Relationship Between Tree Size, Sediment Mud Content, Oxygen Levels, and Pneumatophore Abundance in the Mangrove Tree Species Avicennia Marina (Forssk.) Vierh


Abstract
1. Introduction
2. Materials and Methods
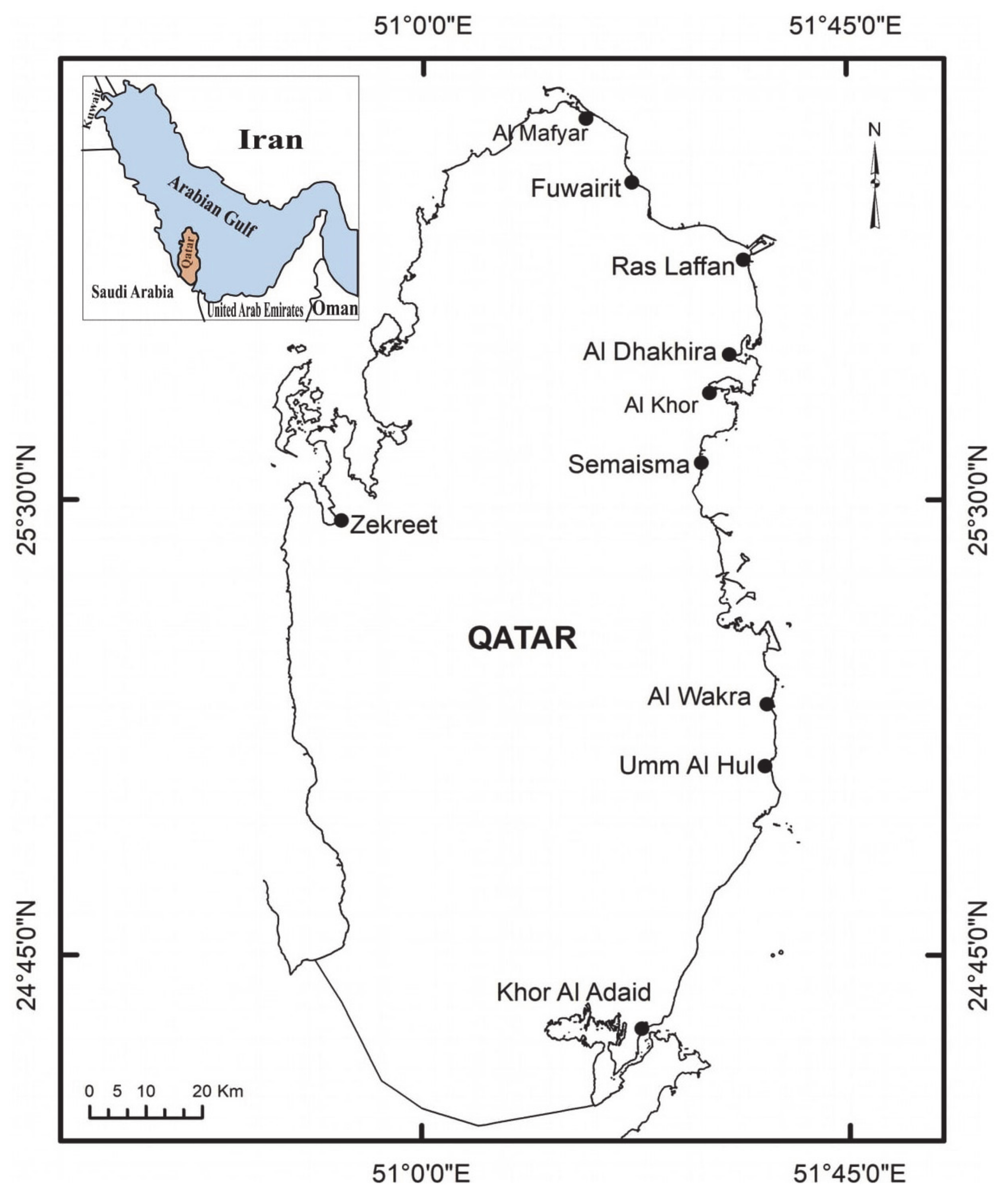
2.1. Study Species and Area
2.2. Data Collection
2.2.1. Environmental Conditions
2.2.2. Pneumatophore Abundance and Crab Burrows
2.2.3. Statistical Analyses
3. Results
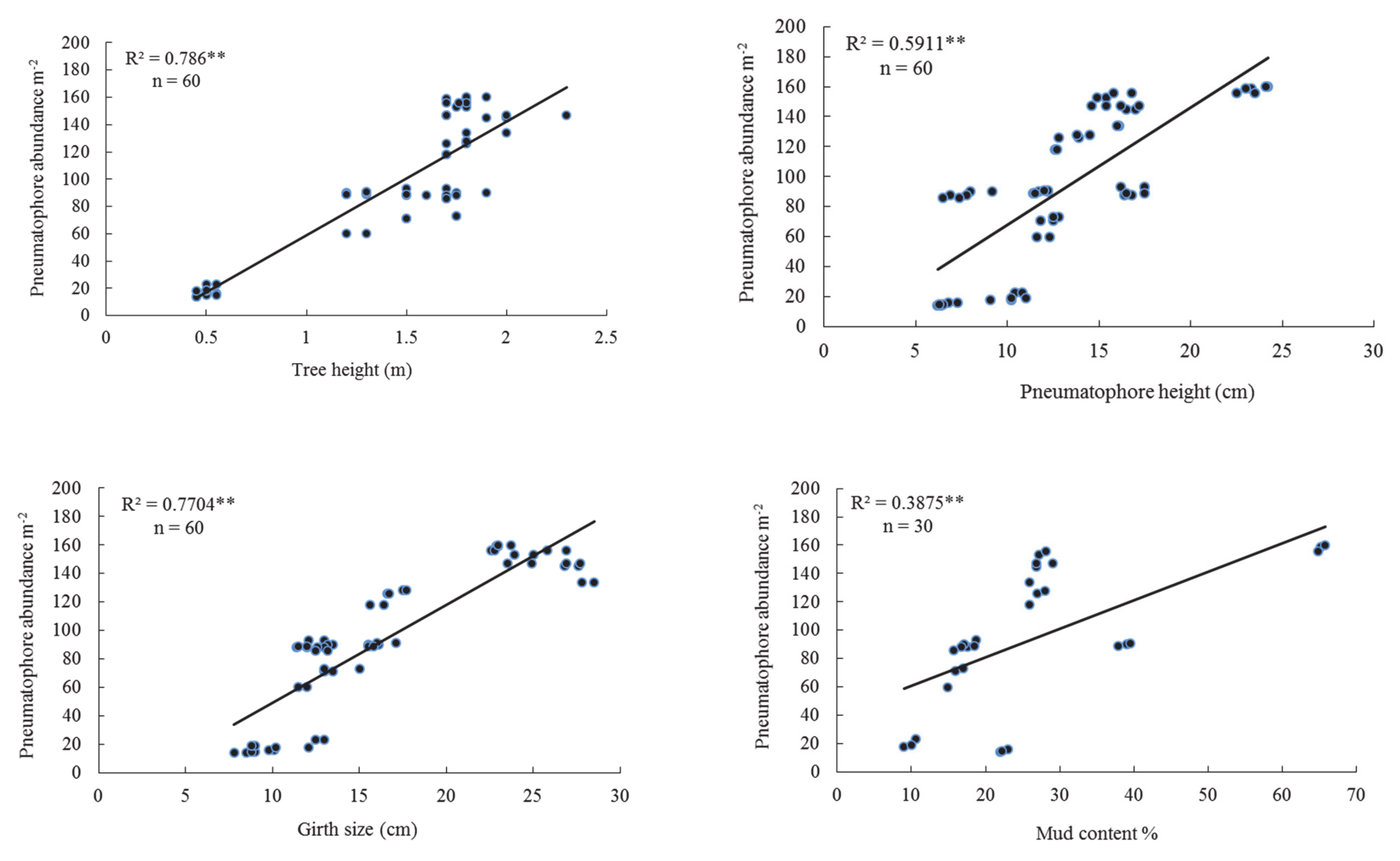
3.1. Upper Tidal Level
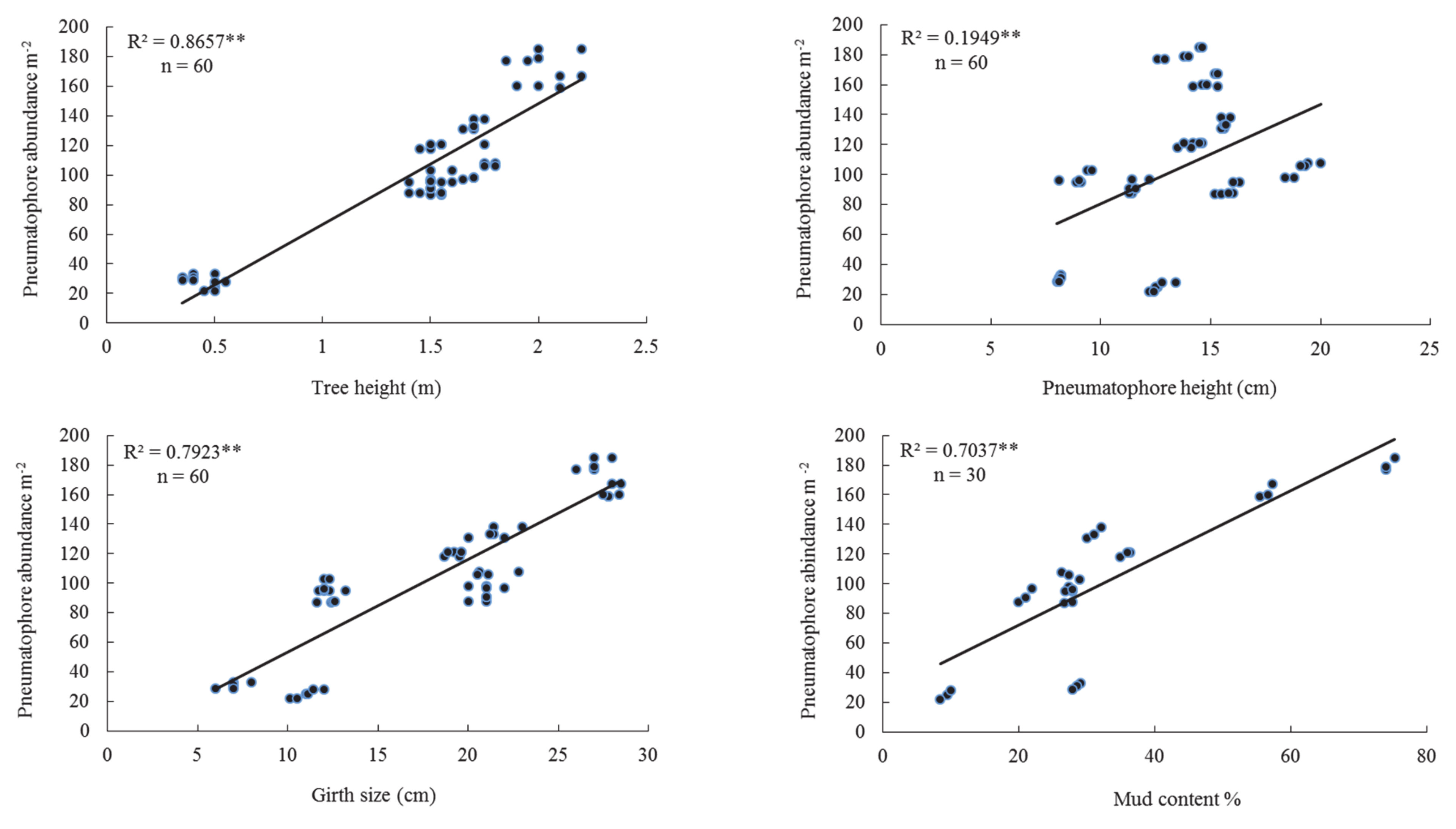
3.2. Middle Tidal Level
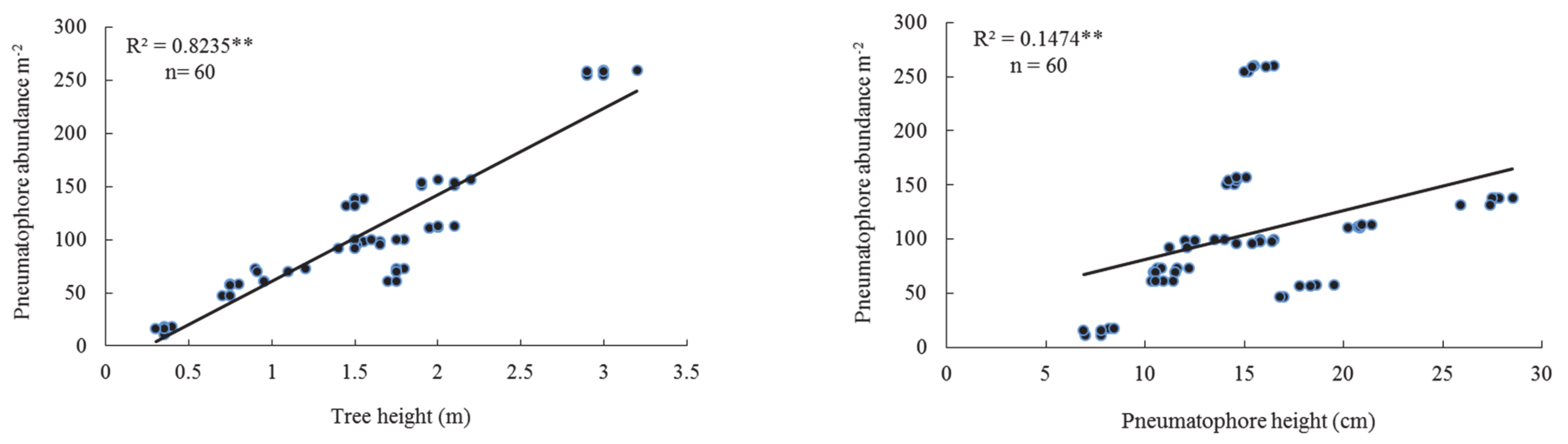
3.3. Lower Tidal Level
3.4. All Tidal Level Sites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alongi, D.M. The Impact of Climate Change on Mangrove Forests. Curr. Clim. Chang. Rep. 2015, 1, 30–39. [Google Scholar] [CrossRef]
- Xia, P.; Meng, X.; Li, Z.; Feng, A. Sedimentary Records of Mangrove Evolution during the Past One Hundred Years Based on Stable Carbon Isotope and Pollen Evidences in Maowei, SW China. J. Ocean Univ. China 2016, 15, 447–455. [Google Scholar] [CrossRef]
- Toosi, N.B.; Soffianian, A.R.; Fakheran, S.; Pourmanafi, S.; Ginzler, C.; Waser, L.T. Comparing Different Classification Algorithms for Monitoring Mangrove Cover Changes in Southern Iran. Glob. Ecol. Conserv. 2019, 19, e00662. [Google Scholar] [CrossRef]
- Almahasheer, H.; Duarte, C.M.; Irigoien, X. Phenology and Growth Dynamics of Avicennia Marina in the Central Red Sea. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef]
- Clough, B.B. Mangrove Ecosystems in Australia: Structure, Function and Management; Australian National University Press: Canberra, Australian, 1982. [Google Scholar]
- Kathiresan, K.; Bingham, B.L. Biology of Mangroves and Mangrove Ecosystems. Adv. Mar. Biol. 2001, 40, 81–251. [Google Scholar]
- Srikanth, S.; Lum, S.K.Y.; Chen, Z. Mangrove Root: Adaptations and Ecological Importance. Trees 2016, 30, 451–465. [Google Scholar] [CrossRef]
- Wada, H. Soil-Water-Plant Relationships of Mangroves in Thailand. Galaxea 1988, 7, 257–270. [Google Scholar]
- Allaway, W.G.; Curran, M.; Hollington, L.M.; Ricketts, M.C.; Skelton, N.J. Gas Space and Oxygen Exchange in Roots of Avicennia Marina (Forssk.) Vierh. Var. Australasica (Walp.) Moldenke Ex NC Duke, the Grey Mangrove. Wetl. Ecol. Manag. 2001, 9, 221–228. [Google Scholar] [CrossRef]
