3. A Tie-Up Theory of Mating
In this section, we present our approach that unifies sex-focused and emotion-focused streams of literature on mating and couple formation and maintenance within a common framework, whose dynamic structure consists of sexual-emotional feedback loops, thus abridging the particular theoretical perspectives of SST and AT. For reasons that will become clear shortly, we refer to our model as the Tie-Up Theory. We regard it as a parsimonious representation and explanation of certain aspects of the dynamic interaction of heterosexual partners in romantic relationships. In particular, in the spirit of [
46], our scheme is designed to analyze open-loop mating strategies where long-term
vs. short-term orientation of the relationship is the endogenous outcome of the interaction itself, rather than a dispositional precondition of subjects—where such an outcome is the result of a complex interplay of the sexual and emotional dimensions. Likewise, our approach develops within a biosocial perspective that fully acknowledges the role of biological factors in determining male-female differences regarding the sexual and emotional aspects of mating, but also takes into account the regulatory role of social factors, in the spirit of [
99].
Our approach is consistent with much of the literature that has been presented in
Section 2. However, to ease readability and to keep the argument compact, in presenting it we will not emphasize all the correspondences that can be found with the literature review, as this would make the paper difficult to read and would subtract clarity and conciseness to the explanation of new concepts. As mere examples, the asymmetry in male
vs. female roles and psychological structures in our model can be easily traced back to the concepts and findings of SST, and we will not generally restate them during our presentation. The same applies to more specific aspects such as, say, courtship as a seduction strategy, sexual exploitation as the result of affective insecurity, deferment of sexual intercourse to incite attachment, and so on. We will limit ourselves to spare references, when they are of special importance, and will leave to the reader the task of re-formulating single aspects and implications of our model in the context of the literature review of
Section 2. As can be expected, not all aspects or implications of our approach find a specific corroboration in the existing literature, and further experimental and empirical work will be needed to this purpose, as it is common when a new theoretical approach is proposed.
3.1. Basic Concepts
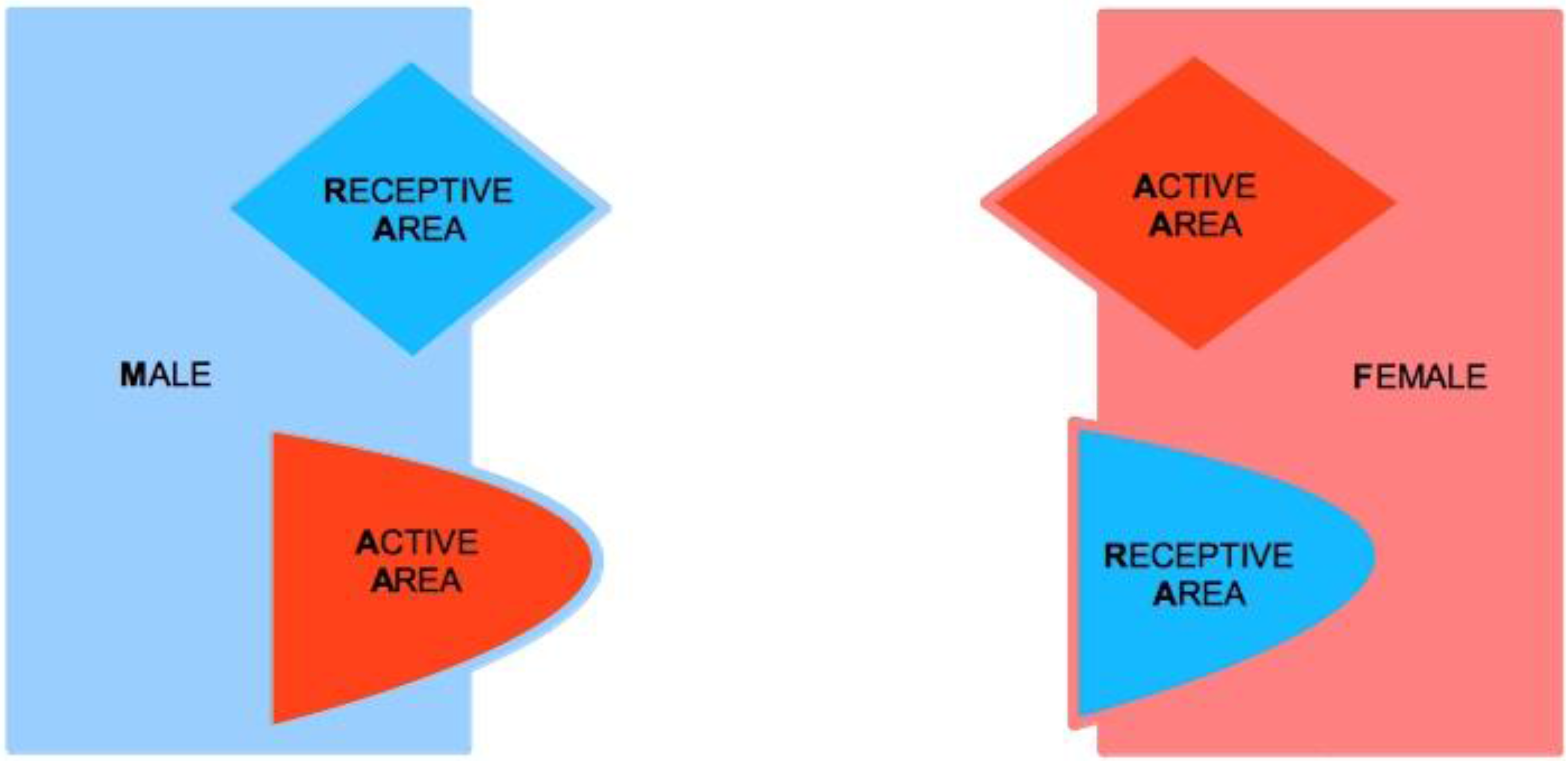
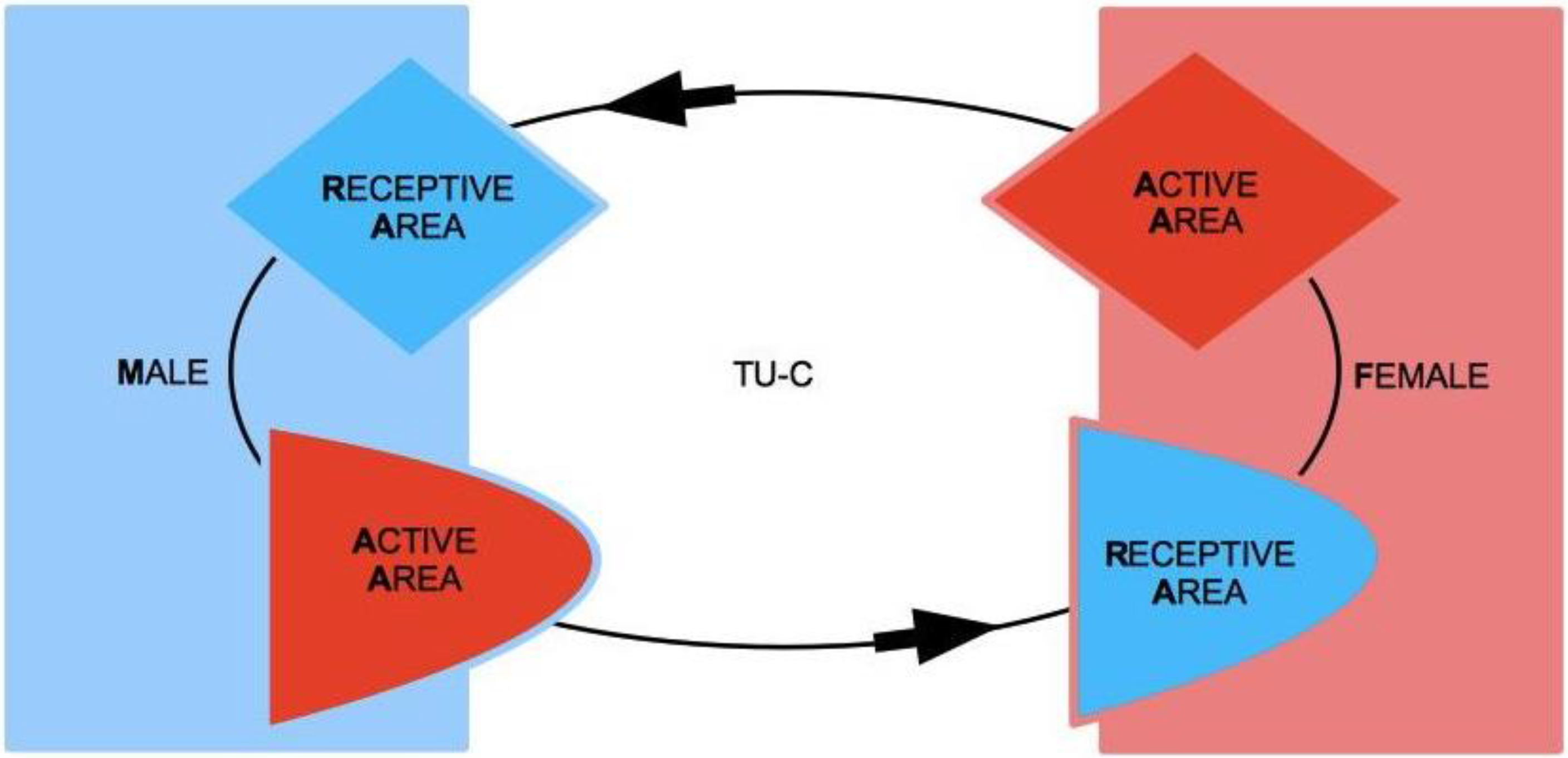
Let us consider two subjects, Male (M) and Female (F), both of which characterized by two complementary psychological structures, which we call the Active Area (AA) and the Receptive Area (RA), respectively. We therefore speak of Male Active Area (M-AA) and Male Receptive Area (M-RA), and likewise of Female Active Area (F-AA) and Female Receptive Area (F-RA)—see
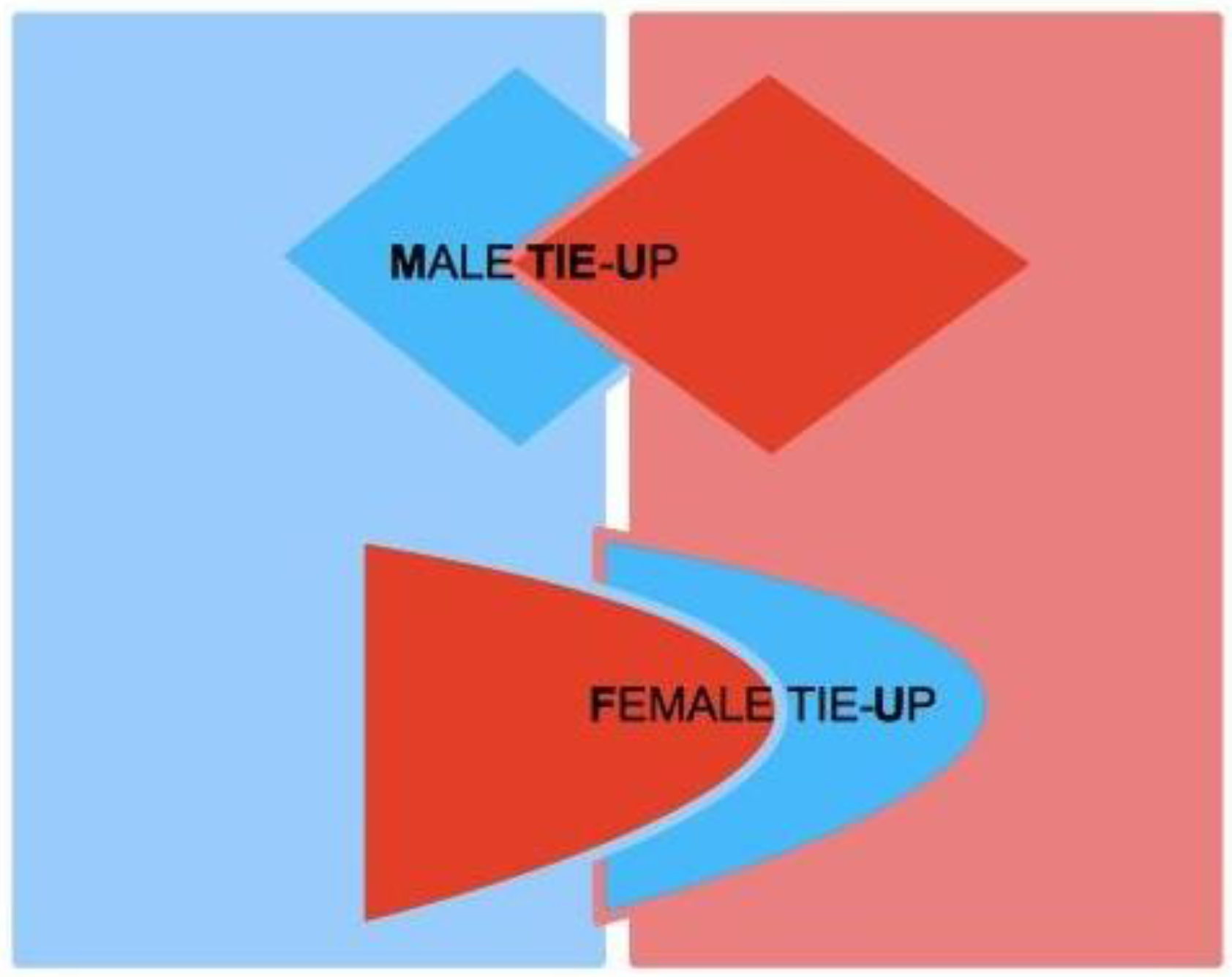
Figure 1, where red codes AAs, and blue codes RAs. AAs and RAs are the components of a dynamical system that determines whether or not M and F are bonded by some type of more or less stable, prolonged (sexual and/or emotional) relationship. If the nature of this bond gets stabilized from M’s point of view, from F’s point of view or from the point of view of both, we say that a Tie-Up (TU) has occurred. If the bond only concerns F, we speak of a Female Tie-Up (F-TU); if it only concerns M, we speak of a Male Tie-Up (M-TU). If a bond has been created for both, we speak of a Double Tie-Up (D-TU)—see
Figure 2.
AAs are psychological structures that build up attraction for and to the opposite sex. They are defined as active in that they elicit behaviors that are aimed at seducing partners, and that, in particular, are aimed at “tying up” potential mates. AAs are regulated by a psychological reward mechanism, which is connected to their mere usage. The optimal reward is reached when the partner’s TU is attained, for either M or F. In particular, M’s reward is optimal when the action of M-AA causes F to tie-up, i.e., F-TU is created, and vice versa for F-AA when M is caused to be tied-up and M-TU is created.
A key feature of our approach lies in the asymmetric characterization of the nature of AAs and RAs in the two sexes. For M, AA corresponds to M’s sexual dimension (see
Figure 3). M can be regarded as sexually active in that his active strategy toward the other sex is deployed on a sexual,
i.e., physical level, and is accompanied by physical correlates of sexual arousal [
100] and primarily aimed at genital contact [
101]. A component of M-AA (sexual) reward is pride for sexual performance: M’s ability in the usage of his AA results in a sexual seduction of F, which at its best outcome causes F to be tied-up (F-TU). M’s reward is increasing in the level of F’s outward display of sexual availability response, as well as in the effective willingness to engage in a sexual intercourse.
For F, AA corresponds to F’s emotional dimension. F elaborates her attraction toward the opposite sex from a mental-emotional viewpoint. F’s active seductive approach has an emotional nature, and reward is accompanied by physical correlates of emotional activation (e.g., [
102]). In female sexual function, the emotional and sexual dimensions are deeply connected [
103], and female implicit sexual desire increases once being romantically stimulated, whereas the same does not occur to men [
104]. The reason of such a difference is that women associate with the romantic situation, and in particular to the first physical contacts aimed at the creation of a possible intimacy, a sexual connotation, while men on the contrary in such a situation are led to momentarily inhibit their sexual response. In both cases, as we will see, these reactions are finalized to each one carrying out the respective compatibility test for the sake of the future formation of the couple. For F, causing M to tie-up (M-TU) as a consequence of the action of F-AA means involving M in her mental process of emotional bonding construal. When M gets bonded in turn, the reward of F reaches its peak, as this signals that F-AA has been able to cause the Tie-Up of M. It is significant to note that Ms’ preference for sexual over-romantic stimuli (and the other way round for Fs) does not depend on pleasantness of the association—in fact, in terms of pleasantness,
both Ms and Fs associate romantic stimuli more strongly with the pleasant condition than sexual ones, which further strengthens the idea of a biological programming of the content of each sex’s AAs [
105]. Additionally, from the cognitive point of view, the structure of conceptual associations about sex-related matters is distinctively more focused on the emotional dimension for women and on the physical one for men [
106], and the corresponding sex-related differences in word frequency and vocabulary use are clearly reflected in communication styles [
107]. However, concluding from the previous discussion that Fs are emotionally oriented whereas Ms are sexually oriented would be misleading. For both men and women, the sexual and emotional dimensions are equally central for mating purposes. What changes substantially is their relative role and functioning. To understand this better, we now turn to a closer examination of RAs.
RAs are passive psychological structures where the Tie-Up happens or not, that is, where F gets sexually seduced and/or M gets emotionally involved. RAs test effective compatibility with mates. For this reason, it is not granted that RAs are activated, even when the opposite-sex potential partner actively engages the subject to this purpose. AAs command more personal (and social) awareness than RAs, so that RAs are effectively perceived only when they are successfully activated by the opposite-sex partner. The reason is that AAs function as safety systems in that they guarantee the perpetuation of the species, even if they do not manage to activate RAs, and become thus potentially conducive to a lasting mating. RAs, on the contrary, are mating stabilizers, which come into function when TU-oriented stimulation is effective, and their activation peaks when the TU takes place.
For M, RA has an emotional nature. M gets tied-up by F on a mental-emotional level, and is seduced in his imagination and feelings. M gets tied-up to F only if she is able to involve him emotionally. Otherwise, once the sexual intercourse has taken place and an optimal reward has been reached, M walks away from the relationship. M-RA has the task of checking the psychological compatibility with the female partner, as well as the right level of complementarity in pursuing joint couple goals such as child rearing, with the consequent evolutionary benefits.
