1. Introduction
The economies of Mediterranean countries show a high dependence on the tourism industry. This dependence leads to overexploitation of coastal resources and especially the beach-dune systems. Human actions with an impact on the sandy coasts and their dunes are gathered in five groups [
1]: (1) Housing and recreation; (2) industrial and commercial use; (3) waste disposal; (4) agriculture, aquaculture, and fisheries; and (5) military activities. The effects of tourism on coastal ecosystems and specifically on the degradation of coastal dune systems have been treated by different authors [
2,
3,
4], among others. The coastal squeeze [
4,
5] to which these environments are subjected, due to the long-term processes as sea-level rise, land subsidence, sediment deficit, and anthropic infrastructures jointly to the different tourism activities that take place there (bath, bars and restaurants, toilets and showers) and the management needs (mechanical cleaning of the beaches, construction of breakwaters, artificial feeding of the beaches...) cause its slow and progressive physical and biological degradation. These activities alter natural processes and coastal dynamics, eliminate topographic variability, fragment, degrade or eliminate habitats, reduce diversity and threaten endemic species [
6,
7].
In Spain, studies on the degradation of coastal dunes date back to the 1970s, see the compilations from References [
8,
9] for an overview, and in recent years the interest and popular pressure for the protection and restoration of coastal dunes are exemplified by the publication of the “Coastal dune restoration manual” [
10] by the Ministry of the Environment. In recent years, and due to the number of dune restoration projects that have been carried out on the Spanish coast, different works have been published on the state of vulnerability, and the assessment of different restoration projects, both on the Atlantic [
11,
12,
13,
14] and Mediterranean coasts [
15,
16,
17,
18].
In littoral geomorphology, the restoration projects of coastal dunes try to achieve a recovery of the exchange of sediment between the beach and the dune (foredune) and recover the dynamic profile of the beach-dune system. Natural ecosystem restoration occurs through similar processes that affect the integrity, functionality, and resilience of the ecosystem [
6]. After Putsy and Silveira [
19], the objective of foredune restoration is to restore the heterogeneity of features in the temporal context. This heterogeneity is achieved by the continual evolution of the beach-dune system. According to SER and UICN [
20,
21], the main methods used in the evaluation of restoration include the study of the ecosystem through: (i) The structure and composition of the plant community; (ii) species during plant succession; or (iii) the resilience after natural regeneration of the restored ecosystem.
These systems, due to their high dynamism, would have to present a high degree of resilience at the first and early stages. The term “resilience” [
22] is used here as “the ability to recover the initial state from a disturbed state to the cessation of the disturbance that caused the change of state” [
23]. Coastal managers are interested in landform development (abiotic barrier) over yearly to decadal temporal scales [
21,
24], but coastal dunes are systems where multiple interacting controls, internal and external to the system, occur at different scales. In a dune system, the goal of the different measures applied is to recover both the geomorphological dynamics, which includes the recovery of morphologies of this type of system, and the activity of the sedimentary dynamics represented by the transport and deposition of sediment, as well as the development capacity of its plant community, characteristic of each of its sub-environments. The recovery and/or restoration of a disturbed site can be approached through different methodologies, in relation to its initial state, the degree of affectation, and the time that has elapsed since anthropic activity ceases [
25].
In this case, spontaneous succession or passive re-vegetation was sought, where colonization and natural succession of plants are allowed, and human action did not intervene. The use of spontaneous vegetation succession in an ecological restoration program requires different points to be addressed [
26], such as: (1) Objectives, processes, and time frame within which the restoration objectives must be achieved; (2) environmental characteristics of the site and how these may or may not influence the succession; (3) the degree of possible generalization and the level of predictability between sites and patterns of succession; (4) communication between the different acting parties (researchers, professionals and authorities, including education and public awareness; and (5) post-project monitoring and evaluation. The monitoring allows us to monitor the development of the restored dune vegetation and to compare the composition and abundance of exclusive dune species with those of the reference dune systems [
12].
The measures proposed for the recovery and restoration of the coastal dune system were and are those of soft management: (i) Prohibition of visitors from walking or crossing the dunes; (ii) delimitation of the dune zone with posts and ropes. Visitors can use the beach for swimming or sunbathing, provided that they use the permitted trails and do not enter the dune area; and (iii) manual cleaning of anthropic remains and inappropriate litter that reaches the beach. In the dune zone, cleaning actions are carried out even more carefully—i.e., less frequently. The proposed actions represent a greater effort of supervision by the environmental authorities, but the dune systems may have more value as habitats if they host sub-environments representative of natural gradients [
27].
In this work, we studied the resilience capacity of the coastal dune system from the protected area of the beach of Remolar (Viladecans, Barcelona), taking into account the evolution of morphology and vegetation of the dune system at decennial term. This started in 2003, when a drastic change in the use of this beach took place, since the enlargement of the Barcelona airport led to closing the direct arrival paths to the beach, and thus, to a dramatic decrease of the leisure use of the beach. The following year, a study was carried out consisting of the realization of topographic profiles and a study of the vegetation of the dune system [
28]. Ten years later, a comprehensive study was performed to evaluate the dynamics of the dune system.
2. Study Area
Remolar beach has a well-developed dune-beach system, linked to the fluvial plain of the Llobregat River [
15,
29]. It is located on the southern lobe of the Llobregat River delta, 5.7 km SW from the mouth. The Llobregat river has a typical Mediterranean regime [
30], influenced by seasonal (autumn) and sporadic torrential rainfalls. The southern lobe of the Llobregat river delta has a shoreline about 17 km long. The construction of infrastructures, notably the Barcelona airport, the extension of Barcelona harbor, and the growth of industrial and urban areas in the delta and its surroundings, transformed their landscape [
30].