- Augustinus, P.G.E.F. The Changing Shoreline of Suriname (South America); Utrecht University: Utrecht, The Netherlands, 1978; Volume 95. [Google Scholar]
- Jimenez, J.A.; Lugo, A.E.; Cintron, G. Tree Mortality in Mangrove Forests. Biotropica 1985, 177–185. [Google Scholar] [CrossRef]
- Breen, C.M.; Hill, B.J. A Mass Mortality of Mangroves in the Kosi Estuary. Trans. R. Soc. S. Afr. 1969, 38, 285–303. [Google Scholar] [CrossRef]
- Dicks, B. Oil and the Black Mangrove, Avicennia Marina in the Northern Red Sea. Mar. Pollut. Bull. 1986, 17, 500–503. [Google Scholar] [CrossRef]
- Saifullah, S.M.; Elahi, E. Pneumatophore Density and Size in Mangroves of Karachi, Pakistan. Pak. J. Bot. 1992, 24, 5. [Google Scholar]
- Dahdouh-Guebas, F.; Kairo, J.G.; De Bondt, R.; Koedam, N. Pneumatophore height and density in relation to micro-topography in the grey mangrove avicennia marina. Belg. J. Bot. 2007, 140, 213–221. [Google Scholar]
- Susilo, A.; Ridd, P.V. The Bulk Hydraulic Conductivity of Mangrove Soil Perforated with Animal Burrows. Wetl. Ecol. Manag. 2005, 13, 123–133. [Google Scholar] [CrossRef]
- Pülmanns, N.; Nordhaus, I.; Diele, K.; Mehlig, U. Artificial Crab Burrows Facilitate Desalting of Rooted Mangrove Sediment in a Microcosm Study. J. Mar. Sci. Eng. 2015, 3, 539–559. [Google Scholar] [CrossRef]
- Heron, S.F.; Ridd, P.V. The Use of Computational Fluid Dynamics in Predicting the Tidal Flushing of Animal Burrows. Estuar. Coast. Shelf Sci. 2001, 52, 411–421. [Google Scholar] [CrossRef]
- Susilo, A.; Ridd, P.V.; Thomas, S. Comparison between Tidally Driven Groundwater Flow and Flushing of Animal Burrows in Tropical Mangrove Swamps. Wetl. Ecol. Manag. 2005, 13, 377–388. [Google Scholar] [CrossRef]
- Smith, N.F.; Wilcox, C.; Lessmann, J.M. Fiddler Crab Burrowing Affects Growth and Production of the White Mangrove (Laguncularia Racemosa) in a Restored Florida Coastal Marsh. Mar. Biol. 2009, 156, 2255–2266. [Google Scholar] [CrossRef]
- Al-Khayat, J.A.; Jones, D.A. A Comparison of the Macrofauna of Natural and Replanted Mangroves in Qatar. Estuar. Coast. Shelf Sci. 1999, 49, 55–63. [Google Scholar] [CrossRef]
