
A Study of the Potential Impact of Dredging the Corpus Christi Ship Channel on Passive Particle Transport



Abstract
:1. Introduction
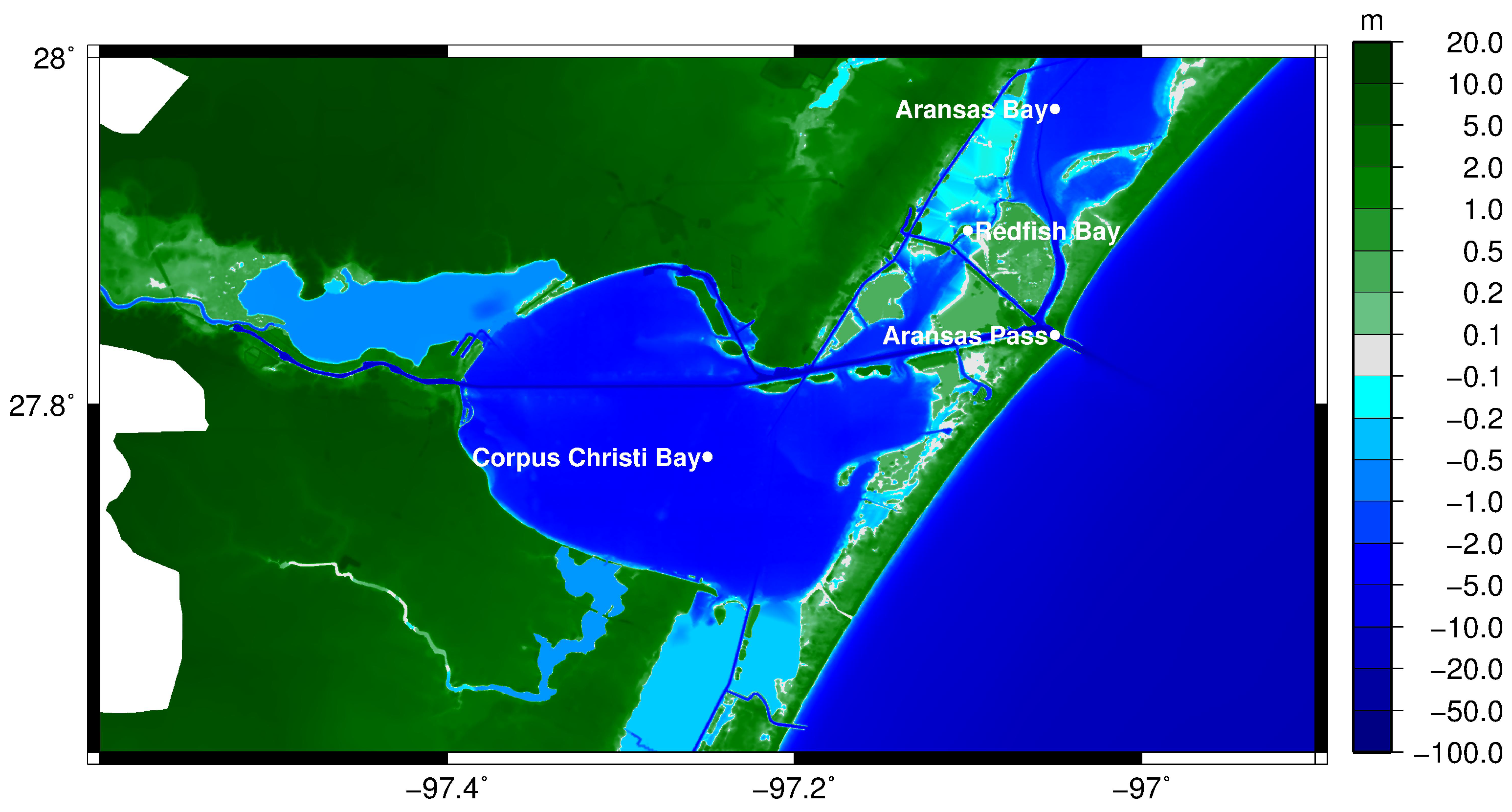
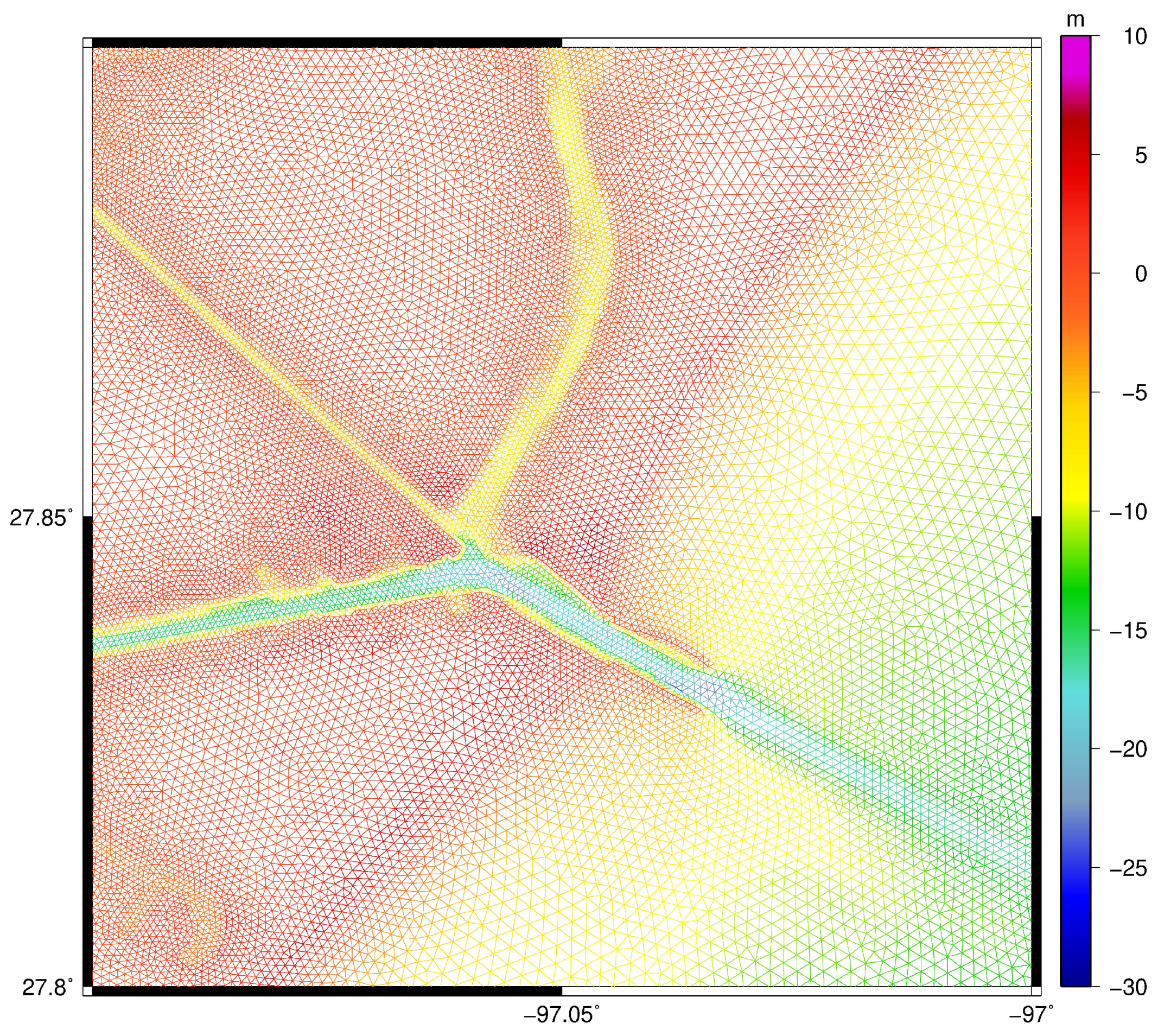
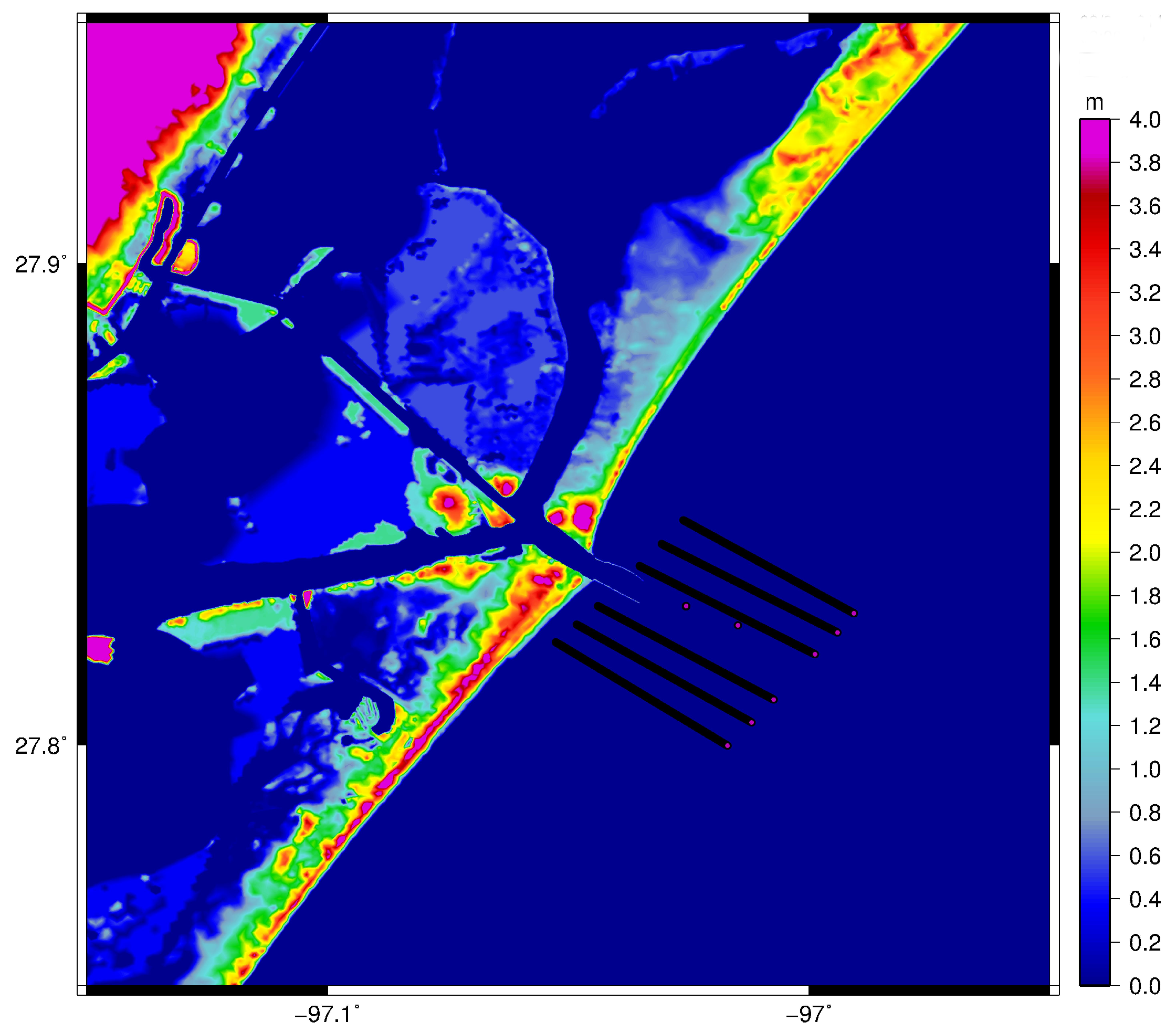
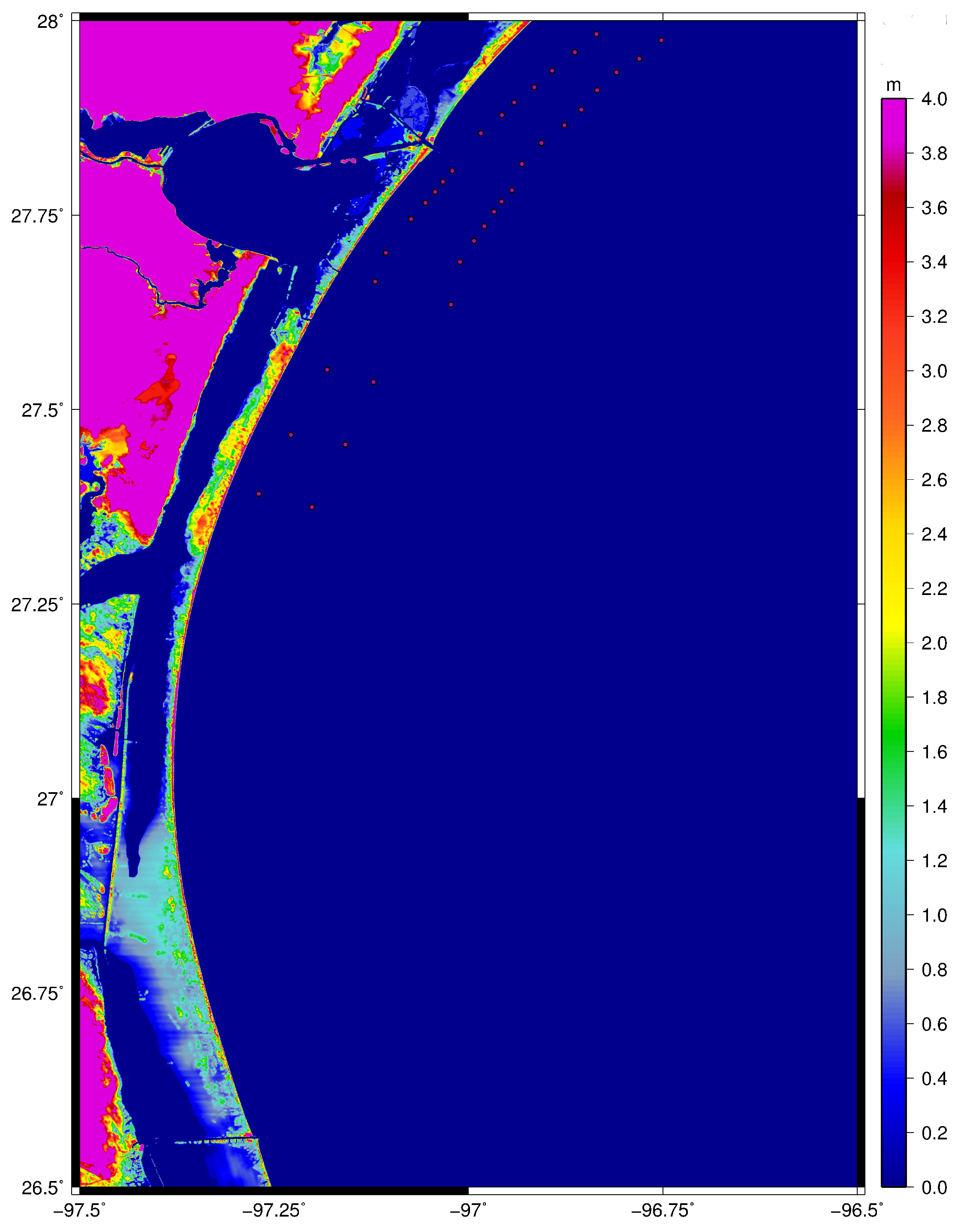
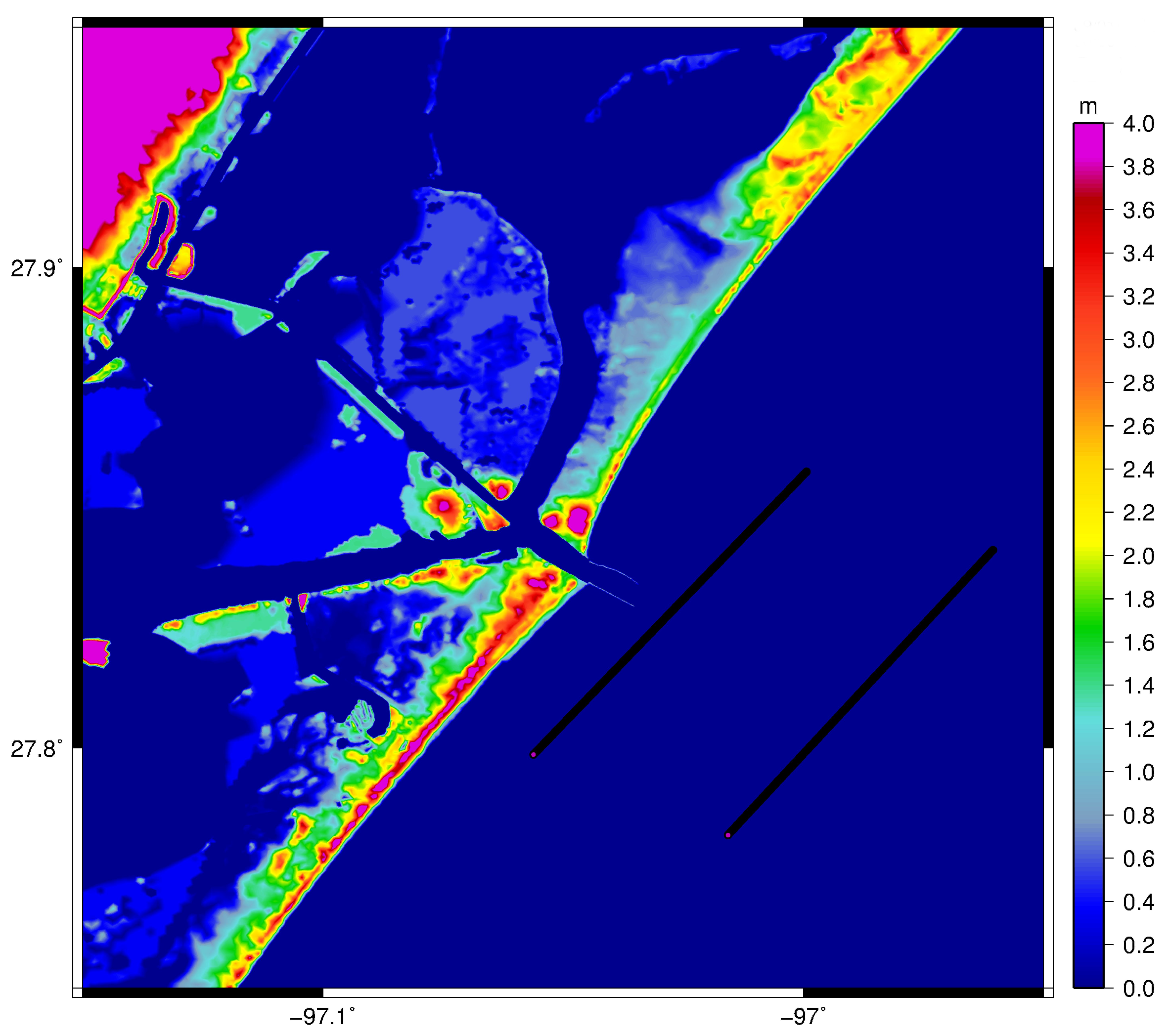
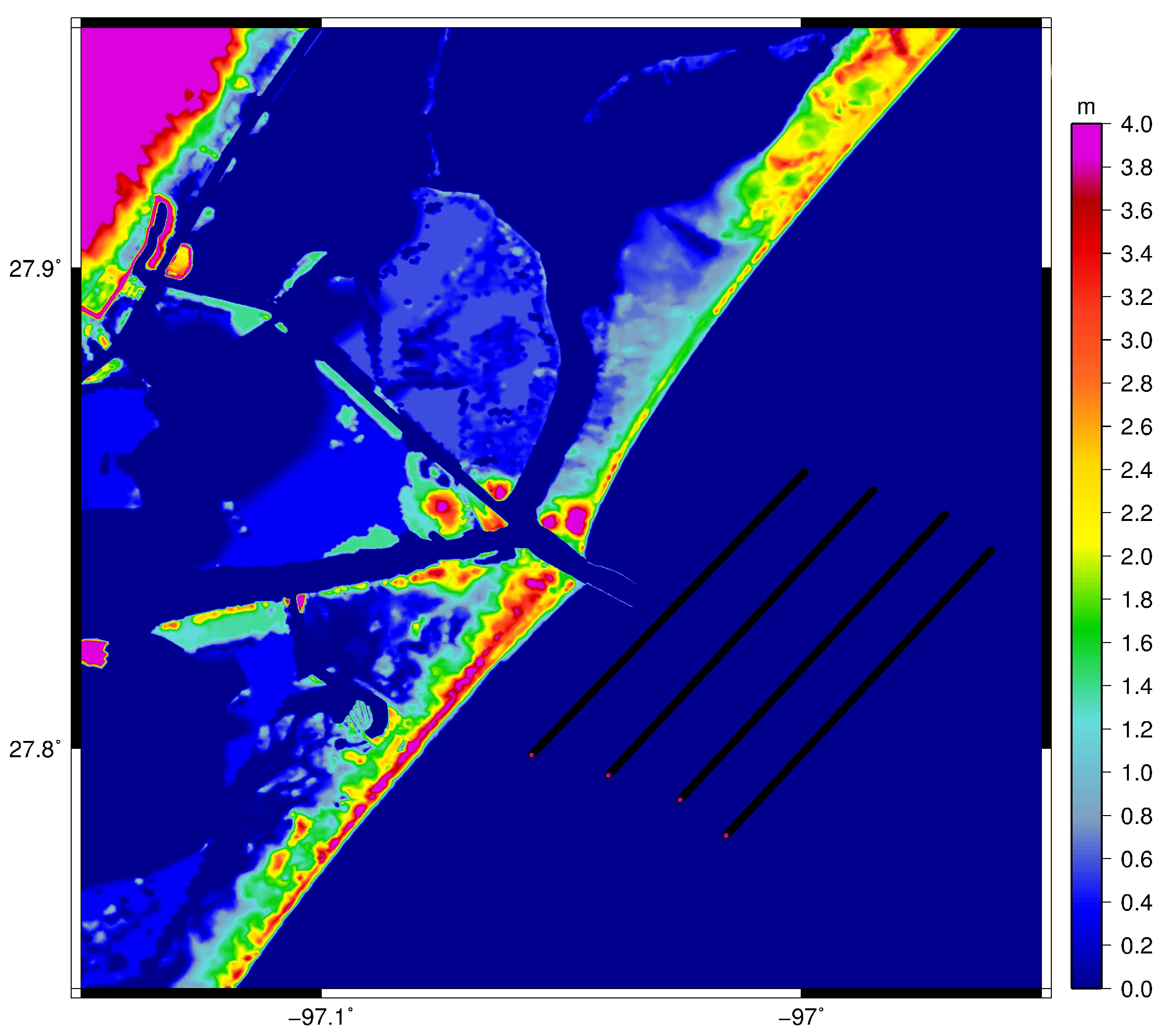
2. Materials and Methods
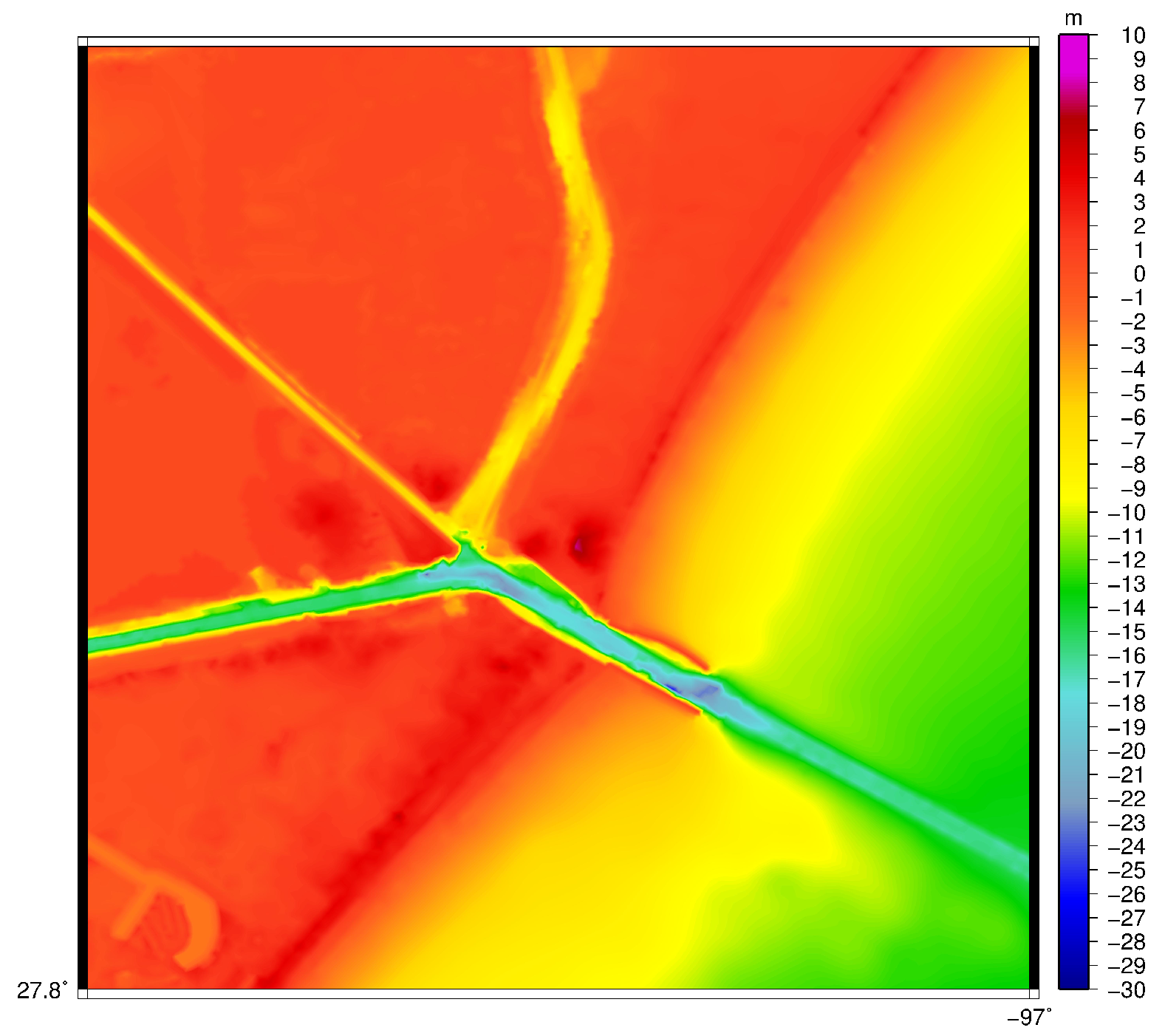
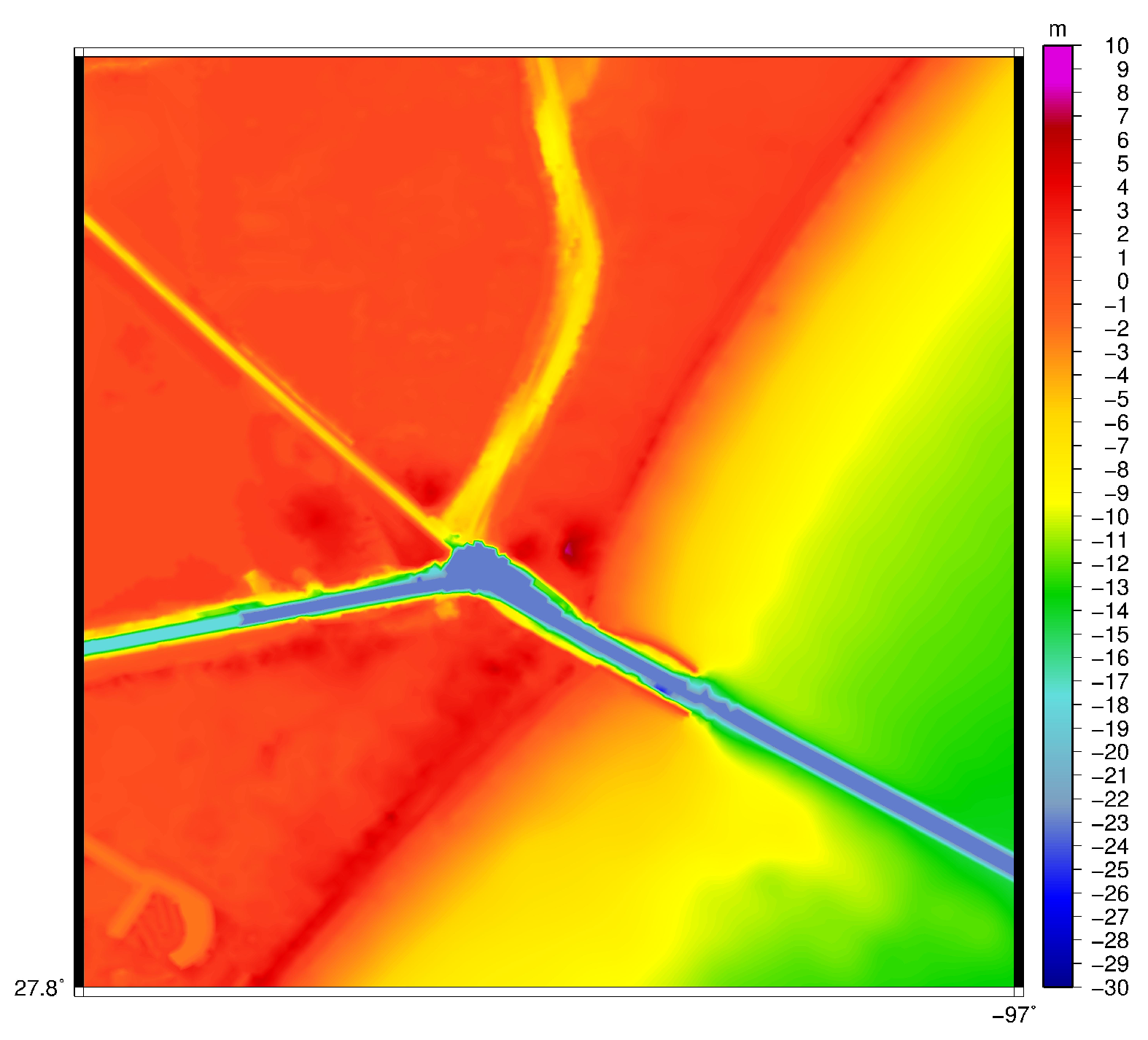
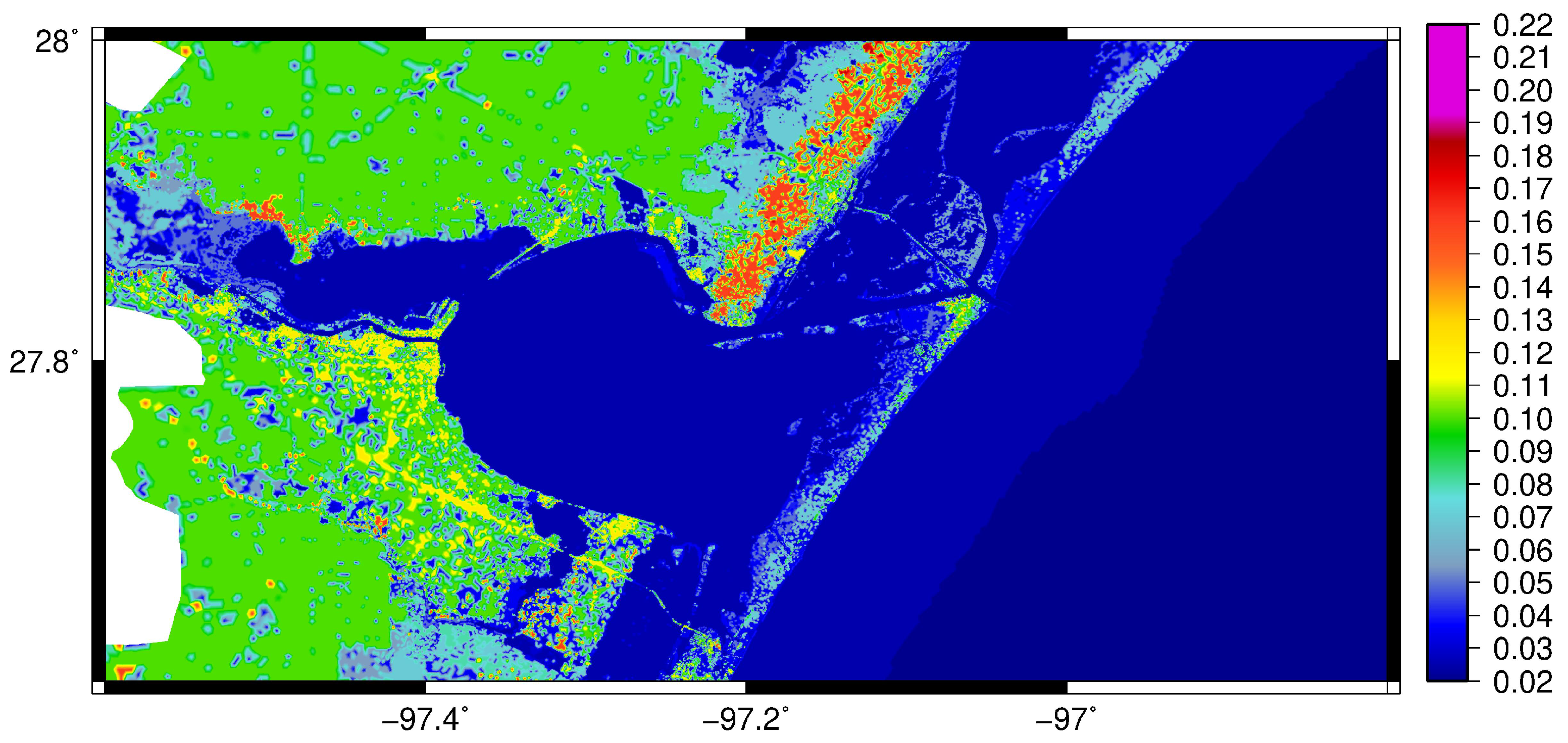
2.1. Hydrodynamics
2.2. Particle Transport
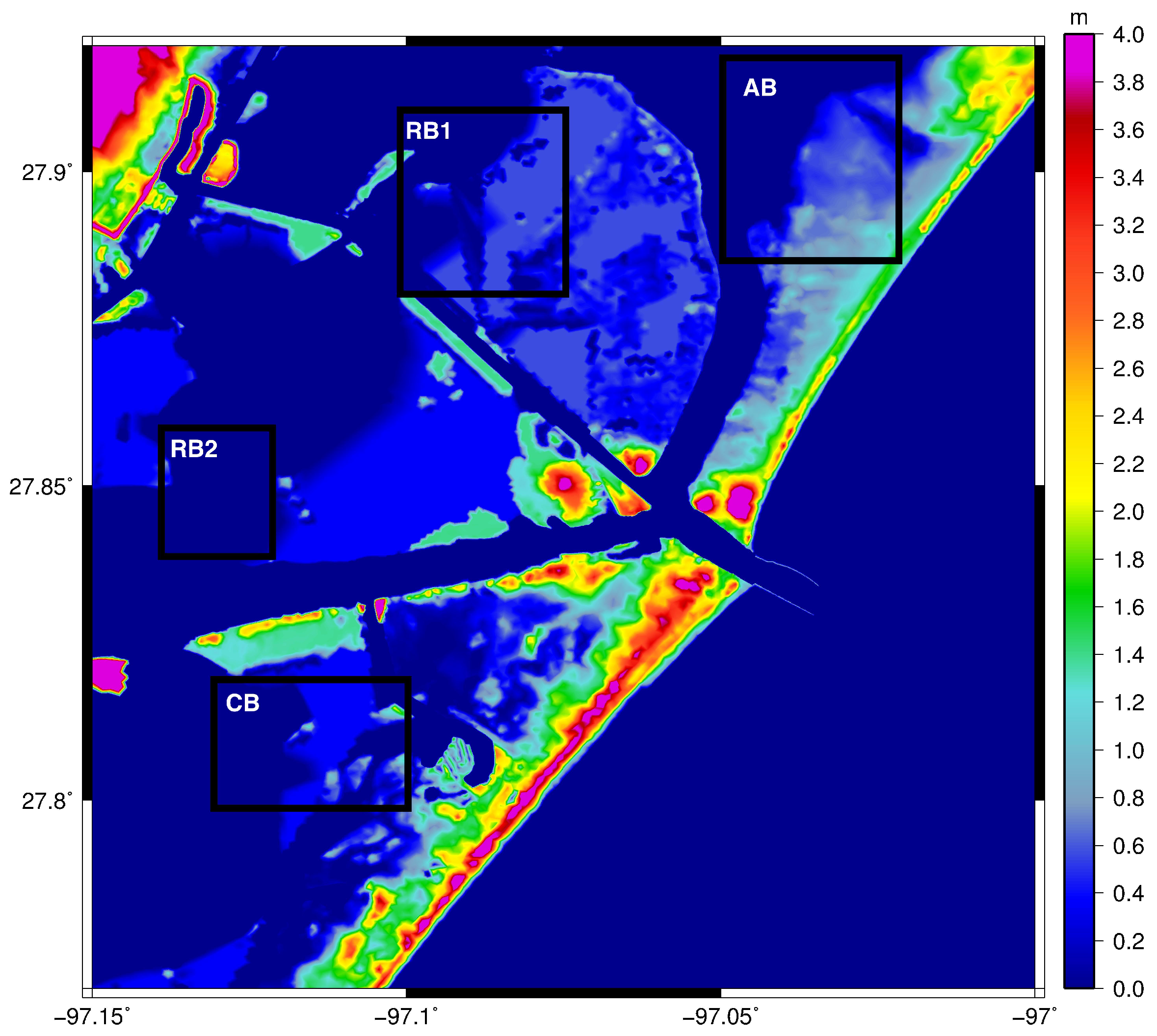
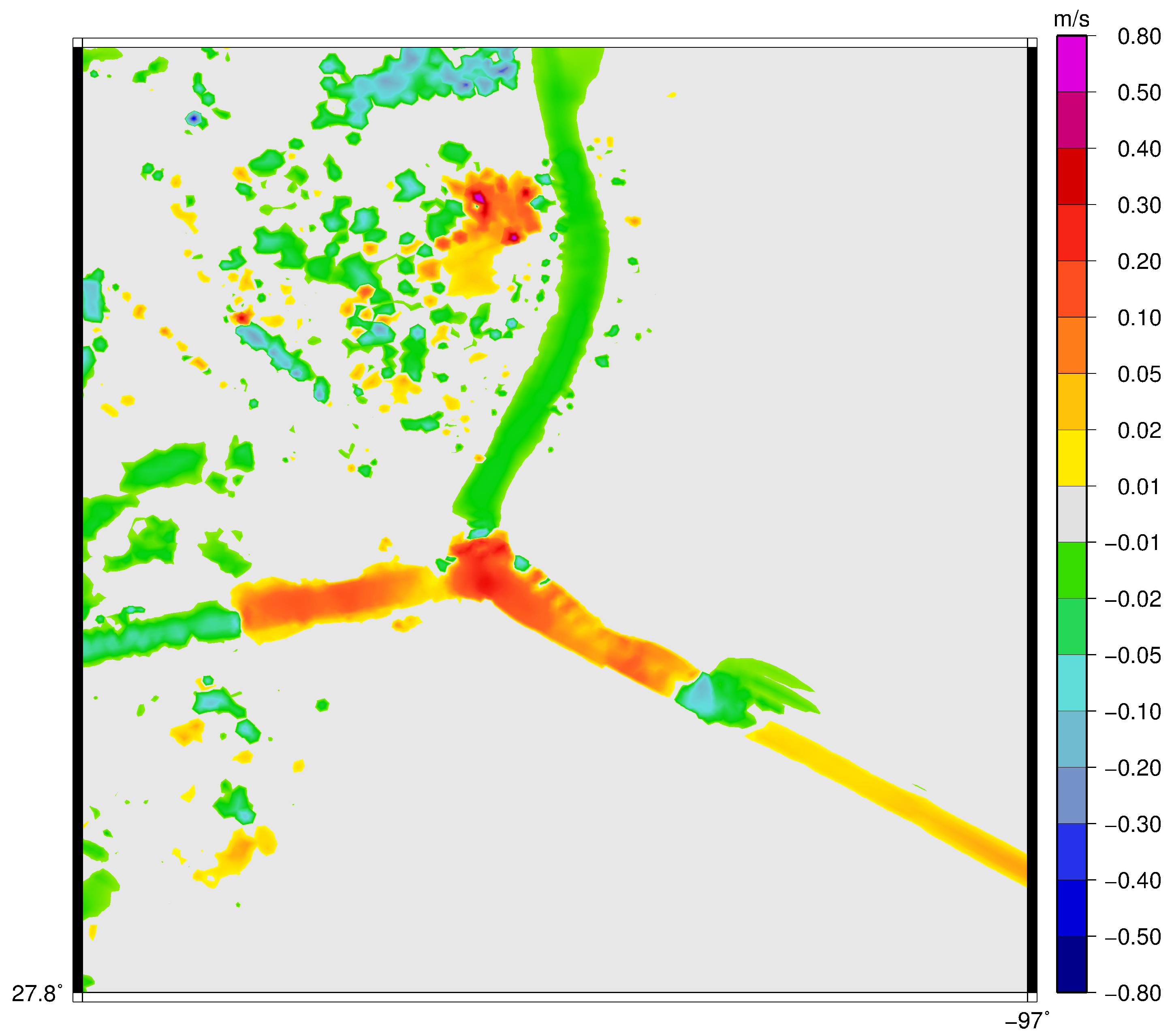
3. Results
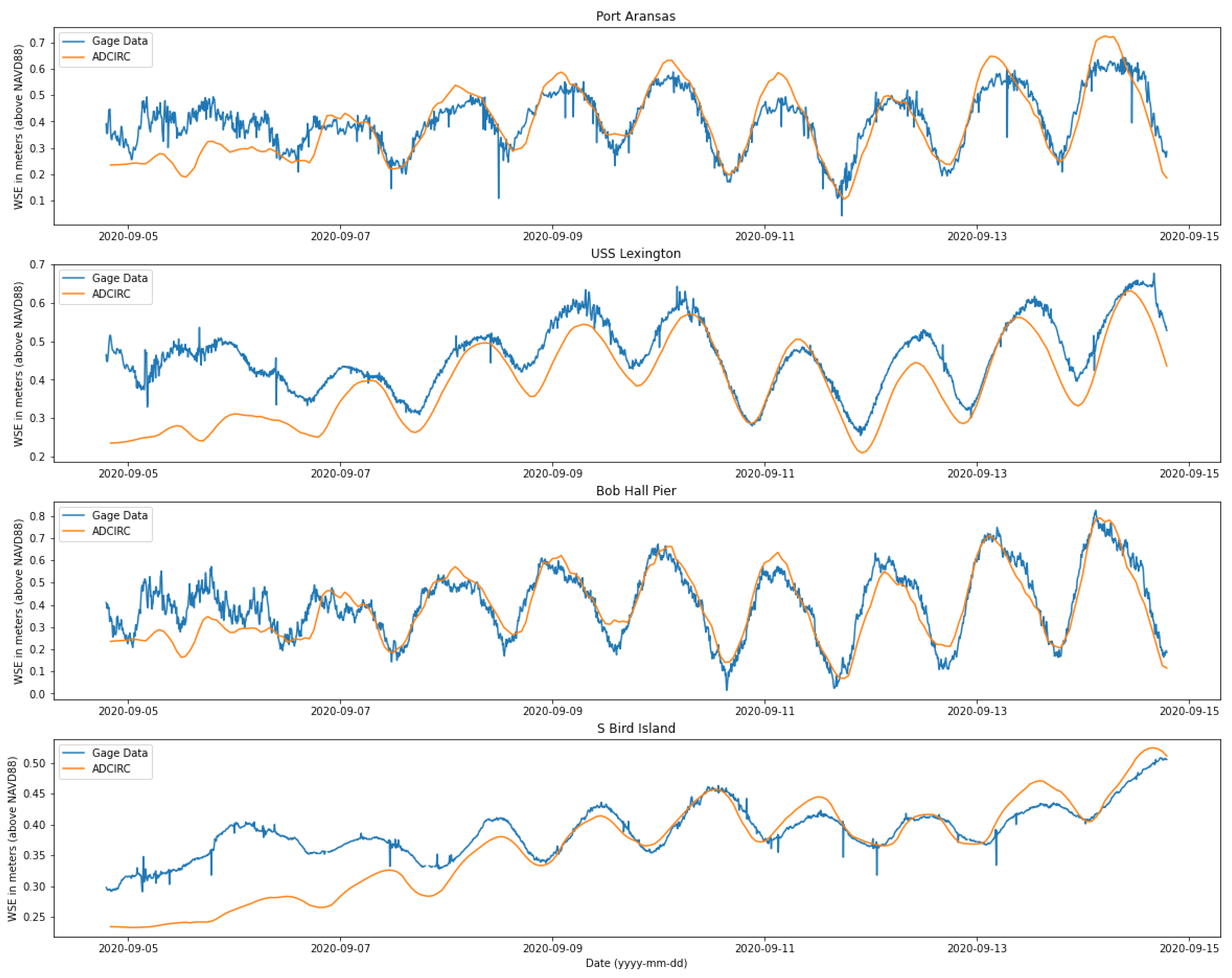
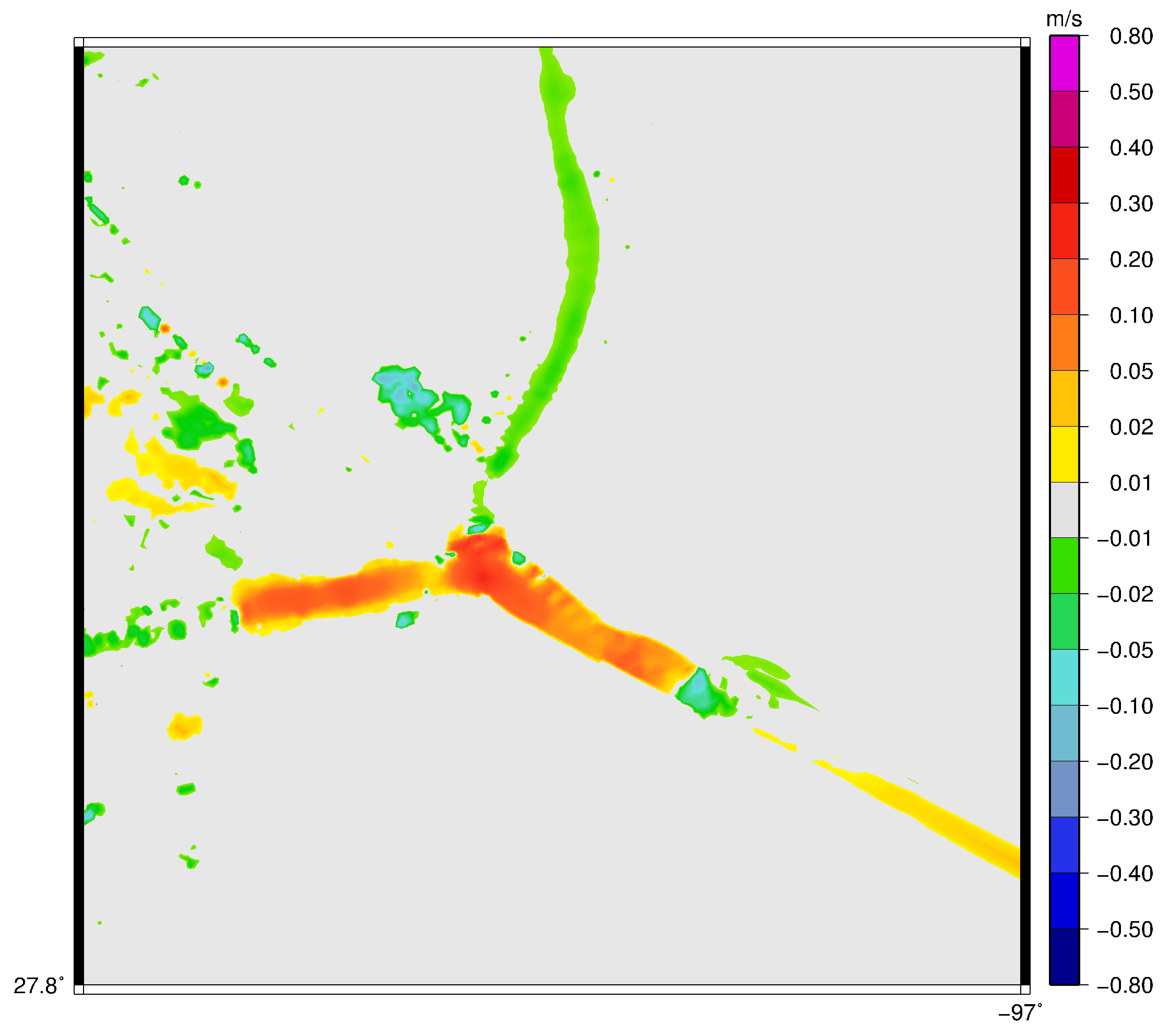
3.1. Velocity in the Aransas Pass
3.2. Particle Tracking
