
Iron-Coupled Anaerobic Oxidation of Methane in Marine Sediments: A Review



{kind=link}
{kind=link}
Abstract
:1. Introduction
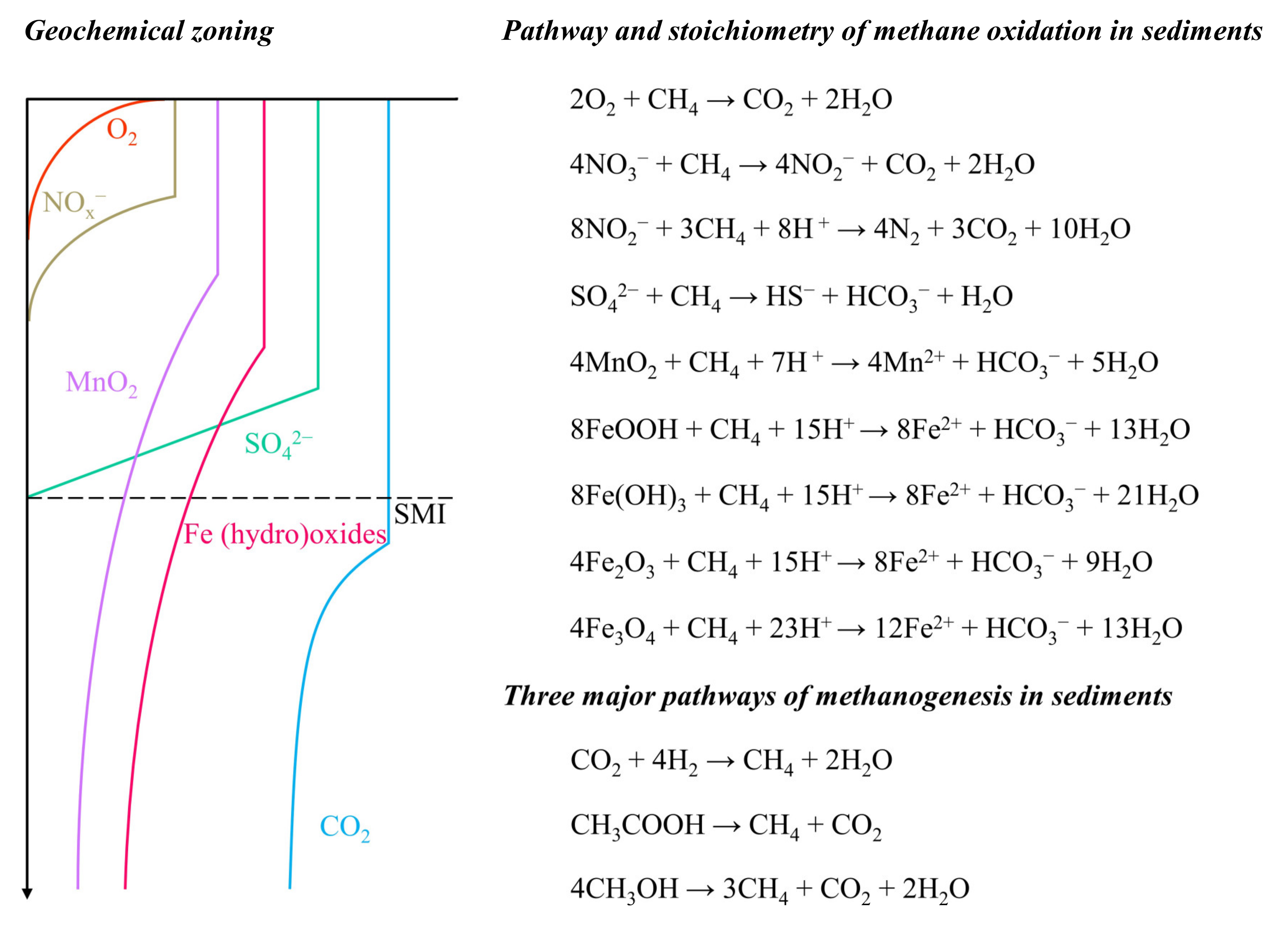
1.1. Conditions for Fe-AOM in Marine Sediments
1.2. Potential Forms of Iron Oxides for Fe-AOM
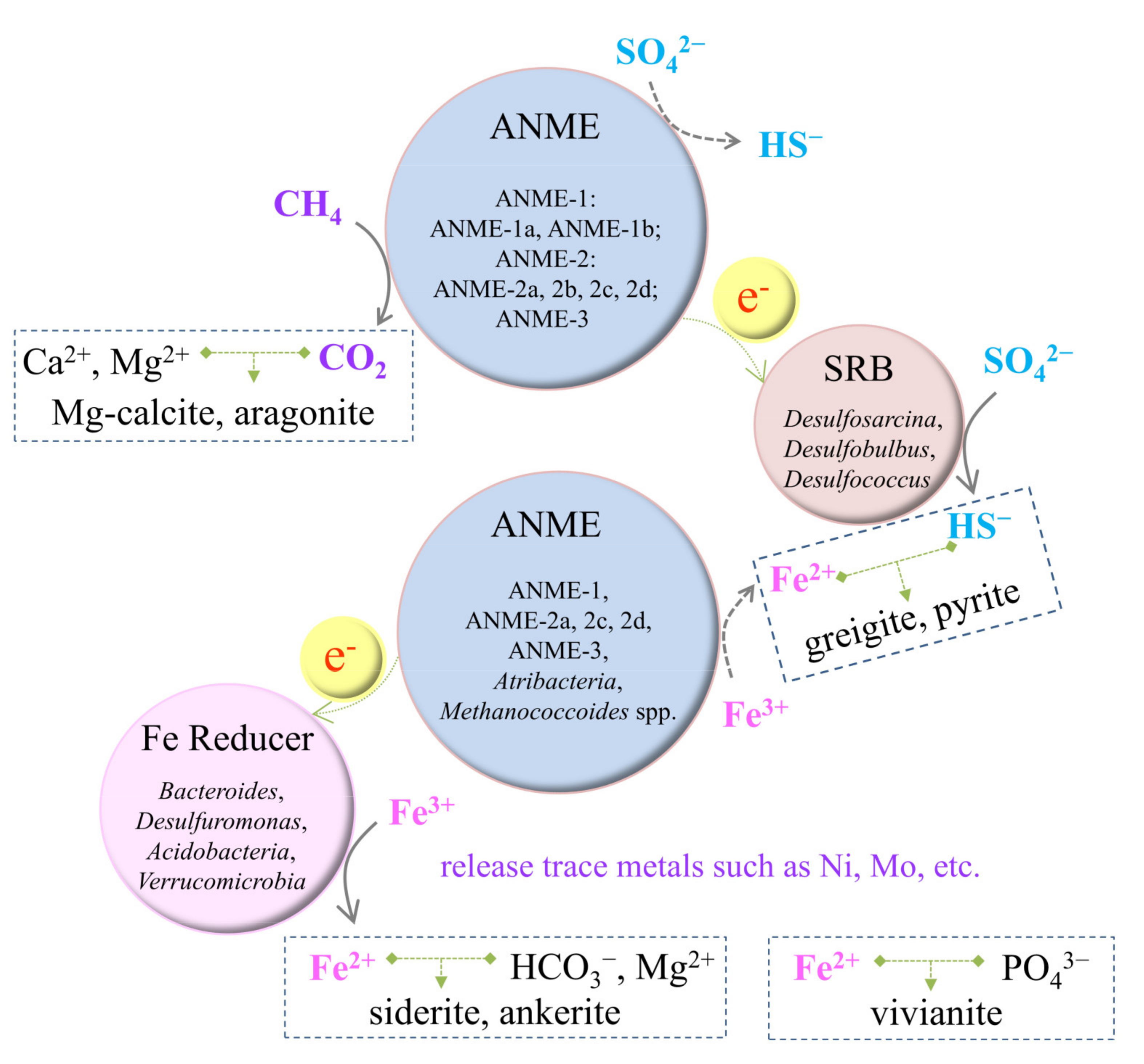
1.3. Microbial Mechanisms
2. Geochemical and Environmental Significance
2.1. Impact on Methane Dynamics
2.2. Impact on Iron Cycling
2.3. Impact on Calcium, Sulfur, Phosphorus, and Other Elements
3. Conclusions and Challenges
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reeburgh, W.S. Oceanic methane biogeochemistry. Chem. Rev. 2007, 107, 486–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciais, P.; Sabine, C.; Bala, G.; Bopp, L.; Brovkin, V.; Canadell, J.; Chhabra, A.; Defries, R.; Galloway, J.; Heimann, M.; et al. Carbon and other biogeochemical cycles. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; pp. 465–570. [Google Scholar]
- Weller, S.; Kraus, D.; Ayag, K.R.P.; Wassmann, R.; Alberto, M.C.R.; Butterbach-Bahl, K.; Kiese, R. Methane and nitrous oxide emissions from rice and maize production in diversified rice cropping systems. Nutr. Cycl. Agroecosyst. 2015, 101, 37–53. [Google Scholar] [CrossRef]
- Shakoor, A.; Ashraf, F.; Shakoor, S.; Mustafa, A.; Rehman, A.; Altaf, M.M. Biogeochemical transformation of greenhouse gas emissions from terrestrial to atmospheric environment and potential feedback to climate forcing. Environ. Sci. Pollut. Res. 2020, 27, 38513–38536. [Google Scholar] [CrossRef]
- Fernandez-Cortes, A.; Cuezva, S.; Alvarez-Gallego, M.; Garcia-Anton, E.; Pla, C.; Benavente, D.; Jurado, V.; Saiz-Jimenez, C.; Sanchez-Moral, S. Subterranean atmospheres may act as daily methane sinks. Nat. Commun. 2015, 6, 7003. [Google Scholar] [CrossRef] [Green Version]
- Bohrmann, G.; Torres, M.E. Methane in marine sediments. In Encyclopedia of Marine Geosciences; Harff, J., Meschede, M., Petersen, S., Thiede, J., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 1–7. [Google Scholar]
- Hinrichs, K.U.; Boetius, A. The anaerobic oxidation of methane: New insights in microbial ecology and biogeochemistry. In Ocean Margin Systems; Wefer, G., Billett, D., Hebbeln, D., Jørgensen, B.B., Schlüter, M., van Weering, T.C.E., Eds.; Springer: Berlin/Heidelberg, Germany, 2003; pp. 457–477. [Google Scholar]