- Chang, H.; Han, S.H.; Kim, S.; An, J.; Alatalo, J.M.; Son, Y. Interactions between Topsoil Properties and Ecophysiological Responses of Mangroves (Avicennia Marina) along the Tidal Gradient in an Arid Region in Qatar. Turk. J. Agric. For. 2019, 43, 121–126. [Google Scholar]
- Buchanan, J.B.; Kain, J.M. Measurement of the physical and chemical environment. In Methods for the Study of Marine Benthos; Blackwell Scientific Publications Oxford: Oxford, UK, 1971; Volume 16, pp. 30–58. [Google Scholar]
- Dahdouh-Guebas, F.; De Bondt, R.; Abeysinghe, P.D.; Kairo, J.G.; Cannicci, S.; Triest, L.; Koedam, N. Comparative Study of the Disjunct Zonation Pattern of the Grey Mangrove Avicennia Marina (Forsk.) Vierh. in Gazi Bay (Kenya). Bull. Mar. Sci. 2004, 74, 237–252. [Google Scholar]
- Macia, A.; Abrantes, K.G.S.; Paula, J. Thorn Fish Terapon Jarbua (Forskal) Predation on Juvenile White Shrimp Penaeus Indicus, H. Milne Edwards and Brown Shrimp Metapenaeus Monoceros (Fabricius): The Effect of Turbidity, Prey Density, Substrate Type and Pneumatophore Density. J. Exp. Mar. Biol. Ecol. 2003, 291, 29–56. [Google Scholar] [CrossRef]
- Toma, T.; Nkamura, K.; Patanaponpaiboon, P.; Ogino, K. Effect of Flooding Water Level and Plant Density on Growth of Pneumatophore of Avicennia Marina. Tropics 1991, 1, 75–82. [Google Scholar] [CrossRef]
- Susilo, A. A New Method in Determining Bulk Hydraulic Conductivity of Mangrove Forest Sediment. Wit Trans. Ecol. Environ. 2005, 83, 10. [Google Scholar]
- Tomlinson, P.B. The Botany of Mangroves; Cambridge University Press: Cambridge, UK, 2016. [Google Scholar]
- Nguyen, H.T.; Stanton, D.E.; Schmitz, N.; Farquhar, G.D.; Ball, M.C. Growth Responses of the Mangrove Avicennia Marina to Salinity: Development and Function of Shoot Hydraulic Systems Require Saline Conditions. Ann. Bot. 2015, 115, 397–407. [Google Scholar] [CrossRef]
- Al-Khayat, J.A.; Abdulla, M.A.; Alatalo, J.M. Diversity of Benthic Macrofauna and Physical Parameters of Sediments in Natural Mangroves and in Afforested Mangroves Three Decades after Compensatory Planting. Aquat. Sci. 2019, 81, 4. [Google Scholar] [CrossRef]