For F, RA has a sexual nature, but its physical correlate is not limited to the genital area [
108]. A wide range of cues, including subliminal ones [
109], can generate a significant activation of RA [
110]. RA has a passive function; thus, the fact that F’s sexuality resides in F-RA tends to (erroneously) characterize F as ‘sexually passive’ when compared to M’s AA-driven sexual initiative. F-TU is therefore the outcome of physical contact with potential sexual implications. If F’s physical interaction with M is rewarding, the conditions for F-TU will be created; otherwise, F will divert attention toward other potential partners. F-RA is devoted to checking the biological compatibility with the M potential partner. In evolutionary terms, the role of F-RA is the screening of the appropriateness of M’s genetic endowment; thus, as one could expect, for Fs the spectrum of motivational factors that determine sexual satisfaction is wider and more heterogeneous than for men, as the sexual dimension plays a more relevant role in mate selection than for men [
111]. It would be interesting to test whether the opposite holds for emotional satisfaction,
i.e., satisfaction related to the dimension that is more relevant for M’s mate selection. To our knowledge, no results in this regard are available as yet, but this is an interesting subject for future inquiry. To sum up, when RA is stimulated (sexually for F, emotionally for M), a potential interest is sparked for the opposite-sex subject who has performed the activation. Humans have evolved special abilities to recognize signs of romantic interest from others, and such abilities extend cross-culturally [
112], although other research provides evidence of higher accuracy in detecting a
lack of romantic interest than its actual presence [
113]. Even minimal stimuli might function in principle as an activation. For instance, F could be stimulated by a visual excitement without any close contact, an olfactory stimulus such as pheromone signals, or a casual touch entailing an energetic perception under the form of body heat, or vocal features such as modulation, tone, frequency, rhythm, vigor, and so on. Such signals, before they intensify or cumulate, anticipate and prepare F-RA to the biological compatibility test. For M, instead, a typical effective stimulation of M-RA might take the form of (non-sexual) curiosity toward an opposite-sex subject, but equally effective might be strong emotional states such as disappointment, scorn, or even hatred, as far as they manage to convey attention toward F, whether positive or negative in that all that is needed is the emotional drive that sparks the psychological test to be carried out by M. The more M considers F in non-sexual terms, the more concrete the possibility of M-TU becomes. M-RA might be stimulated even by a gesture, a posture, a way of walking or of addressing others by an opposite-sex subject, or by emotional cues such as a smile, a laughter, or an uncommon or intriguing situation. In other cases, similarities with a previous situation, or associations with another opposite-sex subject with which an emotional bond was once established, might also work. What drives attention is affinity, in addition to perception of commonality of interests and passions. For some male personalities, M-RA might also be activated by a challenge that creates a clash of opposite wills, or by a mystery that arouses curiosity. M’s psychological state and emotional context have a role. A refusal of attention by F may induce a psychological interest in M, with a consequent activation of M-RA as the effect of frustration. Generally, for both M and F, testing mode is favored by the absence or deterioration of TU.
It is important to point out here the complex interplay between two different, interacting oppositions in the functioning of AAs and RAs: active/passive vs. conscious/unconscious. In particular, identifying active character with consciousness and passive character with unconsciousness would not be accurate. RAs, given their passive character, are often operating below the level of consciousness, but they can become the object of conscious attention, as, for example, when someone realizes s/he has become tied-up to another subject. AAs, on the contrary, given their active character are often functioning consciously, but they could also operate unconsciously in certain conditions, as, for example, when one unintentionally sends signals (e.g., staring at someone) that others can mistakenly interpret as evidence of romantic interest. Therefore, all possible combinations of conscious/unconscious and active/passive modes of functioning of AAs and RAs are possible. We will return on this point later on, when the full dynamic unfolding of the interaction between AAs and RAs will be presented.
3.2. M-TU vs. F-TU
TU is a psychological state where an emotional bond toward an opposite-sex individual is created with a potential purpose of long-term mating. If the bond is solid enough, the subject is tied-up, i.e., is constrained into the relationship until some major force or perturbing event intervenes. In F, this bond is mediated by the physical response of F-RA; in M, by the emotional response of M-RA. TU always contemplates a passive and an active element—one player who gets tied-up, and another player that elicits the Tie-Up. TU takes place when the subject’s AA successfully stimulates the opposite-sex’s partner RA. If this does not happen, TU fails.
Stable mating occurs when each partner gets tied-up to the other, and a D-TU occurs (
Figure 2 above). Stable mating thus calls for two distinct TUs: one where F is active and M passive, and one where M is active and F passive. If only one TU occurs, then one of the two partners is not tied-up, and as a consequence the mating is not stable and poses a dissolution threat to the couple, if previously formed (
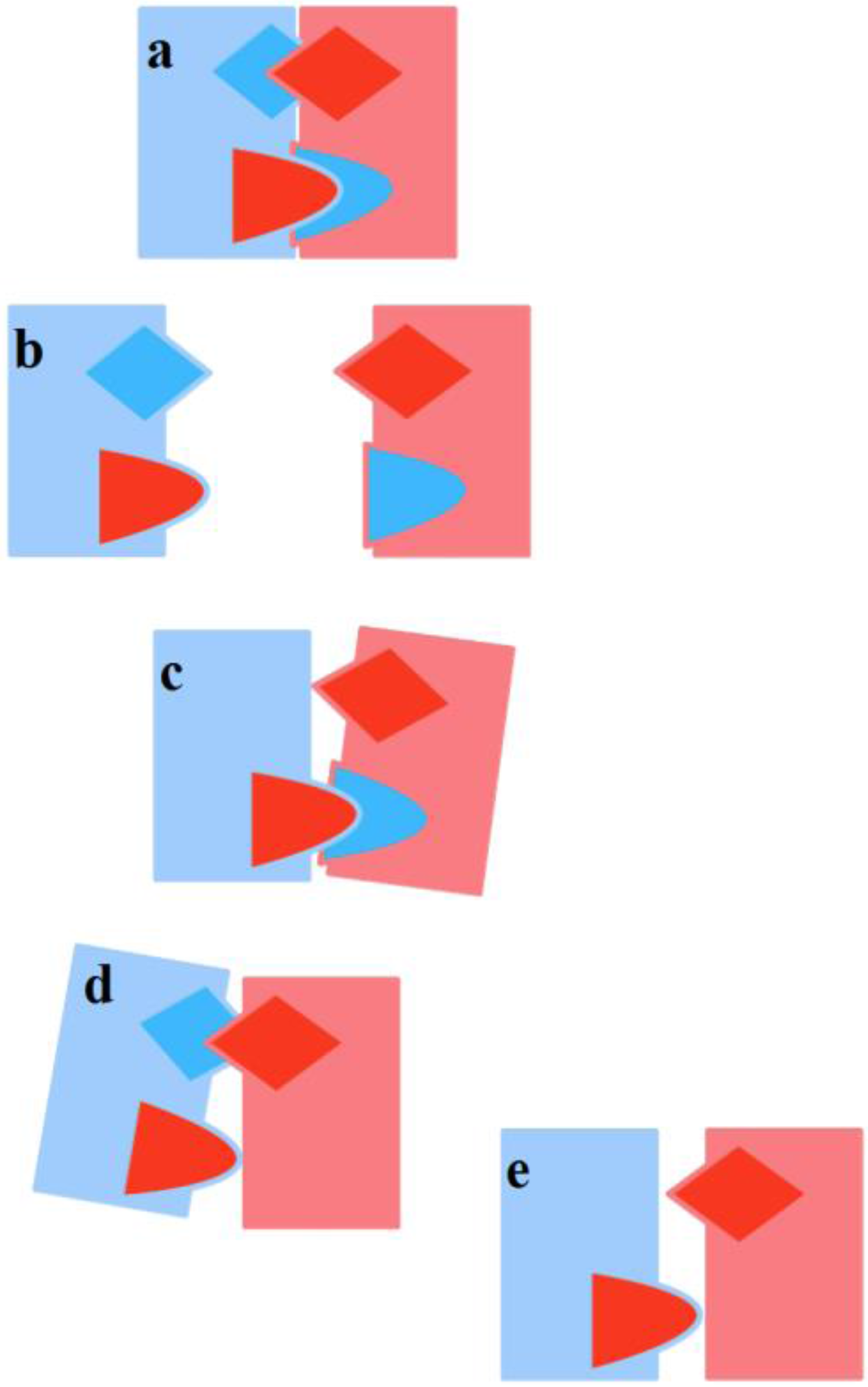
Figure 4). When, instead, in a previously stable mating, one of the TUs deteriorates for some reason, the couple stability is threatened, and once again there is a risk of dissolution.
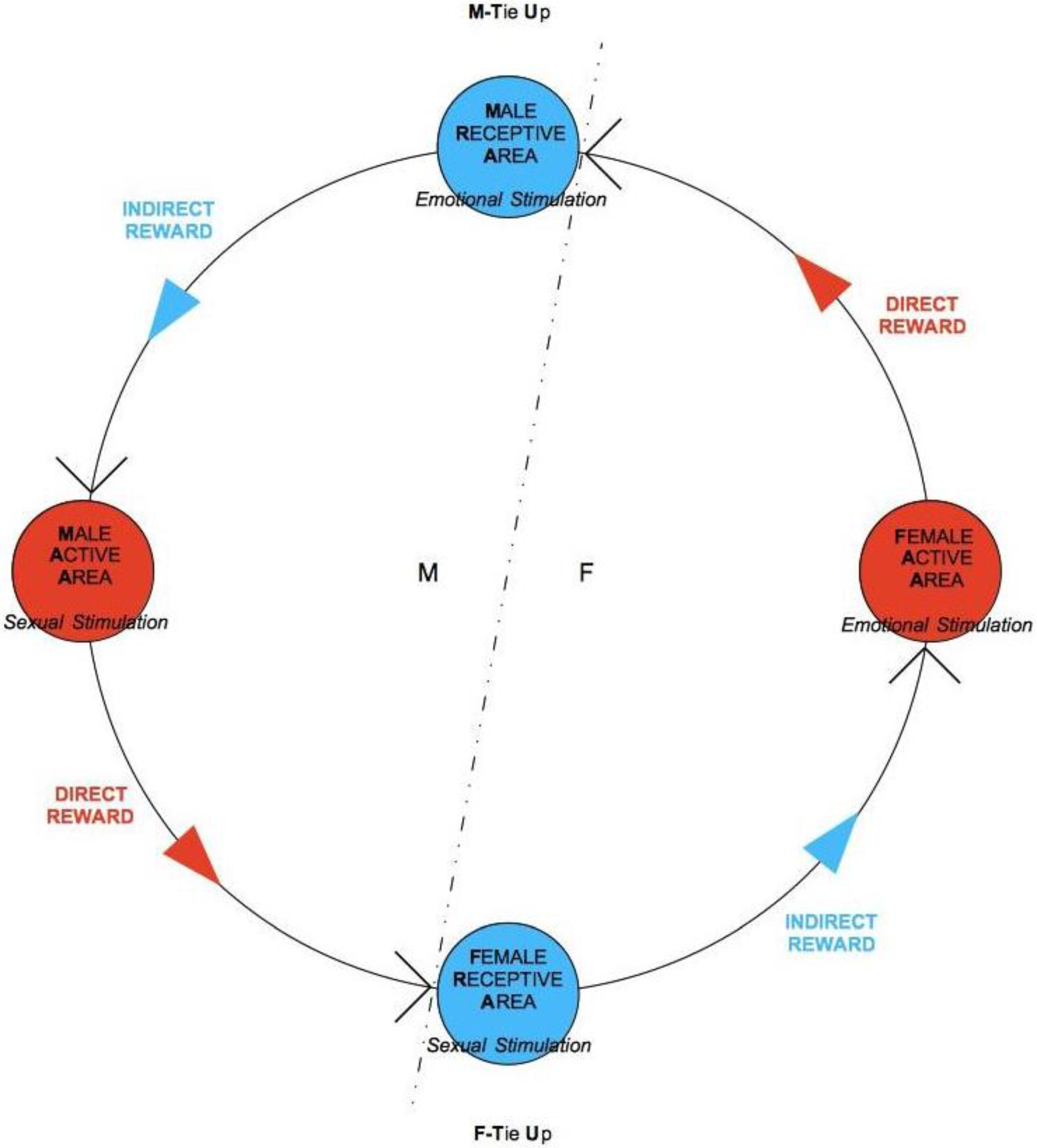
When RA is successfully stimulated by the other-sex partner’s action, as a result of the consequent positive response, there is a feedback toward the acting partner’s AA itself. Such positive response leads the active subject to intensify his/her action, and the process starts to self-catalyze. It must be pointed out that the positive signal arrives from RA in an indirect way, RA being a passive component of the subject who needs the active one, AA, to communicate. An effectively stimulated RA, thus, will send a positive signal to the same subject’s AA, which will be activated to respond to the opposite-sex partner. When RA’s response signal is elaborated by the AA of the same subject (who was originally passive), the subject now becomes active, and starts in turn to stimulate the partner who receives the new signal, this time passively, through his/her own RA. In this way, a flow of transmission/reception of signals is created that goes through AAs and RAs of both subjects, being progressively amplified at every passage, thus favoring the occurrence of the compatibility tests and the formation of the TUs and, due to their occurrence, feeding and strengthening them.
To sum up, the process requires the AA to send signals to the RA of the opposite-sex partner and that the RA directly communicates only with the same subject’s AA. In this way, each subject is passive in his/her own RA and active in his/her own AA, and the alternation between the two phases determines a communication flow, anti-clockwise and cyclical, that self-catalyzes (see
Figure 5). It may happen that, despite the correct and repeated occurrence of the transmission/reception flow, a one-sided TU by one subject only is determined and persists. Moreover, if there are sufficient incentives for each one to keep the relationship alive anyway, the result may be an opposite-sex friendship. If a F-TU occurs, this may lead to a (short-term) sexual friendship, without the formation of a proper couple in that M is not tied-up (see
Figure 4c above). A sexual friendship may be frustrating for F-AA. On the contrary, an emotional friendship where only M-TU occurs and F is not tied-up may frustrate M-AA (see
Figure 4d above).
F-TU is sexual: In this type of TU, F gets bonded to M, through her being tied-up at F-RA, which has a sexual nature, as a consequence of the active stimulation by M. F-TU can only happen when F-RA has successfully tested M’s biological compatibility. As the whole female body reacts sensitively to the stimulation of her RA, F starts to test M’s compatibility from the very first physical and chemical contacts (smell, taste through kissing, bodily energy through touch). In this way, F can carry out the screening without having to undergo an excessive number of sexual intercourses (in the limit, even none) with different potential mates, minimizing the risk of inefficient selection of genetic endowment. For instance, in some societies where women are sexually secluded before marriage, dancing may represent a relatively effective way to carry out the biological compatibility test in the absence of sexual intercourse [
114].
Significantly, women consistently manifest more negative feelings for sex than men both at the implicit and explicit level [
115]; moreover, although women display a relatively more liberal attitude toward sexual invitations in low-risk social environments then in high-risk ones, a significantly higher propensity of men to casual sex still remains in the most favorable conditions [
116]. Men also display a more positive affective response to sexual rejection than women [
117]. F can be mentally and emotionally attracted to M, but physical attraction is not consequential if the biological compatibility test is not successfully passed. Thus, M can emotionally seduce F without causing F-TU to occur. This is the case where F is only interested in a friendship relationship with M, as she feels rewarded in her F-AA by the signal coming from M, without, however, having an impact upon F-RA.
M-TU is emotional: In this type of TU, M is the passive player who gets tied-up in M-RA by the seductive action of F. This form of mental seduction compounds with the physical one, although the two components may decouple to ensure that M-AA keeps on functioning: M can be sexually attracted to F, but the emotional compatibility with F cannot be taken for granted. If the two types of seduction are decoupled, TU does not take place for M: M-AA is strongly stimulated, but M-RA is not. Sperm is distributed anyway if sexual intercourse takes place, thus guaranteeing a “mechanical” perpetuation of the species, but it is not granted that a stable couple that would secure safer child rearing will form. F is therefore able to seduce both emotionally and sexually, but it is only the emotional seduction that is able to induce M-TU in M-RA. M, on the other hand, can also be stimulated in M-AA as a consequence of a merely emotional stimulation in that the emotional M-TU affects M-AA anyway. This explains the difficulties in-built in an opposite-sex friendship: M gets emotionally tied-up; thus, as a consequence, it is possible that his sexual M-AA gets stimulated in turn. Only if M is already stably tied-up in another couple relationship, or if F has a very low short-term mate value for M, does it become possible for M to take part in the friendship with a limited level of frustration of M-AA.
3.3. The Tie-Up Cycle
The interplay between M and F’s AAs and RAs determines what we call the Tie-Up Cycle (TU-C). To understand how it takes place, it is important to distinguish two different kinds of rewards to M and F actions: direct (active) reward and indirect (passive) reward. Reward is what sparks TU-C and keeps it working. Direct reward is originated in AA and depends on its functioning. The more used, well operating, and successful AA is in eliciting (and feeding) the other-sex partner’s TU, the larger the direct reward. Indirect reward is again perceived in AA but is the consequence of a stimulation of RA and thus comes from RA. Indirect reward thus induces a further stimulation of AA and, in particular, desire for a more direct reward. To appreciate the different nature of direct
vs. indirect rewards, one can think of them as rewards linked to carrying out an action (e.g., the dog’s satisfaction from grabbing the thrown stick and bringing it back)
vs. rewards linked to passively enjoying a benefit (e.g., the dog’s satisfaction from being offered biscuits and eating them), respectively. Skillful modulation of active
vs. passive responses to mating-related situations is crucial in the dynamic unfolding of TU-C, and the two different kinds of rewards may be thought of as evolved, complementary psychological incentives that regulate mating interaction. One can thus regard our approach as a first hint of a behavioral theory to make sense of the neurophysiology of mating-related rewards, and which of course calls for substantial further development [
118]. Some preliminary remarks in this direction may be found in
Section 4 below. It is also useful to remark that the specific content of both direct and indirect rewards is subject to cultural variation: What can be perceived as a signal of emotional attention or of sexual interest that is conducive to a certain kind of reward clearly varies in different socio-cultural settings. However, the functioning of rewards in the context of the mating interaction is largely independent of the form or content of the signal that caused the reward.