- Iversen, N.; Jorgensen, B.B. Anaerobic methane oxidation rates at the sulfate-methane transition in marine sediments from Kattegat and Skagerrak (Denmark). Limnol. Oceanogr. 1985, 30, 944–955. [Google Scholar] [CrossRef]
- Boetius, A.; Ravenschlag, K.; Schubert, C.J.; Rickert, D.; Widdel, F.; Gieseke, A.; Amann, R.; Jørgensen, B.B.; Witte, U.; Pfannkuche, O. A marine microbial consortium apparently mediating anaerobic oxidation of methane. Nature 2000, 407, 623–626. [Google Scholar] [CrossRef] [PubMed]
- Wallenius, A.J.; Dalcin Martins, P.; Slomp, C.P.; Jetten, M.S.M. Anthropogenic and environmental constraints on the microbial methane cycle in coastal sediments. Front. Microbiol. 2021, 12, 631621. [Google Scholar] [CrossRef]
- Beal, E.J.; House, C.H.; Orphan, V.J. Manganese- and iron-dependent marine methane oxidation. Science 2009, 325, 184–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slomp, C.P.; Mort, H.P.; Jilbert, T.; Reed, D.C.; Gustafsson, B.G.; Wolthers, M. Coupled dynamics of iron and phosphorus in sediments of an ooligotrophic coastal basin and the impact of anaerobic oxidation of methane. PLoS ONE 2013, 8, e62386. [Google Scholar] [CrossRef]
- Savvichev, A.S.; Kokryatskaya, N.M.; Zabelina, S.A.; Rusanov, I.I.; Zakharova, E.E.; Veslopolova, E.F.; Lunina, O.N.; Patutina, E.O.; Bumazhkin, B.K.; Gruzdev, D.S.; et al. Microbial processes of the carbon and sulfur cycles in an ice-covered, iron-rich meromictic lake Svetloe (Arkhangelsk region, Russia). Environ. Microbiol. 2017, 19, 659–672. [Google Scholar] [CrossRef]
- Crowe, S.A.; Katsev, S.; Leslie, K.; Sturm, A.; Magen, C.; Nomosatryo, S.; Pack, M.A.; Kessler, J.D.; Reeburgh, W.S.; Roberts, J.A.; et al. The methane cycle in ferruginous Lake Matano. Geobiology 2011, 9, 61–78. [Google Scholar] [CrossRef]
- Amos, R.T.; Bekins, B.A.; Cozzarelli, I.M.; Voytek, M.A.; Kirshtein, J.D.; Jones, E.J.P.; Blowes, D.W. Evidence for iron-mediated anaerobic methane oxidation in a crude oil-contaminated aquifer. Geobiology 2012, 10, 506–517. [Google Scholar] [CrossRef]
- Wankel, S.D.; Adams, M.M.; Johnston, D.T.; Hansel, C.M.; Joye, S.B.; Girguis, P.R. Anaerobic methane oxidation in metalliferous hydrothermal sediments: Influence on carbon flux and decoupling from sulfate reduction. Environ. Microbial. 2012, 14, 2726–2740. [Google Scholar] [CrossRef]
