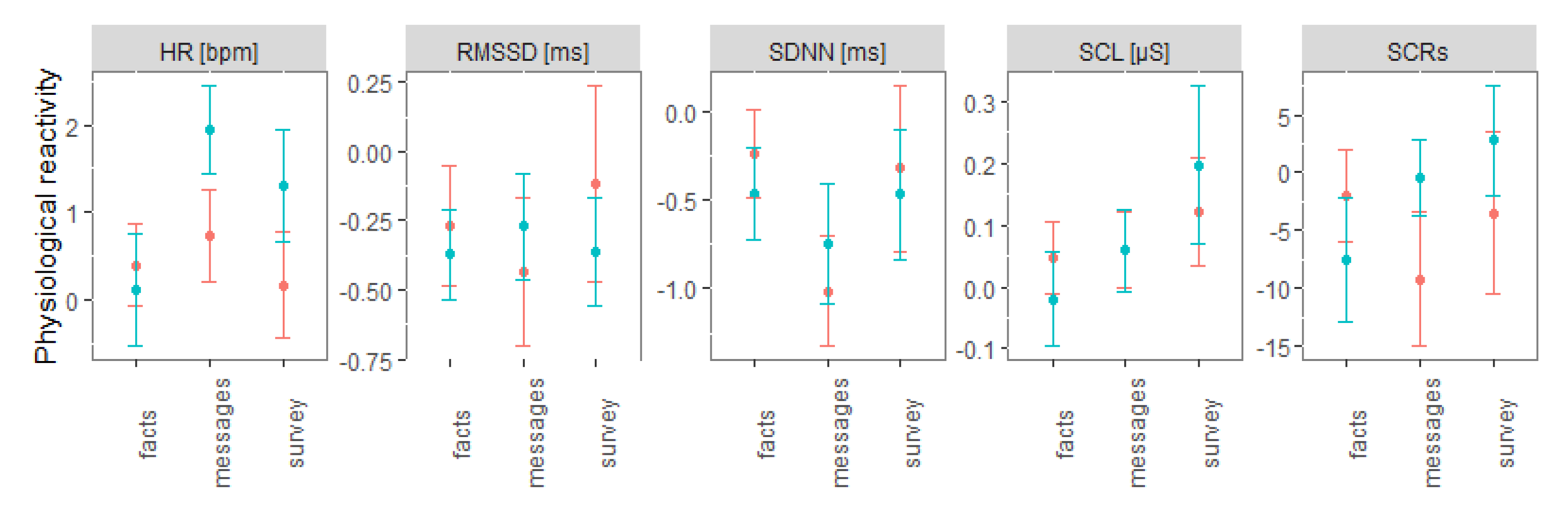
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Miron, A.M.; Brehm, J.W. Reactance Theory—40 Years Later. Z. Sozialpsychol. 2006, 37, 9–18. [Google Scholar] [CrossRef]
- Kreibig, S.D. Autonomic nervous system activity in emotion: A review. Biol. Psychol. 2010, 84, 394–421. [Google Scholar] [CrossRef] [PubMed]
- Sittenthaler, S.; Steindl, C.; Jonas, E. Legitimate vs. illegitimate restrictions–A motivational and physiological approach investigating reactance processes. Front. Psychol. 2015, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Picard, R.W.; Vyzas, E.; Healey, J. Toward machine emotional intelligence: Analysis of affective physiological state. IEEE Trans. Pattern Anal. Mach. Intell. 2001, 23, 1175–1191. [Google Scholar] [CrossRef]
- Steindl, C.; Jonas, E.; Sittenthaler, S.; Traut-Mattausch, E.; Greenberg, J. Understanding psychological reactance: New developments and findings. Z. Psychol. 2015, 223, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Fairclough, S.H. Fundamentals of physiological computing. Interact. Comput. 2009, 21, 133–145. [Google Scholar] [CrossRef]
- Dillard, J.P.; Shen, L. On the nature of reactance and its role in persuasive health communication. Commun. Monogr. 2005, 72, 144–168. [Google Scholar] [CrossRef]
- Rains, S.A. The nature of psychological reactance revisited: A meta-analytic review. Hum. Commun. Res. 2013, 39, 47–73. [Google Scholar] [CrossRef]
- Brehm, J.W. A Theory of Psychological Reactance; Academic Press: Oxford, UK, 1966. [Google Scholar]
- Ajzen, I. Constructing a TPB Questionnaire: Conceptual and Methodological Considerations. Available online: people.umass.edu/aizen/pdf/tpb.measurement.pdf (accessed on 22 August 2018).