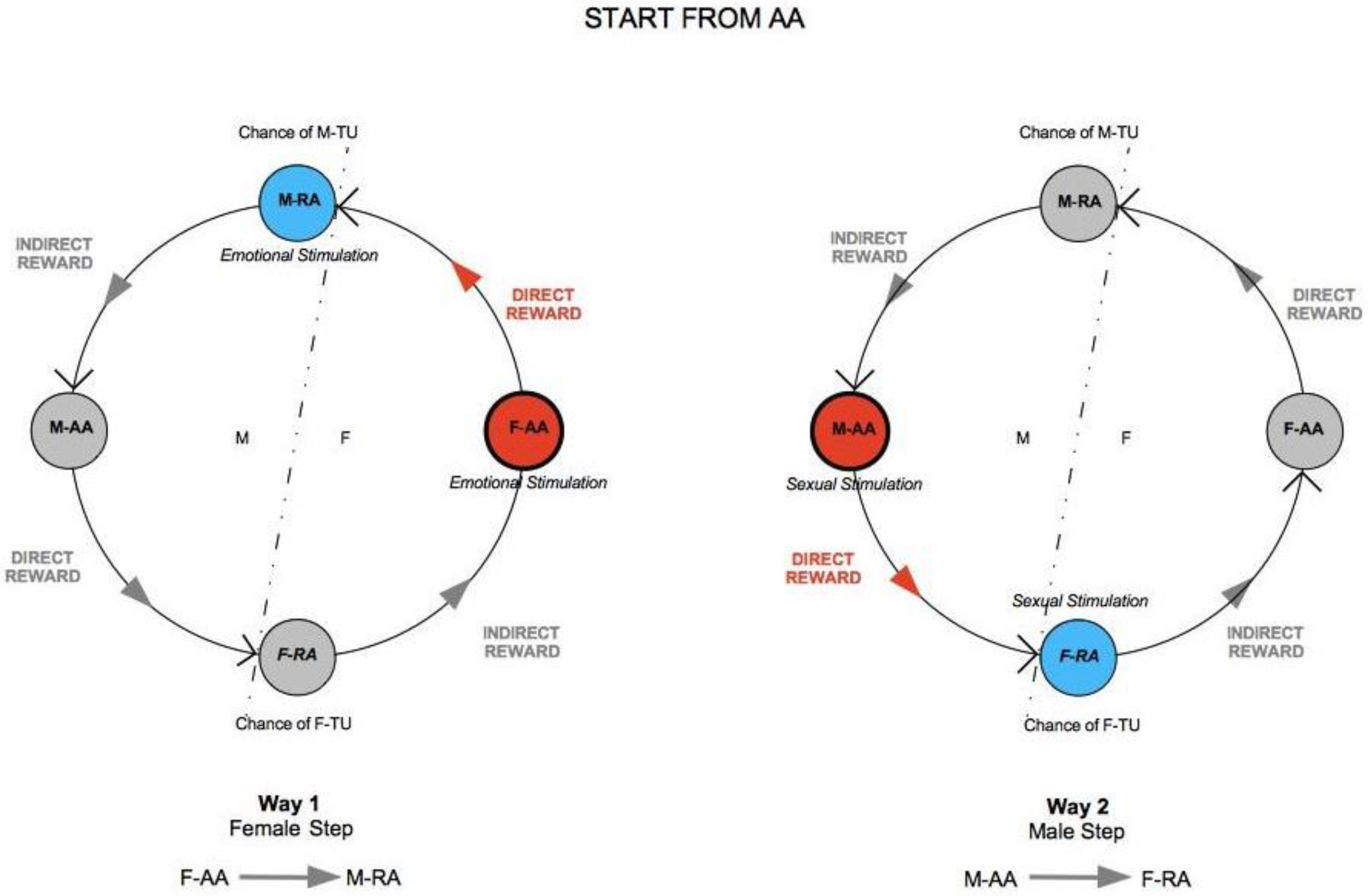
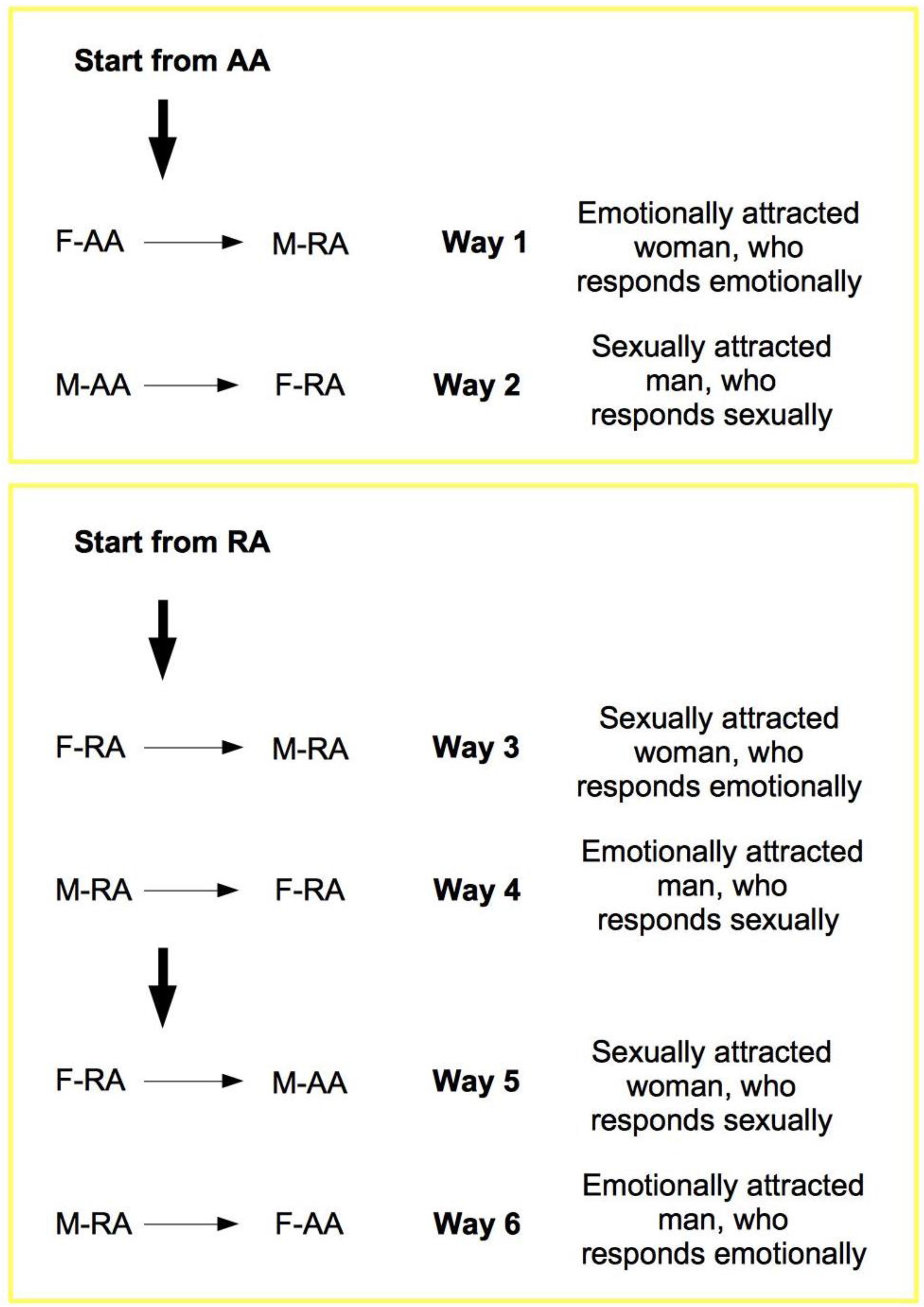
The elements from the previous discussion can now be summarized in the TU-C diagram shown in
Figure 6, which presents the M/F-AA/RA interaction in terms of a specific cycle with a typical structure. In the cycle, a subject’s AA is always linked to the opposite-sex partner’s RA, whereas a subject’s RA is always linked to the same subject’s AA—two rules which are implicit in the very definitions of AA and RA. This amounts to requiring that the cycle proceeds anti-clockwise only. The cycle can moreover be started in principle at any point (although with different consequences, as we will discuss below), and, in order to successfully build up, it must not be interrupted. A full cycle is a precondition for a D-TU, and more cycles correspond to the progressive consolidation of the couple relationship. How many cycles are actually needed for a stable D-TU to happen depends on a variety of factors. If RA is activated, and the potential mate shows an interest as well, one or more test TU-Cs can be tried out. For each full iteration, the possibility of a TU for at least one of the subjects substantially increases. Activations of M/F-AA/RA may occur at different levels of intensity, response, and reward, and interactions themselves may occur at different speeds that depend on the actual frequency of stimulation. In particularly intense situations, several TU-Cs may take place very quickly, and repeated stimulation causes and strengthens TUs as a consequence of the pattern of activation of RAs. However, a slow movement along a TU-C may present advantages in terms of reinforcement of the desire for reward. Both patterns probably have an optimal interval of contextual conditions in which they work best. The varying intensities of interaction and contextual situations in which the cycle happens may cause biological/psychological testing to happen already at the very first iteration, or in the repeated, quick kick-off iterations of TU-C and, if successful, may bring about TU. In other cases, several TU-C iterations might be called for just to carry out the test, which always precedes TU, if there are resistance factors or obstacles at work, both by the subjects themselves and in the external environment.
The dynamic structure of TU-C allows us to illustrate the basic difference between the active/passive vs. conscious/unconscious functioning of AAs and RAs. M could, for example, get tied-up to a F with apparent low mate value insofar as the emotional compatibility test has been unconsciously carried out by M and successfully passed by F. M could therefore inexplicably feel incapable of drawing attention away from F despite his perception of F’s low mate value. At this point, RA is functioning in its unconscious mode. Once M realizes he has become tied-up, however, the functioning of RA moves to the conscious level; however, because of RA’s passive character, there is nothing that M can do to break the Tie-Up insofar as his RA continues to be effectively stimulated by F. Conversely, F could, for example, unconsciously stimulate M’s RA with some unintentional attitude (e.g., a certain way of speaking or paying attention). However, once she realizes that M has been stimulated and reacts accordingly, she could easily refrain from going on with the stimulation if she is not interested in tying-up M. F’s stimulation of M came from F-AA; thus, the passage from the unconscious to the conscious level is all it takes to put the stimulus under control and to block it if needed. A conscious functioning of one’s AA thus corresponds to a situation of full control in the context of the mating interaction. An unconscious functioning of the AA corresponds to a redeemable failure of control. A conscious functioning of one’s RA corresponds to an awareness of a lack of control. Finally, an unconscious functioning of one’s RA corresponds to exposure to external control. If one-sided, it leaves the subject susceptible to possible manipulation.
One can read couple dynamics as a sequence of iterations of the cycle so that interaction among partners may further consolidate D-TU or threaten its stability. If D-TU fails to be achieved or is broken at some point, the cycle dies down. Moreover, there is no consequentiality between TUs: They occur separately and therefore need not happen concurrently, but may even happen at long distances in time from one another. Therefore, the fact that a TU occurs never guarantees that D-TU will eventually take place as well. If D-TU does not occur, TU-C does not unfold effectively. However, there is a possibility that one of the two subjects simulates his/her own TU, pretending s/he is tied-up. In this case, TU-C kicks off but does not really work: Maintaining the simulation calls for continued effort, which subtracts energy from TU-C. The issue with the subject simulating TU is that his/her RA is not really activated; thus, s/he goes through TU-C by benefit only of direct reward in that RA does not yield any indirect reward. Although direct reward is attractive for the subjects in that it confirms their sex role, the true engine of TU-C, the one that guarantees its self-catalytic regeneration, is the indirect reward that further feeds the subjects’ AAs.
One interesting vantage point to understand the interaction dynamics between M and F is the moment in which sexual intercourse takes place so as to evaluate possible evolution scenarios on the basis of TU-C. Let us choose this reference point to fix ideas and to better illustrate the structure and properties of TU-C. The direction of movement along TU-C is what determines the behaviors of M and F after the sexual intercourse. If sex has been rewarding, F-RA gets stimulated and F-TU occurs (bottom circle in the cycle in
Figure 6). The sexual activation that results stimulates F-AA by means of an
indirect (passive) reward, because F-TU has occurred so that sexual stimulation also becomes emotional stimulation, and F-AA is prompted to generate further direct (active) reward to the benefit of M, who has been the cause of F’s indirect reward. If, however, M-TU has not occurred (top circle in the cycle), F-AA gets frustrated, TU-C is interrupted, and the couple is threatened to dissolve.
The anti-clockwise direction of movement of TU-C tells us that, for M, sexual stimulation is by no means conducive to emotional stimulation as for F in that the latter is antecedent to the former in the cycle’s flow of movement. Therefore, if M gets sexually stimulated, emotional M-TU is far from granted, while at the same time not precluding the possibility of sexual intercourse for M. In the case of M, it is emotional stimulation that transforms into sexual stimulation in the TU-C, and not vice versa. For F, likewise, emotional stimulation does not spontaneously evolve into sexual stimulation, as it is still necessary that the cycle’s flow of movement passes through M and his sexual initiative. It is, in other words, necessary that the emotionally stimulated F has the opportunity to test M’s biological compatibility, that is, the quality of the genetic endowment that would be transmitted with impregnation; thus, sexual stimulation exclusively depends upon this test, which takes place in F-RA, through a physical-chemical-energetic sampling of M which does not primarily concern M’s genitals. The kind of test taking place in M-RA instead is related to psychological (and not biological) compatibility; thus, such a test does not influence the possibility of sexual intercourse that can occur for M without the need of emotional involvement.
If M, along TU-C, moves from M-AA, he will proceed toward an increase of his direct reward, of a sexual nature, totally ignoring indirect, emotional reward. If he will manage to elicit F-TU, his level of reward will reach the optimal level, and will inevitably plunge after the intercourse as a consequence of the attainment of the goal, not evolving into any other kind of reward, as the indirect one is behind his shoulders along TU-C, and not in front of him. This is why M, having depleted any form of further reward, will break up TU-C, leaving F to realize, along with a frustration of her F-AA, that no M-TU has occurred. If, instead, M takes the move from his M-RA, that is, when M-TU has occurred, thus experiencing an emotional stimulation, his indirect reward will evolve into a direct reward (circle on the left in
Figure 6), and emotional stimulation will turn into sexual stimulation and, if he manages to cause F-TU to occur, sexual intercourse; thus, F’s active stimulation will bring about further stimulation of M-RA with a consequent indirect reward that will keep the overall level of reward high even after the intercourse. The cycle closes up and becomes self-catalytic, and the premises for a stable couple are set.
For F, it is therefore always safer to enter TU-C from her F-AA in that if M does not tie-up, the inevitable frustration that results is easily solved as F is in turn not tied-up yet; it is therefore easier for her to step down the relationship and to look for other partners. If, instead, F-TU takes place via sexual intercourse to a M who is in turn not tied-up, the frustration of F-AA will be much stronger, and it will take time to solve it, as F is now emotionally linked to a M who refuses her. From M’s point of view, the situation is much different, as he can go through the sexual intercourse without having to be tied-up and without getting any frustration from this—and this guarantees female impregnation, even though a stable reproductive couple has not formed. The stable reproductive couple is, in fact, a higher-level relational arrangement that guarantees not only quantity, but also quality in child rearing, with an evolutionary advantage for offspring reared by both parents. M’s test of psychological compatibility then serves to assess F’s emotional intelligence and personality traits—two crucial aspects of a fully successful child rearing.
F-TU may occur independently of the sexual intercourse: Once the biological compatibility test is completed, if there are no obstacles in the first iterations of TU-C, that is, if rewards are correctly activated and M’s emotional and sexual responses are also satisfactory in terms of F’s active reward, with the unfolding of TU-C, sooner or later, F-TU occurs. However, if the sexual intercourse takes place, if there is biological compatibility, and if an optimal level of indirect reward is generated, F-TU is guaranteed. M-TU, on the other hand, is more likely to occur without sexual intercourse because, as pointed out earlier, if TU-C is started off with intercourse without an involvement of M-RA, M will be subsequently induced to leave TU-C to compensate for his own sudden drop in the after-peak direct reward. This is a typical problem of the earlier phases of TU-C. However, if TU-C kicks off and D-TU is established, there will no longer be a drop in reward for M within the cycle in that the indirect reward generated by M-RA will compensate and ensure a fair level of overall reward for M, stimulating M-AA to generate more reward for F.
The different implications of the sexual intercourse from the viewpoint of F vs. M thus depend on the fact that the intercourse fits into TU-C in a way that favors F-TU, but, on the contrary, plays against M-TU. M has thus an incentive toward pursuing sexual intercourse in the early stages of the interaction, whereas F has an incentive to postpone it. These two incentives are targeting two different adaptation problems: M’s drive ensures that biological reproduction may occur also in the absence of M-TU, whereas F’s conduct has the effect of restricting sexual contacts to the Ms who meet the criteria for effective child rearing, as revealed by the fact that they are willing to cooperate with F in the TU-C kickoff instead of practicing sex without attachment. By going through the first iterations of TU-C without sexual intercourse happening, M and F gradually build up their way to their peak levels of direct vs. indirect reward, respectively, and by so doing they have the possibility of effectively synchronizing their cooperative mating strategies, while at the same time being pulled in by the expectation of higher rewards in the subsequent steps. On the other hand, when D-TU has occurred, there is no need for F to strategically limit sexual intercourse any longer, and there is no reason of impatience for M in achieving it. In other words, one can regard the positioning of the sexual intercourse with respect to F-TU vs. M-TU in a proper TU-C dynamics as a sophisticated, evolved mechanism for optimal mate selection by F, which balances the evolutionary necessity of M’s attachment-free sex as a no-brainer hedging strategy against the threat of demographic decline, and, at the same time, for the sake of M’s partner selection, who is able to check the strategic-emotional intelligence of F as a psychologically (and evolutionarily) useful resource for an effective joint child rearing.
In view of the above, it is possible to better understand the role of generally acknowledged sources of mate attractiveness, namely physical (beauty) and material (wealth, power, etcetera) resourcefulness. Being beautiful, rich or powerful in itself cannot cause a TU, as the latter is regulated by biological (for F) or emotional (for M) compatibility tests, respectively, whose outcome is neither mechanically related to beauty nor to intelligence or social prestige. However, certainly resourcefulness makes a subject more attractive as a possible mate, as the carrier of a highly desirable asset with a clear reproductive value. The main effect of being rich, powerful, or beautiful is therefore that of dramatically increasing the probability that the appropriate conditions are created to be tested out by the other party in terms of close physical interaction or deeper psychological probing, according to cases. Independently of the test’s outcome, the other party could be strongly incentivized to tie-up the resourceful subject for instrumental reasons and therefore to simulate one’s own TU, relying upon direct reward as the only real incentive. If, however, the test is successful, the party attempting to tie-up the resourceful subject will enjoy an indirect reward as well, and the mating interaction will at least partially lose its instrumental character, improving the chances of a successful TU-C and of a stable mating.
3.4. M vs. F Rewards
According to sentimental common sense, the partner who suffers the most in a couple is the one who is most attached. In fact, from what we have learned from our previous analysis, the partner who suffers the most is likely to be the one who is tied-up to someone who pretends to be tied-up, but in fact is not. The reason is that, if a partner is not tied-up, once s/he has got the desired reward, s/he will show very little concern for the reward of the other. A failed TU in a couple may be in turn the response to a specific constellation of incentives. Some men, for instance, systematically avoid hooking up with a woman, as they expect a progressive consummation of direct reward through repeated sexual intercourse, and are unwilling to face the risky prospects of an emotional commitment. Looking for another partner is a possible solution, but it is once again necessary not to tie-up, i.e., not establishing emotional bonds with the new F as well. The ideal target for attachment-free sex is a sexually attractive F who, however, does not pair well with M’s personality and intimate world and possibly lacking strategic mating intelligence, so that it will be easier to walk away once M’s direct reward takes the downward slope, without having to overcome sophisticated mate retention tactics by F, or to resist the temptation of settling in with an emotionally rewarding mate. For M, therefore, the indirect, emotional reward coming from M-RA may look risky and unattractive if M is convinced that life satisfaction depends upon unrestricted sexual choice and lack of couple-related responsibility.