- Nordi, K.A.; Thamdrup, B.; Schubert, C.J. Anaerobic oxidation of methane in an iron-rich Danish freshwater lake sediment. Limnol. Oceanogr. 2013, 58, 546–554. [Google Scholar] [CrossRef]
- Maignien, L.; Parkes, R.J.; Cragg, B.; Niemann, H.; Knittel, K.; Coulon, S.; Akhmetzhanov, A.; Boon, N. Anaerobic oxidation of methane in hypersaline cold seep sediments. FEMS Microbiol. Ecol. 2013, 83, 214–231. [Google Scholar] [CrossRef]
- Segarra, K.E.A.; Comerford, C.; Slaughter, J.; Joye, S.B. Impact of electron acceptor availability on the anaerobic oxidation of methane in coastal freshwater and brackish wetland sediments. Geochim. Cosmochim. Acta 2013, 115, 15–30. [Google Scholar] [CrossRef]
- Riedinger, N.; Formolo, M.J.; Lyons, T.W.; Henkel, S.; Beck, A.; Kasten, S. An inorganic geochemical argument for coupled anaerobic oxidation of methane and iron reduction in marine sediments. Geobiology 2014, 12, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Treude, T.; Krause, S.; Maltby, J.; Dale, A.W.; Coffin, R.; Hamdan, L.J. Sulfate reduction and methane oxidation activity below the sulfate-methane transition zone in Alaskan Beaufort Sea continental margin sediments: Implications for deep sulfur cycling. Geochim. Cosmochim. Acta 2014, 144, 217–237. [Google Scholar] [CrossRef] [Green Version]
- Oni, O.; Miyatake, T.; Kasten, S.; Richter-Heitmann, T.; Fischer, D.; Wagenknecht, L.; Kulkarni, A.; Blumers, M.; Shylin, S.I.; Ksenofontov, V.; et al. Distinct microbial populations are tightly linked to the profile of dissolved iron in the methanic sediments of the Helgoland mud area, North Sea. Front. Microbial. 2015, 6, 365. [Google Scholar] [CrossRef]
- Egger, M.; Hagens, M.; Sapart, C.J.; Dijkstra, N.; van Helmond, N.A.G.M.; Mogollon, J.M.; Risgaard-Petersen, N.; van der Veen, C.; Kasten, S.; Riedinger, N.; et al. Iron oxide reduction in methane-rich deep Baltic Sea sediments. Geochim. Cosmochim. Acta 2017, 207, 256–276. [Google Scholar] [CrossRef]
- Vigderovich, H.; Liang, L.; Herut, B.; Wang, F.; Wurgaft, E.; Rubin-Blum, M.; Sivan, O. Evidence for microbial iron reduction in the methanic sediments of the oligotrophic southeastern Mediterranean continental shelf. Biogeosciences 2019, 16, 3165–3181. [Google Scholar] [CrossRef] [Green Version]