- Tamin, N.M.; Zakaria, R.; Hashim, R.; Yin, Y. Establishment of Avicennia Marina Mangroves on Accreting Coastline at Sungai Haji Dorani, Selangor, Malaysia. Estuar. Coast. Shelf Sci. 2011, 94, 334–342. [Google Scholar] [CrossRef]
- MacNae, W. A General Account of the Fauna and Flora of Mangrove Swamps and Forests in the Indo-West-Pacific Region. Adv. Mar. Biol. 1969, 6, 73–270. [Google Scholar]
- Stokes, D.J.; Healy, T.R.; Cooke, P.J. Expansion Dynamics of Monospecific, Temperate Mangroves and Sedimentation in Two Embayments of a Barrier-Enclosed Lagoon, Tauranga Harbour, New Zealand. J. Coast. Res. 2010, 26, 113–122. [Google Scholar] [CrossRef]
- Mandura, A.S.; Khafaji, A.K.; Saifullah, S.M. Ecology of a Mangrove Stand of a Central Red Sea Coast Area: Ras Hatiba (Saudi Arabia). Proc. Saudi Biol. Soc. 1988, 11, 85–112. [Google Scholar]
- Park, S.G. Aspects of Mangrove Distribution and Abundance in Tauranga Harbour; Environment Bay of Plenty: Huakatani, New Zealand, 2004. [Google Scholar]
- Zhou, X.; Cai, L.; Fu, S. Comparison of Meiofaunal Abundance in Two Mangrove Wetlands in Tong’an Bay, Xiamen, China. J. Ocean Univ. China 2015, 14, 816–822. [Google Scholar] [CrossRef]
- Chen, X.; Cai, L.; Zhou, X.; Rao, Y. Geographical Variation in Oligochaete Density and Biomass in Subtropical Mangrove Wetlands of China. J. Ocean Univ. China 2017, 16, 925–931. [Google Scholar] [CrossRef]
- Al-Khayat, J.A.; Giraldes, B.W. Burrowing Crabs in Arid Mangrove Forests on the Southwestern Arabian Gulf: Ecological and Biogeographical Considerations. Reg. Stud. Mar. Sci. 2020, 39, 101416. [Google Scholar] [CrossRef]
- Stieglitz, T.; Ridd, P.; Müller, P. Passive Irrigation and Functional Morphology of Crustacean Burrows in a Tropical Mangrove Swamp. Hydrobiologia 2000, 421, 69–76. [Google Scholar] [CrossRef]
- Dunn, R.J.K.; Welsh, D.T.; Teasdale, P.R.; Gilbert, F.; Poggiale, J.-C.; Waltham, N.J. Effects of the Bioturbating Marine Yabby Trypaea Australiensis on Sediment Properties in Sandy Sediments Receiving Mangrove Leaf Litter. J. Mar. Sci. Eng. 2019, 7, 426. [Google Scholar] [CrossRef]
- Crona, B.I.; Holmgren, S.; Rönnbäck, P. Re-Establishment of Epibiotic Communities in Reforested Mangroves of Gazi Bay, Kenya. Wetl. Ecol Manag. 2006, 14, 527–538. [Google Scholar] [CrossRef]
