
An Autonomous Platform for Near Real-Time Surveillance of Harmful Algae and Their Toxins in Dynamic Coastal Shelf Environments

 ,
,
Abstract
1. Introduction
2. Materials and Methods
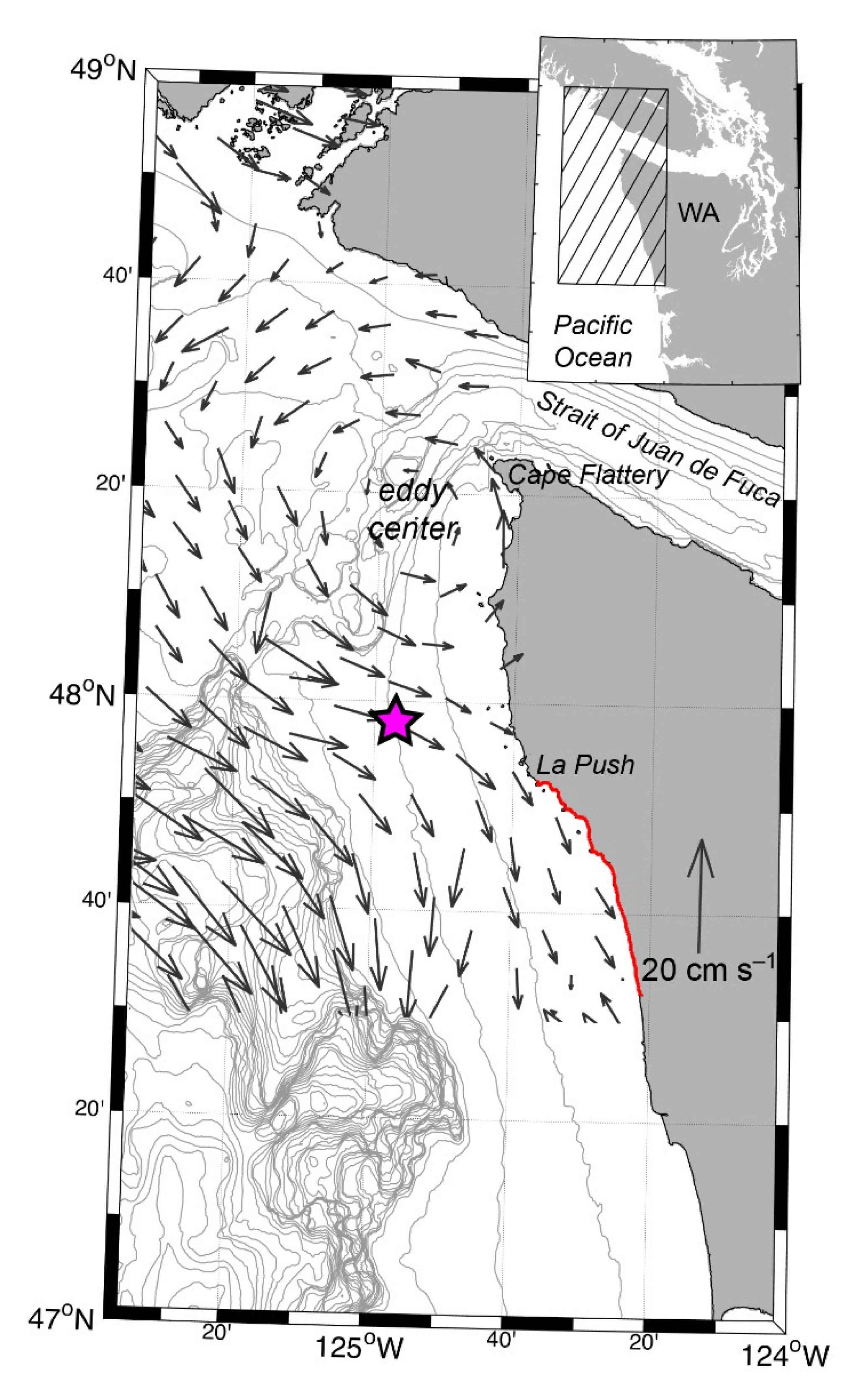
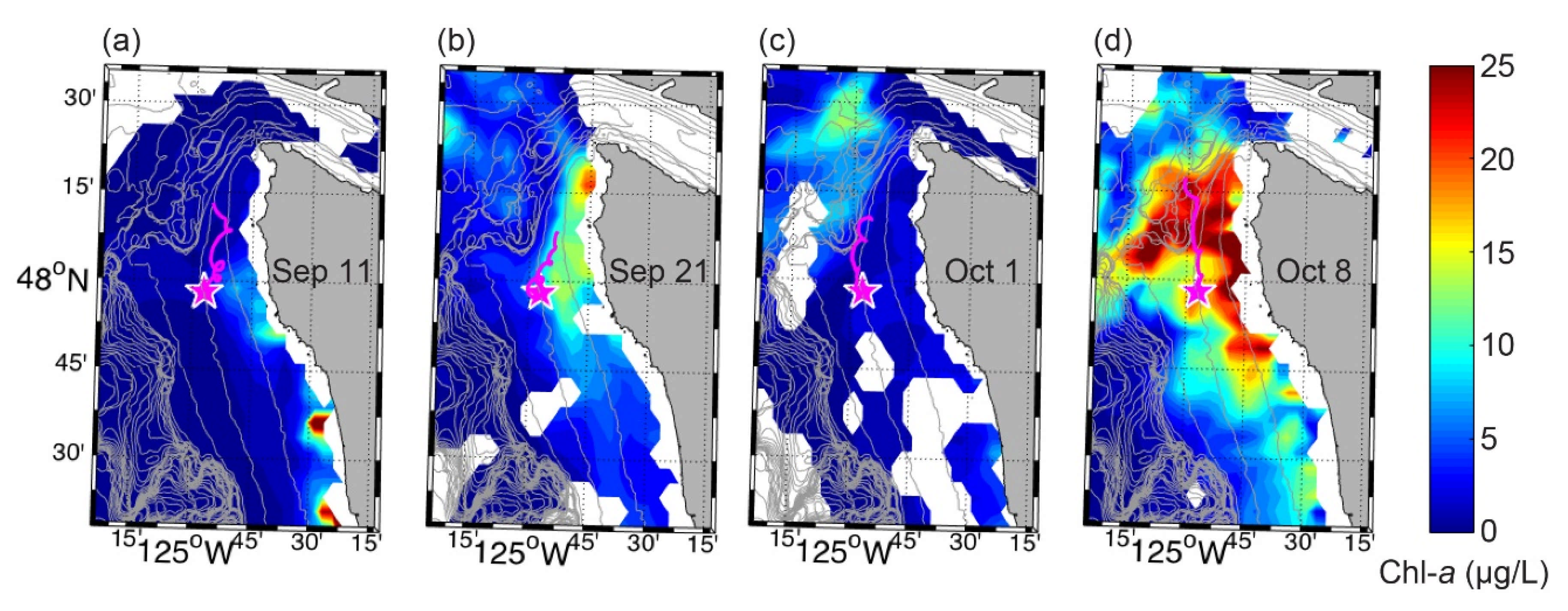
2.1. Study Region
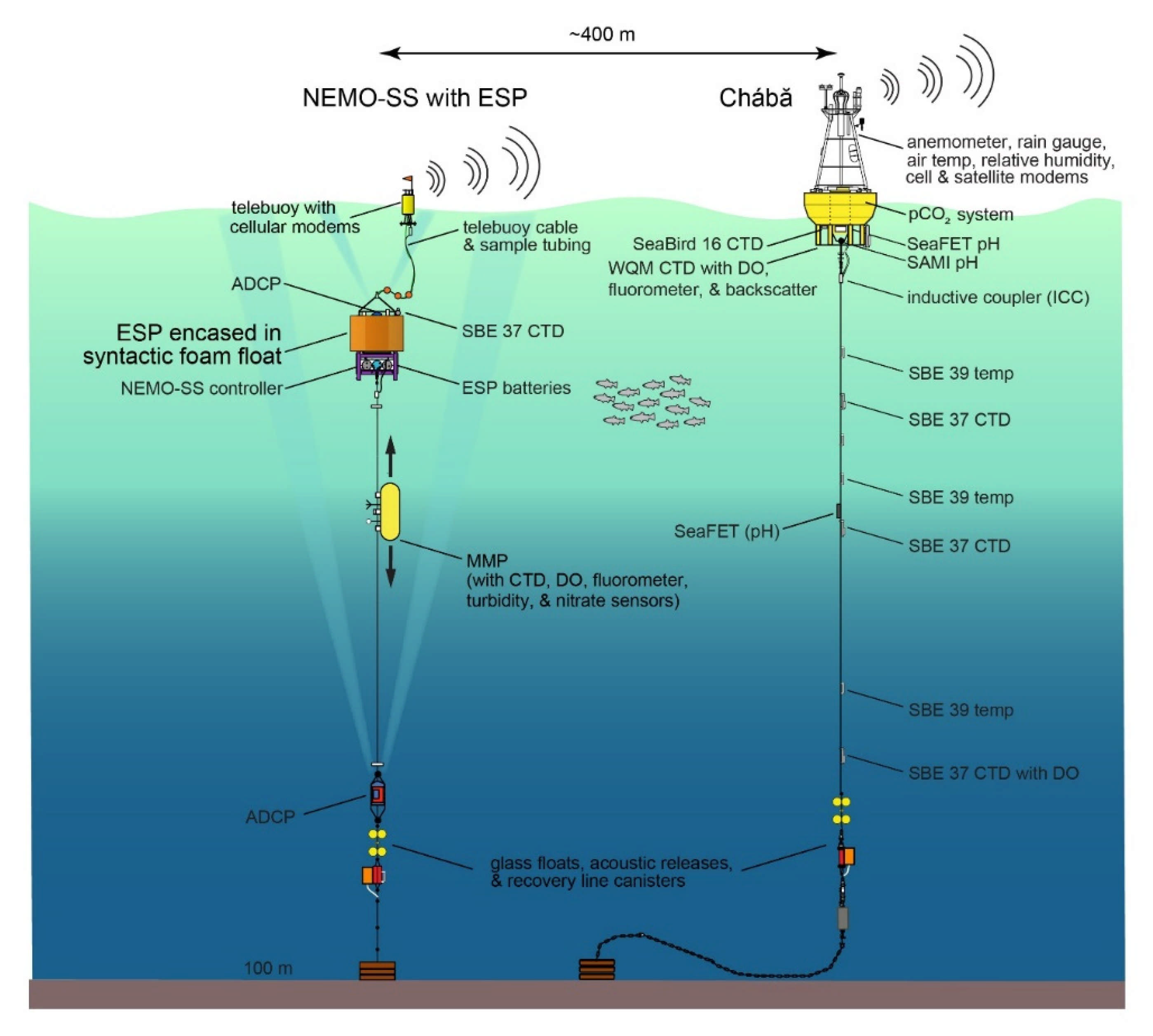
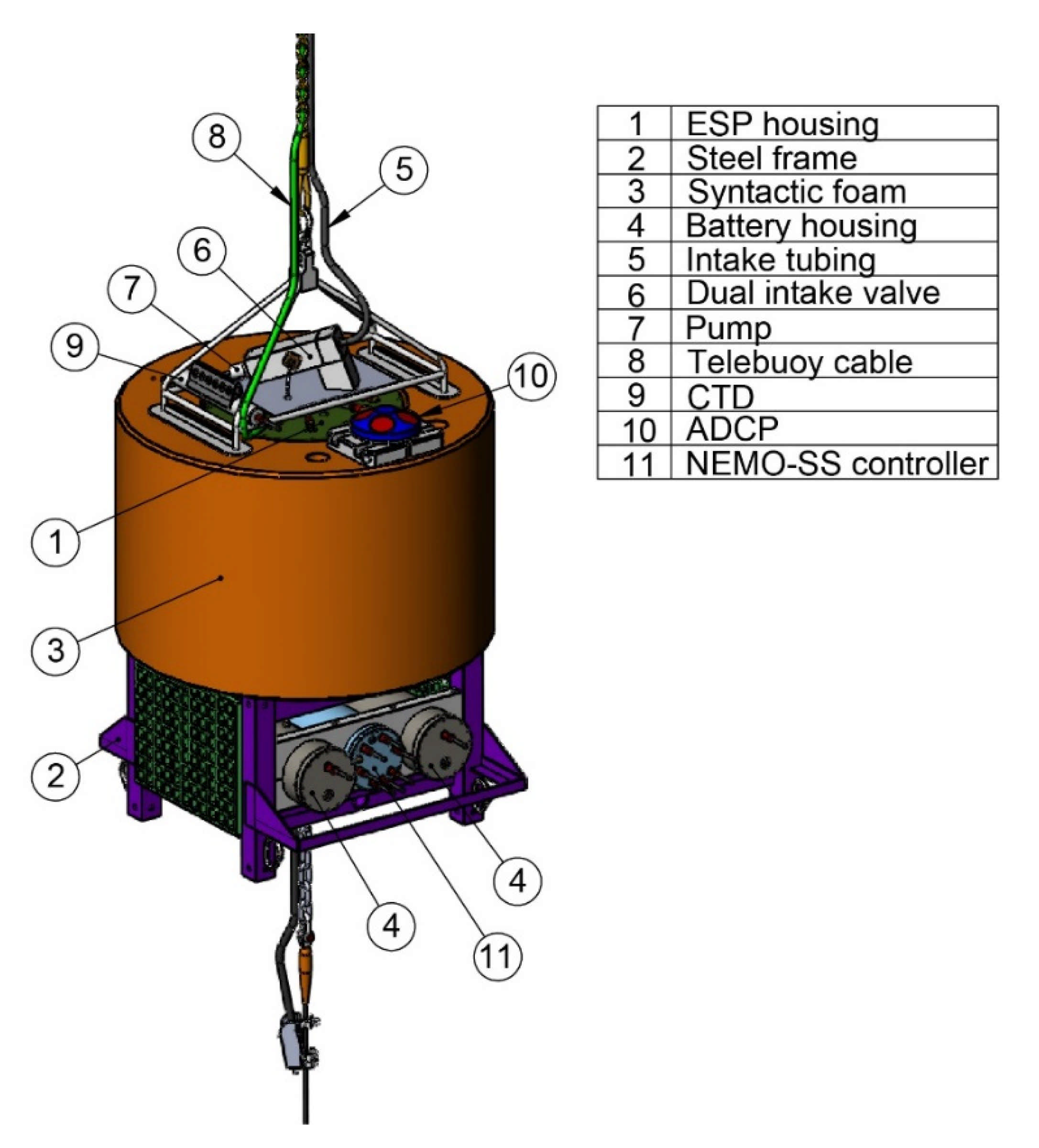
2.2. ESP Mooring Design
- Minimizing ESP tilts and accelerations (particularly on recovery) so that they do not exceed 30 degrees from vertical and 2g (where g is the acceleration of gravity and is equal to 9.8 m/s2) to prevent damage to ESP components and spills/leaks of ESP reagents/waste;
- providing at least 5000 watt-hours of power for the ESP and its telemetry and pump systems for the ~60-day deployments;
- supporting constant, relatively high-bandwidth, two-way communications with a shore-side server;
- sampling near-surface water (1–2 m), where the highest concentrations of Pseudo-nitzschia cells were expected, without drawing air into the pumped sampling system;
- facilitating straightforward and safe deployments and recoveries;
- minimizing drag to decrease mooring knock-down in expected currents of up to 0.75 m/s;
- locating the ESP below the strong accelerations of surface wave motions, but shallower than the pressure case rating of 50-m, and ideally at pressures of ~16–18 dbars to facilitate sample collection; and
- attaining a minimum buoyancy of 226 kg to allow sufficient wire tension for a McLane Moored Profiler (MMP) to operate.