Another common belief, that is, that women are more prone to be bonded to men than the other way round, illustrates a constellation of incentives that is the mirror image of the former, in terms of a failed TU, this time from F’s side: Social pressures, the biological clock, or the achievement of economic security urge them to find a mate no matter what and thus, to keep biologically undemanding standards for mate choice to prioritize instrumental mating. Occurrence of F-TU is therefore not a precondition. However, men who marry a non-tied-up woman face no less trouble than women marrying a non-tied-up man. What is at stake for M is not only his direct reward, but his very masculine identity: Living together without being tied-up entails a very high temptation to frustrate the partner, and this is typically what will happen during marital conflict. If M cares for having children and a family without being exposed to frustration on a daily basis in both his direct and indirect reward, it is not enough to look for a physically attractive woman, but it is necessary that she is also emotionally intelligent, and well-matched in terms of feelings and personality, world views, and ideals, so that indirect rewards can be generated for M. Moreover, F will have to be tied-up in turn so as to generate also direct (sexual) reward. A non-tied-up F is sexually detached and could easily deny, or more generally strategically manipulate, sexual access by M. The existence of a D-TU is therefore a precondition for a TU-C where the continued exchange of rewards between M and F will make marital life satisfactory and desirable.
Notice how, despite the strong reproductive rationality of direct and indirect rewards, mating interaction leaves space for cultural variation. For women, the test has a biological nature, but the conditions under which women can actually carry out the test are often strongly regulated, if not restricted, in ways that clearly depend on the socio-cultural environment. For men, the test has an emotional nature; therefore, the criteria with which psychological compatibility is defined and established are clearly culturally influenced. The limitation of women’s discretional power in mating choices observed in many cultures could therefore be regarded as a male-controlled form of limitation of the scope of the female compatibility test, whose content can be otherwise culturally manipulated only to a limited extent.
A sphere in which the interaction of M and F rewards lends itself to interesting exemplification is that of chivalric courtship. When M chivalrously courts F, he is feeding F’s direct reward. Here, courting is nothing but emotionally pleasing F, hoping for an active response from her. Courtship can thus be regarded as a persuasive technique of M, targeting F-AA. F is certainly pleased by it, also in the absence of any level of sexual involvement or even interest. Such kind of male courtship simulates, or is a concrete manifestation of, male romantic love. This is always gratifying for F in that it confirms the effective functioning of her F-AA, aimed at conquering M’s emotional attentions (not necessarily of the M actually courting her). Some direct reward for F occurs even if it is apparent that M’s courtship is not really aimed at sexual conquering, and this is because in this case courtship purely acts upon F-AA, and not upon F-RA. F perceives a discreet courting from M who appreciates femininity in general, as a gentle, refined form of attention and respect. Chivalrous courting could thus also be directed at women with very low mating value for M (e.g., a very elder woman) while still making sense for him as a way to test his courtship abilities.
If M expresses sexual attention instead, with explicit appreciations and no courtship, no direct reward will be generated for F, and not because F is not interested in sex, but because such an un-mediated approach does not gratify her F-AA, which has an emotional nature. M’s crude sexual approach clearly signals not only no real propensity to tie-up, but an exclusive desire to gratify his own M-AA, totally ignoring F-AA: In other words, a plain intention of sexual exploitation. Frustrating F’s direct reward, in the absence of any F-TU, leads to an abrupt interruption of the flow in TU-C. F ceases to show interest toward M, who signals he is not orientated toward M-TU and thus rejects him all the more in sexual terms, even if he turned out to be biologically compatible. If, instead, M shows an ability to generate a direct reward for F, F will in turn activate F-AA so as to stimulate him and to eventually arrive, through the steps of TU-C, to test his biological characteristics.
In case of F’s affective insecurity and/or low self-appreciation, sexual attention from M can be misunderstood for emotional interest, leading to classical sexual exploitation dynamics. In this case, M’s crude sexual request generates an “anomalous” direct reward for F. This sparks a rather problematic TU-C, which will lead with difficulty to a stable couple. Affective insecurity might in particular make F willing to sexually comply with the requests of a non-tied-up M simply to get some manipulative emotional, or even merely sexual, attention.
In the absence of self- or hetero-directed manipulative distortions, we can claim in general that F’s direct reward sparks an emotional interest toward M, and F’s indirect reward sparks sexual attraction toward M. The actual starting point of TU-C will depend on which area of F will be stimulated first: AA or RA. If F meets a physically attractive male, appreciating his posture, gesture, prowess, voice, smell, energy, touch, even merely through a simple, close handshake, her F-RA gets passively stimulated, and the reward that follows is an indirect one. The signs of an indirect reward for F are all the more evident in the early phases of her psycho-sexual development, when her capacity of strategic dissimulation is still limited: blushing, embarrassment, giggling, smiles, laughter, excitement—a typical attitude, for instance, of young fans of pop stars at their idols’ live acts. Their hysteria is the result of an indirect reward that reaches paroxysm. Indirect reward being passive, if there is no F-TU, it does not generate any frustration and is therefore psychologically innocuous for F. It is just F’s body that responds to a sensory stimulus, even a mere visual one, and F can decide to stop at this point and not to go for a direct reward. This is just the premise of a potential TU, and not an actual one in that the biological test has just started. This is what is commonly called an infatuation.
If, in general, the early phases of an interaction escalate to kissing, and if the experience is pleasing, F’s indirect reward reaches a first peak. For teenagers, kissing may actually cause TU, and it may be highly persuasive even for adult women as a very direct and effective way to sample M’s chemistry and energy. The more the indirect reward generated by the kiss, the more promptly TU-C will start. If kissing in itself will most likely not be enough to cause a TU, provided that it generates enough indirect reward for F, it will certainly stimulate F-AA, thus producing a direct reward. Beyond physical response, there will be an emotional response as well. F will begin to act emotionally, that is, actively, with respect to the M who kissed her, even by means of complex signals that may lead to significant strategic interaction. For instance, the face slap that M may receive in certain circumstances after the kiss is a concrete signal of an active emotional response by F and not necessarily with a negative valence. If F did not like the kiss, she would clearly show disgust and would try to escape; however, if M, say, gets simply slapped, the message may be more ambiguous, and it is, however, a signal that an indirect reward has reached F-AA. The impulse behind the slap may then possibly be a way for F to brake or interrupt a process that has generated some level, however small, of indirect reward. In this case, if M wants to take advantage of the kiss with the intent of getting closer to F, the best move is not to react to the physical offence and to stop the sexual approach immediately. Surrendering makes a winning move here in that it amplifies the overall reward for F as a sum of the indirect sexual reward from the kiss and of the direct emotional reward from realizing how her active response has proven effective.
The fact that there is a fundamental asymmetry between M and F with regard to the positioning of their sexual/emotional sphere into the AA/RA reflects, as briefly anticipated in
Section 3.1, different modes of management of the relationship between the two spheres in each sex. As to such differences, there are also substantial misunderstandings though, especially about the role of emotions in the female sexual response. The fact that women get sexually excited by a romantic situation (for instance, a movie scene) is not, as it is erroneously believed, the consequence of the activation of her emotional sphere, but on the contrary derives from the excitement of her RA due to the simulation (with the movie’s male lead character as the potential mate) of the test of biological compatibility that is connected to romantic physical contact. This is the reason why the actors who play the male lead character in romantic movies (from Rodolfo Valentino to Leonardo DiCaprio) turn into sex symbols; the female audience gets potentially tied-up to them. The excitement of the F-RA is immediately processed by the F-AA, which receives in turn a considerable increase of its direct reward, thus generating the emotional response, which, however,
follows the sexual one. Thus, contrary to what is commonly believed, the driver of mating-oriented attraction for women has a
sexual nature. In the case of the man, instead, the romantic situation excites his emotional RA, and the man thus interprets this situation as the simulation of a test of psychological compatibility with the partner (in this case, the female lead character of the movie) that calls for a momentary inhibition of his sexual response, which will possibly
follow the emotional one should the test be successful. Again, contrary to common beliefs, the driver of mating-oriented attraction for men has thus an
emotional nature. The romantic movie, being centered upon the creation of the conditions for the possible formation of a couple (and thus of a D-TU) is in other words designed in order to activate the RAs of both sexes, which will however react in opposite ways because of the different sphere of pertinence of the respective RAs. The consequence of such asymmetry is that, in the romantic situation, in the case of the woman, the rapid firing of the sexual response becomes synergetic to the emotional one that follows, whereas for the man the sexual response must be momentarily inhibited so that the emotional signal may become legible to him for the purpose of the possible formation of the couple.
If M’s direct reward calls for continued shots of confirmation about his masculinity, F’s direct reward calls for attention and consideration by M, who must show sensitivity toward F’s emotional world and, more generally, a genuine interest for her. Many Ms are able to simulate such interest without an actual TU, but in case an easy capitulation from F is not obtained, they may get easily discouraged, revealing their lack of TU and their mere appetite for a direct reward. The only possibility for F, at the beginning of a TU-C, to expose a manipulative tactic of this sort is thus to take time, that is, being able to go through several iterations of TU-C without conceding to sexual intercourse: An optimal mechanism to frustrate a sexually opportunistic M and to dissuade him from insisting further. For a tied-up M, instead, the same situation is less difficult to manage, as he can benefit anyway from the indirect reward deriving from the activation of his M-RA, that is, the gratification experienced from the emotional involvement with F. In this case, then, postponing the intercourse may even result in an amplification of future sexual pleasure, when F will become stably tied-up, thus generating an even greater direct reward for M. This request to wait, however stressing for him, further produces in the tied-up M, as already remarked, a reassurance as to having chosen a F with an appropriate level of emotional intelligence. If M is really tied-up, the resulting indirect reward will induce him to explore F’s psychological world in search of more emotional stimulation, and the interaction with F will then be able to generate further psychological and emotional advantage.
The well-known gender differences as to the sensitivity to sexual
vs. emotional forms of infidelity may thus also be read, in terms of M
vs. F rewards, as gender-specific sensitivity toward the kind of infidelity that directly threatens the partner’s TU, that is, that attacks the betrayed subject’s monopoly of the partner’s indirect reward: M’s emotional infidelity for F, and F’s sexual infidelity for M. It is interesting to remark that such differences are still distinctively found in societies with very high levels of gender egalitarianism like Norway [
119].
3.5. Frustration
The positioning of M-TU in M-RA explains how M can easily abandon F after having obtained his optimal level of direct reward. The positioning of F-TU in F-RA explains why some F bind themselves to some M who are totally unfit at the emotional level. Therefore, the M–F interaction may easily degenerate from the point of view of a stable mating. However, an effective strategy to achieve TU may be found in frustration. Frustrating the direct reward in an early phase of the M–F interaction leaves two possibilities open, depending on whether an indirect reward of some sort has already been generated or not, and the implications are different for M and F. When M is sexually attracted to F, his M-AA starts to be stimulated, that is, direct reward begins to be produced, which leads M to attempt a sexual approach. If, however, F refuses the approach, M’s direct reward gets frustrated. The two possibilities are then the following: M gives up and turns attention toward another F, or M insists. The difference lies in indirect reward, that is, in the actual starting point of TU-C. If M, even without being tied-up, has been stimulated in his M-RA by F, even unintentionally, some level of indirect reward (however small) will be generated for him. If direct reward thus gets coupled to indirect reward, the frustration of the former will induce an urge for the achievement and consolidation of the latter. It is as if frustration calls for a compensation via the type of available gratification that is more proximate to the frustrated one, which in this case is provided by indirect reward—or otherwise is sought for elsewhere.
Therefore, if M is intrigued by F, the denial of his desire will end up intensifying it, leading M to persist, and to seek emotional attention from F. In this way, frustration encourages further strategic effort by M to achieve his goal, temporary shifting attention from his own direct reward to that of F. M thus goes searching for an entrance way to F’s psychology, for an access key to her interiority in that he instinctively, and sometimes consciously, knows that stimulating F emotionally, raising her active interest, and thus aiming at her direct reward, provided that the indirect one seems blocked, amplifies his chances of mating with her. What M fails to realize, however, is that in so doing he also amplifies his own chances to get tied-up in turn, as in this way he cultivates his own indirect, passive reward. In practice, M gets exposed to F’s emotional fascination, in addition to the sexual one, and carries out his own compatibility test on F, which if successful will expose M to a further, increasing indirect reward, that is, to a possible M-TU.
The activation of M-RA, as a precondition for the TU, is thus for M the ideal starting point of a long-lasting TU-C in that the lack of indirect reward, if the direct one gets frustrated, causes M to lose any interest for F.
A minimum level of indirect reward achievable by M is, for instance, feeling courted and the object of female emotional attention as, at least, M will in this way feel pleased to arise an active interest in F. However, if F looks like she did not even notice M, to obtain any level of indirect reward it will not be enough for M that F be sexually attractive, but it will also be needed that F has noticeable qualities in terms of emotional fascination—for instance, a particular sense of style and elegance, or inspiring tenderness and sense of protection, or, to the opposite, displaying a challenging attitude or an unconventional personality, and so on. The mere display of such non-sexual qualities may cause in some M an indirect reward linked to psychological curiosity and to the propensity to experiment with mystery. Lack of sympathy, or a strong initial disappointment, or even hatred, provided that they cause a destabilization of M’s certainties, such as questioning his emotional equilibrium and at the same time prospecting a different, unexpected one, might in principle become the basis for M’s emotional entrapment. Whether this kind of circumstance may lead to a M-TU ultimately depends upon the context, partners’ personality traits, their personal histories, and the details of the interaction itself. M might be willing to defend himself from the possibility of a TU by developing some form of emotional insensitivity toward F, which might be due to an exclusive interest for short-term mating, but also reveal pathological personality traits (for instance, a narcissistic, Machiavellian, or psychopathic syndrome [
120]). In this latter case, the simulation of an interaction aiming at the TU might be brought forward through a purely instrumental psychological exploration to enable a more effective manipulation of F, also by means of sophisticated usage of the signals and strategies of attraction [
121,
122]. Frustration might be thus the first step of a risky manipulative strategic interaction for the opposite sex partner [
123].
Similar remarks to those concerning manipulative frustration can be made, in a mirror-like manner, for rewards. Here, manipulation does not amount to taking the reward away but, on the contrary, to instrumentally providing it to get something from the partner. One must consider that the effect upon TU-C of reward increases is no longer that of a suspension of the flow, but of a flow acceleration within the cycle, which will, however, become increasingly demanding for the manipulative partner and will inevitably imply for him some simulation effort. The effort to sustain an accelerated rhythm of iterations of TU-C, though they are not so pleasing for those with merely manipulative goals, may unveil with time the instrumental intent. If the ultimate goal is that of inflicting some suffering to the other person, as in the above-mentioned pathological cases, the partner of the manipulative subject will be exposed to sudden accelerations followed by equally sudden brakes of the TU-C flow. The manipulator will make use of rounds of reward combined with frustration, thus mixing up the effects of both, and making the interruption of the cycle even more painful in its abrupt reversal of the acceleration of the flow of rewards along TU-C. Unlike frustration, which becomes effective for manipulation purposes only if targeting a direct reward, a manipulative action that makes use of the partner’s reward increases is equally effective whether it targets direct or indirect reward.
As to F, in this case an effective strategy to achieve F-TU may also be found in frustration. One, however, must keep in mind the actual functioning of TU-C in that if F does not feel herself sexually considered by a M she is interested in and is therefore sexually frustrated, in her case resentment will be felt not on a merely sexual level, as it happens to M, but will be felt on an emotional level, whereas M will perceive himself as sexually inadequate, F will not feel herself emotionally appreciated. This occurs because, in TU-C, F’s sexual area lies in RA and not in AA as for M. Sexually frustrating F means interrupting the flow of female indirect reward so that the affected area will still be F-AA, which will register the overall decrease of its reward, depleted of its indirect component coming from RA, and reacts on its (pertinent) emotional level: F will feel frustrated in her aspiration to perceive herself as emotionally and not sexually desired.
It is now appropriate to ask why, in an initial phase of TU-C, we only focus upon frustration of direct reward, and not upon frustration of indirect reward. Indeed, frustration of indirect reward, both for F and M, in this delicate phase of the TU build-up, would amount to a sort of suicide for the subject who frustrates with the ultimate goal of kicking off TU-C in that in this case one would undermine from the start the possibility of TU itself by blocking the compatibility tests for both sexes. Sexually frustrating a F while aiming to seduce her sexually is obviously counterproductive, and likewise it is far from convenient, to psychologically conquer M, to highlight an apparent conflict of character or even worse an inadequacy in terms of intelligence and mental capacity, be it the partner’s or the subject’s. More generally, if F frustrates M’s indirect reward, this means that F is no longer emotionally stimulating M, that is, she becomes emotionally repelling—therefore, the only reward that F may now offer to M has a sexual nature and is devoid of emotional attachment, as it happens, for instance, in prostitution. If M frustrates F’s indirect reward, this is equally counterproductive in that he devaluates his own sexual profile, thus only becoming able to offer F an emotional reward close to the one that is characteristic of same-sex friendship. The momentary, targeted frustration of direct reward, instead, does not endanger TU in that it does not act upon RAs, but rather addresses the AAs of both sexes, stimulating their active response.
The strategy also remains the same as far as F is concerned: denying a desire only to intensify it. If one wants to use frustration to induce F-TU, it must be realized that, in this latter case, it is necessary to administer the signal more parsimoniously than for M, and this derives from the basic difference between M-AA and F-AA, which are the actual targets of frustration so as to carry out an adequate calibration of the intensity of the effect brought about by frustration. The sexual nature of M-AA requires that the F willing to frustrate M exerts a sharp, peremptory action, with unequivocal implications and matching the sexual energy to be countervailed with an opposite push of equal intensity. However, the emotional nature of F-AA calls instead for greater precision on M’s side so as to modulate a much more sensitive, susceptible response. The finer emotional sensitivity of women with respect to men [
124] may be interpreted in our context in terms of the association of the emotional dimension with women’s AA and with men’s RA. M will thus have to avoid any form of frustration that derives from tactics and behaviors that have been validated and are generally accepted in male-only relational environments (in which agonistic fight, real or symbolic, has its own evolutionary rationale), such as name calling or ironical mocking of the other. To emotionally frustrate F, then, a simple, momentary suspension of interest may be enough already.
The frustration of F-AA based upon a temporary suspension of interest is, for instance, a common strategy among M teenagers, sometimes as an involuntary consequence of affective insecurity in relating to attractive potential female partners. A typical scenario of interaction is the following: When F enters the room, M acts as if F were not there, not speaking to her and not even looking at her, or, in more strategically sophisticated cases, even courting other Fs. In fact, with these premises, the young M’s strategy might prove ineffective unless he did not previously display at least a minimum level of emotional involvement toward F, or attract her attention, for example, through a non-incidental eye contact, before carrying out the suspension. Without a preexisting emotional anchoring, a suspension becomes entirely meaningless. Once framed into an appropriate emotional context, though, the suspension strategy of the M teenager carries over practically unaltered to the adult age: After having intensely courted F, M momentarily ceases to show attention, as if any interest would have suddenly died down. F’s frustration would be immediate, and all the more so provided that F previously grew accustomed to the constant direct reward from the courtship.
Additionally, in this case, frustration of F’s direct reward will open up two different, alternative scenarios, depending upon the implications for F’s indirect reward. If F has no sexual interest in M, the frustration of F’s indirect reward by M will have no consequence other than provoking a short-lived disappointment in F, which will be soon wiped out by new opportunities for reward. If, on the other hand, F-RA was effectively stimulated during the previous courting by M, and F experienced some level of indirect reward (however small), thus beginning or even completing the biological compatibility test, for instance, as a consequence of an incidental physical contact with M, then frustration would turn out to be considerably amplified, and F would suddenly miss the reward value of that experience. Therefore, when direct and indirect reward are coupled, frustrating the former practically amounts to intensifying the latter, and F will consequently be sexually attracted to the potential, biologically compatible other sex partner she fears to have lost. The extent of the attraction will depend also upon a possible preexisting TU situation with another partner, which might have been weakened through time, making F susceptible to new opportunities of mating. The more solid the preexisting TU condition, the stronger the needed impulse in terms of indirect reward in order to make frustration effective and to open up further relational developments.