- Young, B.M.; Harvey, E.L. A Spatial Analysis of the Relationship Between Mangrove (Avicennia Marinavar.Australasica) Physiognomy and Sediment Accretion in the Hauraki Plains, New Zealand. Estuar. Coast. Shelf Sci. 1996, 42, 231–246. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elevation Location/Hydrochemical Data | Upper Mangroves | Middle Mangroves | ||
|---|---|---|---|---|
| Tree | Seedling | Tree | Seedling | |
| Natural Mangroves East Coast Al Khor | ||||
| (a) Tree density no./ha | 2800 | 1100 | 3100 | 1100 |
| (b) Tree height range | 1–3 m | 5–60 cm | 1.5–6 m | 10–70 cm |
| (c) Pnematophore/m2 | 86 | - | 516 | - |
| Al Dhakhira | ||||
| (a) Tree density no./ha | 2800 | 1333 | 3000 | 1000 |
| (b) Tree height range | 1–3 m | 10–70 cm | 1.7–6 m | 10–80 cm |
| (c) Pnematophore/m2 | 158 | - | 308 | - |
| Planted Mangroves East Coast Semaisma | ||||
| (a) Tree density no./ha | 1100 | 933 | 1700 | 1000 |
| (b) Tree height range | 1.5–2.7 m | 10–40 cm | 1–2.5 m | 10–80 cm |
| (c) Pnematophore/m2 | 124 | - | 272 | - |
| Fuwairit | ||||
| (a) Tree density no./ha | 1600 | 1270 | 1900 | 1233 |
| (b) Tree height range | 1.5–3 m | 5–30 cm | 1.4–4.5 m | 5–60 cm |
| (c) Pnematophore/m2 | 308 | - | 194 | - |
| Al Mafyar | ||||
| (a) Tree density no./ha | 2100 | 1466 | 2600 | 1100 |
| (b) Tree height range | 0.6–2 m | 5–40 cm | 1.5–4 m | 10–40 cm |
| (c) Pnematophore/m2 | 88 | - | 88 | - |
| West Coast Zekreet | ||||
| (a) Tree density no./ha | Nil | Nil | 200 | Nil |
| (b) Tree height range | Nil | Nil | 0.5–1.0 m | Nil |
| (c) Pnematophore/m2 | 14 | - | Nil | - |
| North East Coast Ras Laffan | ||||
| (a) Tree density no./ha | 1100 | 920 | 3100 | 1250 |
| (b) Tree height range | 1–3 m | 10–50 cm | 1.5–3 m | 10–70 cm |
| (c) Pnematophore/m2 | 68 | - | 90 | - |
| East South Al Wakra | ||||
| (a) Tree density no./ha | 1500 | 1040 | 2200 | 1050 |
| (b) Tree height range | 1–3 m | 5–45 cm | 1.0–6 m | 10–75 cm |
| (c) Pnematophore/m2 | 142 | 134 | ||
| Umm Al Hul | ||||
| (a) Tree density no./ha | 2300 | 1200 | 2500 | 1500 |
| (b) Tree height range | 1.5–2.7 m | 10–40 cm | 1.5–3 m | 10–80 cm |
| (c) Pnematophore/m2 | 90 | 104 | ||