2.2.1. ESP Power Subsystem
2.2.2. ESP Telemetry Subsystem
2.2.3. ESP Pump Subsystem
2.3. ESP Configuration and Calibration
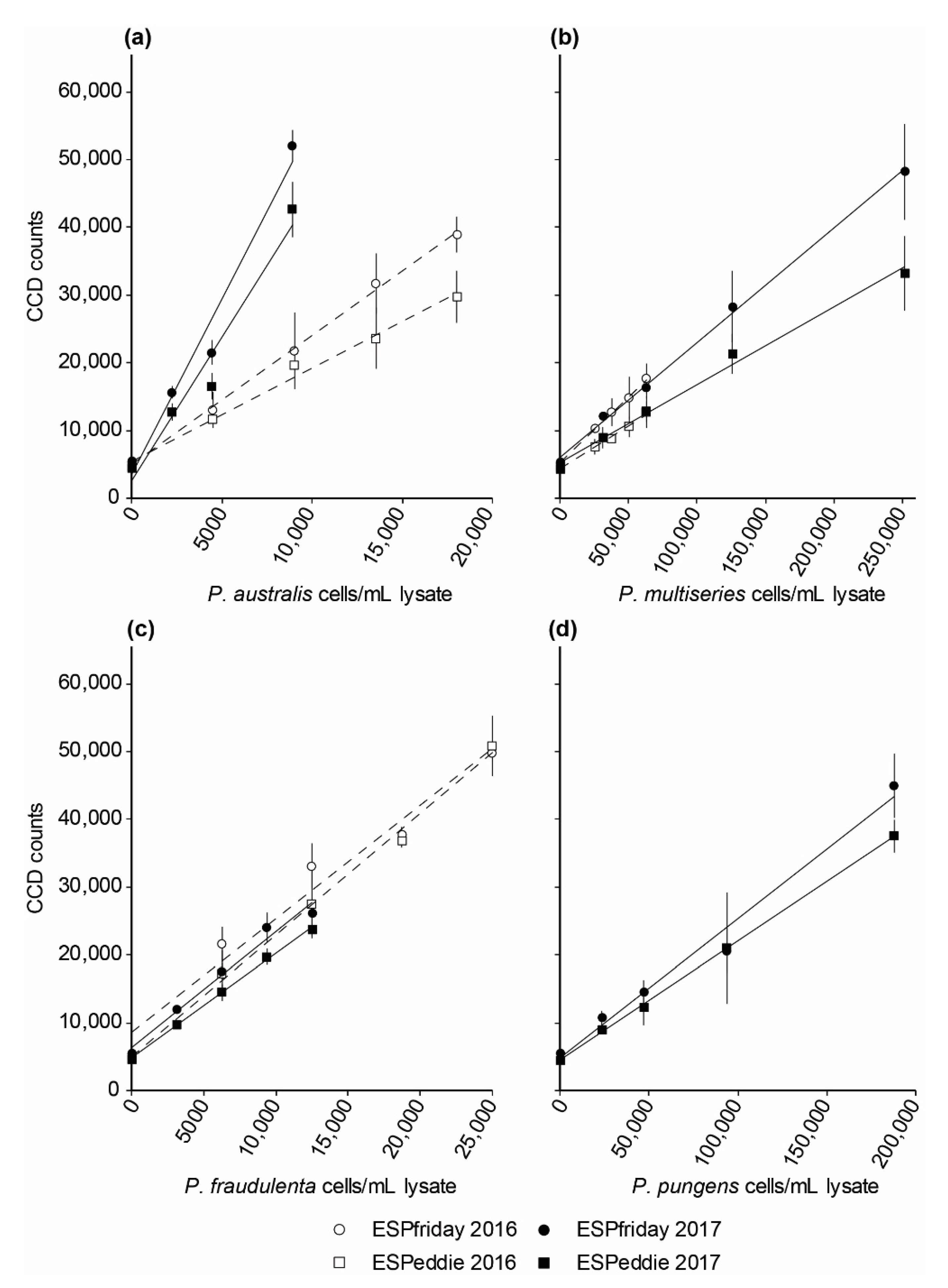
2.3.1. Harmful Algal Bloom Species Calibration
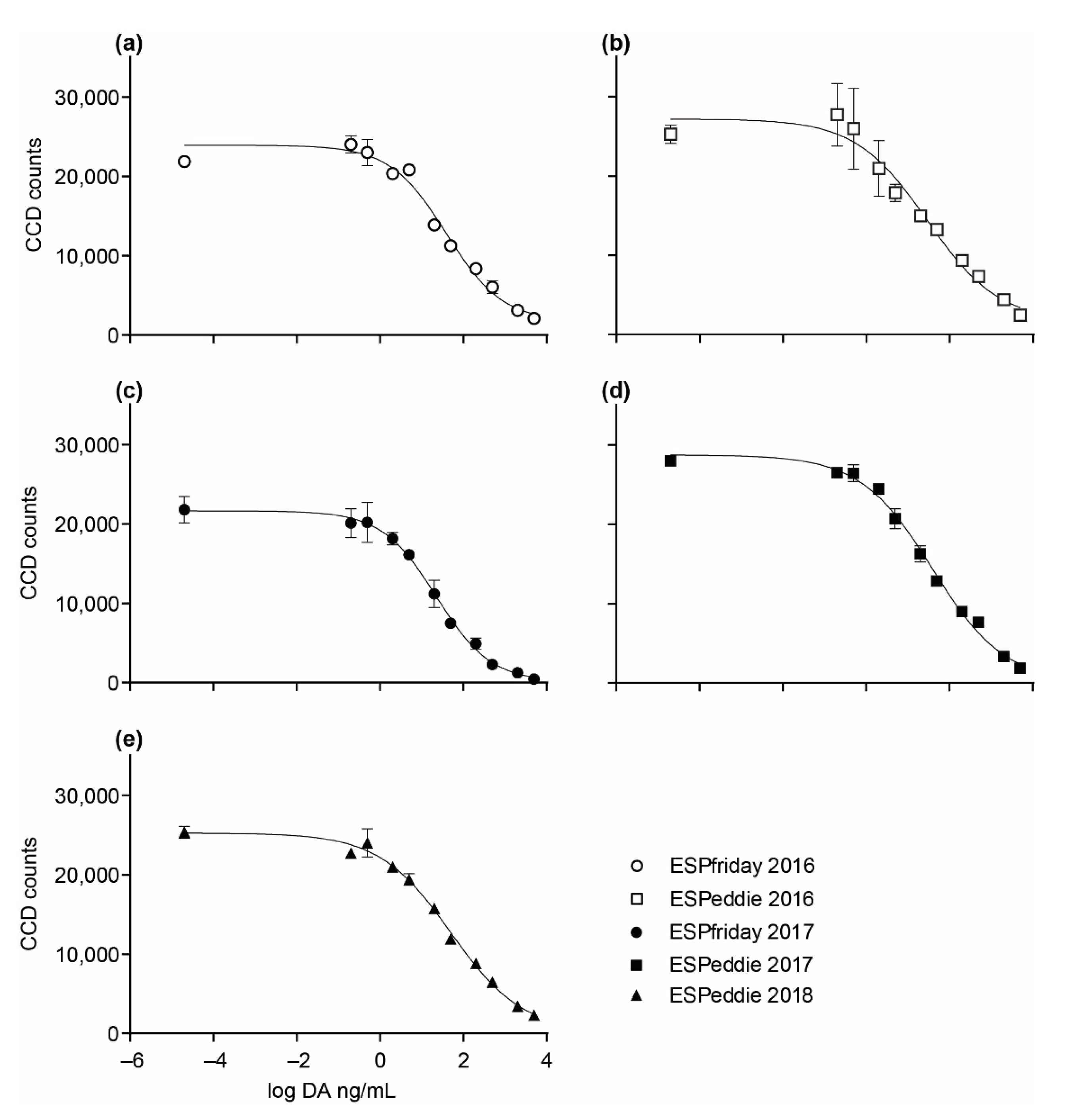
2.3.2. Domoic Acid Calibration
2.4. ESP Deployments
3. Results
3.1. Overview of ESP Observations and Mooring Performance
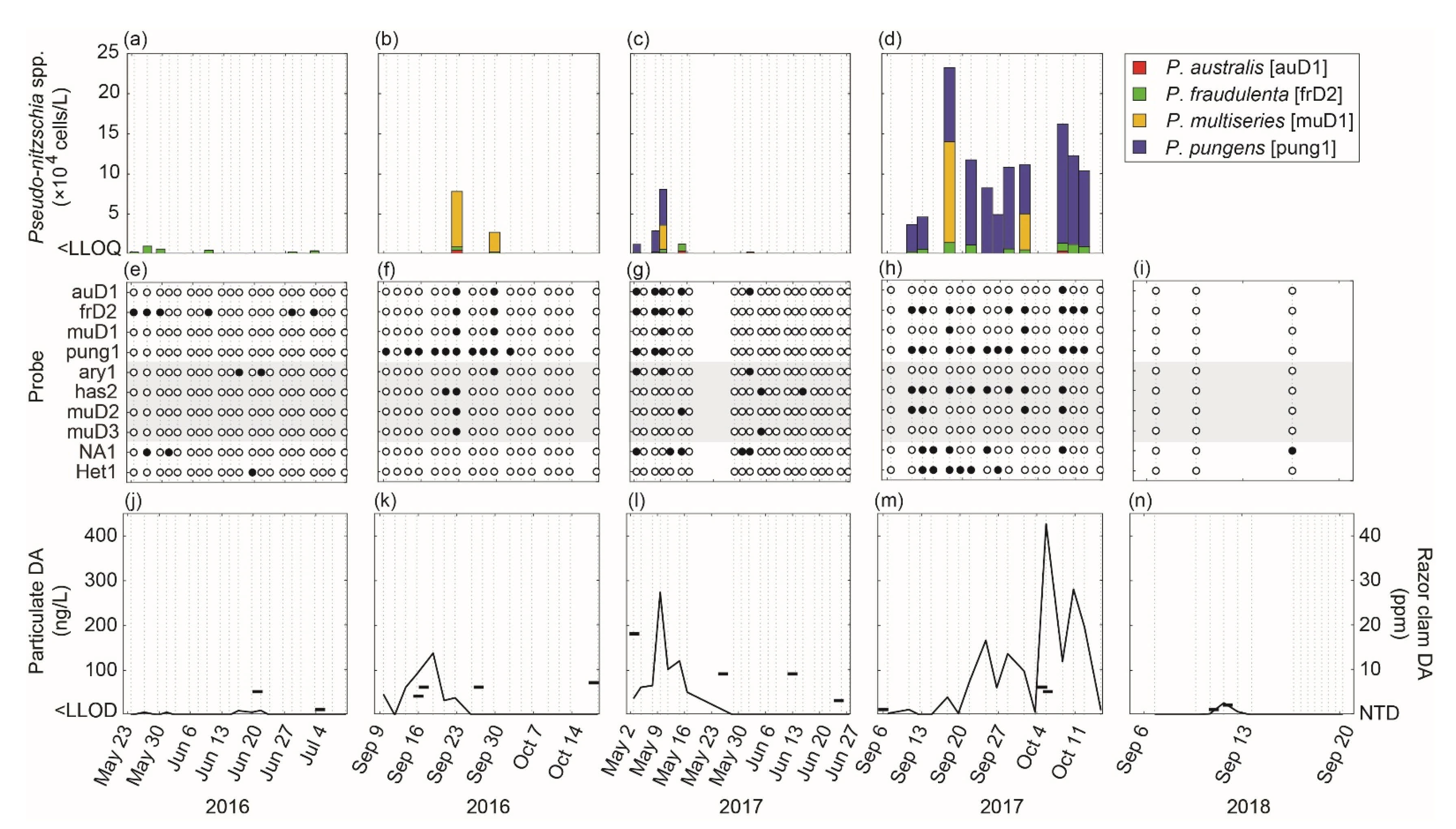
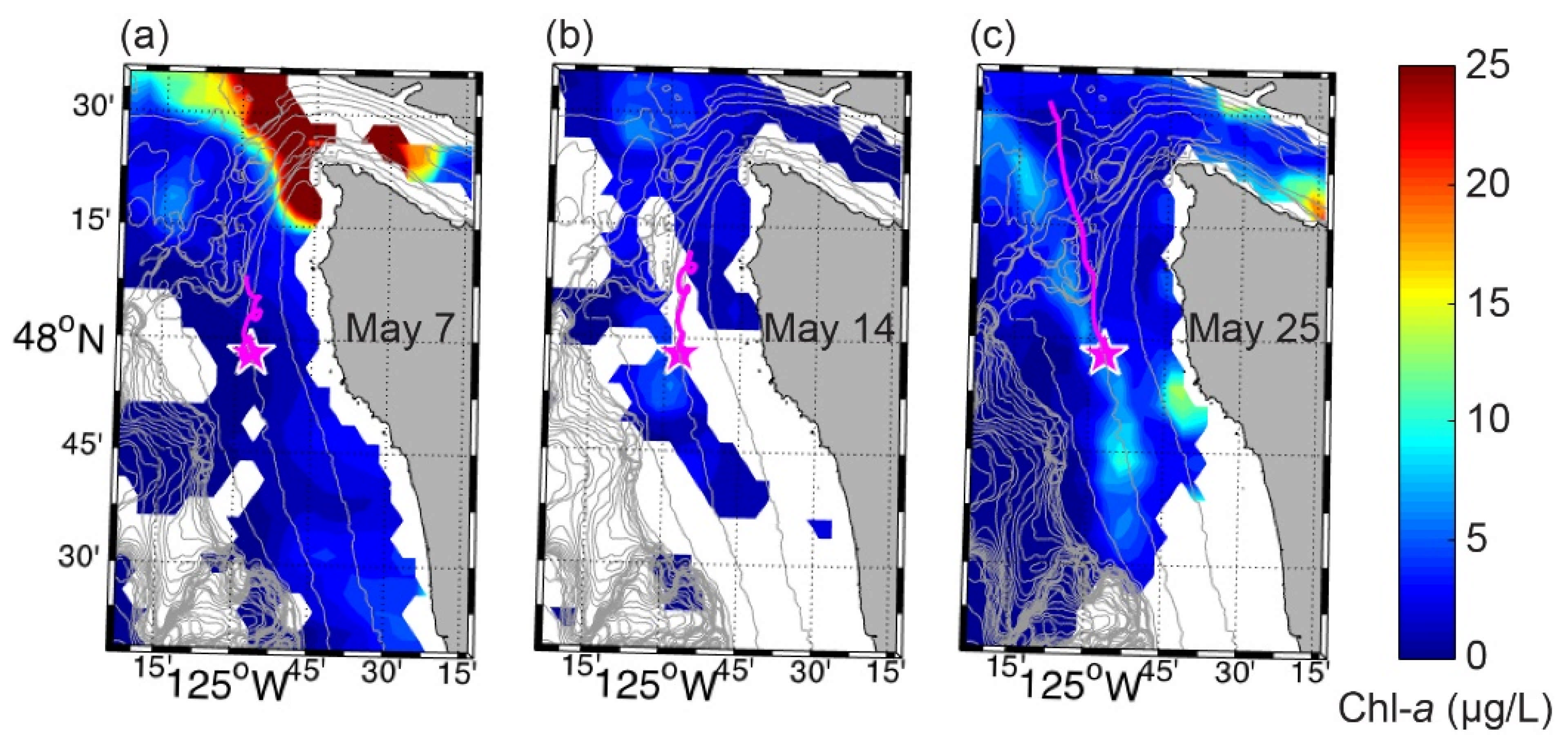
3.2. Spring 2017 Synthesis
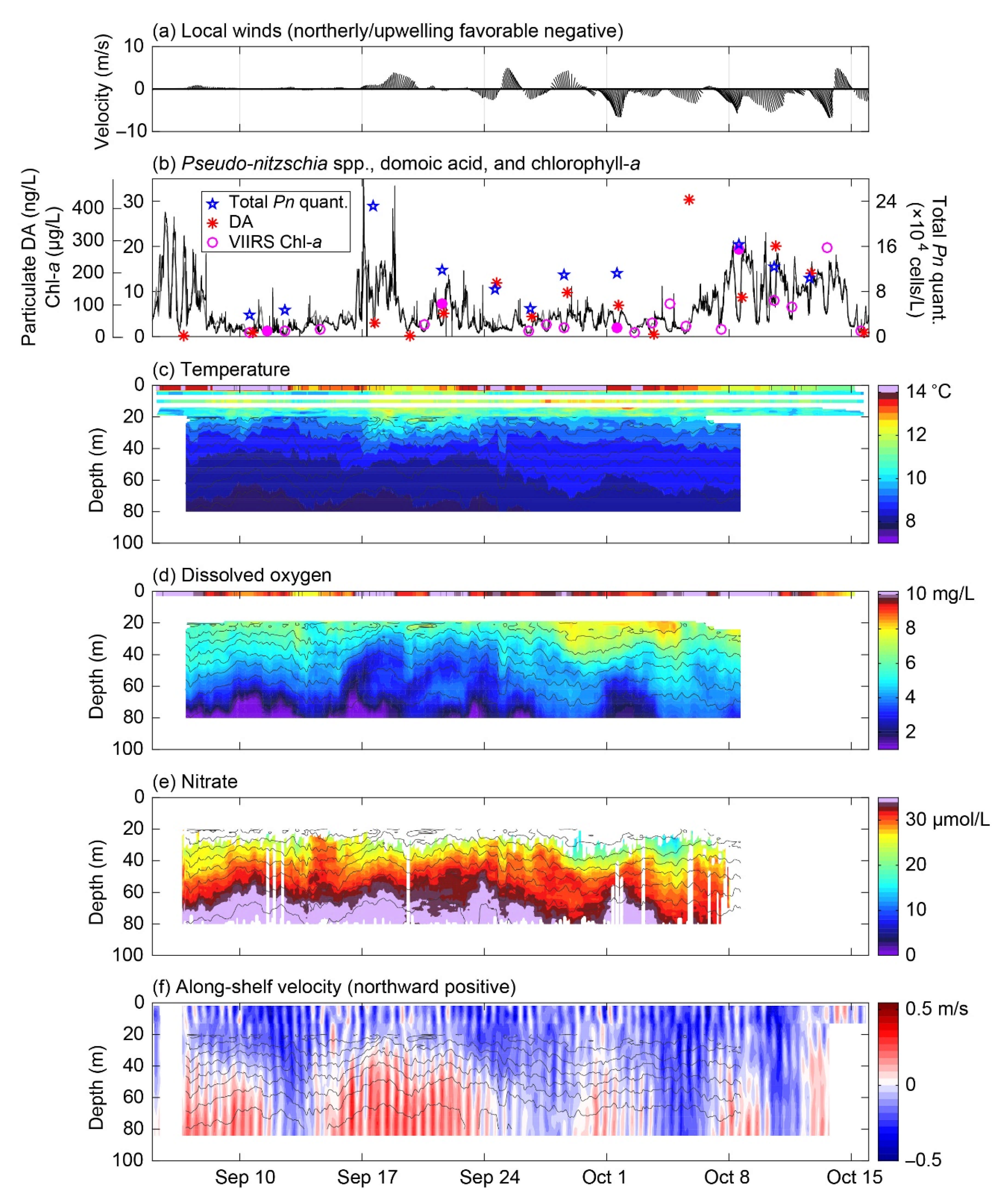
3.3. Fall 2017 Synthesis
4. Discussion
4.1. Episodic Blooms of Pseudo-nitzschia spp. Revealed by ESP
4.2. ESP Mooring Performance and Improvements
4.3. Future Directions for Identifying HAB Response to Climate Change Using ESP
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Dedication
References
- Wells, M.L.; Trainer, V.L.; Smayda, T.J.; Karlson, B.S.O.; Trick, C.G.; Kudela, R.M.; Ishikawa, A.; Bernard, S.; Wulff, A.; Anderson, D.M.; et al. Harmful algal blooms and climate change: Learning from the past and present to forecast the future. Harmful Algae 2015, 49, 68–93. [Google Scholar] [CrossRef]
- Moore, S.K.; Trainer, V.L.; Mantua, N.J.; Parker, M.S.; Laws, E.A.; Fleming, L.E.; Backer, L.C. Impacts of climate variability and future climate change on harmful algal blooms and human health. Environ. Health 2008, 7 (Suppl. 2). [Google Scholar] [CrossRef]
- Hallegraeff, G.M. Ocean climate change, phytoplankton community responses, and harmful algal blooms: A formidable predictive challenge. J. Phycol. 2010, 46, 220–235. [Google Scholar] [CrossRef]
- Gobler, C.J.; Doherty, O.M.; Hattenrath-Lehmann, T.K.; Griffith, A.W.; Kang, Y.; Litaker, R.W. Ocean warming since 1982 has expanded the niche of toxic algal blooms in the North Atlantic and North Pacific oceans. Proc. Natl. Acad. Sci. USA 2017, 114, 4975–4980. [Google Scholar] [CrossRef]
- McCabe, R.M.; Hickey, B.M.; Kudela, R.M.; Lefebvre, K.A.; Adams, N.G.; Bill, B.D.; Gulland, F.M.D.; Thomson, R.E.; Cochlan, W.P.; Trainer, V.L. An unprecedented coastwide toxic algal bloom linked to anomalous ocean conditions. Geophys. Res. Lett. 2016, 43, 10366–10376. [Google Scholar] [CrossRef] [PubMed]
- Trainer, V.L.; Kudela, R.M.; Hunter, M.V.; Adams, N.G.; McCabe, R.M. Climate Extreme Seeds a New Domoic Acid Hotspot on the US West Coast. Front. Clim. 2020, 2, 571836. [Google Scholar] [CrossRef]
- Wells, M.L.; Karlson, B.; Wulff, A.; Kudela, R.; Trick, C.; Asnaghi, V.; Berdalet, E.; Cochlan, W.; Davidson, K.; Rijcke, M.D.; et al. Future HAB science: Directions and challenges in a changing climate. Harmful Algae 2020, 91, 101632. [Google Scholar] [CrossRef] [PubMed]
- Brosnahan, M.L.; Velo-Suárez, L.; Ralston, D.K.; Fox, S.E.; Sehein, T.R.; Shalapyonok, A.; Sosik, H.M.; Olson, R.J.; Anderson, D.M. Rapid growth and concerted sexual transitions by a bloom of the harmful dinoflagellate Alexandrium fundyense (Dinophyceae). Limnol. Oceanogr. 2015, 60, 2059–2078. [Google Scholar] [CrossRef] [PubMed]
- Fu, F.-X.; Tatters, A.O.; Hutchins, D.A. Global change and the future of algal blooms in the ocean. Mar. Ecol. Prog. Ser. 2012, 470, 207–233. [Google Scholar] [CrossRef]
- Trainer, V.L.; Moore, S.K.; Hallegraeff, G.; Kudela, R.M.; Clement, A.; Mardones, J.I.; Cochlan, W.P. Pelagic harmful algal blooms and climate change: Lessons from nature’s experiments with extremes. Harmful Algae 2020, 91, 101591. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.; Connell, P.; Evans, R.H.; Gellene, A.G.; Howard, M.D.A.; Jones, B.H.; Kaveggia, S.; Palmer, L.; Schnetzer, A.; Seegers, B.N.; et al. A decade and a half of Pseudo-nitzschia spp. and domoic acid along the coast of southern California. Harmful Algae 2018, 79, 87–104. [Google Scholar] [CrossRef]