- Egger, M.; Rasigraf, O.; Sapart, C.J.; Jilbert, T.; Jetten, M.S.M.; Rockmann, T.; van der Veen, C.; Banda, N.; Kartal, B.; Ettwig, K.F. Iron-mediated anaerobic oxidation of methane in brackish coastal sediments. Environ. Sci. Technol. 2015, 49, 277–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rooze, J.; Egger, M.; Tsandev, I.; Slomp, C.P. Iron-dependent anaerobic oxidation of methane in coastal surface sediments: Potential controls and impact. Limnol. Oceanogr. 2016, 61, S267–S282. [Google Scholar] [CrossRef] [Green Version]
- Egger, M.; Kraal, P.; Jilbert, T.; Sulu-Gambari, F.; Sapart, C.J.; Röckmann, T.; Slomp, C.P. Anaerobic oxidation of methane alters sediment records of sulfur, iron and phosphorus in the Black Sea. Biogeosciences 2016, 13, 5333–5355. [Google Scholar] [CrossRef] [Green Version]
- Aromokeye, D.A.; Kulkarni, A.C.; Elvert, M.; Wegener, G.; Henkel, S.; Coffinet, S.; Eickhorst, T.; Oni, O.E.; Richter-Heitmann, T.; Schnakenberg, A.; et al. Rates and microbial players of iron-driven anaerobic oxidation of methane in methanic marine sediments. Front. Microbiol. 2019, 10, 3041. [Google Scholar] [CrossRef] [PubMed]
- Crowe, S.A.; Jones, C.; Katsev, S.; Magen, C.; O’Neill, A.H.; Sturm, A.; Canfield, D.E.; Haffner, G.D.; Mucci, A.; Sundby, B.; et al. Photoferrotrophs thrive in an Archean Ocean analogue. Proc. Natl. Acad. Sci. USA 2008, 105, 15938–15943. [Google Scholar] [CrossRef] [Green Version]
- Sivan, O.; Adler, M.; Pearson, A.; Gelman, F.; Bar-Or, I.; John, S.G.; Eckerte, W. Geochemical evidence for iron-mediated anaerobic oxidation of methane. Limnol. Oceanogr. 2011, 56, 1536–1544. [Google Scholar] [CrossRef] [Green Version]
- Sivan, O.; Antler, G.; Turchyn, A.V.; Marlow, J.J.; Orphan, V.J. Iron oxides stimulate sulfate-driven anaerobic methane oxidation in seeps. Proc. Natl. Acad. Sci. USA 2014, 111, E4139–E4147. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Lu, H.; Ruffine, L. Geochemical characteristics of iron in sediments from the Sea of Marmara. Deep Sea Res. Part II 2018, 153, 121–130. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Izon, G.; Wang, J.; Antler, G.; Wang, Z.; Zhao, J.; Egger, M. Vivianite formation in methane-rich deep-sea sediments from the South China Sea. Biogeosciences 2018, 15, 6329–6348. [Google Scholar] [CrossRef] [Green Version]
- Roberts, A.P. Magnetic mineral diagenesis. Earth Sci. Rev. 2015, 151, 1–47. [Google Scholar] [CrossRef]