However potentially effective, though, the suspension of interest strategy needs skillful implementation: If the suspension is too evident and emphasized, exposing its manipulative character, F will realize this, and all the emotional impact will be lost, possibly together with the chance to give it another try. The crucial aspect of the successful frustration of F’s direct reward is its nuanced character, at the border with non-intentionality so as to feel natural and authentic. A similar effect can be generated by a sudden absence of M preceded by a period of successful courting. Additionally, in this case, the effect is the activation of F-RA in that M’s absence is perceived as equivalent to the frustration of F’s direct reward. In the case of benign manipulation, this sort of strategizing is used only once, in the most appropriate moment to foster F-TU, that is, when F is emotionally pleased and has surrendered to the pleasure of the emotional reward despite not having to feel any obligation to concede at the sexual level. By experiencing a frustration of her direct reward, F turns attention toward the only viable reward left, that is, the indirect, sexual one, which is still attainable through the memory of M’s rewarding physical approaches and of F’s bodily reactions.
If M frequently adopts a suspension of interest strategy, however, his manipulative intention may be more radical and less benign: M could only be interested in his own direct reward, the sexual one; in this case, the name of the game is rapidity in that a prolonged phase of indirect reward not adequately supported by a direct one would not be sustainable for long. If the suspensive technique is instead applied after the sexual intercourse and F-TU has taken place, the manipulative intention is potentially even more dangerous in that the frustration of F’s direct reward would in this case amount to an outright act of (implicit) hostility, as typical, in extreme cases, of pathological narcissistic personalities. Misuse of frustration of a tied-up F is therefore tantamount to sending her a negative signal of lack of attachment and of manipulative intention. Likewise, if F abuses the frustration of M’s direct reward, letting him believe that she will sexually concede, only to perpetually postpone the intercourse, we find the female equivalent of the manipulative male narcissistic personality just mentioned. If F’s manipulative intention is (benignly) limited to inducing M-TU, she will frustrate M in an explicit, clearly interpretable way.
In already formed couples engaging in conflict and/or subject to emotional distress, frustration can be used as a proper marital weapon. In these cases, not only direct reward, but also indirect reward can be attacked by the partner, with the specific purpose of inflicting emotional pain. This is a typical consequence of the breakup of TU and of the consequent interruption of TU-C. Analyzing these issues in depth is beyond the scope of the present paper. However, what may be of interest here is the implication that an effective maintenance of the TU is strictly connected to refraining from frustrating one’s partner, for whatever reason, whether intentionally or not. Once this erroneously happens, apologizing becomes necessary to prevent the negative impact of the action on either direct or indirect reward, or on both. A basic level of emotional intelligence as to preventing and detecting unnecessary frustration seems to be fundamental for couple maintenance and stability.
In a nutshell, we may conclude that:
- -
M’s indirect reward begins to be frustrated when F loses interest and consideration for her male partner and when M consequently feels he is no longer important for his female partner;
- -
F’s indirect reward begins to be frustrated when F feels no longer sexually desired by M and when M consequently avoids approaching her and maintaining physical contact;
- -
M’s direct reward begins to be frustrated when F subtracts herself, physically and mentally, to the pleasure of physical contact with her male partner;
- -
F’s direct reward begins to be frustrated when M does not pay attention anymore to what F says and shows no sign of interest toward her.
Without a continued interest in the partner, TU is seriously threatened, and the situation may easily escalate into a sequence of frustration, wrongdoing, provocation, and revenge, where of course past emotional openness offers to each party plenty of opportunity to harm the partner in a carefully targeted, extremely effective way. On the contrary, reciprocal care strengthens TUs, creating with time couple solidity and psychological enrichment, as continued reciprocal reward within the couple is a clear sign of psychological well-being and personal development [
125].
To sum up, frustration is an interruption in the flow of rewards within TU-C. If such an interruption persists, the cycle is irreparably compromised as TUs are dissolved. Frustration causes distress in AA, in RA, or in both, depending on whether the object of frustration is direct or indirect reward, and may unfold in time in many different ways. A crucial difference is whether the TU has already taken place or not when frustration occurs. Without TU, frustrating indirect reward, the one concerning (passive) RA, does not cause significant distress in both sexes, whereas frustrating direct reward, even in the absence of TU, causes real distress in that it is the AA which is being hit, and the offence is then perceived by the subject, M or F, as a personal attack to one’s own identity—of sexual nature for M, and of emotional nature for F. The situation is turned upside down once TU has taken place. In this case, frustrating indirect reward causes a devastating distress, which deeply undermines F’s sexual self-confidence and M’s affective self-confidence, with serious repercussions upon esteem, trust, balance, and so on. Attacking RA equals to attaching TU itself, as RA is the area where TU occurs.
Once TU-C is started, there are self-regulating mechanisms that come into play in case TU-C is threatened by some intervening situation—in particular in the case of a momentary interruption of the flow of direct rewards, that is to say, of compromised AAs. If, instead, the compromised areas are the RAs, and the interruption affects indirect reward, the issue rapidly escalates to a critical level, and the dissolution of TU may arrive quickly and painfully. In non-secularized societies, this sort of issue is solved by means of socio-cultural compensation mechanisms, generally acting upon Fs who, even when compromised in their sexual RAs, are kept bound to their partner by religious constraints and social stigma, while at the same time socially incentivizing AAs for both couple members, through family affects and protection for F, and through the acknowledgement to M of his masculine rule, which also guarantees a buffer of social acceptance for out-of-wedlock sexual conduct. In secularized societies, such mechanisms have been gradually dismantled, and today TU dissolution as a consequence of interruption of indirect rewards is typically conducive to couple disintegration.
3.6. Inversion of TU-C Flow
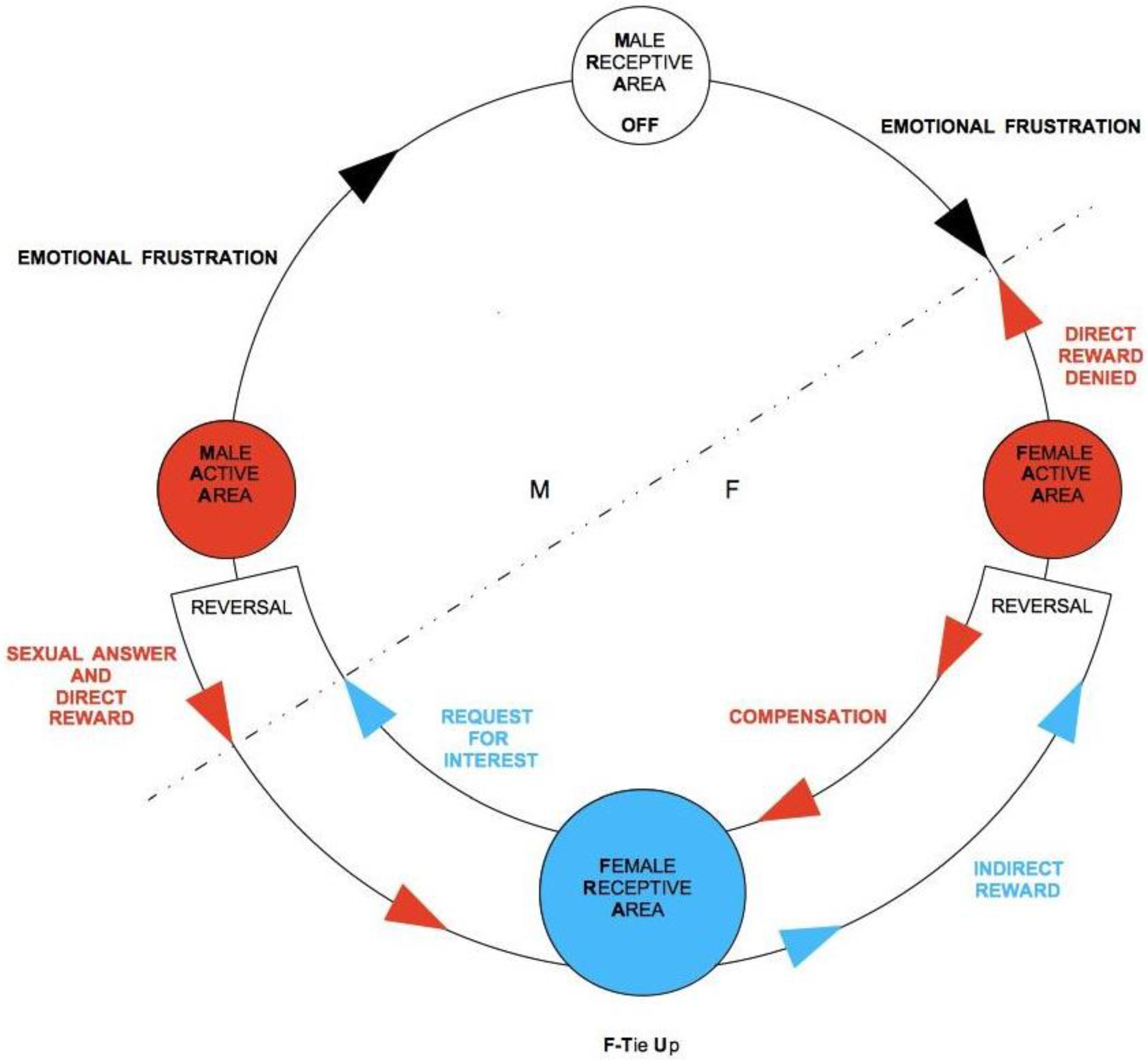
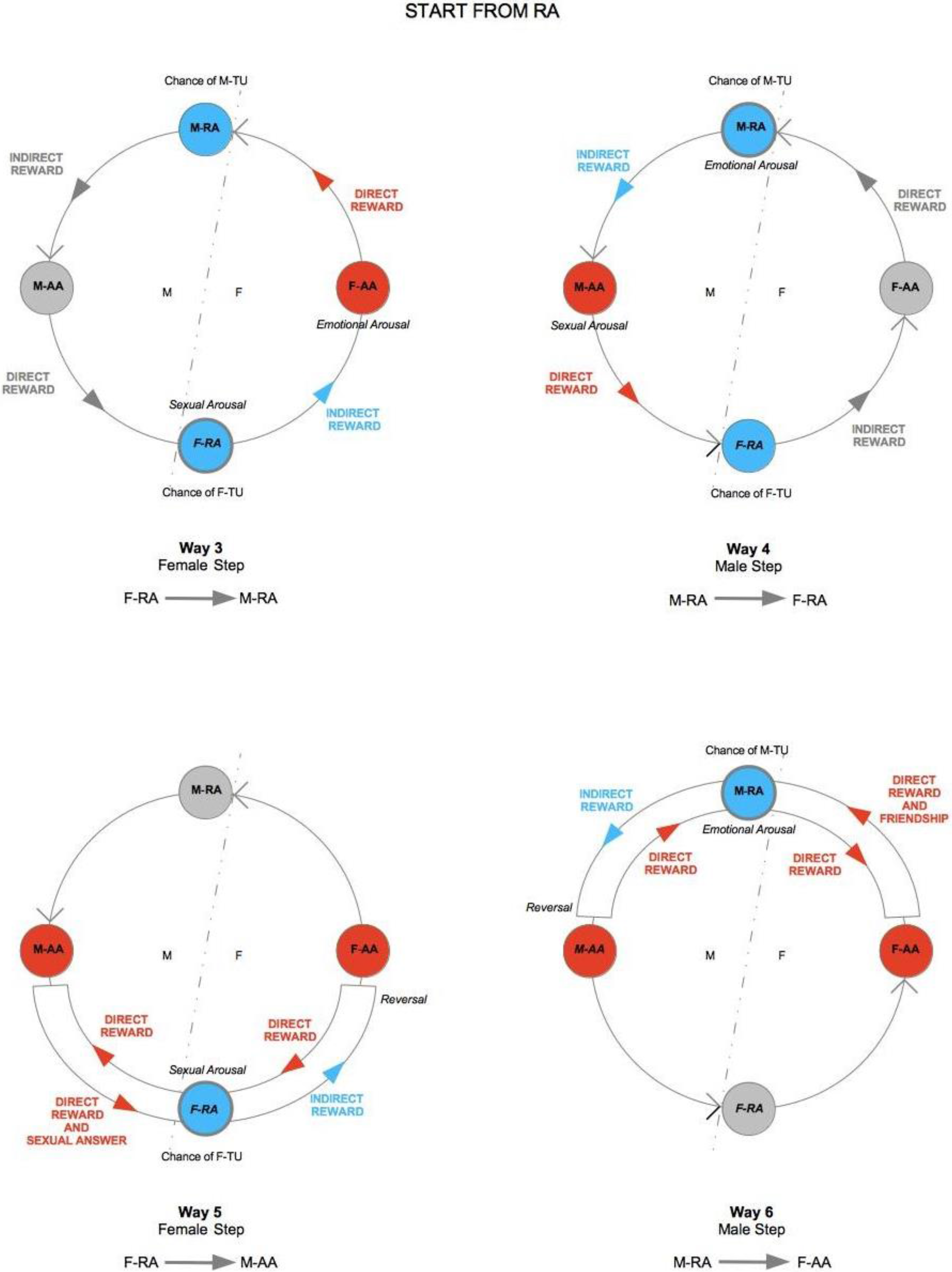
The practical consequence of frustration of direct reward from the point of view of TU-C is a blockage of the flow of rewards, which tends to be compensated with an inversion in the direction of the flow itself. If frustration is momentary, the cycle is therefore able to self-regulate effectively and to restore normal anti-clockwise flow direction. If it persists, the whole cycle is compromised. In terms of our TU-C diagram, interruption of the flow amounts to turning off one of the two RAs, which closes itself to the interaction with the other areas, stopping any circulation of rewards. In this case, the first area to be exposed to distress is the RA that is opposed to the one that has closed down. The cycle’s self-regulation mechanisms are immediately activated, trying to produce a compensation, by exposing the distressed area to an increase in stimulation in order to restore the correct direction of the flow that has been broken. The acting components are AAs, as frustration emanates from an AA to hit the opposed AA. For instance, let us imagine that it is M who is willing to frustrate F. M-AA sends an impulse in the opposite direction with respect to the normal flow (
Figure 7), which, once his own M-RA is reached, turns it off and moves forward, always in opposition to the normal flow direction, to hit its final target, F-AA. We speak in this case of emotional frustration. F’s emotional frustration, in turn, contrasts and blocks F’s direct reward, preventing it from reaching M-RA. The repression of F’s direct reward sparks a regulatory mechanism of the system, which consequently leads F-AA to send, always in the opposite direction with respect to the normal one, a balancing signal to F-RA, which we will simply call compensation (the inner flow in the bottom part of the diagram in
Figure 7). In this case, compensation is an impulse of sexual drive that stimulates F-RA to generate sexual desire. The direction of the flow is still inverted, as F-RA, once stimulated, instead of sending signals to F-AA, as would ordinarily happen, now sends a signal—more specifically, a request for sexual attention, to M-AA. If M-AA responds sexually, then the anti-clockwise direction of the cycle is re-established and starts again flowing in the normal direction. A further passage through F-AR, at this point, may also cause F-TU (if it was not previously established yet) in that F-RA has been highly stimulated. With the sexual response of M-AA, and with its landing onto F-RA, an indirect reward is generated, which, once F-AA is reached, in turn causes a further direct reward, targeting M-RA. At this point, M-AA, which with its sexual response has generated a direct reward for M, could then suspend its emotional frustration impulse toward F and allow F’s direct reward to activate its own M-RA again. If this happens, TU-C is safely restored. If, instead, M persists in his action of frustration, the flow will be inverted again, exposing F-RA to further stress. The repeated exposure to frustration may thus lead to a dysfunctional dynamic, where M aims at the domination of F through a stable condition where only F ends up being tied-up and D-TU is strategically avoided by M.