- Irwin, A.J.; Finkel, Z.V.; Müller-Karger, F.E.; Ghinaglia, L.T. Phytoplankton adapt to changing ocean environments. Proc. Natl. Acad. Sci. USA 2015, 112, 5762–5766. [Google Scholar] [CrossRef]
- Ryan, J.P.; Kudela, R.M.; Birch, J.M.; Blum, M.; Bowers, H.A.; Chavez, F.P.; Doucette, G.J.; Hayashi, K.; Marin, R., III; Mikulski, C.M.; et al. Causality of an extreme harmful algal bloom in Monterey Bay, California, during the 2014–2016 northeast Pacific warm anomaly. Geophys. Res. Lett. 2017, 44, 5571–5579. [Google Scholar] [CrossRef]
- Anderson, D.M.; Cembella, A.D.; Hallegraeff, G.M. Progress in understanding harmful algal blooms: Paradigm shifts and new technologies for research, monitoring, and management. Annu. Rev. Mar. Sci. 2012, 4, 143–176. [Google Scholar] [CrossRef] [PubMed]
- Scholin, C.A.; Doucette, G.; Jensen, S.; Roman, B.; Pargett, D.; Marin, R., III; Preston, C.M.; Jones, W.; Feldman, J.; Everlove, C.; et al. Remote detection of marine microbes, small invertebrates, harmful algae, and biotoxins using the Environmental Sample Processor (ESP). Oceanography 2009, 22, 158–167. [Google Scholar] [CrossRef]
- Bowers, H.A.; Ryan, J.P.; Hayashi, K.; Woods, A.L.; Marin, R., III; Smith, G.J.; Hubbard, K.A.; Doucette, G.J.; Mikulski, C.M.; Gellene, A.G.; et al. Diversity and toxicity of Pseudo-nitzschia species in Monterey Bay: Perspectives from targeted and adaptive sampling. Harmful Algae 2018, 78, 129–141. [Google Scholar] [CrossRef]
- Anderson, D.M.; Keafer, B.A.; McGillicuddy, D.J., Jr.; Solow, A.R.; Kleindhinst, J.L. Improving the accuracy and utility of harmful algal bloom forecasting systems. In Biological and Geological Perspectives of Dinoflagellates; Lewis, J.M., Marrett, F., Bradley, L.R., Eds.; Geological Society for the Micropalaeontological Society: Bath, UK, 2013; pp. 141–148. [Google Scholar]
- Ryan, J.; Greenfield, D.; Marin, R., III; Preston, C.M.; Roman, B.; Jensen, S.; Pargett, D.; Birch, J.; Mikulski, C.; Doucette, G.; et al. Harmful phytoplankton ecology studies using an autonomous molecular analytical and ocean observing network. Limnol. Oceanogr. 2011, 56, 1255–1272. [Google Scholar] [CrossRef]
- Horner, R.A.; Garrison, D.L.; Plumley, F.G. Harmful algal blooms and red tide problems on the U.S. west coast. Limnol. Oceanogr. 1997, 42, 1076–1088. [Google Scholar] [CrossRef]
- Davis, K.A.; Banas, N.S.; Giddings, S.N.; Siedlecki, S.A.; MacCready, P.; Lessard, E.J.; Kudela, R.M.; Hickey, B.M. Estuary-enhanced upwelling of marine nutrients fuels coastal productivity in the U.S. Pacific Northwest. J. Geophys. Res. 2014, 2014, 8778–8799. [Google Scholar] [CrossRef]
- Lewitus, A.J.; Horner, R.A.; Caron, D.A.; Garcia-Mendoza, E.; Hickey, B.M.; Hunter, M.; Huppert, D.D.; Kudela, R.M.; Langlois, G.W.; Largier, J.L.; et al. Harmful algal blooms along the North American west coast region: History, trends, causes, and impacts. Harmful Algae 2012, 19, 133–159. [Google Scholar] [CrossRef]
- FDA. Fish and Fishery Products Hazards and Controls Guidance; Department of Health and Human Services, Public Health Service, Food and Drug Administration, Center for Food Safety and Applied Nutrition, Office of Food Safety: Silver Spring, MD, USA, 2011. [Google Scholar]
- Trainer, V.L.; Suddleson, M. Monitoring approaches for early warning of domoic acid events in Washington State. Oceanography 2005, 18, 228–237. [Google Scholar] [CrossRef]