- Ettwig, K.F.; Zhu, B.; Speth, D.; Keltjens, J.T.; Jetten, M.S.M.; Kartal, B. Archaea catalyze iron-dependent anaerobic oxidation of methane. Proc. Natl. Acad. Sci. USA 2016, 113, 12792–12796. [Google Scholar] [CrossRef] [Green Version]
- Scheller, S.; Yu, H.; Chadwick, G.L.; McGlynn, S.E.; Orphan, V.J. Artificial electron acceptors decouple archaeal methane oxidation from sulfate reduction. Science 2016, 351, 703–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovley, D.R.; Phillips, E.J. Novel mode of microbial energy metabolism: Organic carbon oxidation coupled to dissimilatory reduction of iron or manganese. Appl. Environ. Microb. 1988, 54, 1472–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bligh, M.W.; Waite, T.D. Formation, aggregation and reactivity of amorphous ferric oxyhydroxides on dissociation of Fe(III)-organic complexes in dilute aqueous suspensions. Geochim. Cosmochim. Acta 2010, 74, 5746–5762. [Google Scholar] [CrossRef]
- Ding, L.-J.; Su, J.-Q.; Xu, H.-J.; Jia, Z.-J.; Zhu, Y.-G. Long-term nitrogen fertilization of paddy soil shifts iron-reducing microbial community revealed by RNA-C-13-acetate probing coupled with pyrosequencing. ISME J. 2015, 9, 721–734. [Google Scholar] [CrossRef]
- Hori, T.; Aoyagi, T.; Itoh, H.; Narihiro, T.; Oikawa, A.; Suzuki, K.; Ogata, A.; Friedrich, M.W.; Conrad, R.; Kamagata, Y. Isolation of microorganisms involved in reduction of crystalline iron(III) oxides in natural environments. Front. Microbial. 2015, 6, 386. [Google Scholar] [CrossRef] [Green Version]
- Bosch, J.; Heister, K.; Hofmann, T.; Meckenstock, R.U. Nanosized iron oxide colloids strongly enhance microbial iron reduction. Appl. Environ. Microb. 2010, 76, 184–189. [Google Scholar] [CrossRef] [Green Version]
- Boetius, A.; Wenzhoefer, F. Seafloor oxygen consumption fuelled by methane from cold seeps. Nat. Geosci. 2013, 6, 725–734. [Google Scholar] [CrossRef]
- Ruff, S.E.; Biddle, J.F.; Teske, A.P.; Knittel, K.; Boetius, A.; Ramette, A. Global dispersion and local diversification of the methane seep microbiome. Proc. Natl. Acad. Sci. USA 2015, 112, 4015–4020. [Google Scholar] [CrossRef] [Green Version]
- Orphan, V.J.; Hinrichs, K.U.; Ussler, W.; Paull, C.K.; Taylor, L.T.; Sylva, S.P.; Hayes, J.M.; Delong, E.F. Comparative analysis of methane-oxidizing archaea and sulfate-reducing bacteria in anoxic marine sediments. Appl. Environ. Microbiol. 2001, 67, 1922–1934. [Google Scholar] [CrossRef] [Green Version]