3.3. Influence of Extreme Flow Conditions
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hamilton, P.; Wood, E.; Lin, L.; Campbell, T.; Olson, L.; Jones, S.; Howard, S.; Skalbeck, K. Alternatives to Manage Sediment at the Intersection of the Gulf Intracoastal Waterway (GIWW) and the Corpus Christi Ship Channel (CCSC); Technical Report; Engineer Research and Development Center Vicksburg: Vicksbourg, MS, USA, 2018. [Google Scholar]
- Brown, C.; Holt, S.; Jackson, G.; Brooks, D.; Holt, G. Simulating larval supply to estuarine nursery areas: How important are physical processes to the supply of larvae to the Aransas Pass Inlet? Fish. Oceanogr. 2004, 13, 181–196. [Google Scholar] [CrossRef]
- Jenkins, G.P.; Black, K.P.; Wheatley, M.J.; Hatton, D.N. Temporal and spatial variability in recruitment of a temperate, seagrass-associated fish is largely determined by physical processes in the pre-and post-settlement phases. Mar. Ecol. Prog. Ser. 1997, 148, 23–35. [Google Scholar] [CrossRef]
- Matlock, G.C. The life history of the red drum. In Manual of Red Drum Aquaculture; Chamberlain, G.W., Miget, R.J., Haby, M.G., Eds.; Texas Agricultural Extension Service and Sea Grant College Program; Texas A&M University: College Station, TX, USA, 1987; pp. 1–47. [Google Scholar]
- Holt, G.J.; Holt, S.A.; Arnold, C.R. Diel periodicity of spawning in sciaenids. Mar. Ecol. Prog. Ser. 1985, 27, 7. [Google Scholar] [CrossRef]
- Holt, S.A.; Kitting, C.L.; Arnold, C.R. Distribution of young red drums among different sea-grass meadows. Trans. Am. Fish. Soc. 1983, 112, 267–271. [Google Scholar] [CrossRef]