- Baugh, K.A.; Bush, J.M.; Bill, B.D.; Lefebvre, K.A.; Trainer, V.L. Estimates of specific toxicity in several Pseudo-nitzschia species from the Washington coast, based on culture and field studies. Afr. J. Mar. Sci. 2006, 28, 403–407. [Google Scholar] [CrossRef]
- WDOH. Shellfish Poisoning: Paralytic, Domoic Acid, or Diarrhetic. 2016. Available online: https://www.doh.wa.gov/Portals/1/Documents/5100/420-077-Guideline-ShellfishPoisoning.pdf (accessed on 17 March 2021).
- Ritzman, J.; Brodbeck, A.; Brostrom, S.; McGrew, S.; Dreyer, S.; Klinger, T.; Moore, S.K. Economic and sociocultural impacts of fisheries closures in two fishing-dependent communities following the massive 2015 U.S. West Coast harmful algal bloom. Harmful Algae 2018, 80, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.K.; Dreyer, S.J.; Ekstrom, J.A.; Moore, K.; Norman, K.; Klinger, T.; Allison, E.H.; Jardine, S.L. Harmful algal blooms and coastal communities: Socioeconomic impacts and actions taken to cope with the 2015 U.S. West Coast domoic acid event. Harmful Algae 2020, 96, 101799. [Google Scholar] [CrossRef] [PubMed]
- Trainer, V.L.; Hickey, B.M.; Lessard, E.J.; Cochlan, W.P.; Trick, C.G.; Wells, M.L.; MacFadyen, A.; Moore, S.K. Variability of Pseudo-nitzschia and domoic acid in the Juan de Fuca eddy region and its adjacent shelves. Limnol. Oceanogr. 2009, 54, 289–308. [Google Scholar] [CrossRef]
- Hickey, B.M.; Banas, N. Oceanography of the Pacific Northwest coastal ocean and estuaries with application to coastal ecosystems. Estuaries 2003, 26, 1010–1031. [Google Scholar] [CrossRef]
- Hickey, B.M.; Trainer, V.L.; Kosro, P.M.; Adams, N.G.; Connolly, T.P.; Kachel, N.B.; Geier, S.L. A springtime source of toxic Pseudo-nitzschia cells on razor clam beaches in the Pacific Northwest. Harmful Algae 2013, 25, 1–14. [Google Scholar] [CrossRef]
- MacFadyen, A.; Hickey, B.M.; Foreman, M.G.G. Transport of surface waters from the Juan de Fuca eddy region to the Washington coast. Cont. Shelf Res. 2005, 25, 2008–2021. [Google Scholar] [CrossRef]
- MacFadyen, A.; Hickey, B.M. Generation and evolution of a topographically linked, mesoscale eddy under steady and variable wind-forcing. Cont. Shelf Res. 2010, 30, 1387–1402. [Google Scholar] [CrossRef]
- Olson, M.B.; Lessard, E.J.; Cochlan, W.P.; Trainer, V.L. Intrinsic growth and phytoplankton grazing on toxigenic Pseudo-nitzschia spp. diatoms from the coastal Northeast Pacific. Limnol. Oceanogr. 2008, 53, 1352–1368. [Google Scholar] [CrossRef]
- MacFadyen, A.; Hickey, B.M.; Cochlan, W.P. Influences of the Juan de Fuca Eddy on circulation, nutrients, and phytoplankton production in the northern California Current System. J. Geophys. Res. 2008, 113, C08008. [Google Scholar] [CrossRef]
- Greenfield, D.I.; Marin, R., III; Jensen, S.; Massion, E.; Roman, B.; Feldman, J.; Scholin, C.A. Application of environmental sample processor (ESP) methodology for quantifying Pseudo-nitzschia australis using ribosomal RNA-targeted probes in sandwich and fluorescent in situ hybridization formats. Limnol. Oceanogr. Methods 2006, 4, 426–435. [Google Scholar] [CrossRef]
- Greenfield, D.I.; Marin, R., III; Doucette, G.J.; Mikulski, C.; Jones, K.; Jensen, S.; Roman, B.; Alvarado, N.; Feldman, J.; Scholin, C. Field applications of the second-generation Environmental Sample Processor (ESP) for remote detection of harmful algae: 2006–2007. Limnol. Oceanogr. Methods 2008, 6, 667–679. [Google Scholar] [CrossRef]