The southern Llobregat delta functions as a littoral cell. With the contribution of sediment at the mouth of the river, the port of Barcelona with its piers makes it impossible to transport sediment from the north, which on the southern hemi-delta coast is from ENE to WSW. The end of the littoral cell is the port of Ginesta (built from 1986) acts as a littoral drift barrier, where we find the widest beaches in the delta, reaching more than 150 m [
31].
The beach of Remolar has a length of 700 m and is limited by the breakwater dams that protect the inlet from the lagoon of Remolar to the NE and the channelized stream of Sant Climent to the SW. The beach-dune system has a width greater than 110 m, of which between 70–93 m are occupied by the dune system and the rest by the beach (
Figure 1). In total, the Remolar beach-dune system takes an area of about 75,000 m
2, of which 53,000 m
2 were occupied by the dune system in 2013. From 1960 to 2004, the dune system was limited on its landward side by a fence, still existing, that delimited a campground. This fence had several direct exits to the beach. The result (
Figure 1) was a degradation of the dune system in the occupied area (decreasing until 42,000 m
2 in 2003), in the dune morphology, and in its vegetation cover. The extension of the Barcelona airport involved the cessation of the activity of the camping and the entrance as manager of the beach of the Environmental area of the City Council of Viladecans.
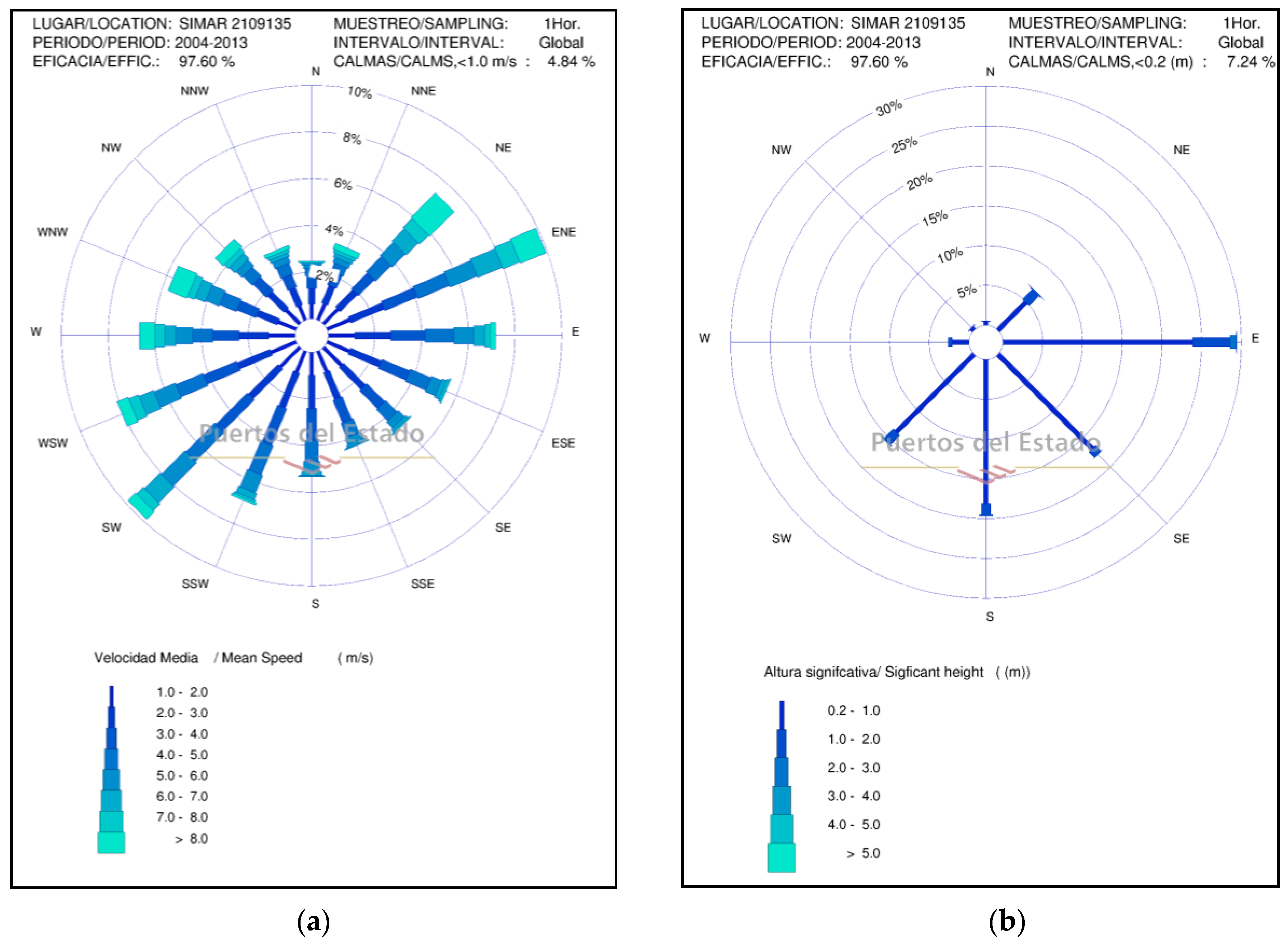
The characterization of the dynamic agents (wind and waves) of the study area has been carried out using data from the official web “Puertos del Estado” of the Ministry of Transport, Mobility and Urban Agenda of the Government of Spain (
http://www.puertos.es/es-es/oceanografia/). The wind system in the area is dominated by the dipole ENE (10%)-NE (7.5%) and SW (9%)-WSW (8%) directions (
Figure 2a). The modal wind speed is between 2–3 m·s
−1 (7.2–10.8 km·h
−1), the maximum values exceed 22 m·s
−1 (79.2 km·h
−1) and centered its presence during December and January. Waves (
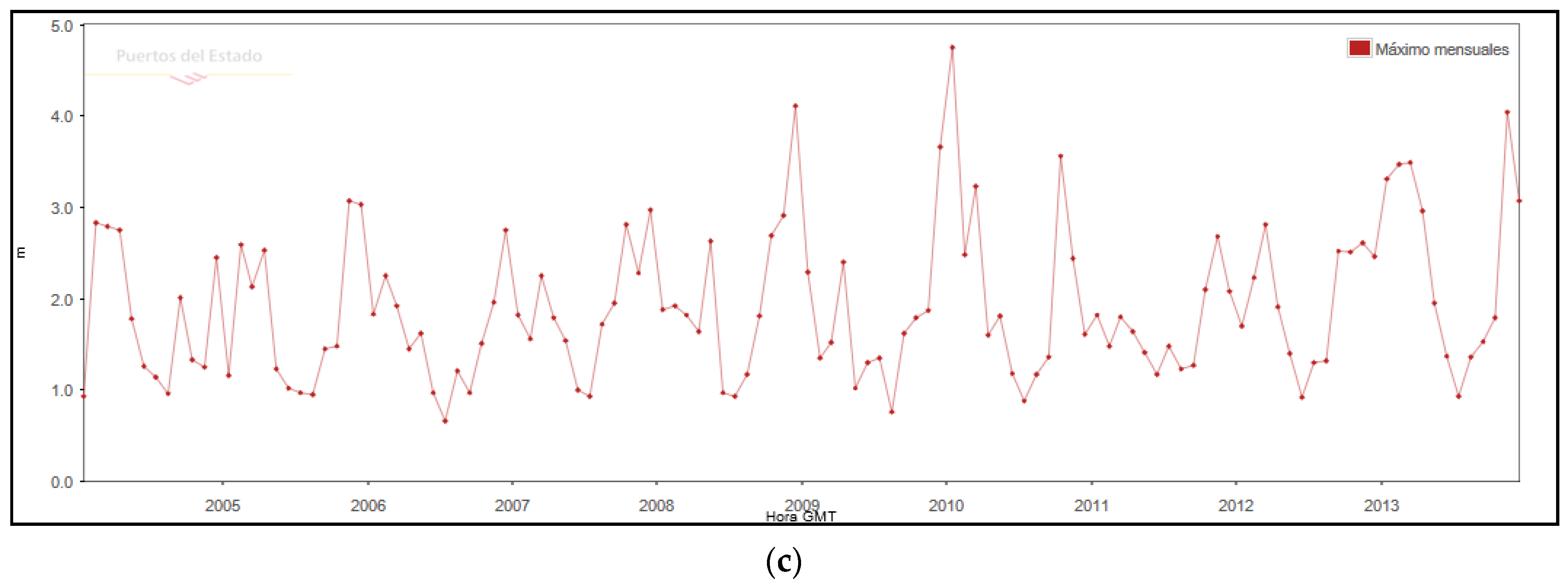
Figure 2b) are characterized by a significant height (Hs) of less than 0.5 m (51%) and a period of between 4–5 s. The predominant directions are from SW (16%) and E (15%), and only 4% of the waves exceed 1.5 m, the predominant storms being in E-ENE direction. The activity of the storms has been controlled based on the maximum Hs time series, between the periods 2004–2013 (
Figure 2c). From the series, it is extracted that in the decade studied, there have been 39 storms greater than 2.0 m of Hs, wave height that is considered a storm on the coast of the Llobregat delta [
32,
33], 14 with a height greater than 3.0 m, 5 greater than 3.5 m, and 3 greater than 4.0 m. These storms cause a run-up [