- Esther, J.; Sukla, L.B.; Pradhan, N.; Panda, S. Fe(III) reduction strategies of dissimilatory iron reducing bacteria. Korean J. Chem. Eng. 2015, 32, 1–14. [Google Scholar] [CrossRef]
- Weber, H.S.; Habicht, K.S.; Thamdrup, B. Anaerobic Methanotrophic Archaea of the ANME-2d cluster are active in a low-sulfate, iron-rich freshwater sediment. Front. Microbiol. 2017, 8, 619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, C.; Leu, A.O.; Xie, G.-J.; Guo, J.; Feng, Y.; Zhao, J.-X.; Tyson, G.W.; Yuan, Z.; Hu, S. A methanotrophic archaeon couples anaerobic oxidation of methane to Fe(III) reduction. ISME J. 2018, 12, 1929–1939. [Google Scholar] [CrossRef] [PubMed]
- Roland, F.A.E.; Borges, A.V.; Darchambeau, F.; Lliros, M.; Descy, J.-P.; Morana, C. The possible occurrence of iron-dependent anaerobic methane oxidation in an Archean Ocean analogue. Sci. Rep. 2021, 11, 1597. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Li, S.W.; Ding, Z.W.; Ding, J.; Lu, Y.Z.; Zeng, R.J. Iron reduction in the DAMO/Shewanella oneidensis MR-1 coculture system and the fate of Fe(II). Water Res. 2016, 88, 808–815. [Google Scholar] [CrossRef]
- Folgosa, F.; Tavares, P.; Pereira, A.S. Iron management and production of electricity by microorganisms. Appl. Microbiol. Biotechnol. 2015, 99, 8329–8336. [Google Scholar] [CrossRef]
- Wegener, G.; Krukenberg, V.; Riedel, D.; Tegetmeyer, H.E.; Boetius, A. Intercellular wiring enables electron transfer between methanotrophic archaea and bacteria. Nature 2015, 526, 587–590. [Google Scholar] [CrossRef]
- Konhauser, K.O.; Newman, D.K.; Kappler, A. The potential significance of microbial Fe(III) reduction during deposition of Precambrian banded iron formations. Geobiology 2005, 3, 167–177. [Google Scholar] [CrossRef]
- Lyu, Z.; Shao, N.; Akinyemi, T.; Whitman, W.B. Methanogenesis. Curr. Biol. 2018, 28, R727–R732. [Google Scholar] [CrossRef] [Green Version]
- Reiche, M.; Torburg, G.; Kuesel, K. Competition of Fe(III) reduction and methanogenesis in an acidic fen. FEMS Microbiol. Ecol. 2008, 65, 88–101. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Xu, J.; Yang, G.; Zhuang, L. Methanogenesis affected by the co-occurrence of iron(III) oxides and humic substances. FEMS Microbiol. Ecol. 2014, 88, 107–120. [Google Scholar] [CrossRef]