- Ghazali, A.; Ham, J.; Barakova, E.; Markopoulos, P. The influence of social cues in persuasive social robots on psychological reactance and compliance. Comput. Human Behav. 2018, 87, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.H.; Lane, L.T.; Deatrick, L.M.; Young, A.M.; Potts, K.A. Psychological reactance and promotional health messages: The effects of controlling language, lexical concreteness, and the restoration of freedom. Hum. Commun. Res. 2007, 33, 219–240. [Google Scholar] [CrossRef]
- Picard, R.W. Affective Computing Research; MIT Media Laboratory: Cambridge, MA, USA, 1995. [Google Scholar]
- Cacioppo, J.T.; Berntson, G.G.; Larsen, J.T.; Poehlmann, K.M.; Ito, T.A. The psychophysiology of emotion. In Handbook of Emotions; Lewis, M., Haviland-Jones, J.M., Feldman Barrett, L., Eds.; Guilford Publications: New York, NY, USA, 2000; Volume 2, ISBN 978-1593856502. [Google Scholar]
- Jänig, W. The autonomic nervous system and its coordination by the brain. In Handbook of Affective Sciences; Davidson, R.J., Scherer, K.R., Goldsmith, H.H., Eds.; Oxford University Press: Oxford, UK, 2003; pp. 135–186. [Google Scholar]
- Cacioppo, J.T.; Tassinary, L.G.; Berntson, G.G. The Handbook of Psychophysiology; Cacioppo, J.T., Tassinary, L.G., Berntson, G.G., Eds.; Cambridge University Press: New York, NY, USA, 2007; Volume 44, ISBN 9780521844710. [Google Scholar]
- Shoemaker, J.K.; Norton, K.N.; Baker, J.; Luchyshyn, T.A. Forebrain organization for autonomic cardiovascular control. Auton. Neurosci. Basic Clin. 2015, 188, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Thayer, J.F.; Åhs, F.; Fredrikson, M.; Sollers, J.J.; Wager, T.D. A meta-analysis of heart rate variability and neuroimaging studies: Implications for heart rate variability as a marker of stress and health. Neurosci. Biobehav. Rev. 2012, 36, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Camm, A.J.; Malik, M.; Bigger, J.T.; Breithardt, G.; Cerutti, S.; Cohen, R.J.; Coumel, P.; Fallen, E.L.; Kennedy, H.L.; Kleiger, R.E.; et al. Heart rate variability: Standards of measurement, physiological interpretation and clinical use. Circulation 1996, 1043–1065. [Google Scholar]
- Thayer, J.F.; Lane, R.D. Claude Bernard and the heart-brain connection: Further elaboration of a model of neurovisceral integration. Neurosci. Biobehav. Rev. 2009, 33, 81–88. [Google Scholar] [CrossRef]
- Boucsein, W. Electrodermal Activity, 2nd ed.; Springer Science+Business Media: New York, NY, USA, 2012; ISBN 9781461411253. [Google Scholar]
- Jonas, E.; Graupmann, V.; Kayser, D.N.; Zanna, M.; Traut-Mattausch, E.; Frey, D. Culture, self, and the emergence of reactance: Is there a “universal” freedom? J. Exp. Soc. Psychol. 2009, 45, 1068–1080. [Google Scholar] [CrossRef]
- Sittenthaler, S.; Jonas, E.; Traut-Mattausch, E. Explaining self and vicarious reactance. Personal. Soc. Psychol. Bull. 2016, 42, 458–470. [Google Scholar] [CrossRef]
- Rogers, R.W. Cognitive and psychological processes in fear appeals and attitude change: A revised theory of protection motivation. In Social Psychophysiology: A Source Book; Cacioppo, J.T., Shapiro, D., Eds.; Guilford Press: New York, NY, USA, 1983; pp. 153–176. [Google Scholar]
- Loughnan, S.; Bastian, B.; Haslam, N. The psychology of eating animals. Curr. Dir. Psychol. Sci. 2014, 23, 104–108. [Google Scholar] [CrossRef]
- Overbeek, T.J.M.; van Boxtel, A.; Westerink, J.H.D.M. Respiratory sinus arrhythmia responses to induced emotional states: Effects of RSA indices, emotion induction method, age, and sex. Biol. Psychol. 2012, 91, 128–141. [Google Scholar] [CrossRef]
- RStudio: Integrated Development Environment for R 2016, version 3.5.2; Eggshell Igloo, Rstudio Team: Boston, MA, USA, 2016.