34] of between 0.6–1.5 m for Hs > 2.5 m and a rise of 1.0–2.4 m in storms of Hs > 4.0 m. Such run-up heights are capable of producing complete erosion of the embryonic dunes and producing erosion scarps and washover deposits in the primary dune.
Following the aforementioned recommendations by Reference [
26], we present a reference model of the spontaneous vegetation of the dunes in the studied area of the Remolar beach, and it is important to consider the distribution of plant communities on a beach where disturbance has been minimal. The dunes studied in the Remolar area are classified within the type of “dune ridges with semi-fixed dunes” according to Reference [
29]. Following several authors [
29,
35] the model of plant communities found is: (1)
Agropyretum mediterraneum, the grass-sand community in the embrionary dunes; (2)
Ammophiletum arundinaceae, the community of marram grass located on the top of the dunes, with species, such as
Ammophila arenaria—marram grass- and
Pancratium maritimum; (3)
Crucianelletum maritimae, the community located on the leeward side of the primary dune, the secondary dune and more stabilized dunes; and these give way to (4) pinewood (stands of
Pinus pinea, at places planted by man) or to juniper scrub.
3. Methods
To assess the impact of the measures implemented on the dune ecosystem, between 2013 and 2014, we carried out a study of the state of the (1) morphology, through topographic surveys; (2) grain-size distribution of the dune system; (3) internal structure of the dunes, using GPR (ground-penetrating radar) profiles; and (4) dune vegetation, in terms of plant communities. All this information has been compared with the work of Gonzalez et al. [
28]. The methodology used to carry out the topographic profiles and vegetation studies has also been the same.
The methodology to carry out the topographic profiles is the same used by Reference [
28] in a previously performed study, namely, we performed five transects located perpendicular to the coast (
Figure 1). A spatial reference system was created using 2 m high wooden rods, firmly fixed every 10 m along with the profile, and joined by a rope that was kept horizontal with the help of a bubble level. Height measurements were taken from the rope to the sand every 20 cm, considering the sea-level as 0 m high. The validity of the system is verified by the good correlation in the profiles of the different years in the first meters, an area crossed by a path of compacted material that has not changed over time (
Figure 1). Due to the soft behavior of sand, topographic measurements can have up to 1% percent error, which is carried over into volume calculations. The topographic profiles of the two study years have been compared both volumetrically and morphologically. Volume calculations referred to sand gains, or losses have been made per linear meter of profile (m
3·m
−1).