| South Khor Al Adaid | ||||
| (a) Tree density no./ha | Nil | Nil | 15 | Nil |
| (b) Tree height range | Nil | Nil | 0.5–0.75 m | Nil |
| (c) Pnematophore/m2 | 20 | 25 | ||
| Site | Position on Shore | Pneumat. Abundance (m2) | Tree Height (m) | Pneumat. Height (cm) | Pneumat. Dia. (mm) | Girth Size (cm) | Mud Content % | Burrows no/m2 |
|---|---|---|---|---|---|---|---|---|
| Umm Al Hul | Upper | 90 | 1.5 | 16.82 | 9 | 12 | 18.25 | 10 |
| Semaisma | Upper | 124 | 1.75 | 13.38 | 10.5 | 16.75 | 26.97 | 12 |
| Al Wakra | Upper | 142 | 2 | 16.5 | 12.65 | 27.5 | 26.5 | 9 |
| Al Khor | Upper | 86 | 1.25 | 11.83 | 8 | 16 | 38.78 | 10 |
| Ras Laffan | Upper | 68 | 1.5 | 12.25 | 8.3 | 13 | 15.92 | 8 |
| Al Dhakhira | Upper | 158 | 1.8 | 23.43 | 10 | 23 | 65.23 | 11 |
| Al Mafyar | Upper | 88 | 1.75 | 7.63 | 6.5 | 13 | 16.54 | 4 |
| Fuwairit | Upper | 152 | 1.75 | 15.48 | 10 | 25 | 28.15 | 12 |
| Zekreet | Upper | 14 | 0.45 | 6.58 | 8 | 9 | 22.42 | 0 |
| KhorAl Adaid | Upper | 20 | 0.5 | 10.28 | 6.2 | 12 | 9.85 | 16 |
| Maximum | 158.00 | 2.00 | 23.43 | 12.65 | 27.50 | 65.23 | 16.00 | |
| Minimum | 14.00 | 0.45 | 6.58 | 6.20 | 9.00 | 9.85 | 0.00 | |
| Mean | 94.20 | 1.43 | 13.42 | 8.91 | 16.73 | 26.86 | 9.20 | |
| SD | 50.92 | 0.54 | 4.94 | 1.94 | 6.29 | 15.73 | 4.47 | |
| SE | 15.81 | 0.17 | 1.56 | 0.61 | 1.99 | 4.97 | 1.41 | |
| Umm Al Hul | Middle | 208 | 1.75 | 19.17 | 9.0 | 21 | 27.37 | 9 |
| Semaisma | Middle | 240 | 1.55 | 14.125 | 9.4 | 19.25 | 35.7 | 6 |
| Al Wakra | Middle | 268 | 1.7 | 15.64 | 9.2 | 21.5 | 31.03 | 22 |
| Al Khor | Middle | 324 | 2.07 | 14.9 | 9.2 | 28 | 56.44 | 8 |
| Ras Laffan | Middle | 180 | 1.55 | 15.8 | 9.0 | 12.4 | 27.46 | 4 |
| Al Dhakhira | Middle | 360 | 2 | 13.73 | 9.0 | 27 | 74.46 | 12 |
| Al Mafyar | Middle | 184 | 1.5 | 11.53 | 8.0 | 21 | 20.93 | 9 |
| Fuwairit | Middle | 196 | 1.5 | 9.15 | 7.0 | 12 | 27.88 | 14 |
| Zekreet | Middle | 62 | 0.4 | 8.1 | 7.0 | 7 | 28.43 | 0 |
| Khor Al Adaid | Middle | 50 | 0.5 | 12.64 | 6.9 | 11 | 9.23 | 14 |
| Maximum | 360.00 | 2.07 | 19.17 | 9.35 | 28.00 | 74.46 | 22.00 | |
| Minimum | 50.00 | 0.40 | 8.10 | 6.90 | 7.00 | 9.23 | 0.00 | |
| Mean | 207.20 | 1.45 | 13.48 | 8.37 | 18.02 | 33.89 | 9.80 | |