- Wickham, H. Tidyverse: Easily Install and Load the “Tidyverse”; RStudio: Boston, MA, USA, 2017. [Google Scholar]
- Revelle, W. Psych: Procedures for Psychological, Psychometric, and Personality Research; Northwestern University: Evanston, IL, USA, 2017. [Google Scholar]
- Zeileis, A.; Grothendieck, G.; Ryan, J.A.; Ulrich, J.M.; Andrews, F. Zoo: S3 Infrastructure for Regular and Irregular Time Series (Z’s Ordered Observations); RStudio: Boston, MA, USA, 2019. [Google Scholar]
- Carezia, A.; Dupuis, P.; Eaton, J.W.; Habel, K.; Hornik, K.; Krey, S.; Lash, B.; Leisch, F.; Smith, J.O.; Stewart, D.; et al. Signal Processing 2015; RStudio: Boston, MA, USA, 2015. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Chambers, J., Eddy, W., Härdle, W., Sheather, S., Tierney, L., Eds.; Springer Science+Business Media: New York, NY, USA, 2003; ISBN 978-1-4419-3008-8. [Google Scholar]
- Wagenmakers, E.; Farrell, S. AIC model selection using Akaike weights. Psychon. Bull. Rev. 2004, 11, 192–196. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Stoess, C.; Rozin, P.; Markwith, M. Moralization and becoming a vegetarian: The transformation of preferences into values and the recruitment of disgust. Psychol. Sci. 1997, 8, 67–73. [Google Scholar]
- Ajzen, I. The theory of planned behavior. Organ. Behav. Hum. Decis. Process. 1991, 50, 179–211. [Google Scholar] [CrossRef]
- Balconi, M.; Falbo, L.; Brambilla, E. BIS/BAS responses to emotional cues: Self report, autonomic measure and alpha band modulation. Pers. Individ. Dif. 2009, 47, 858–863. [Google Scholar] [CrossRef]
- Takahashi, T.; Murata, T.; Hamada, T.; Omori, M.; Kosaka, H.; Kikuchi, M.; Yoshida, H.; Wada, Y. Changes in EEG and autonomic nervous activity during meditation and their association with personality traits. Int. J. Psychophysiol. 2005, 55, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Koelsch, S.; Enge, J.; Jentschke, S. Cardiac signatures of personality. PLoS ONE 2012, 7, e31441. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Segment | Example Sentences |
|---|---|
| Threat-to-health/environment (202 words, presented to both groups) |
|
| Recommendation in HCL (226 words, presented to group 1) |
|
| Recommendation in LCL (224 words, presented to group 2) |
|
| Feelings of Anger | Perceived Threat to Freedom | |
|---|---|---|
| 1 | I was irritated | The video tried to make a decision for me |
| 2 | I was angry | The video tried to manipulate me |
| 3 | I was annoyed | The video tried to pressure me |
| 4 | I was aggravated | The video threatened my freedom to choose |
| Condition | HCL | LCL | |||||
|---|---|---|---|---|---|---|---|
| Scale | α | m | s.d. | m | s.d. | U | p |
| Anger | 0.91 | 3.45 | 1.35 | 2.84 | 1.12 | 280 | 0.034 |
| PTTF | 0.92 | 4.20 | 1.43 | 2.70 | 1.45 | 164 | 0.002 |
| HR | SDNN·102 | RMSSD·102 | SCL | SCRs | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Predictors | Est.1 | p | Est. | p | Est. | p | Est. | p | Est. | p |
| Persuasive messages (Intercept) | 1.30 | 0.001 | −0.89 | <0.001 | −0.36 | 0.040 | 0.06 | 0.294 | −4.56 | 0.190 |
| Factual video | −1.03 | 0.018 | 0.54 | 0.038 | 0.04 | 0.829 | −0.06 | 0.317 | −0.32 | 0.948 |
| Survey | −0.60 | 0.165 | 0.50 | 0.058 | 0.13 | 0.519 | 0.10 | 0.112 | 4.74 | 0.330 |
| Random Effects | ||||||||||
| σ2 | 5.25 | 2.26 | 1.10 | 0.11 | 639.99 | |||||
| ICC | 0.43 | 0.37 | 0.33 | 0.31 | 0.03 | |||||
| Obs. | 171 | 171 | 171 | 164 | 165 | |||||
| R2/Cond. R2 | 0.025 / 0.449 | 0.010 / 0.376 | 0.001 / 0.336 | 0.028 / 0.327 | 0.008 / 0.040 | |||||
| AIC | 847.072 | 692.645 | 563.667 | 163.913 | 1549.650 | |||||
| Null Model | State Model | Full Model | ||||
|---|---|---|---|---|---|---|
| Predictors | Est.1 | p | Est. | p | Est. | P |
| (Intercept) | 6.63 | <0.001 | 8.47 | <0.001 | 8.73 | <0.001 |
| Intention to limit meat consumption | −0.54 | 0.012 | −0.58 | 0.006 | ||
| HR reactivity to persuasive messages | −0.23 | 0.056 | ||||
| RMSSD reactivity to persuasive messages | −0.52 | 0.055 | ||||
| Obs. | 56 | 56 | 56 | |||
| R2 / Adj. R2 | 0.000 / 0.000 | 0.112 / 0.095 | 0.201 / 0.155 | |||
| AIC | 262.250 | 257.627 | 255.693 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spelt, H.; Dijk, E.K.-v.; Ham, J.; Westerink, J.; IJsselsteijn, W. Psychophysiological Measures of Reactance to Persuasive Messages Advocating Limited Meat Consumption. Information 2019, 10, 320. https://doi.org/10.3390/info10100320
Spelt H, Dijk EK-v, Ham J, Westerink J, IJsselsteijn W. Psychophysiological Measures of Reactance to Persuasive Messages Advocating Limited Meat Consumption. Information. 2019; 10(10):320. https://doi.org/10.3390/info10100320
Chicago/Turabian StyleSpelt, Hanne, Elisabeth Kersten-van Dijk, Jaap Ham, Joyce Westerink, and Wijnand IJsselsteijn. 2019. "Psychophysiological Measures of Reactance to Persuasive Messages Advocating Limited Meat Consumption" Information 10, no. 10: 320. https://doi.org/10.3390/info10100320