The characterization of the surface grain size was performed in profiles 1 and 5, collecting samples distributed along with each topographic profile, 15 and 14 samples per profile, respectively. The samples were analyzed by a COULTER LS230 laser diffractometer, and their system software supplied the main statistical parameters as the mean (
), the mode, and the standard deviation (σ). The grain size analysis implies a pre-treatment of the sample with H
2O
2, for the elimination of organic matter, and shaking the sample with sodium pyrophosphate avoids flocculation. To know the dynamics of the sediment and its classification, due to transport using the wind, the sorting or classification parameter was calculated based on the mean (
) and standard deviation (σ) (Thus:
+ σ/
− σ), similar to the sorting parameter of Trask [
36], where lower values indicate better sediment classification. The relationship between the mean grain size and the dispersion of the sample is visualized by the coefficient of variation (CV: (
)*100).
The present work gives us the opportunity to study the regenerated dunes both in their morphological and sedimentological dynamics. For this and to know the internal structure of the new dune bodies, GPR transects were made in profiles 1 and 2. A Georadar (SIR 3000) was used, equipped with two types of antenna that provide data of the signal that was processed, filtered, and amplified. This allowed us to analyze and visualize the different sedimentary levels in the subsoil. In this work, an antenna of 400 (MHz) was used. Thus, the RPM (reflection profiling mode) configuration was used. Said configuration consists of two mobile antennas that are separated by a fixed distance along the surface of the terrain. These antennas send pulses (shots) to the ground and capture the reflection signal. All shots were stacked and displayed in the GPR profile. The signals obtained have been clear throughout the profile. GPR data is processed for correction of low-frequency and DC bias in data (Dewow), for correction of start time to match with surface position (time zero correction) and filtered to remove noise from the acquired signal and to improve and visual quality (filtering 1D and 2D). Furthermore, the radargram is adapted to the terrain topography and selected for the appropriate gains for data display and interpretation. The penetration depths depend on the propagation velocities, and these on the dielectric properties of the sediments. For sands, such as those of dune-beach systems, propagation velocities of 0.10 to 0.20 m·s
−1 have been proposed [
37,
38]. In coastal dunes, changes in the dielectric properties of sediments are produced by saturation with seawater [
38]. In the present work, to favor resolution against penetration, a 400 kHz antenna has been used, as indicated above, and we consider a penetration of 0.2 m·ns
−1 and a resolution of less than 20 cm. With this resolution, the reflections can be interpreted as the product of primary depositional fabric and sedimentary structures [
39].
Sampling and collection of vegetation data were carried out at different visits during May 2014, within the period of maximum growth and flowering (March-July). For the sampling of plant communities, we set squares of 0.5 0.5 m every 2 m in each of the profiles, from the point of origin (inland fence) to the point of absence of vegetation, 70–100 m length, and between 39–47 quadrants per profile with a total of 225 quadrants for the entire study area. In each square, we obtained a vegetation inventory, namely, we recorded a complete list of the species present and evaluated their abundance in terms of cover. For this, we used a simplified scale of three value ranges, inspired in the phytosociological method, with the following meaning: (1) For a covering between 0–10%; (2) for a covering between 10–50%; and (3) for a covering between 50–100%.
Since vegetation analysis was through Fuzzy groups, where the species cover values have a low weight in front of their presence, we preferred this value simplification to obtain more inventories. The analysis of the plant communities has been carried out using the statistical software GINKGO [
40,
41], in order to form groups of related inventories according to their plant species composition. Then, we assembled these vegetation groups to the phytosociological schemes of vegetation [
28,
40]. For the allocation of the inventories to each group (plant association), we used the GINKGO fuzzy clustering analysis (Fuzzy-c Means algorithm). The grouping criterion to obtain the optimal fuzzy partition (number of inventory groups) is associated with the least squared error function, which produces a Bray-Curtis distance matrix. The descriptive determination of the communities has been carried out considering the results of the matrix of the
Phi coefficient of fidelity (φ) [
42]. “Fidelity” is the degree of preference of each species to each given plant association.
The representation of the results obtained in the statistical analysis on the vegetation has been plotted on each profile. Then, with the support of field assessment and current orthophotos, we interpolated the distribution of the vegetation units between profiles, to draw up a vegetation map. The data treatment has been processed with the Geographic Information Systems software gvSIG.
5. Discussion
The abiotic and biotic characterization of the Remolar coastal dune system through geomorphology, grain size, internal sedimentary structure, and the distribution of vegetation carried out between the years 2013–2014 and its comparison with existing data from 2004 [
28] allow us to study the evolution of the ecosystem in the last 10 years and evaluate the effectiveness of the restoration measures applied.