- Doucette, G.J.; Mikulski, C.M.; Jones, K.L.; King, K.L.; Greenfield, D.I.; Marin, R., III; Jensen, S.; Roman, B.; Elliott, C.T.; Scholin, C.A. Remote, sub-surface detection of the algal toxin domoic acid onboard the Environmental Sample Processor: Assay development and field trials. Harmful Algae 2009, 8, 880–888. [Google Scholar] [CrossRef]
- Scholin, C.A.; Marin, R., III; Miller, P.E.; Doucette, G.J.; Powell, C.L.; Haydock, P.; Howard, J.; Ray, J. DNA probes and a receptor-binding assay for detection of Pseudo-nitzschia (Bacillariophyceae) species and domoic acid activity in cultured and natural samples. J. Phycol. 1999, 35, 1356–1367. [Google Scholar] [CrossRef]
- Bowers, H.A.; Marin, R., III; Birch, J.M.; Scholin, C.A. Sandwich hybridization probes for the detection of Pseudo-nitzschia (Bacillariophyceae) species: An update to existing probes and a description of new probes. Harmful Algae 2017, 70, 37–51. [Google Scholar] [CrossRef]
- Tyrrell, J.V.; Scholin, C.A.; Berguist, P.R.; Berguist, P.L. Detection and enumeration of Heterosigma akashiwo and Fibrocapsa japonica (Raphidophyceae) using rRNA-targeted oligonucleotide probes. Phycologia 2001, 40, 457–467. [Google Scholar] [CrossRef]
- Scholin, C.A.; Doucette, G.J.; Cembella, A.D. Prospects for Developing Automated Systems for In Situ Detection of Harmful Algae and Their Toxins; UNESCO Publishing: Paris, France, 2008; pp. 413–462. [Google Scholar]
- Guillard, R.R.L.; Ryther, J.H. Studies of marine planktonic diatoms: I. Cyclotella nana Hustedt, and Detonula confervacea (Cleve). Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, J.E.; Maritorena, S.; Mitchell, B.G.; Siegel, D.A.; Carder, K.L.; Garver, S.A.; Kahru, M.; McClain, C.R. Ocean color chlorophyll algorithms for SeaWiFS. J. Geophys. Res. 1998, 103, 24937–24953. [Google Scholar] [CrossRef]
- Battisti, D.S.; Hickey, B.M. Application of Remote Wind-Forced Coastal Trapped Wave Theory to the Oregon and Washington Coasts. J. Phys. Oceanogr. 1984, 14, 887–903. [Google Scholar] [CrossRef]
- Wang, D.-P.; Mooers, C.N.K. Long Coastal-Trapped Waves off the West Coast of the United States, Summer 1973. J. Phys. Oceanogr. 1977, 7, 856–864. [Google Scholar] [CrossRef][Green Version]
- Alford, M.H.; Mickett, J.B.; Zhang, S.; MacCready, P.; Zhao, Z.; Newton, J.A. Internal waves on the Washington continental shelf. Oceanography 2012, 25, 66–79. [Google Scholar] [CrossRef]
- Omand, M.M.; Feddersen, F.; Guza, R.T.; Franks, P.J.S. Episodic vertical nutrient fluxes and nearshore phytoplankton blooms in Southern California. Limnol. Oceanogr. 2012, 57, 1673–1688. [Google Scholar] [CrossRef]
- Villamaña, M.; Mouriño-Carballido, B.; Marañón, E.; Cermeño, P.; Chouciño, P.; Silva, J.C.B.d.; Díaz, P.A.; Fernández-Castro, B.; Gilcoto, M.; Graña, R.; et al. Role of internal waves on mixing, nutrient supply and phytoplankton community structure during spring and neap tides in the upwelling ecosystem of Ría de Vigo (NW Iberian Peninsula). Limnol. Oceanogr. 2017, 62, 1014–1030. [Google Scholar] [CrossRef]
- Alonso-Rodríguez, R.; Ochoa, J.L. Hydrology of winter–spring “red tides” in Bahía de Mazatlán, Sinaloa, México. Harmful Algae 2004, 3, 163–171. [Google Scholar] [CrossRef]
- Herfort, L.; Seaton, C.; Wilkin, M.; Roman, B.; Preston, C.M.; Marin, R.I.; Seitz, K.; Smith, M.W.; Haynes, V.; Scholin, C.A.; et al. Use of continuous, real-time observations and model simulations to achieve autonomous, adaptive sampling of microbial processes with a robotic sampler. Limnol. Oceanogr. Methods 2016, 14, 50–67. [Google Scholar] [CrossRef]
- Ryan, J.P.; McManus, M.A.; Kudela, R.M.; Artigas, M.L.; Bellingham, J.G.; Chavez, F.P.; Doucette, G.; Foley, D.; Godin, M.; Harvey, J.B.J.; et al. Boundary influences on HAB phytoplankton ecology in a stratification-enhanced upwelling shadow. Deep-Sea Res. II 2014, 101, 63–79. [Google Scholar] [CrossRef]