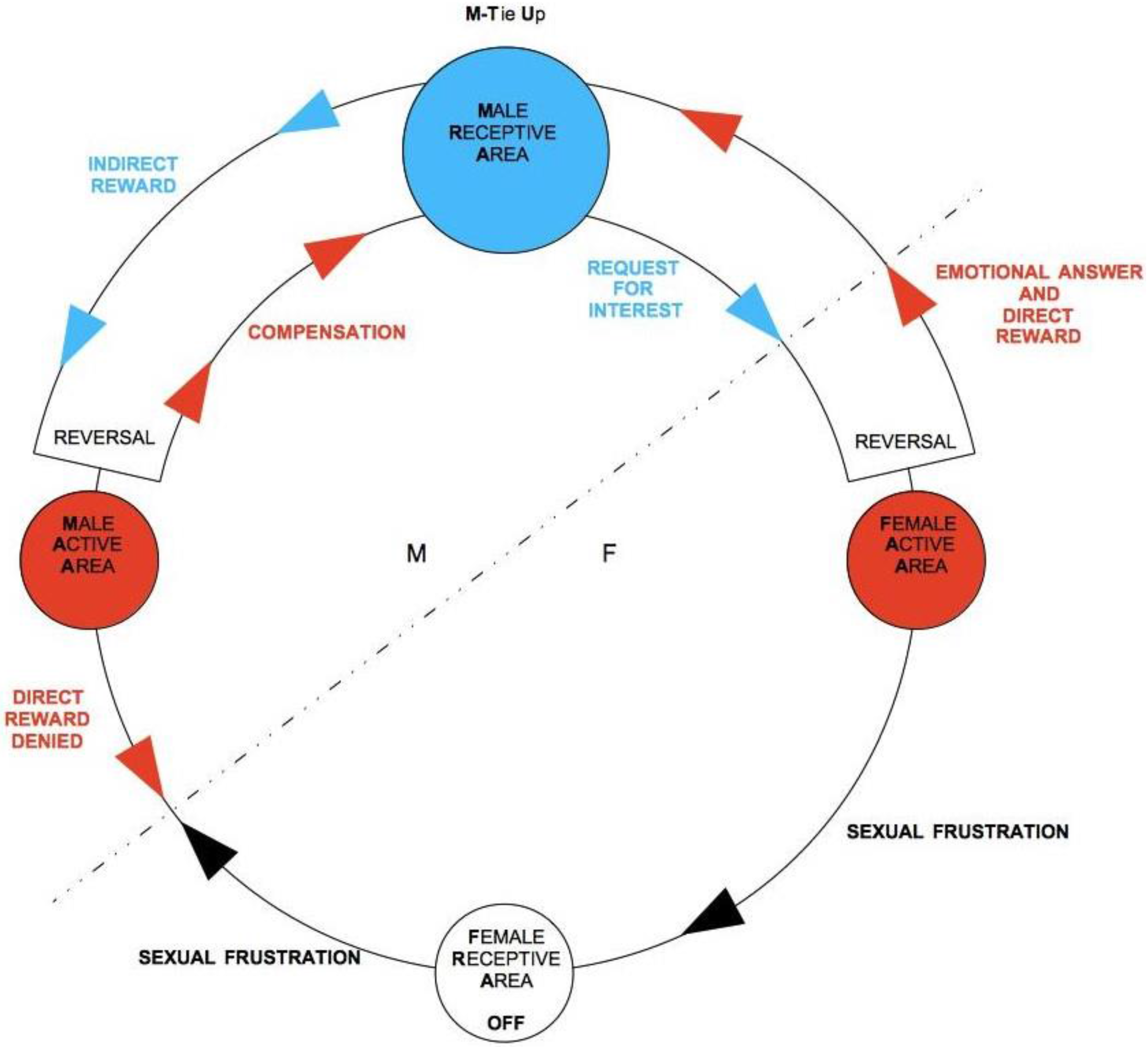
Observation of the graph clarifies what happens when M is not tied-up and is not interested in activating his M-RA in order to get an indirect reward. In this case, a proper TU-C does not even exist, as the flux is persistently confined in the bottom part of the diagram. However, F might interpret this situation as a momentary suspension and, being frustrated by the lack of M’s attention, could end up accommodating the cycle’s regulation mechanism. If several iterations through F-RA occur without causing M to unblock his ongoing emotional frustration of F, F’s distress will tend to become more and more painful at each round. If, in particular, F gets tied-up by means of a sexual intercourse, the suffering may be quite intense. It is unfortunately not easy to discern when a potential partner, whether M or F, is simulating a TU. Particularly for F, whose testing has a merely biological nature and thus does not give any guarantee for the long-term commitment of M, the only truly effective counter-strategy is to slow down the unfolding of the cycle and to frequently screen M’s actual intentions through temporary frustrations of M’s direct reward (see
Figure 8). F must exert special caution and be aware that for her, in the context of TU-C, the sexual intercourse is the situation that exposes her to the higher risk of premature and possibly unilateral TU. On the other hand, deferring the intercourse moves the psychological cost on the shoulders of the manipulating M, who is only interested in his own direct reward. Such a strategy works as a revealing mechanism for M’s true intentions [
126]. In case of a real lack of M-TU, with time the non-existence of TU-C becomes apparent. In the early phases of mating, as a consequence of uncertainty about M’s true intentions, F should learn to modulate with emotional intelligence all possible gradations of sexual reward. A cycle broken through frustration can be correctly restarted without necessarily having to engage in full sexual intercourse. If M shows patience, this increases the likelihood that he is actually engaged and allows him to learn to synchronize to F’s psychological states—an important asset for both sexual and emotional satisfaction. The above analysis tells us that, whereas the evolutionary benchmark for male selection in mating is on the sexual and genetic level, for females it is related to emotional intelligence—women who manage to engage men emotionally through a skillful strategic manipulation of direct rewards and thus to overcome M’s potential sexual opportunism face a higher chance of stable mating by signaling to Ms that, thanks to their psychological qualities, M’s investment in a stable couple may be worthwhile in terms of the quality of marital relationship and of successful child rearing.
The mirror-like situation of a F-driven inversion of TU-C is shown in
Figure 8. When it is F who is willing to frustrate M, it is F-AA that sends an impulse in the opposite direction with respect to the anti-clockwise flow. Once F-RA is reached, the impulse turns it off and proceeds further against the normal flow by hitting its final target, M-AA. We can speak here of sexual frustration. M’s sexual frustration, in turn, blocks M’s direct reward, preventing M-AA from reaching out to F-RA. Additionally, in this case, the repression of M’s direct reward activates a regulatory mechanism of the system, which consequently makes M-AA send an equilibrating signal in the opposite direction with respect to the normal flow for the purpose of compensation (the internal flow in the top part of the diagram in
Figure 8). This time, the compensation is no longer an impulse of a sexual nature, as it was for M-sparked frustration, but rather an emotional impulse that stimulates M-RA. The direction of the flow is still opposite with respect to the normal anti-clockwise one; therefore, M-RA, once stimulated, rather than sending signals to M-AA as it would ordinarily happen, now sends a request for emotional attention to F-AA so as to encourage F to suspend the frustration. If F-AA responds emotionally, the anti-clockwise normal flow is restored. The emotional response of F-AA (external flow in the top part of the diagram in
Figure 8) reaches M-RA and an indirect reward is generated, which moves on to M-AA, generating in turn a direct reward and thus a further impulse toward F-RA. At this point, F-AA, which with its emotional response has generated a direct reward for F, might suspend its sexual frustration, and allow the consequent M’s direct reward to cause M-AA to activate F-RA again. If this happens, TU-C is finally re-established. The structure of this second diagram of flow inversion is thus the perfect mirror image of the former, shown in
Figure 7. The difference is that, in this case, the flow persists in the top part of the diagram rather than in the bottom one. Inspection of the diagram suggests how M’s sexual frustration caused by F produces, as a compensation to restore TU-C, a repeated stimulation of M-RA and an increase of the emotional interest of M toward F. The high level of stimulation of M-RA, analogously to the effect of M-sparked frustration upon F-RA described in
Figure 7, may cause M to carry out the psychological compatibility test and thus lead to M-TU (if not previously established yet). Sexual frustration administered by F may therefore be regarded as expedient to increase M’s possibility of a TU. Moreover,
Figure 8 clarifies what happens when F is not tied-up and is not sexually interested in M, but is nevertheless willing to exploit M’s emotional attentions to her own advantage, for instance, through a promise of future direct reward for M, which will not necessarily occur.
It is useful to remark that sexual frustration of M implies a failed achievement of M’s maximum level of direct reward, i.e., full sexual intercourse. There is, however, a full range of intermediate forms of sexual reward that ensure a gradual satisfaction in the early phases of couple formation, allowing F to check to what extent M is willing to accommodate his frustration with an inversion of the flow of rewards. The more M is willing to compensate the frustration of his direct reward with a request for emotional attention from F, the more likely the production of indirect reward for M and an eventual M-TU. For this reason, in an early phase, going through TU-C without seeking to achieve maximum levels of reward may allow F to check whether M-TU has actually taken place, and M to test in turn F’s psychological compatibility, by amplifying the response of M-RA and consequently the emotional interaction with F-AA.
3.7. The Six Ways: the Role of Entry Points in TU-C
A TU-C may kick off from any of the four Areas, that is, it may start from active Areas (M-AA and F-AA) or from passive ones (M-RA and F-RA). If the Area lies in the F hemicycle, it is the woman that initiates TU-C; if, instead, the Area is in the M hemicycle, it is the man who starts. There are three possibilities for each sex, for a total of six, of which only four will have a chance to bring about the partner’s TU. Whereas starting from AA entails only one possible path, with a quick transition into the partner’s hemicycle, by simply following the anti-clockwise direction of movement of the cycle itself, without crossing other Areas of one’s own hemicycle, starting from RA opens up two possibilities to reach the partner’s hemicycle, one of which calls for the inversion of the cycle’s flow of movement.
In the former case, we have that AA directly communicates with the opposite sex’s RA (Ways 1–2,
Figure 9), and no inversion in the flow of rewards is contemplated, that is, no change in the direction of movement of the cycle, which remains anti-clockwise. This depends on the fact that AA constitutionally relates to the opposite sex’s RA, which always follows up in the TU-C diagram, in addition to representing the most proximate access to the opposite hemicycle, and being excited by AA on its same sphere of pertinence, be it emotional-psychological or sexual. This means that, in the case of the woman, there is an emotional excitement of F-AA toward the male’s psychological sphere, that is, M-RA (Way 1), whereas in the case of the man there is sexual excitement, and the sexual M-AA will directly address the (sexual) F-RA of the woman (Way 2). This represents a classical approach to the other sex and thus depends on the characteristics of each sex’s AAs. If the action of AA is successful, a TU in the opposite sex’s RA will be caused, generating a peak of direct reward for AA—and this may occur at the first iteration or may call for several rounds of the cycle by repeating and strengthening at each passage the excitement of RA, keeping in mind that, at each round, the possibility that the other TU occurs in the starting hemicycle keeps on increasing. In case the compatibility test fails, instead, RA is not activated, and the cycle does not start at all.
When the cycle moves from the AAs, this entails a larger probability of an opposite sex’s TU, while as regards the second TU in the same hemicycle of the starting AA, this depends upon the capacity of the potential partner to complete his/her own half cycle within TU-C, preserving the same anti-clockwise direction of movement. Thus, in Way 1, if M does not succeed in correctly stimulating F-RA, there will be no possibility of a F-TU, and F will only experience a direct reward, maintaining a friendship relation with M, especially if the latter has inverted the direction of the flow. In Way 2, likewise, if F does not manage to correctly stimulate M-RA, there will be no chance of a M-TU. At best, if F inverts the direction of the flow and responds to the sexual offer, M will enjoy his direct reward through a sexual friendship. If the cycle is not successfully completed by the involved partner, then there will be only one possibility of direct reward for the acting AA and thus for the sex of the starting hemicycle, whereas, for the opposite sex partner, in case s/he fails to stimulate the other’s RA or decides to invert the direction of the flow, the lack of direct reward may result in frustration, especially in case his/her TU has occurred.
This mechanism also ensures the efficacy of the system when the cycle is not activated and the couple is not formed in that there will be in any case a high propensity to insemination on M’s side, balanced by a low propensity of F to being frequently inseminated by different partners, consequently preserving the sperm from the possibility of being displaced by that of competing males, and at any rate not being conveniently selected (Way 2). At the same time, F will not be engaged into relations with a bad probability of insemination, remaining available for other mating and TU opportunities, whereas Ms will be concurrently selected in terms of their conformity to the TU-C demands and of transmission of the best generic material (Way 1).
If, instead, the cycle moves from RAs, we need to keep in mind that these are passive areas, which, once activated and excited, must, however, make use of the respective AAs, which are designated for action, to engage into an active interaction with the other hemicycle. If the response of RA takes an active character, RA connects to its own AA. One must realize, however, that RA has its own autonomy of passive response to the stimulating AA, for instance, in female sexual surrender with a strong passive connotation or in sending out invitation signals that M-AA is easily able to decode. Allowing the partner to act sexually in an active fashion, therefore, is a typical response of F-RA to M-AA. When the flow is inverted, instead, it is always F-AA that operates to actively seduce M-AA within the sexual F-RA sphere. If, in particular, we consider male courtship as a form of seduction, when courting has sexual implications, its active nature accommodates the natural flow of TU-C, heading toward F-RA. However, if courting is devoid of sexual implications and only aims at F’s emotional sphere as in chivalry, it will be M-RA that interacts with F-AA, and this happens through an inversion of the flow, that is, when M-RA involves M-AA in an active mode of emotional seduction.
When the cycle starts from a RA, the first step will always be toward one’s own AA (Ways 3, 4, 5, and 6;
Figure 10), but at that point there are two options: reaching the opposite-sex partner’s RA, continuing along the anti-clockwise direction of movement (Way 3, Way 4), or inverting the direction of the flow to head back toward the partner’s AA with a second passage through one’s own RA (Way 5, Way 6). This latter possibility could be regarded as resource economizing, as AA will reach the area that has determined its own excitement more quickly, that is, the other hemicycle’s AA (Way 5, Way 6), whereas in the latter option, despite not having to invert the cycle’s direction of movement, the partner’s AA will only be reachable once the partner’s RA has been crossed, and this implies a switch in the sphere of pertinence, from sexual to emotional or
vice versa, depending on whether we are in the M or F hemicycle, to subsequently return to the initial sphere in which the excitement has originated (Way 3, Way 4).
In other words, we must realize that, if we start from AAs, these get excited through their forward-heading action (in terms of the normal anti-clockwise direction of TU-C) toward the other hemicycle’s RA. However, if we start from RAs and an indirect reward is produced anyway, the AAs will also be excited by this other reward that comes from behind, that is, from the same hemicycle’s RA, for which there is, instead, only a possibility of passive excitement that depends upon the opposite hemicycle’s AA, which still precedes according to the anti-clockwise direction of the flow of rewards.
It is thus understandable that, when we start from RAs, for the same hemicycle’s AAs, there will be a temptation to invert the direction of the flow. Now, the AA, through its own RA, will experience the attraction toward the partner’s AA from which the reward comes, this time no longer direct but indirect, and which it can achieve in two ways: moving along and generating direct reward in the direction of the other hemicycle’s RA (Way 3, Way 4), in the hope of managing to excite it to reach, with a further step, the AA it sought for (so that it commands the production of further indirect reward). Or rather, inverting the flow and generating direct reward in the direction of the partner’s AA (Way 5, Way 6). Obviously, we are not dealing with the same kind of direct reward in the two cases because the one addressing the other hemicycle’s RA will remain within the same sphere of pertinence, be it emotional-psychological or sexual, whereas the direct reward that moves clockwise toward the opposite hemicycle’s AA by crossing its own RA will switch to the latter’s sphere of pertinence so as to be able to dialogue with the partner’s AA.
For more clarity: If, for instance, the cycle has started from the F hemicycle, the direct reward that travels in the direction of M-RA will express itself on an emotional level, whereas the direct reward that moves toward M-AA will adjust to the sphere of pertinence of the F-RA it crosses along the way and will approach M-AA sexually. In a sense, each sex can decide whether to go along the faster, easier path in the economy of the cycle, which entails carrying out the inversion, or choosing the longer, more exacting path, which requires a complete round. From the point of view of couple formation, however, the system remunerates the most expensive option, which is the one that accommodates the TU-C direction of flow, with a concrete possibility of a TU of the partner who has actively sparked the cycle. The inversion of the flow indeed represents the only case that does not contemplate the TU of the partner who has excited, and possibly also tied-up, the initially engaged RA. The inversion of the flow moreover represents a real risk of one-sided TU for the subject that practices it, with a cycle that has not even properly started, in that the opposite sex’s AA, once obtained and its reward enjoyed, does not follow up with more action, whereas the respective RA remains turned off, not having been crossed at all.
In evolutionary terms, the short path ensures the species’ survival in that: if in one case (when it is F who inverts the flow), the sexual intercourse is quickly reached; in the other (when it is M who inverts the flow), what is favored, instead, is the selection of the fittest subjects for insemination purposes, ruling out those M who proved themselves less competitive on sexual grounds, and who therefore did not manage to tie-up any female subject. The long path conversely favors the species’ cultural evolution, as it is the only one that, once the positive outcome of the compatibility test for both sexes is checked, ensures the unfolding of a complete TU-C and the constitution of the stable couple that will rear the offspring.
Some simple exemplification may help to better appreciate the specific characteristics of each Way from the viewpoint of the dynamics of TU-C. Way 1 (F-AA → M-RA) may, for instance, represent the case of a woman who admires a man for his endowment of psychological resources and skills (gentleness, sympathy, intellectual brightness, talent, professionalism), or of material resources (wealth, power, etc.), which constitute for her a source of attraction with a clear evolutionary rationale. In Way 1, F thus admires M to the point of feeling psychologically attracted to him, independently of his aspect and physical characteristics. This psycho-emotional approaching of F allows M to interact on the same sphere and to test his psychological compatibility with F. If the test succeeds and M, in turn, will feel emotionally attracted to F, some level of indirect reward in the M hemicycle will be produced, which will push the cycle toward the sexual sphere (M-AA), but not necessarily—if, for instance, preexisting M-TU and F-TU with other partners are already in place, or if F is not sexually attractive for age reasons or other factors, if M has his own inhibitions or is just not into heterosexual sex, the flow could stop anyway. M could then accept a friendship with F, without even approaching her enough to enable the test of biological compatibility in her own F-RA.
Way 2 (M-AA → F-RA) represents, instead, the classical case when a man notices a woman who attracts him physically, that is, excites him in a sexual way, thus activating his own AA and consequently leading him to deploy all of his seductive ability. M-AA will produce ever increasing shots of direct reward, which will prompt M to approach F physically, and the choice of the how, and of the how much, will be a key part of the seductive strategy, successful or not according to cases. F, on the other hand, could already be happily tied-up to someone else. In this case she will try to keep herself out of (physical) reach to prevent the kickoff of a biological compatibility test (ranging from simple visual, olfactory, and vocal examination to move to a tactile-energetic sampling, and finally a chemical one). The test is automatic, and can be avoided only by maintaining physical distance, but even if it would turn out positive and would generate a shot of indirect reward in the F hemicycle, F might not be interested and avoid accommodating M for various reasons, especially if there is an absence of F-TU toward him. However, for other possible reasons, F could accept the approach of M, actively responding in sexual terms with an inversion of the cycle, or instead following the anti-clockwise direction of movement of the rewards, and become interested in M from a psychological point of view. The cycle, however, will not move further in two cases: if F sexually gratifies M from the very early phase of the approach with full sexual intercourse before M-TU is brought about, or if M does not tie-up because there is not an equivalent psycho-emotional attraction toward the F who attracted him physically, that is, if the psychological compatibility test with F fails.
Way 3 (F-RA → M-RA) is the fortuitous case where a positive test of biological compatibility occurs in the woman without anyone noticing, including the man who is the object of the test. The woman thus starts to feel physical attraction for the man, thereby resolving to seduce him primarily on an emotional, psychological level. This Way concerns examples of women who proceed carefully, without exposing themselves to a F-TU too precociously, but that at the same time emotionally re-elaborate their sexual instincts. The activation of the F-RA is generally quicker the younger she is and the less sexually mature she is, as in this case the test will be less detailed and demanding. For example, in the case of a teenager, it may happen that the sight and the sound of the voice of a potential male partner that corresponds to some ideal standard of underage male attractiveness may be enough to activate her F-RA and to generate indirect reward, even if this will not be conducive to an effective, full-fledged TU, the biological compatibility test not having been completed (that is, what is commonly meant by an experience of Platonic infatuation). What is interesting to observe with very young subjects is that F instinctively starts to experiment and to practice with her own AA by attempting an approach toward the M-RA of the male subject who has stimulated her F-RA. The first teenage love experiences may thus represent an entirely natural way to learn about future romantic interactions, and especially about how to successfully move along a fully functioning TU-C.
In Way 4 (M-RA → F-RA), the man feels psychologically attracted to a woman, up to the point of tying-up, and thus attempts to make her tied-up in turn by means of an appropriate physical approach. This is the Way that best represents a male subject who is fit for long-term mating and child rearing. M instinctively operates the choice of his partner in this regard already, by primarily looking for a psychological compatibility with the F who could interest him sexually, while ruling out instead those Fs who are only interesting on a merely sexual level. The M of Way 4 represents the man who wants to be intrigued by a woman both mentally and sexually, and who moreover feels the confidence to engage himself sexually once he has found the right F partner, showing a stronger persistence and determination in the construction of the relationship with respect to the M of Way 2, who, instead, could just be satisfied with his own (tied-up) achievement of direct reward of a sexual nature.