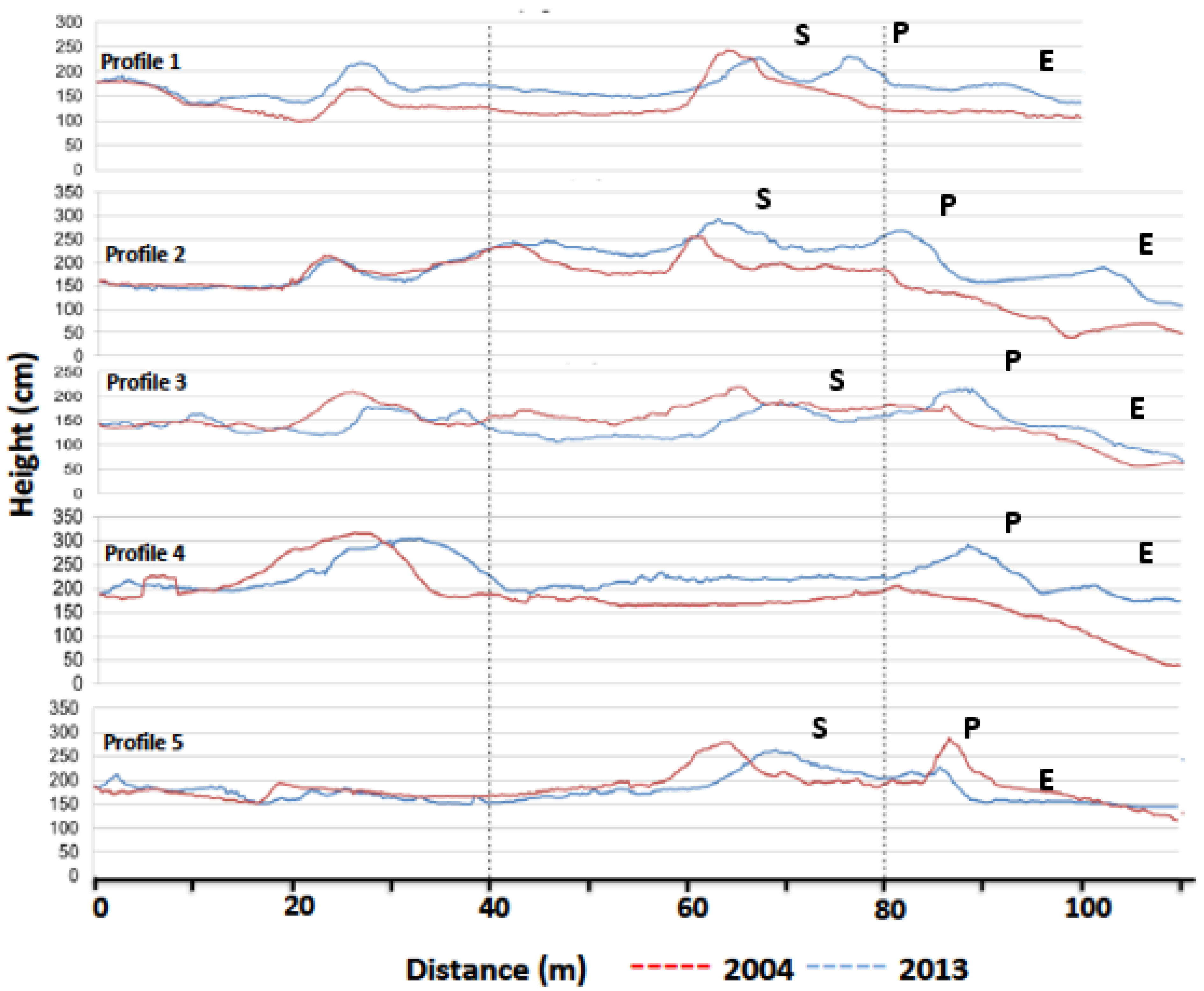
The fundamental requirements for forming coastal sand dunes are an abundant sediment supply and sufficiently strong wind to transport it from the beach to the hinterland [
46]. These requirements are given in the case of the Remolar beach, as shown by the dune system formed in the backshore of the beach. Morphologically, the dune system has originated a new primary dune, present throughout the beach as of 2006 (
http://www.icc.cat/vissir3/) and which remains until the completion of this work, and much of an embryonic dune ridge. This embryonic ridge appeared temporally interrupted by erosion scarps, as a result of winter storms. The profiles also show a volumetric increase (
Table 1) of the dune system, which has resulted in aggradation and progradation processes (
Figure 3) in most of the dune system. In this sense, we consider that the washover processes, originated during storms, are one of the main sediment supply processes at the landside of dunes. The effectiveness of the process is clearly observed in the two GPR profiles (
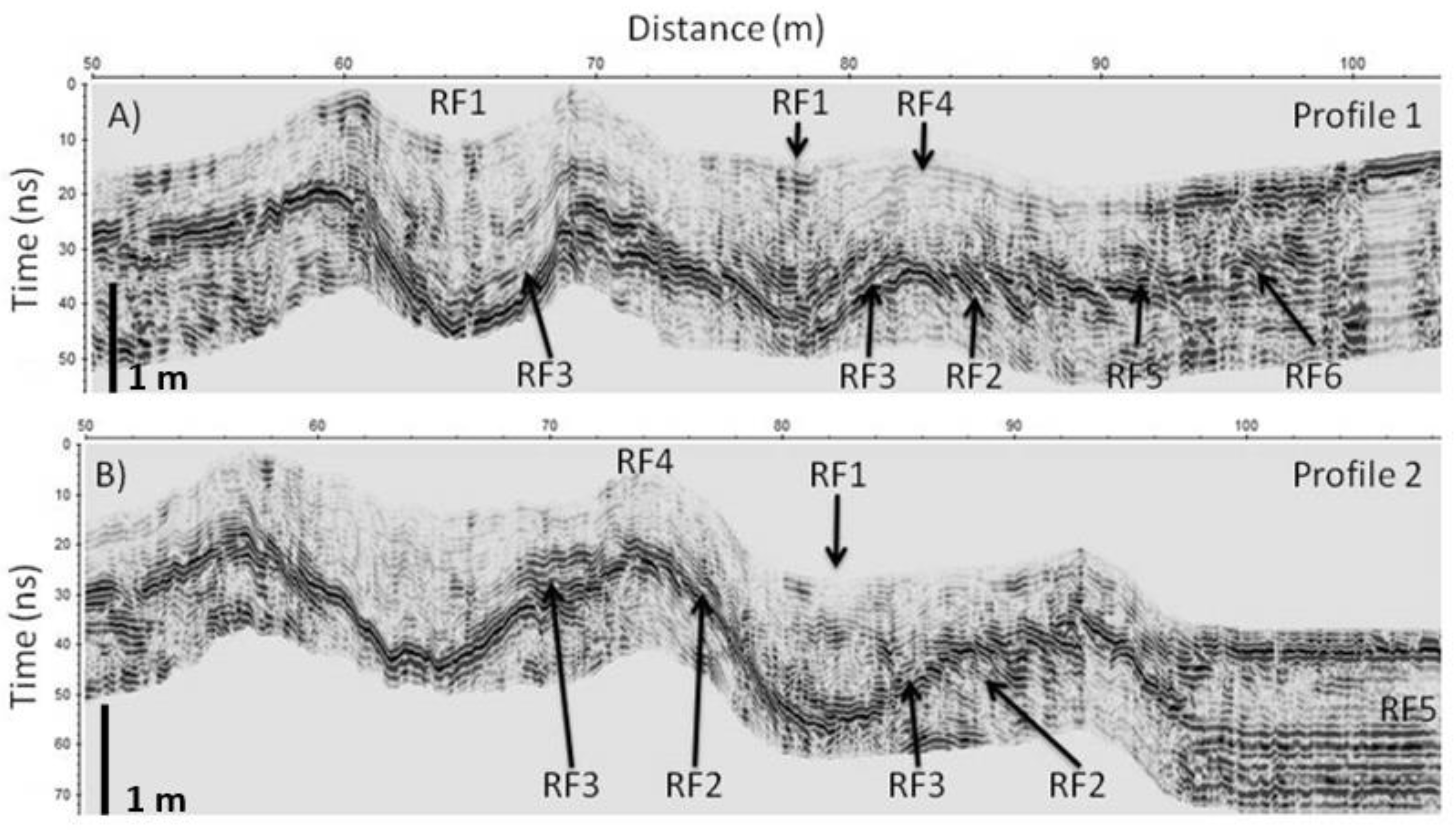
Figure 6), which present reflectors in the embryonic and primary dunes that fill the interdunar spaces through the RF3 facies.
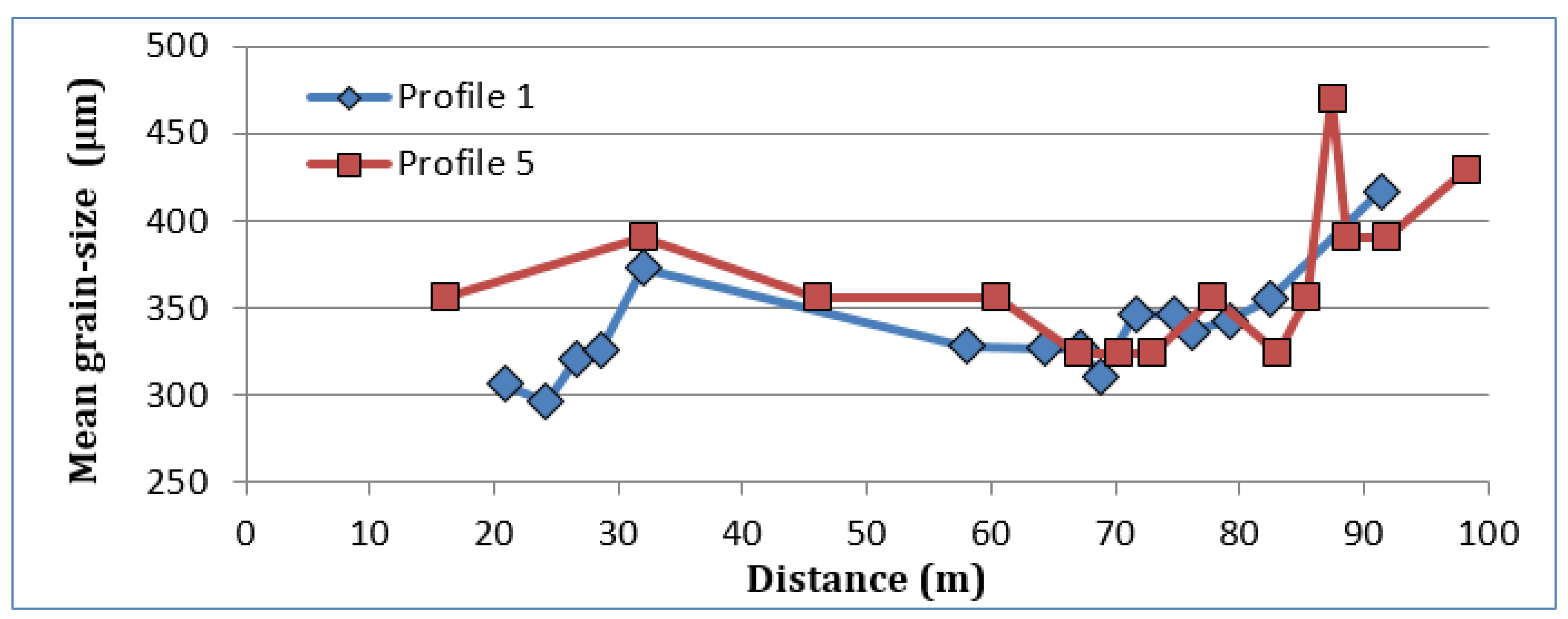
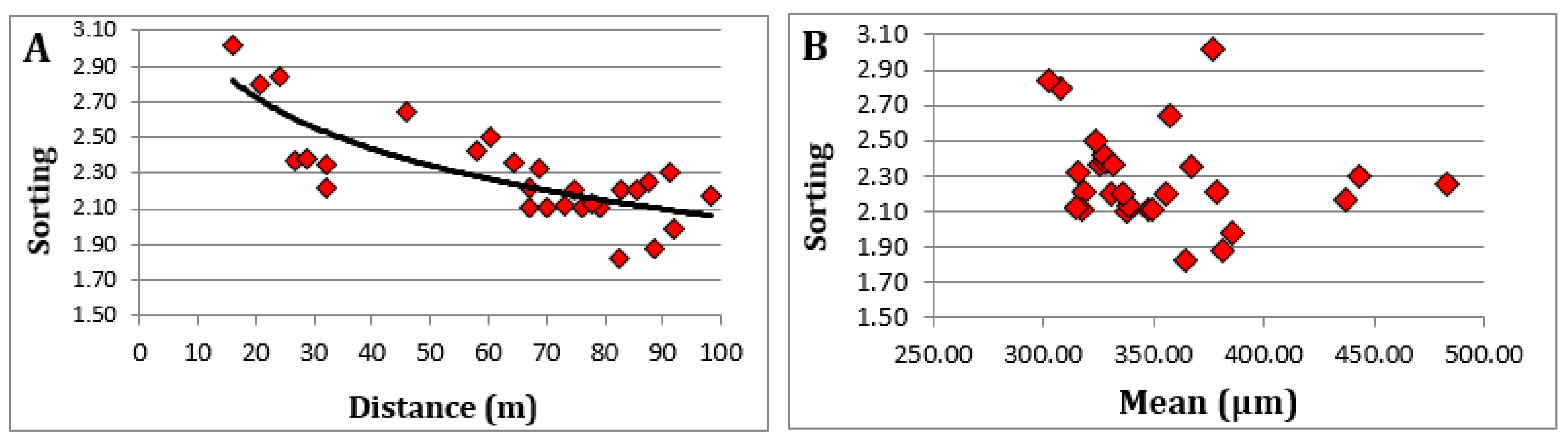
As for the sand, the grain size distribution shows (
Figure 4) a mean grain size cent red over 350 μm and showing an increase at the crest of the dune ridges [
47,
48]. The evolution of the granulometric distribution shows higher values in the dry beach decreasing towards the leeward areas of the primary dune (
Figure 4). We interpret this decrease as the concentration of transport on the seaside of the primary dune [
46,
47,
48]. The result of the concentration of sediment deposition on this side of the primary dune is observed in the GPR profiles with the presence of the RF2 facies [
38]. This facies shows the growth and migration of the new primary dune formed since 2004, as can be seen visually in the 2006 photo (