| SD | 99.42 | 0.56 | 3.28 | 1.03 | 7.07 | 18.53 | 6.12 | |
| SE | 31.44 | 0.18 | 1.37 | 0.33 | 2.24 | 5.86 | 1.94 | |
| Umm Al Hul | Lower | 224 | 2 | 20.79 | 7.6 | 23 | 29.87 | 8 |
| Semaisma | Lower | 272 | 1.5 | 27.5 | 8 | 18 | 36.33 | 3 |
| Al Wakra | Lower | 196 | 1.65 | 15.75 | 9.1 | 18 | 29.89 | 33 |
| Al Khor | Lower | 516 | 3 | 15.62 | 9.3 | 55 | 79.54 | 12 |
| Ras Laffan | Lower | 160 | 1.75 | 11.51 | 8.5 | 11 | 24.57 | 3 |
| Al Dhakhira | Lower | 308 | 2 | 14.52 | 9 | 24 | 79.76 | 18 |
| Al Mafyar | Lower | 132 | 1 | 10.52 | 8 | 14 | 17.91 | 10 |
| Fuwairit | Lower | 194 | 1.5 | 12.55 | 8 | 14 | 28.31 | 8 |
| Zekreet | Lower | 30 | 0.35 | 7.68 | 7 | 8 | 22.21 | 0 |
| Khor Al Adaid | Lower | 108 | 0.75 | 18 | 8.5 | 14 | 14.2 | 18 |
| Maximum | 516.00 | 3.00 | 27.50 | 9.30 | 55.00 | 79.76 | 33.00 | |
| Minimum | 30.00 | 0.35 | 7.68 | 7.00 | 8.00 | 14.20 | 0.00 | |
| Mean | 214.00 | 1.55 | 15.44 | 8.30 | 19.90 | 36.26 | 11.30 | |
| SD | 132.90 | 0.74 | 5.67 | 0.72 | 13.30 | 23.73 | 9.72 | |
| SE | 41.74 | 0.23 | 1.79 | 0.23 | 4.21 | 7.50 | 3.07 | |
| Total mangrove | Max. | 258 | 3 | 27.5 | 1.265 | 55 | 79.76 | 33 |
| Min. | 14 | 0.35 | 6.58 | 0.62 | 7 | 9.23 | 0 | |
| Mean | 101.67 | 1.48 | 14.11 | 0.85 | 18.21 | 32.34 | 10.10 | |
| SD | 54.47 | 0.60 | 4.67 | 0.13 | 9.19 | 19.35 | 6.92 | |
| SE | 9.95 | 0.11 | 0.85 | 0.02 | 1.68 | 3.53 | 1.26 |
| Upper Tidal L. | Middle Tidal L. | Lower Tidal L. | All Tidal L. | |||||
|---|---|---|---|---|---|---|---|---|
| P. ab. | p-Value | P. ab. | p-Value | P. ab. | p-Value | P. ab. | p-Value | |
| T.Ht | 0.786 | 0.000 | 0.866 | 0.000 | 0.824 | 0.000 | 0.822 | 0.000 |
| P.Ht | 0.591 | 0.000 | 0.195 | 0.000 | 0.147 | 0.002 | 0.262 | 0.000 |
| P.Dia | 0.318 | 0.000 | 0.264 | 0.000 | 0.160 | 0.002 | 0.193 | 0.000 |
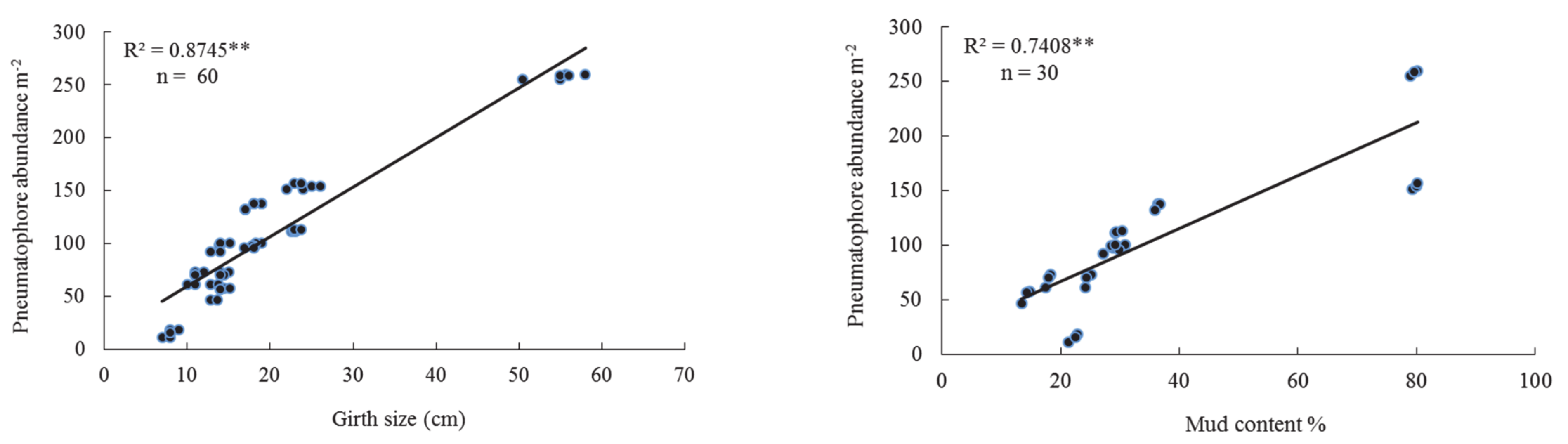
| G.S | 0.770 | 0.000 | 0.792 | 0.000 | 0.874 | 0.000 | 0.800 | 0.000 |
| Md.Ct | 0.388 | 0.000 | 0.704 | 0.000 | 0.741 | 0.000 | 0.629 | 0.000 |
| Br.No | 0.091 | 0.105 | 0.088 | 0.112 | 0.031 | 0.351 | 0.055 | 0.026 |
| Site | Tidal Level | Family | Species | Mean ± SE | Range |
|---|---|---|---|---|---|
| Natural mangroves | Upper | Dotillidae Camptandriidae | Scopimera carbricauda (Alcock, 1900) Nasima dotilliformis (Alcock, 1900) | 10.5 ± 0.5 | 10–11 |