- Zhuang, G.-C.; Heuer, V.B.; Lazar, C.S.; Goldhammer, T.; Wendt, J.; Samarkin, V.A.; Elvert, M.; Teske, A.P.; Joye, S.B.; Hinrichs, K.-U. Relative importance of methylotrophic methanogenesis in sediments of the Western Mediterranean Sea. Geochim. Cosmochim. Acta 2018, 224, 171–186. [Google Scholar] [CrossRef]
- Li, J.; Peng, X.; Bai, S.; Chen, Z.; Nostrand, J.V. Biogeochemical processes controlling authigenic carbonate formation within the sediment column from the okinawa trough. Geochim. Cosmochim. Acta 2018, 222, 363–382. [Google Scholar] [CrossRef]
- Ahmed, M.; Lin, L.-S. Ferric reduction in organic matter oxidation and its applicability for anaerobic wastewater treatment: A review and future aspects. Rev. Environ. Sci. Bio/Techol. 2017, 16, 273–287. [Google Scholar] [CrossRef]
- Weber, K.A.; Achenbach, L.A.; Coates, J.D. Microorganisms pumping iron: Anaerobic microbial iron oxidation and reduction. Nat. Rev. Microbiol. 2006, 4, 752–764. [Google Scholar] [CrossRef] [Green Version]
- Maerz, C.; Hoffmann, J.; Bleil, U.; De Lange, G.J.; Kasten, S. Diagenetic changes of magnetic and geochemical signals by anaerobic methane oxidation in sediments of the Zambezi deep-sea fan (SW Indian Ocean). Mar. Geol. 2008, 255, 118–130. [Google Scholar] [CrossRef]
- Lim, Y.C.; Lin, S.; Yang, T.F.; Chen, Y.-G.; Liu, C.-S. Variations of methane induced pyrite formation in the accretionary wedge sediments offshore southwestern Taiwan. Mar. Pet. Geol. 2011, 28, 1829–1837. [Google Scholar] [CrossRef]
- Hsu, T.-W.; Jiang, W.-T.; Wang, Y. Authigenesis of vivianite as influenced by methane-induced sulfidization in cold-seep sediments off southwestern Taiwan. J. Asian Earth Sci. 2014, 89, 88–97. [Google Scholar] [CrossRef]
- Sun, Z.; Wei, H.; Zhang, X.; Shang, L.; Yin, X.; Sun, Y.; Xu, L.; Huang, W.; Zhang, X. A unique Fe-rich carbonate chimney associated with cold seeps in the Northern Okinawa Trough, East China Sea. Deep Sea Res. Part I 2015, 95, 37–53. [Google Scholar] [CrossRef]
- Peng, X.; Guo, Z.; Chen, S.; Sun, Z.; Xu, H.; Ta, K.; Zhang, J.; Zhang, L.; Li, J.; Du, M. Formation of carbonate pipes in the northern Okinawa Trough linked to strong sulfate exhaustion and iron supply. Geochim. Cosmochim. Acta 2017, 205, 1–13. [Google Scholar] [CrossRef]