Figure S1, Supplementary Material). The distribution range of the mean grain sizes, between 296 and 471 m, within the granulometric class of the medium sand [
43], throughout the dune system explains the capacity of the area’s active winds to transport practically the entire granulometric range of sediments contributed by marine dynamics. It is evident that the areas closer to the marine area (backshore, embryonic and primary dunes) show a better classification of sediments (
Figure 5) with sorting values below 2.30. The importance of the different processes acting in a dune system, such as the edaphization processes, supply of organic matter, and calcium carbonate animal remains (mainly snail shells), explain the low relationship between grain size and sorting of the sediment.
As mentioned above, the Remolar beach-dune system holds the contribution of sediment on the beaches by the inputs of sand by coastal drift from the mouth of the Llobregat River. Currently, the beach shows moderate progradation dynamics (RF5 facies,
Figure 6), which in turn creates space on the dry beach and allows sediment to accumulate (RF2 facies in
Figure 6) on the seaside of the embryonic dune that deposits sediments on the landside of the dune (RF3 facies in
Figure 6). The embryonic and primary dunes of profiles 1 and 2 (
Figure 6) show the formation and growth process by repeating the successive accretion cycles on the seaside and by the deposition of sediments by washover processes in the landside, a process that produces a gradual filling of the interdune depression. In all, reflective beaches, such as Remolar, are characterized by low waves and narrow width; they usually present a single embryonic ridge [
46,
47]. The interrelation between morphology and vegetation is observed in the crest of the different dune ridges, where the RF4 facies clearly appear, identified as facies originated by the deposition of sediment through vegetation and root activity [
38]. In our case (
Figure 7), it would be mainly due to the presence of