- Leis, J.M.; Carson-Ewart, B.M. Orientation of pelagic larvae of coral-reef fishes in the ocean. Mar. Ecol. Prog. Ser. 2003, 252, 239–253. [Google Scholar] [CrossRef]
- Smith, N.P.; Stoner, A.W. Computer simulation of larval transport through tidal channels: Role of vertical migration. Estuar. Coast. Shelf Sci. 1993, 37, 43–58. [Google Scholar] [CrossRef]
- Bowden, K.; Hamilton, P. Some experiments with a numerical model of circulation and mixing in a tidal estuary. Estuar. Coast. Mar. Sci. 1975, 3, 281–301. [Google Scholar] [CrossRef]
- Brown, C.; Jackson, G.; Holt, S.; Holt, G. Spatial and temporal patterns in modeled particle transport to estuarine habitat with comparisons to larval fish settlement patterns. Estuar. Coast. Shelf Sci. 2005, 64, 33–46. [Google Scholar] [CrossRef]
- Brown, C.A.; Jackson, G.A.; Brooks, D.A. Particle transport through a narrow tidal inlet due to tidal forcing and implications for larval transport. J. Geophys. Res. Ocean. 2000, 105, 24141–24156. [Google Scholar] [CrossRef] [Green Version]
- Lynch, D.R.; Ip, J.T.; Naimie, C.E.; Werner, F.E. Comprehensive coastal circulation model with application to the Gulf of Maine. Cont. Shelf Res. 1996, 16, 875–906. [Google Scholar] [CrossRef]
- Lynch, D.R.; Gray, W.G. A wave equation model for finite element tidal computations. Comput. Fluids 1979, 7, 207–228. [Google Scholar] [CrossRef]
- Blanton, B. DROG3D: User’s Manual for 3-Dimensional Drogue Tracking on a Finite Element Grid with Linear Finite Elements; Program in Marine Sciences; University of North Carolina: Chapel Hill, NC, USA, 1995. [Google Scholar]