- Brosnahan, M.L.; Doucette, G.J.; Smith, J.L.; Keafer, B.A.; Mikulski, C.M.; Sanderson, M.P.; Anderson, D.M. Dynamics of PSP toxin production by an inshore bloom of A. catenella observed through co-deployment of complementary, automated in situ phytoplankton sensors. In Proceedings of the 9th United States Symposium on Harmful Algae, Baltimore, MD, USA, 11–17 November 2017. [Google Scholar]
- Saito, M.A.; Bulygin, V.V.; Moran, D.M.; Taylor, C.; Scholin, C. Examination of microbial proteome preservation techniques applicable to autonomous environmental sample collection. Front. Microbiol. 2011, 2, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Scholin, C.A. Ecogenomic Sensors. In Encyclopedia of Biodiversity, 2nd ed.; Levin, S.A., Ed.; Academic Press: Waltham, MA, USA, 2013; Volume 2, pp. 690–700. [Google Scholar]
- Ottesen, E.A.; Young, C.R.; Gifford, S.M.; Eppley, J.M.; Marin, R., III; Schuster, S.C.; Scholin, C.A.; DeLong, E.F. Multispecies diel transcriptional oscillations in open ocean heterotrophic bacterial assemblages. Science 2014, 345, 207–212. [Google Scholar] [CrossRef]
- Bowers, H.A.; Marin, R.I.; Birch, J.M.; Scholin, C.A.; Doucette, G.J. Recovery and identification of Pseudo-nitzschia (Bacillariophyceae) frustules from natural samples acquired using the environmental sample processor. J. Phycol. 2016, 52, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Preston, C.M.; Harris, A.; Ryan, J.P.; Roman, B.; Marin, R., III; Jensen, S.; Everlove, C.; Birch, J.; Dzenitis, J.M.; Pargett, D.; et al. Underwater application of quantitative PCR on an ocean mooring. PLoS ONE 2011, 6, e22522. [Google Scholar] [CrossRef] [PubMed]
- Robidart, J.C.; Church, M.J.; Ryan, J.P.; Ascani, F.; Wilson, S.T.; Bombar, D.; Marin, R.I.; Richards, K.J.; Karl, D.M.; Scholin, C.A.; et al. Ecogenomic sensor reveals controls on N2-fixing microorganisms in the North Pacific Ocean. ISME J. 2014, 8, 1175–1185. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Bellingham, J.G.; Ryan, J.P.; Kieft, B.; Stanway, J. Autonomous four-dimensional mapping and tracking of a coastal upwelling front by an autonomous underwater vehicle. J. Field Robot. 2016, 33, 67–81. [Google Scholar] [CrossRef]
- Zhang, Y.; Ryan, J.P.; Kieft, B.; Hobson, B.W.; McEwen, R.S.; Godin, M.A.; Harvey, J.B.; Barone, B.; Bellingham, J.G.; Birch, J.M.; et al. Targeted sampling by autonomous underwater vehicles. Front. Mar. Sci. 2019, 6, 415. [Google Scholar] [CrossRef]
- Zhang, Y.; Kieft, B.; Hobson, B.; Ryan, J.; Barone, B.; Preston, C.; Roman, B.; Raanan, B.-Y.; Marin, R.I.; O’Reilly, T.; et al. Autonomous tracking and sampling of the deep chlorophyll maximum layer in an open-ocean eddy by a long range autonomous underwater vehicle. IEEE J. Ocean. Eng. 2019, 45, 1308–1321. [Google Scholar] [CrossRef]
- Ussler, W.; Preston, C.; Lingerfelt, L.; Mikulski, C.; Den Uyl, P.; Johengen, T.; Vander Woude, A.; Errera, R.; Ruberg, S.; Goodwin, K.; et al. The 3rd Generation ESP/Long-Range AUV: First tests of autonomous, underway sampling and analysis of microcystin in western Lake Erie. In Proceedings of the 10th US Symposium on Harmful Algae, Orange Beach, AL, USA, 3–8 November 2019. [Google Scholar]
- McKibben, M.; Peterson, W.; Wood, A.M.; Trainer, V.L.; Hunter, M.; White, A.E. Climatic regulation of the neurotoxin domoic acid. Proc. Natl. Acad. Sci. USA 2017, 114, 239–244. [Google Scholar] [CrossRef]