Elymus farctus in the embryonic dune and of
Medicago marina in the primary dune, two species with a large root volume. In addition to the relationship between dune morphology and vegetation, certain studies [
49,
50,
51] have shown the relationship between grain size and plant zonation [
52], in our case the textural differences between the crests of the embryonic and primary dunes contrast with the parts more internal to the dune system, due to the edaphic processes.
According to Reference [
28] in 2004 at Remolar, due to anthropogenic disturbances, the typical disposition of plant communities was strongly altered, highly dislocated, highly impoverished into characteristic species, and bastardized by species that are not properly psammophilous. To solve the problem, in Reference [
28], only the two dominant vegetation types have been differentiated, which are those corresponding to mobile and embryonic dunes (white dunes) and those that develop into more or less fixed or stabilized dunes (gray dunes). Vegetation from mobile and embryonic dunes includes two communities—the
Cypero mucronati-Agropyretum juncei community and the
Medicagini marinae-Ammophiletum australis community. The
Cypero mucronati-Agropyretum juncei community is composed of
Elymus farctus, accompanied by
Cyperus capitatus,
Polygonum maritimum,
Sporobolus pungens,
Euphorbia peplis, and
Calystegia soldanella. The
Medicagini marinae-Ammophiletum australis community is composed of marram grass,
Ammophila arenaria, the most characteristic species of this community, but that in the beach of the Remolar is a rare species that forms some bush in the innermost zones, the remaining species of this community, as
Echinophora spinosa,
Medicago marina or
Eryngium maritimum, being more represented. The vegetation of the more or less stabilized dunes is represented only by the community of
Crucianelletum maritimae, composed of some shrub species, among them
Crucianella maritima, accompanied by species like
Helichryssum stoechas, that announce the transition towards the coastal pine forest.