- Luettich, R.A.; Westerink, J.J.; Scheffner, N.W. ADCIRC: An Advanced Three-Dimensional Circulation Model for Shelves, Coasts, and Estuaries. Report 1, Theory and Methodology of ADCIRC-2DD1 and ADCIRC-3DL; Technical Report; Engineer Research and Development Center Vicksburg: Vicksbourg, MS, USA, 1992. [Google Scholar]
- Dietrich, J.; Trahan, C.; Howard, M.; Fleming, J.; Weaver, R.; Tanaka, S.; Yu, L.; Luettich, R., Jr.; Dawson, C.; Westerink, J.; et al. Surface trajectories of oil transport along the Northern Coastline of the Gulf of Mexico. Cont. Shelf Res. 2012, 41, 17–47. [Google Scholar] [CrossRef]
- Tan, W.Y. Shallow Water Hydrodynamics: Mathematical Theory and Numerical Solution for a Two-Dimensional System of Shallow-Water Equations; Elsevier: Amsterdam, The Netherlands, 1992. [Google Scholar]
- Manning, R.; Griffith, J.P.; Pigot, T.; Vernon-Harcourt, L.F. On the Flow of Water in Open Channels and Pipes; Transaction of the Institution of Civil Engineers of Ireland: Dublin, Ireland, 1890; Volume 20. [Google Scholar]
- Hope, M.E.; Westerink, J.J.; Kennedy, A.B.; Kerr, P.; Dietrich, J.C.; Dawson, C.; Bender, C.J.; Smith, J.; Jensen, R.E.; Zijlema, M.; et al. Hindcast and validation of Hurricane Ike (2008) waves, forerunner, and storm surge. J. Geophys. Res. Ocean. 2013, 118, 4424–4460. [Google Scholar] [CrossRef] [Green Version]
- Pringle, W. Oceanmesh2d: User Guide—Precise Distance-Based Two-Dimensional Automated Mesh Generation Toolbox Intended for Coastal Ocean/Shallow Water. 2018. Available online: https://www3.nd.edu/coast/reports_papers/2018-oceanmesh2d-user-guide.pdf (accessed on 7 October 2020).