- Roberts, A.P.; Weaver, R. Multiple mechanisms of remagnetization involving sedimentary greigite (Fe3S4). Earth Planet. Sci. Lett. 2005, 231, 263–277. [Google Scholar] [CrossRef]
- Lin, Z.; Sun, X.; Harald, S.; Lu, Y.; Böttcher, M.E.; Teichert, B.; Gong, J.; Li, X.; Liang, J.; Lu, H. Multiple sulfur isotopic evidence for the origin of elemental sulfur in an iron-dominated gas hydrate-bearing sedimentary environment. Mar. Geol. 2018, 403, 271–284. [Google Scholar] [CrossRef]
- Wijsman, J.W.M.; Middelburg, J.J.; Heip, C.H.R. Reactive iron in Black Sea sediments: Implications for iron cycling. Mar. Geol. 2001, 172, 167–180. [Google Scholar] [CrossRef]
- Lyons, T.W.; Severmann, S. A critical look at iron paleoredox proxies: New insights from modern euxinic marine basins. Geochim. Cosmochim. Acta 2006, 70, 5698–5722. [Google Scholar] [CrossRef]
- Raiswell, R.; Canfield, D.E. The iron biogeochemical cycle past and present. Geochem. Perspect. 2012, 1, 1–220. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Xu, T.; Shang, S.; Tian, H.; Cao, Y.; Wang, J.; Shi, Z.; Liu, X. Laboratory experimental study on the formation of authigenic carbonates induced by microbes in marine sediments. J. Mar. Sci. Eng. 2021, 9, 479. [Google Scholar] [CrossRef]
- Burstein, G.T. The iron oxides: Structure, properties, reactions, occurrence, and uses. Corros. Sci. 1997, 39, 1499–1500. [Google Scholar] [CrossRef]
- Ma, G.; Zhan, L.; Lu, H.; Hou, G. Structures in shallow marine sediments associated with gas and fluid migration. J. Mar. Sci. Eng. 2021, 9, 396. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Yu, S.; Lu, H. Iron-Coupled Anaerobic Oxidation of Methane in Marine Sediments: A Review. J. Mar. Sci. Eng. 2021, 9, 875. https://doi.org/10.3390/jmse9080875
Yang H, Yu S, Lu H. Iron-Coupled Anaerobic Oxidation of Methane in Marine Sediments: A Review. Journal of Marine Science and Engineering. 2021; 9(8):875. https://doi.org/10.3390/jmse9080875
Chicago/Turabian StyleYang, Hailin, Shan Yu, and Hailong Lu. 2021. "Iron-Coupled Anaerobic Oxidation of Methane in Marine Sediments: A Review" Journal of Marine Science and Engineering 9, no. 8: 875. https://doi.org/10.3390/jmse9080875
APA StyleYang, H., Yu, S., & Lu, H. (2021). Iron-Coupled Anaerobic Oxidation of Methane in Marine Sediments: A Review. Journal of Marine Science and Engineering, 9(8), 875. https://doi.org/10.3390/jmse9080875