- Anderson, C.R.; Berdalet, E.; Kudela, R.M.; Cusack, C.; Silke, J.; O’Rourke, E.; Dugan, D.; McCammon, M.; Newton, J.A.; Moore, S.K.; et al. Scaling Up from Regional Case Studies to a Global Harmful Algal Bloom Observing System. Front. Mar. Sci. 2019, 6. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | ESP | Probe | α | β | R2 | LLOQ (Cells/mL) |
|---|---|---|---|---|---|---|
| 2016 | ESPfriday | auD1 | 5043.6 | 1.8999 | 0.9976 | 434 |
| muD1 | 5362.3 | 0.1932 | 0.9986 | 2622 | ||
| frD2 | 8543.2 | 1.6806 | 0.9693 | 323 | ||
| ESPeddie | auD1 | 5386.4 | 1.3797 | 0.9888 | 152 | |
| muD1 | 4408.0 | 0.1263 | 0.9903 | 9409 | ||
| frD2 | 5021.9 | 1.7922 | 0.9953 | 627 | ||
| 2017 | ESPfriday | auD1 | 3440.0 | 5.1888 | 0.9706 | 484 |
| muD1 | 6090.2 | 0.1693 | 0.9979 | 4099 | ||
| frD2 | 6380.1 | 1.7112 | 0.9784 | 405 | ||
| pung1 | 4843.9 | 0.2051 | 0.9828 | 5396 | ||
| ESPeddie | auD1 | 2555.6 | 4.2469 | 0.9587 | 697 | |
| muD1 | 5347.1 | 0.1144 | 0.9911 | 1471 | ||
| frD2 | 4812.5 | 1.5423 | 0.9980 | 456 | ||
| pung1 | 4466.7 | 0.1758 | 0.9996 | 5964 |
| Year | ESP | Slope | R2 | EC50 | EC80 | EC20 | LLOD (ng/mL) |
|---|---|---|---|---|---|---|---|
| 2016 | ESPfriday | −0.642 | 0.978 | 38.93 | 4.48 | 338.03 | 1.81 |
| ESPeddie | −0.504 | 0.942 | 29.02 | 1.85 | 455.21 | 1.48 | |
| 2017 | ESPfriday | −0.663 | 0.955 | 22.42 | 2.77 | 181.66 | 1.95 |
| ESPeddie | −0.501 | 0.989 | 38.97 | 2.45 | 619.70 | 1.32 | |
| 2018 | ESPeddie | −0.502 | 0.988 | 50.23 | 3.17 | 794.85 | 1.40 |
| Year | Season | ESP | Active Deployment Period | Target Analyte | Total # Samples | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| auD1 | muD1 | frD2 | pung1 | NA−1 | Het1 | pDA | |||||
| 2016 | Spring | ESPfriday | 23 May–10 July | Q | Q | Q | +/− | +/− | +/− | Q | 42 |
| Fall | ESPeddie | 9 September–18 October | Q | Q | Q | +/− | +/− | +/− | Q | 34 | |
| 2017 | Spring | ESPfriday | 2 May–27 June | Q | Q | Q | Q | +/− | +/− | Q | 42 |
| Fall | ESPeddie | 6 September–15 October | Q | Q | Q | Q | +/− | +/− | Q | 34 | |
| 2018 | Fall | ESPeddie | 6–20 6 September | +/− | +/− | +/− | +/− | +/− | +/− | Q | 17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moore, S.K.; Mickett, J.B.; Doucette, G.J.; Adams, N.G.; Mikulski, C.M.; Birch, J.M.; Roman, B.; Michel-Hart, N.; Newton, J.A. An Autonomous Platform for Near Real-Time Surveillance of Harmful Algae and Their Toxins in Dynamic Coastal Shelf Environments. J. Mar. Sci. Eng. 2021, 9, 336. https://doi.org/10.3390/jmse9030336
Moore SK, Mickett JB, Doucette GJ, Adams NG, Mikulski CM, Birch JM, Roman B, Michel-Hart N, Newton JA. An Autonomous Platform for Near Real-Time Surveillance of Harmful Algae and Their Toxins in Dynamic Coastal Shelf Environments. Journal of Marine Science and Engineering. 2021; 9(3):336. https://doi.org/10.3390/jmse9030336
Chicago/Turabian StyleMoore, Stephanie K., John B. Mickett, Gregory J. Doucette, Nicolaus G. Adams, Christina M. Mikulski, James M. Birch, Brent Roman, Nicolas Michel-Hart, and Jan A. Newton. 2021. "An Autonomous Platform for Near Real-Time Surveillance of Harmful Algae and Their Toxins in Dynamic Coastal Shelf Environments" Journal of Marine Science and Engineering 9, no. 3: 336. https://doi.org/10.3390/jmse9030336
APA StyleMoore, S. K., Mickett, J. B., Doucette, G. J., Adams, N. G., Mikulski, C. M., Birch, J. M., Roman, B., Michel-Hart, N., & Newton, J. A. (2021). An Autonomous Platform for Near Real-Time Surveillance of Harmful Algae and Their Toxins in Dynamic Coastal Shelf Environments. Journal of Marine Science and Engineering, 9(3), 336. https://doi.org/10.3390/jmse9030336