- Egbert, E. Efficient inverse modeling of barotropic ocean tides. J. Atmos. Ocean. Technol. 2002, 19, 183–204. [Google Scholar] [CrossRef] [Green Version]
- Blain, C.A.; Rogers, W.E. Coastal Tide Prediction Using the adcirc-2ddi Hydrodynamic Finite Element Model: Model Validation and Sensitivity Analyses in the Southern North Sea/English Channel; Technical Report; Naval Research Lab Stennis Space Center MS Coastal and Semi-Enclosed Seas Section: Hancock County, MS, USA, 1998. [Google Scholar]
- Cyriac, R.; Dietrich, J.; Blain, C.; Dawson, C.; Dresback, K.; Fathi, A.; Bilskie, M.; Graber, H.; Hagen, S.; Kolar, R. Wind and tide effects on the Choctawhatchee Bay plume and implications for surface transport at Destin Inlet. Reg. Stud. Mar. Sci. 2020, 35, 101131. [Google Scholar] [CrossRef]
- Dawson, C.; Loveland, M.; Valseth, E. Potential Effects of Deepening of the Aransas Ship Channel on Particle Transport: Implications for Recruitment of Estuarine Dependent Larvae. 2021. Available online: https://utmsi.utexas.edu/images/MSI/Blog_Research/OdenInstitute_PotentialEffectsofDeepeningoftheAransasShipChannelonParticleTransport.pdf (accessed on 20 June 2021).
- Fuiman, L.; Smith, M.; Malley, V. Ontogeny of routine swimming speed and startle responses in red drum, with a comparison of responses to acoustic and visual stimuli. J. Fish Biol. 1999, 55, 215–226. [Google Scholar] [CrossRef]
- Forward, R.B.; Tankersley, R.A.; Welch, J.M. Selective tidal-stream transport of the blue crab Callinectes sapidus: An overview. Bull. Mar. Sci. 2003, 72, 347–365. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Initial Condition | Current Bathymetry | Proposed Bathymetry | % Difference |
|---|---|---|---|
| I | 442 | 410 | −7.24 |
| II | 144 | 106 | −26.39 |
| III | 351 | 381 | 8.55 |
| IV | 1265 | 1342 | 6.09 |
| Total | 2202 | 2239 | 1.54 |
| Initial Condition | Current Bathymetry | Proposed Bathymetry | % Difference |
|---|---|---|---|
| I | 285 | 294 | 2.81 |
| II | 38 | 27 | −28.95 |
| III | 209 | 216 | 3.35 |
| IV | 959 | 966 | 0.73 |
| Total | 1491 | 1503 | 0.80 |
| Monitoring Location | Current Bathymetry | Proposed Bathymetry | Difference |
|---|---|---|---|
| Initial condition I | |||
| AB | 375 | 333 | −42 |
| CB | 35 | 40 | +5 |
| RB1 | 20 | 26 | +6 |
| RB2 | 12 | 11 | −1 |
| Initial condition II | |||
| AB | 103 | 70 | −33 |
| CB | 28 | 22 | −6 |
| RB1 | 10 | 6 | −4 |
| RB2 | 3 | 8 | +5 |
| Initial condition III | |||
| AB | 278 | 301 | +23 |
| CB | 39 | 48 | +9 |
| RB1 | 19 | 24 | +5 |
| RB2 | 15 | 8 | +7 |
| Initial condition IV | |||
| AB | 918 | 997 | +79 |
| CB | 194 | 188 | −6 |
| RB1 | 98 | 87 | +9 |
| RB2 | 55 | 70 | +15 |
| Monitoring Location | Current Bathymetry | Proposed Bathymetry | Difference |
|---|---|---|---|
| Initial condition I | |||
| AB | 49 | 62 | +13 |
| CB | 135 | 149 | +14 |
| RB1 | 33 | 34 | +1 |
| RB2 | 68 | 48 | −20 |
| Initial condition II | |||
| AB | 15 | 8 | −7 |
| CB | 18 | 12 | −6 |
| RB1 | 2 | 4 | +2 |
| RB2 | 3 | 3 | - |
| Initial condition III | |||
| AB | 50 | 50 | - |
| CB | 82 | 91 | +9 |
| RB1 | 39 | 29 | −10 |
| RB2 | 38 | 46 | +8 |
| Initial condition IV | |||
| AB | 270 | 257 | −13 |
| CB | 378 | 407 | +29 |
| RB1 | 157 | 122 | −25 |
| RB2 | 154 | 180 | +26 |
| Initial Condition | Current Bathymetry | Proposed Bathymetry | % Difference |
|---|---|---|---|
| I | 166 | 161 | −3.0 |
| II | 23 | 20 | −13.0 |
| III | 125 | 144 | 15.2 |
| IV | 499 | 510 | 2.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valseth, E.; Loveland, M.D.; Dawson, C.; Buskey, E.J. A Study of the Potential Impact of Dredging the Corpus Christi Ship Channel on Passive Particle Transport. J. Mar. Sci. Eng. 2021, 9, 935. https://doi.org/10.3390/jmse9090935
Valseth E, Loveland MD, Dawson C, Buskey EJ. A Study of the Potential Impact of Dredging the Corpus Christi Ship Channel on Passive Particle Transport. Journal of Marine Science and Engineering. 2021; 9(9):935. https://doi.org/10.3390/jmse9090935
Chicago/Turabian StyleValseth, Eirik, Mark D. Loveland, Clint Dawson, and Edward J. Buskey. 2021. "A Study of the Potential Impact of Dredging the Corpus Christi Ship Channel on Passive Particle Transport" Journal of Marine Science and Engineering 9, no. 9: 935. https://doi.org/10.3390/jmse9090935
APA StyleValseth, E., Loveland, M. D., Dawson, C., & Buskey, E. J. (2021). A Study of the Potential Impact of Dredging the Corpus Christi Ship Channel on Passive Particle Transport. Journal of Marine Science and Engineering, 9(9), 935. https://doi.org/10.3390/jmse9090935