Way 5 (F-RA → M-AA) represents the case, rather frequent today, of single women who finally find the recognition of their right to make their own sexual choices freely and autonomously. However, this option does not necessarily guarantee fulfillment from the psycho-emotional viewpoint, and many women still realize, bitterly, how it may be difficult to achieve a satisfactory level of direct reward in the sentimental sphere with the same ease of men. The reason is because, for F, the inversion of the cycle according to the logic of Way 5 indeed allows an immediate access to sexuality, but at the same time, in the absence of a tied-up male, strongly limits the opportunity for direct reward and thus the full social acknowledgement of her sex identity, which is linked to the expression of her AA. The male partner, on the contrary, receives a full direct reward from a relationship centered upon the sexual dimension, consequently feeling perfectly socially confirmed in his sex identity. The only reward which F can access in Way 5 is thus the indirect one, with a perennial exposure to the risk of a one-sided TU. There is moreover a further complication of a social nature: A woman who freely makes her sexual choices outside the context of a stable couple must still face, even in the most permissive social environments, a strong social prejudice, and may, for instance, encounter difficulties to enforce her rights and her reasons vis-à-vis retaliations from former partners who aim at devaluating her identity and social dignity via stigmatization of her previous sexual availability toward them (think, for instance, of the sadly escalating phenomenon of revenge porn [
127]). The achievement of a real sex parity in this regard is far from simple, and, to this purpose, women need to acquire specific relational competences, maturing a deep awareness of their as well as of male nature, and of the associated social perceptions, and to avoid surrender to the simplification that identifies emancipation with a mechanical imitation of the respective male role models.
Way 6 (M-RA → F-AA), finally, is, for instance, the classical case of the shy, immature teenager who, having fallen in love, does not know how to make use of his M-AA to win the sexual attention of his loved one, and attempts an inversion of the flow in the hope that she will make the first step. This is a passive attitude typical of M-RA, which is rarely remunerated by selection in couple formation. The problem is exacerbated when M must face the competition of more expert and proactive males. The possibilities opened by Way 6 may, however, be widely utilized to gratify women, even with a simple exercise in chivalry or a bland courtship which will please women of any age and social extraction. The man who practices Way 6 skillfully will thus be able to obtain an ascendant on many women, consolidating a situation that might also be conducive to a relational development in the sexual sphere. Way 6, in the absence of competition with other males, may represent a good preparation strategy to obtain feminine esteem and respect before the sexual approach, but at the same time is very risky in that it may confine M into the role of the friend-confident of F, making the passage to the sexual sphere increasingly more difficult the more the relationship gets locked into its original mode.