| Middle | Camptandriidae Macrophthalmidae | Manningis arabicum (Jones and Clayton, 1983) Macrophthalmus depressus (Rǔppell, 1830) | 10.0 ± 2.0 | 8–12 | |
| Lower | Macrophthalmidae | Macrophthalmus depressus (Rǔppell, 1830) | 15.0 ± 3.0 | 12–18 | |
| Planted mangroves | Upper | Dotillidae | Scopimera carbricauda (Alcock, 1900) | 8.88 ± 1.8 | 0–16 |
| Middle | Dotillidae Macrophthalmidae | Scopimera carbricauda (Alcock, 1900) Macrophthalmus depressus (Rǔppell, 1830) | 9.75 ± 2.4 | 0–22 | |
| Lower | Macrophthalmidae | Macrophthalmus depressus (Rǔppell, 1830) | 10.4 ± 3.5 | 0–33 |
| Source | Type III Sum of Squares | df | Mean Square | F | p-Value |
|---|---|---|---|---|---|
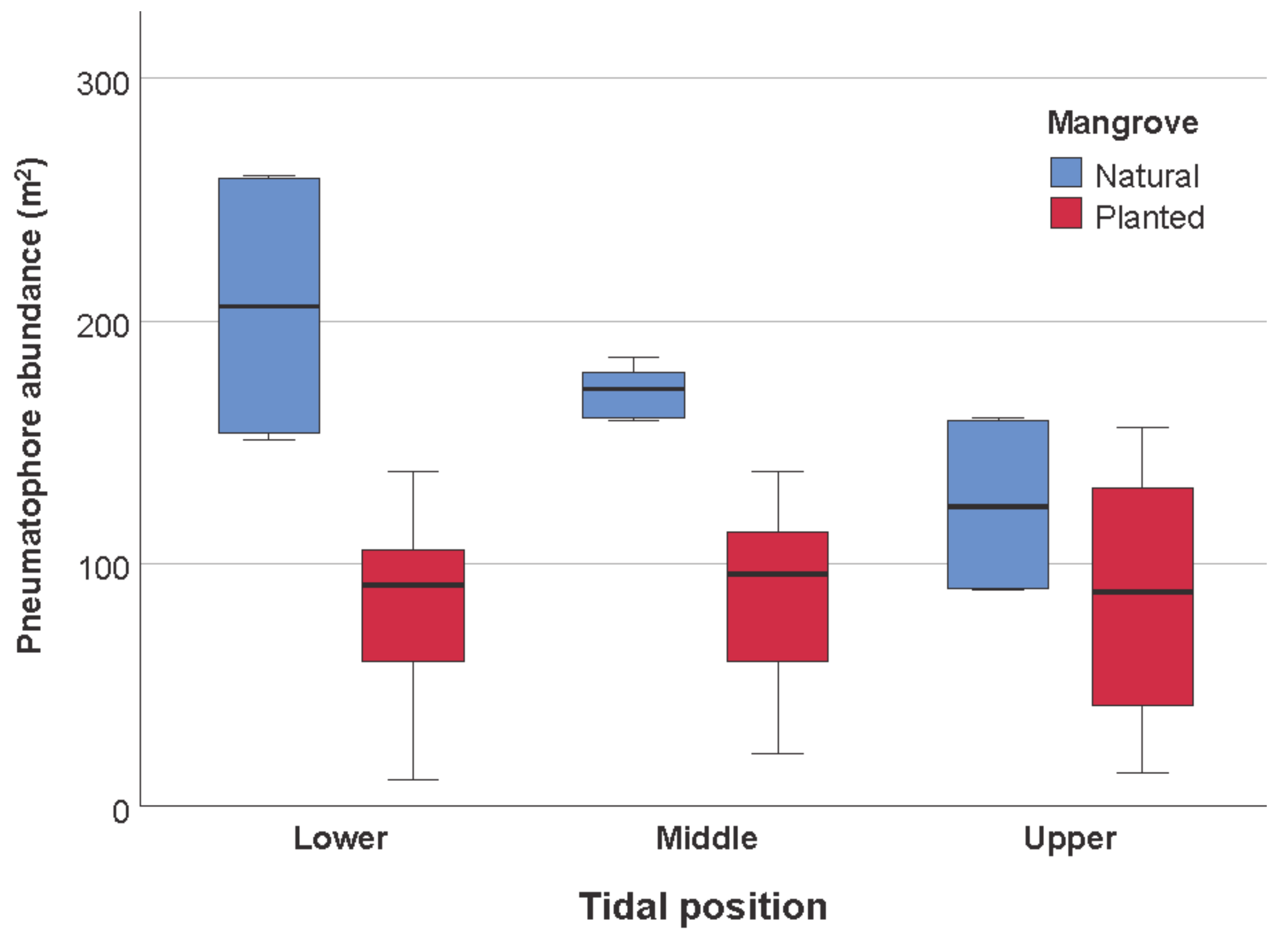
| Mangrove | 95,811.469 | 1 | 95,811.469 | 56.551 | 0.000 |
| Tidal position | 14,413.706 | 2 | 7206.853 | 4.254 | 0.017 |
| Mangrove × Tidal position | 18,102.372 | 2 | 9051.186 | 5.342 | 0.007 |
| Total | 1,192,413.000 | 90 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Khayat, J.A.; Alatalo, J.M. Relationship Between Tree Size, Sediment Mud Content, Oxygen Levels, and Pneumatophore Abundance in the Mangrove Tree Species Avicennia Marina (Forssk.) Vierh. J. Mar. Sci. Eng. 2021, 9, 100. https://doi.org/10.3390/jmse9010100
Al-Khayat JA, Alatalo JM. Relationship Between Tree Size, Sediment Mud Content, Oxygen Levels, and Pneumatophore Abundance in the Mangrove Tree Species Avicennia Marina (Forssk.) Vierh. Journal of Marine Science and Engineering. 2021; 9(1):100. https://doi.org/10.3390/jmse9010100
Chicago/Turabian StyleAl-Khayat, Jassim A., and Juha M. Alatalo. 2021. "Relationship Between Tree Size, Sediment Mud Content, Oxygen Levels, and Pneumatophore Abundance in the Mangrove Tree Species Avicennia Marina (Forssk.) Vierh" Journal of Marine Science and Engineering 9, no. 1: 100. https://doi.org/10.3390/jmse9010100
APA StyleAl-Khayat, J. A., & Alatalo, J. M. (2021). Relationship Between Tree Size, Sediment Mud Content, Oxygen Levels, and Pneumatophore Abundance in the Mangrove Tree Species Avicennia Marina (Forssk.) Vierh. Journal of Marine Science and Engineering, 9(1), 100. https://doi.org/10.3390/jmse9010100