In our study, we can observe that the identified plant communities showed (
Figure 7), consistently between the different transects, an arrangement in parallel longitudinal bands, or zonation following the previously presented model [
29,
35]. In this sense, the interrelation between geomorphic controls and the spatial distribution of vegetation is highlighted as a main determinant of the development of the system [
12,
38,
53,
54,
55,
56].
The position of the vegetation of embryonic dunes (
Cypero mucronati-Agropyretum juncei) stands up in our study in the form of an almost-continuous strip near the coast. The plants of this community are characterized by having a great capacity for colonization, given the harsh environmental conditions that determine the foredune [
53,
55]. González et al. [
28] they already found this community then, but then it occupied a wider strip with a more inland location. It is worth mentioning the importance of this community as a nesting place for the Kentish plover (
Charadrius alexandrinus), a species recorded in the Spanish Red Book of Birds [
57] as a vulnerable species.
The vegetation characteristic of primary dunes (
Medicagini marinae-Ammophiletum australis) was found in its expected location, occupying a continuous strip along the entire beach of Remolar, where the environmental conditions are not as extreme as in the foredune. The plant species of this community develop systems that guarantee efficient water use (xeromorphic leaves, extensive root systems), that allows a medium vegetation cover, thus stabilizing the primary dunes, increasing sedimentary deposition, and favoring their growth [
51,
53,
55]. In 2004, González et al. [
28] located this community in the same area of the Remolar where we currently found it, except in some areas where its location occupied a more interior position. The absence of the typical species of this community
Ammophila arenaria and
Euphorbia paralias has been confirmed, but there is a good representation of the rest of the species of this community, such as
Medicago marina, Silene niceensis, or
Eryngium maritimum.
The secondary dune community with
Crucianella maritima is the best-represented plant assemblage on the Remolar beach, occupying the back of the primary dune community. Both in our study and in that of Gonzalez et al. [
28], this community in the Remolar beach includes a good deal of
Cyperus capitatus, a species most typically found in less evolved dunes. Probably, the legacy of continued trampling affects negatively the populations of
Crucianella maritima and associated species, and favors the persistence of
Cyperus capitatus. This plant, due to its rhizome system giving aerial tillers, better resists frequentation and human disturbance, and contributes to effectively restoring the dunes. This community appears widely distributed in the Remolar, occupying a large part of the area that would correspond to the
Crucianelletum, in which has enlarged from 2005.
Behind the dunes, vegetation is more protected from the action of the wind and marine aerosols, and nutrients and water are better conserved by more efficient soils. Moreover, despite the access to the dune system is currently restricted; we still found a large area occupied by ruderal and opportunistic communities, as a vestige of the ancient anthropic pressure (
Figure 7). All this allows for greater diversity, both in species and in ecological groups. Plant communities include shrubby species (chamaephytes) together with plants related to anthropogenic influence. Among these species, we found
Plantago coronopus,
Salsola kali,
Bromus madridensis, and introduced species, such as
Carpobrotus edulis. The highest degree of ruderalization is found in areas where the allien
Ambrosia coronopifolia predominates, which (due to its rhizomatous nature) almost forms monospecific populations.
Finally, González et al. [
28] mention “an incipient pine forest with low elevation but vigorous pine trees” see
Figures S1 and S2 (Supplementary Material), which has been transformed into well-formed pine stands in a maquia shape, which is practically mono-specific in nature with a high density and in which only shade-tolerant species appear in the understory, such as
Pleurochaete squarrosa or
Aetheorhiza bulbosa.
6. Conclusions
The restored coastal dunes of El Remolar beach have undergone a process of resilience. If we consider the definitions given above (see
Section 1), the dune system studied would show a high degree of resilience, having recovered much of the initial characteristics of a coastal dune system: Morphology, vegetation, dynamic behavior, the interrelation between morphology and vegetation, and heterogeneity. Therefore, the good resilience ability made that the measures of restoration and regeneration applied have had success.
Specifically, at the sedimentological level, the system works in a “natural” way, with the formation of a new dune ridge (primary dune) present along the entire beach and parallel to the coastline, and an embryonic dune absent only in the sections eroded by sea storms. The restoration measures applied have proven to be an effective measure for increasing the volume of sand accumulated in the dune system, without the need for artificial nourishment actions. The GPR profiles show the binomial of progradation and aggradation that occurs in the formation and growth of the dune ridges, with accretion on the side of the dune and deposition by slides or washover processes on the land side of it.
The vegetation in the dune system has spontaneously improved during the last decade. The phytosociological results have differentiated seven well-established plant communities, occurring on the dune system in a functional way, i.e., set in bands parallel to the coast. But despite the imposed restoration measures, the anthropic influence manifested by the opportunistic and ruderal communities is still alive after a decade.
The dynamics of the beach-dune system, represented in the Remolar beach by the presence of the new dune ridges of primary and embryonic dunes and their specific plant communities, demonstrates the need for the geomorphological connectivity of the chain formed by the beach, the dry beach, the embryonic dune, and the primary dune. The interruption of this concatenation of morphologies produces a degradation of the entire system and its affectation both to the morphology and to the entire vegetation of the system.
Therefore, we consider that after ten years of maintaining the simple and cheap measures of restoration and protection, the effects of trampling have disappeared, and that the ecosystem has recovered and contains the biotic and abiotic resources to continue its development [
20]. The aforementioned problems of introduction of typically anthropic plant types are currently generalized to most of the dune systems in the area [
29].


 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}