A last possibility, which is not, however, contemplated among the six Ways just presented, but that may happen anyway, is that of a simulated TU-C sparked by F, in a sort of manipulative mirror reflection of Way 1. Here F starts from her AA but moves in the opposite direction with respect to the normal anti-clockwise direction. The reason why we do not include it among the Ways is that, in this case, interaction is programmatically started in a logic of manipulation in which TU-C is entirely simulated, from the very beginning and with purely instrumental purposes. An F who moves from her F-AA to launch a cycle of interaction in the opposite direction of the anti-clockwise movement corresponds, for instance, to the case of a sexual approach toward M to get material benefits, in the absence of a F-TU, or even of a simple activation of F-RA (for example, if M has considerable financial or power resources). Moving in the opposite direction with respect to TU-C, F first crosses her inactive F-RA to proceed further toward M-AA and stimulate M sexually so as to amplify as much as possible his direct reward, and therefore the whole interaction is shaped by a manipulative logic. To the contrary, a manipulative M has no need to move from the beginning in the opposite direction with respect to the natural flow of TU-C to cause an instrumental, one-sided TU in a F with considerable material resources, as this goal can simply be achieved by following the flow of TU-C and going, first of all, for an active sexual seduction of F-RA and subsequently for a passive one, by simulating M-TU once F emotionally responds on the F-AA sphere.
When F sexually seduces M in an instrumental way, it is not necessary that M gets unilaterally tied-up, but it is, however, necessary that the relationship be stabilized to allow F to extract the material benefits she craves for. If F would limit herself to stimulate M-AA, once obtained his reward M would simply abandon the relationship in search of more rewards elsewhere. F has therefore two alternatives. The first, risky in the short term but convenient in the long term, is that of attempting M-TU, to stabilize the relationship and hedge against the competition of other Fs who could approach M with a similar tactic. In this case, once having sexually stimulated M-AA, F could follow the natural direction of flow of the cycle and emotionally activate her own F-AA to stimulate M-RA. The risk lies in the fact that, in case M’s test of the psychological compatibility of F fails, F could be discarded by M. F should in this case invert the flow again and attempt a new sexual seduction of M through a further stimulation of M-AA. If the attempt turned out successful, F would have gained more time for the pursuit of her personal material advantage. The alternative strategy is that of not undergoing the psychological test by M, thus minimizing the risk of abandonment in the short term, but remaining exposed to the substantial risk of being discarded sooner or later in favor of other competing Fs.
A summing up of the basic characteristics of each Way is presented in
Figure 11.



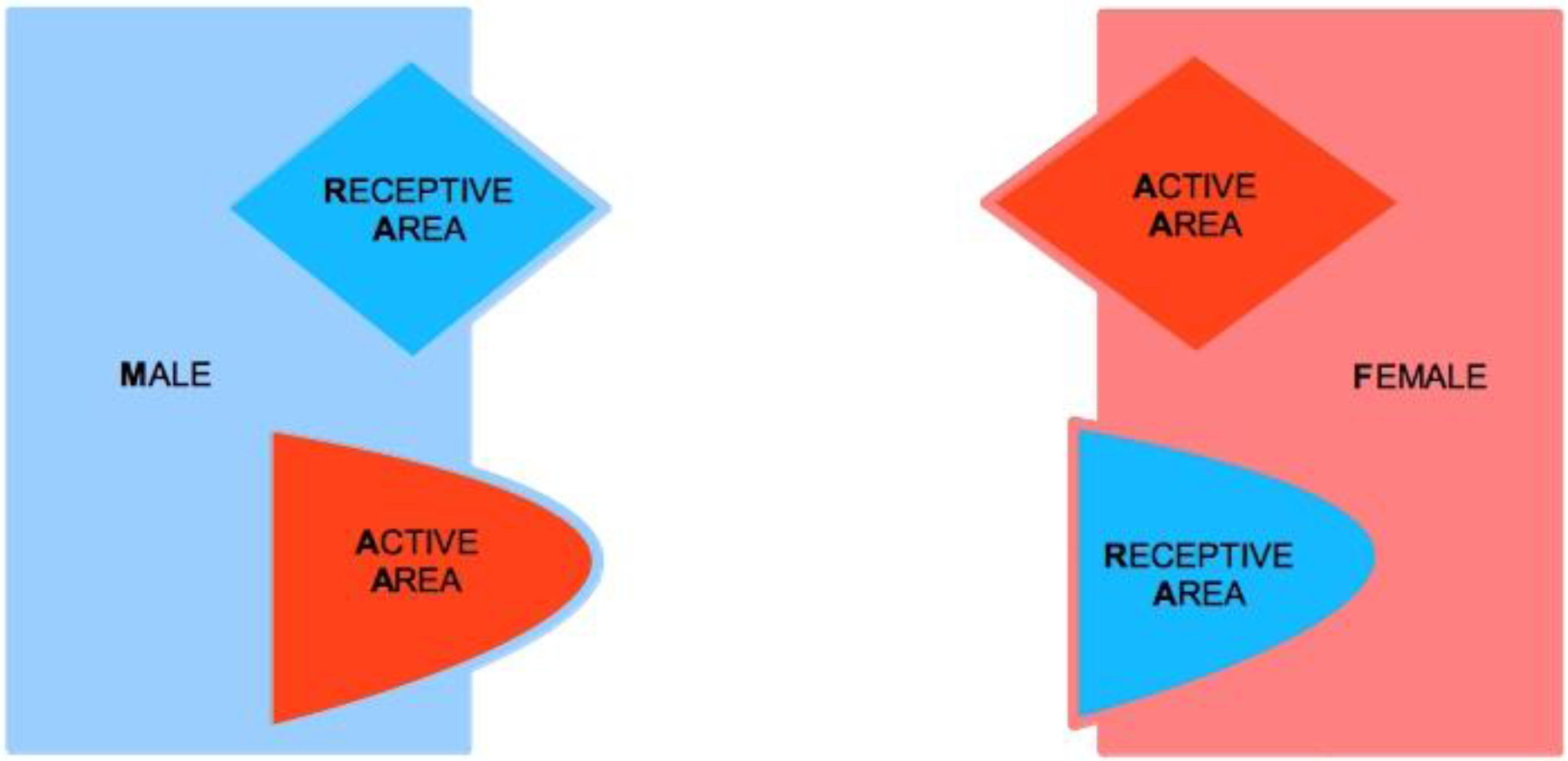{kind=link}
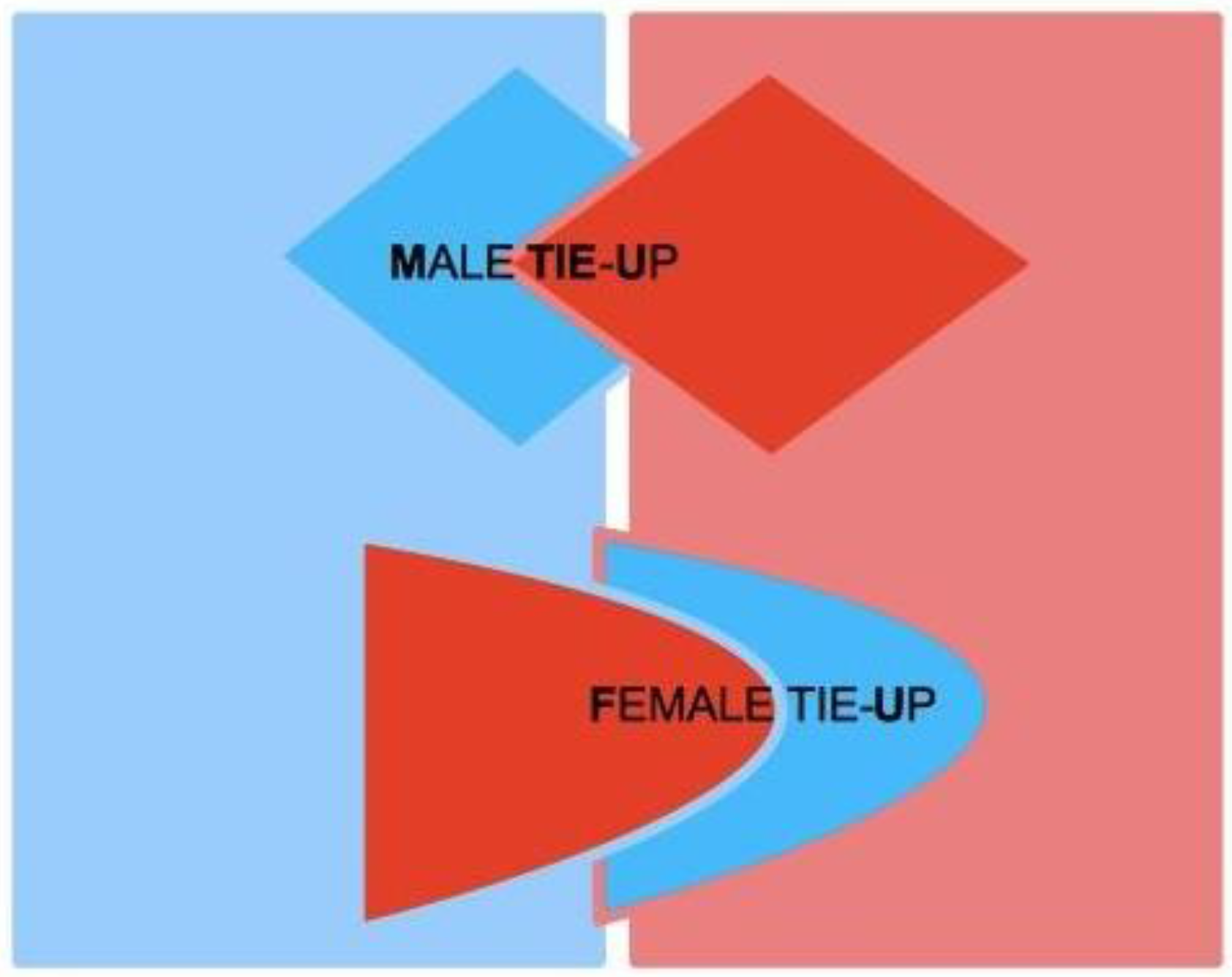{kind=link}
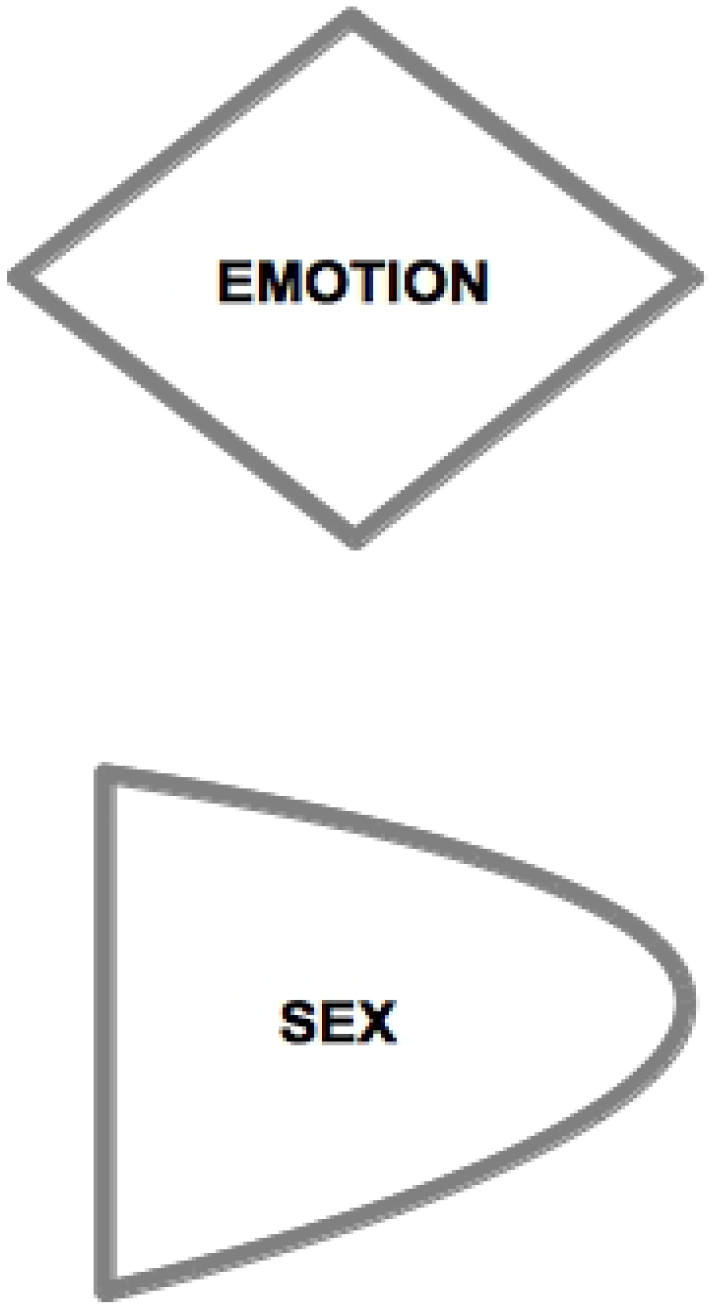{kind=link}
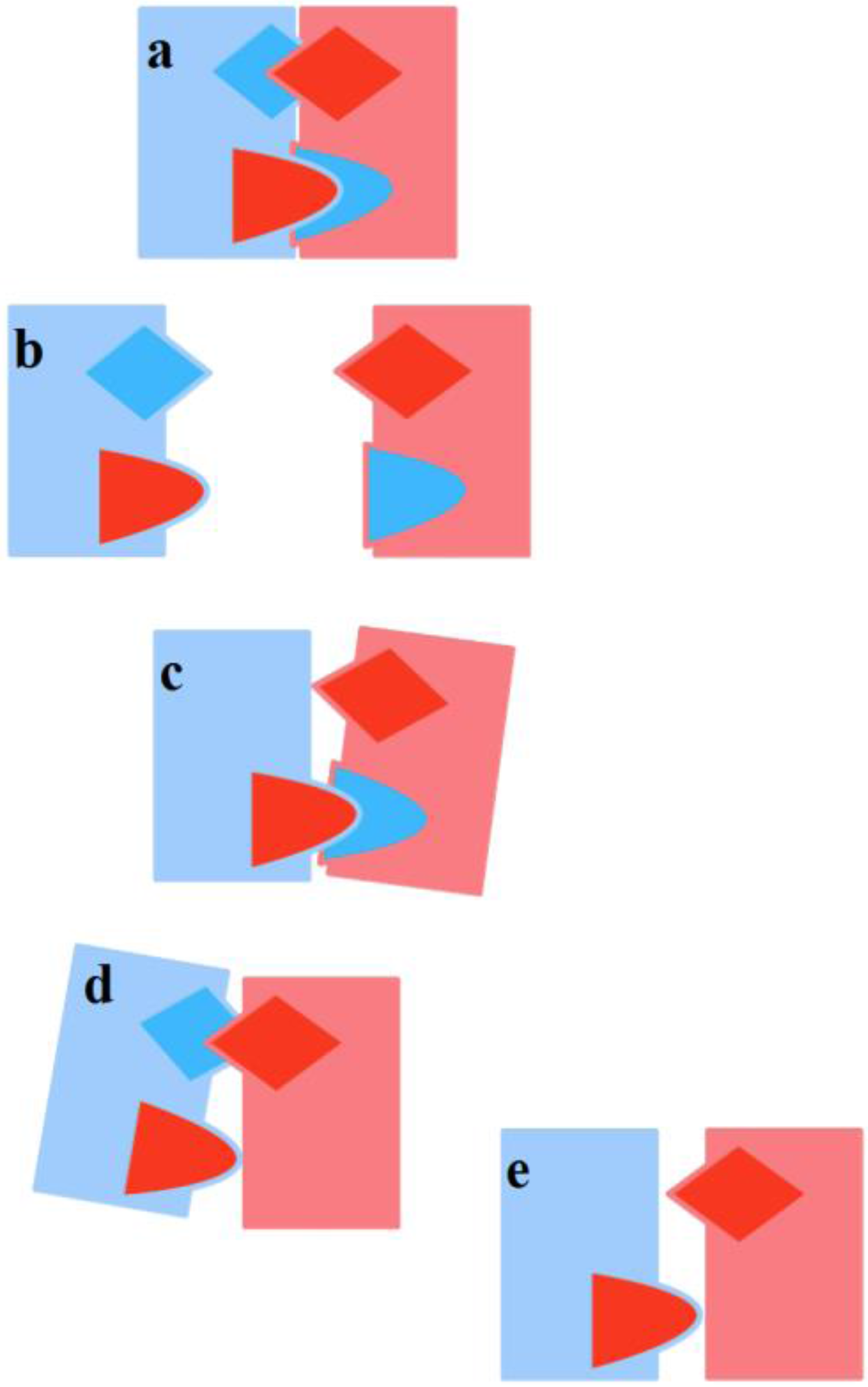{kind=link}
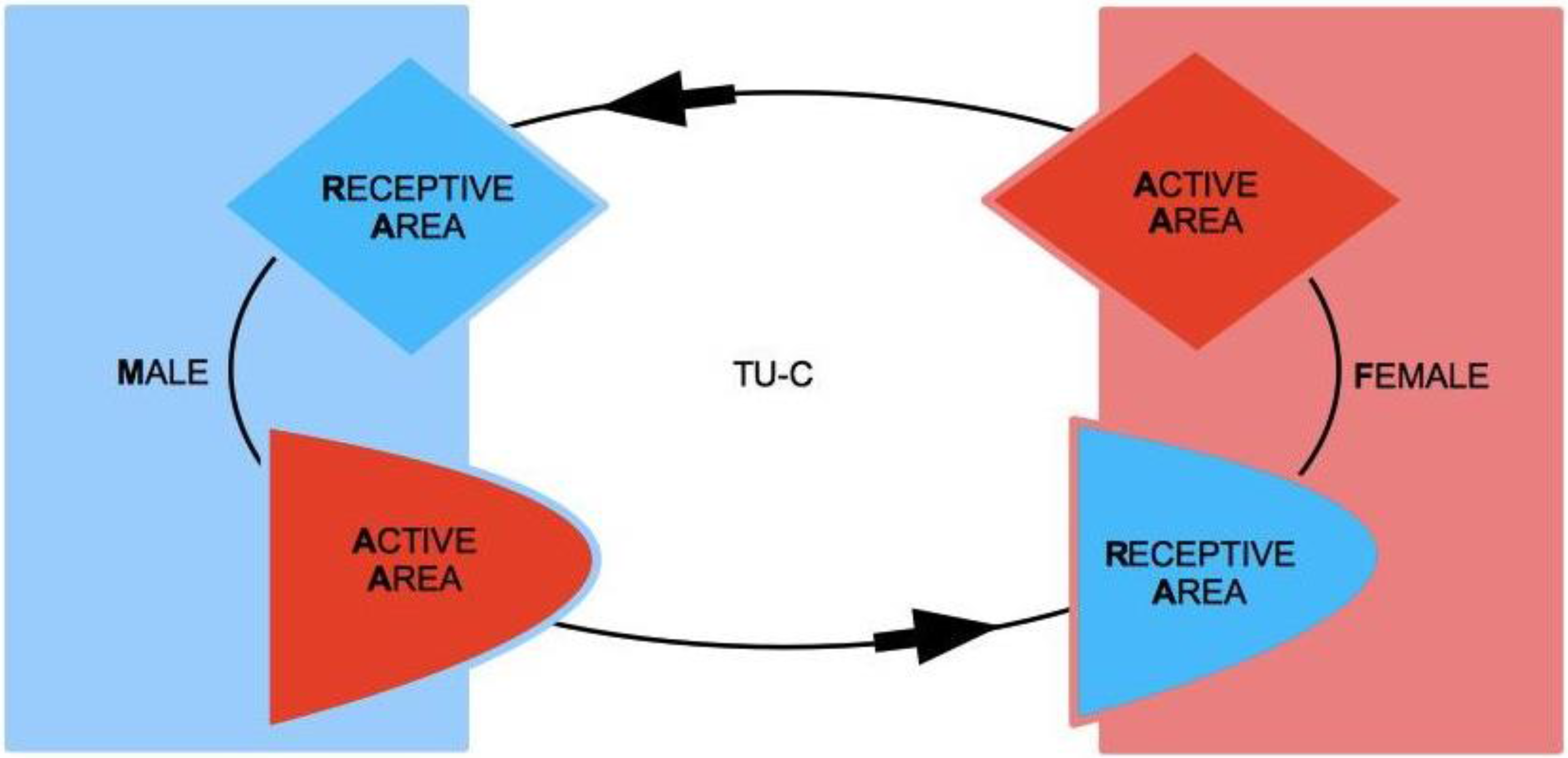{kind=link}
{kind=link}
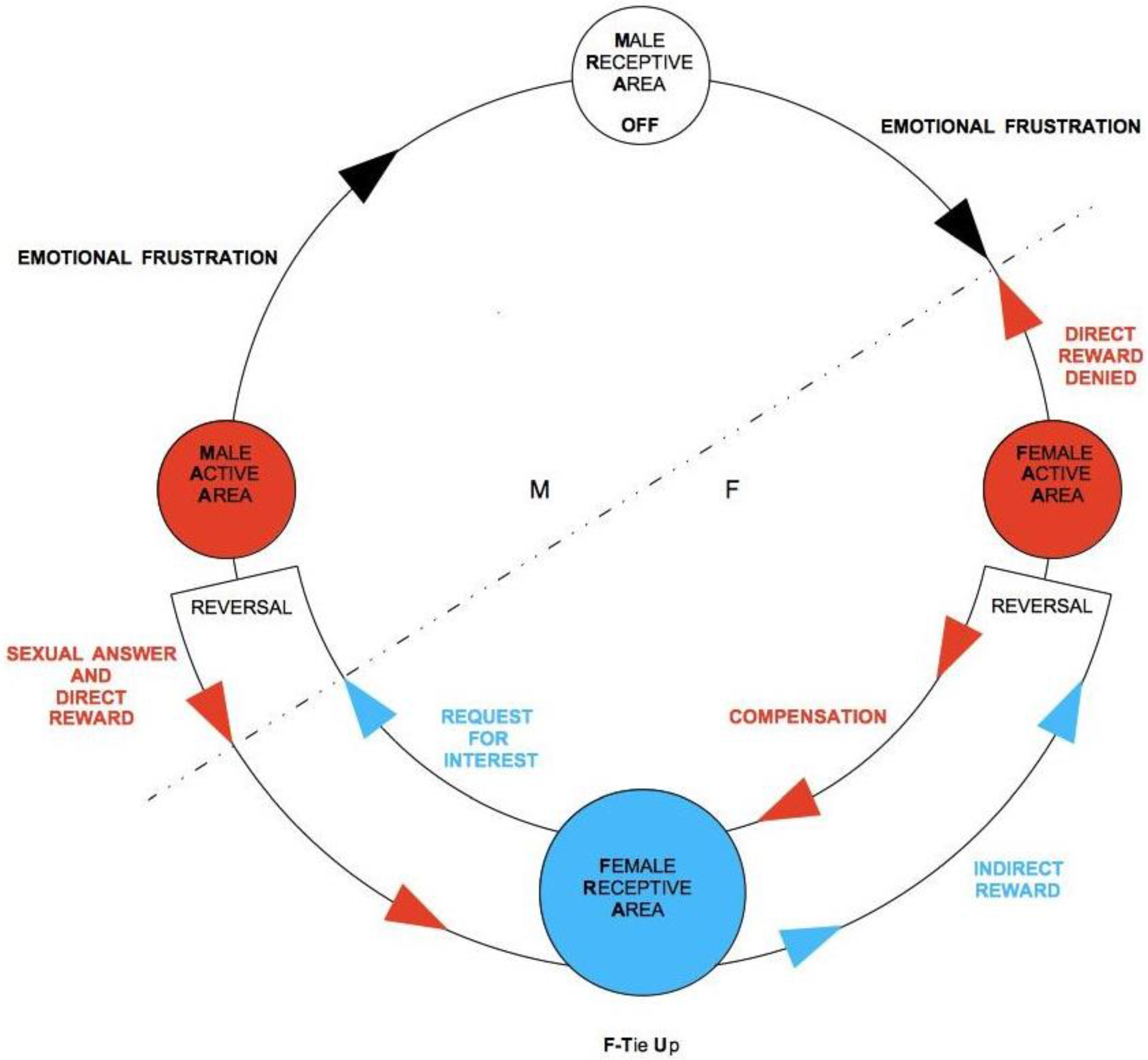{kind=link}
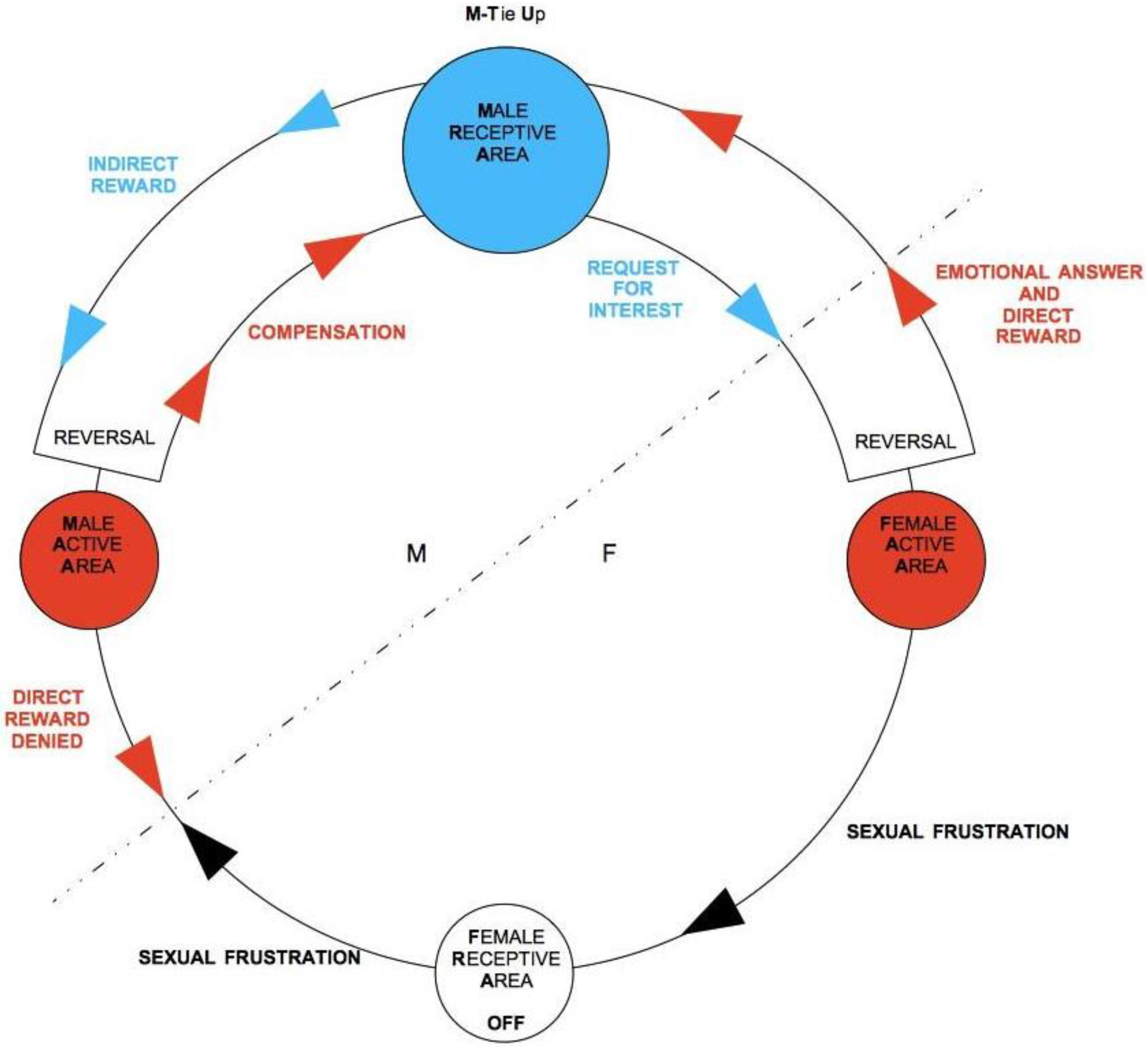{kind=link}
{kind=link}
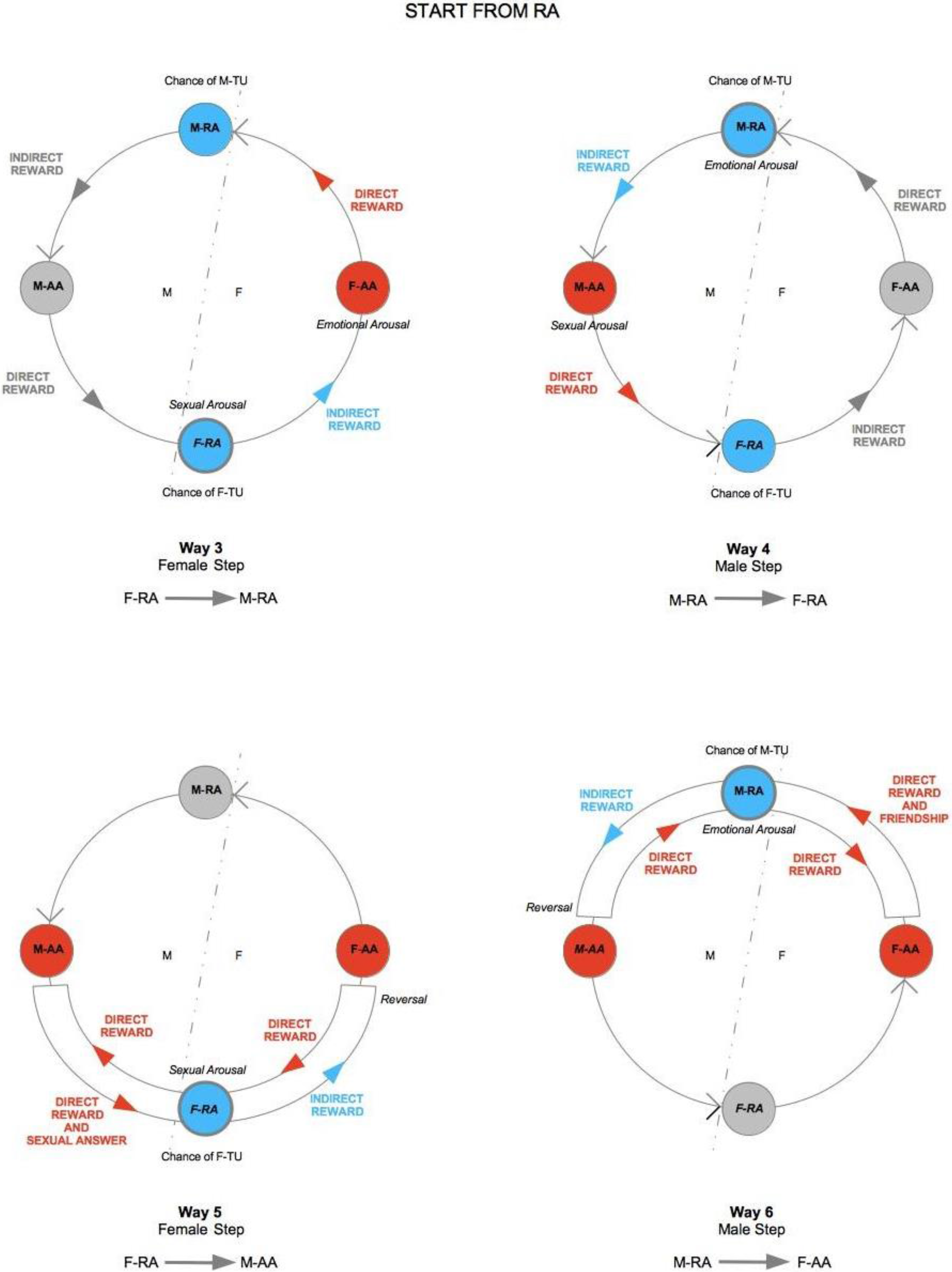{kind=link}
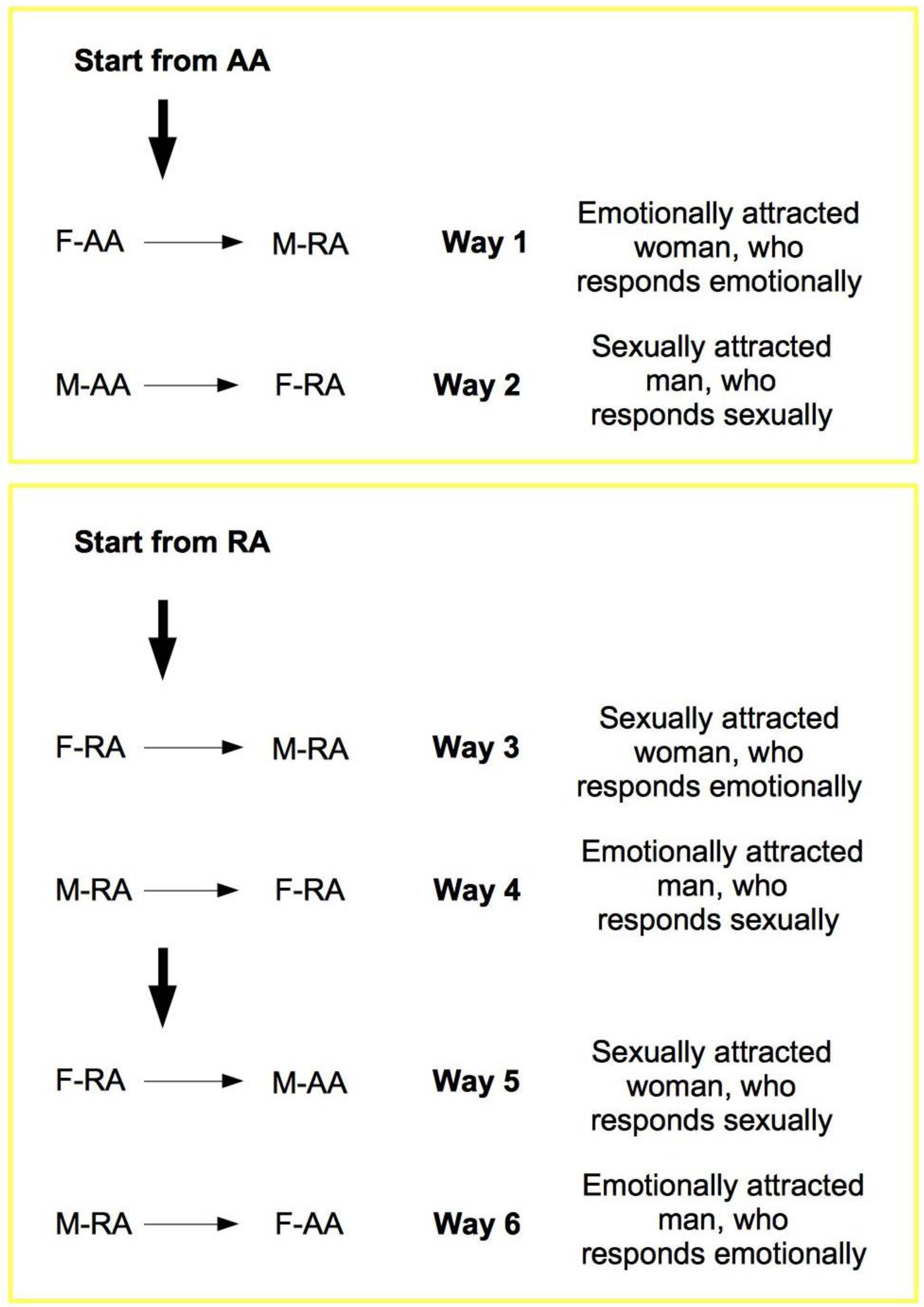{kind=link}