Severe Coastal Hypoxia Interchange with Ocean Acidification: An Experimental Perturbation Study on Carbon and Nutrient Biogeochemistry

 ,
,  ,
,
Abstract
1. Introduction
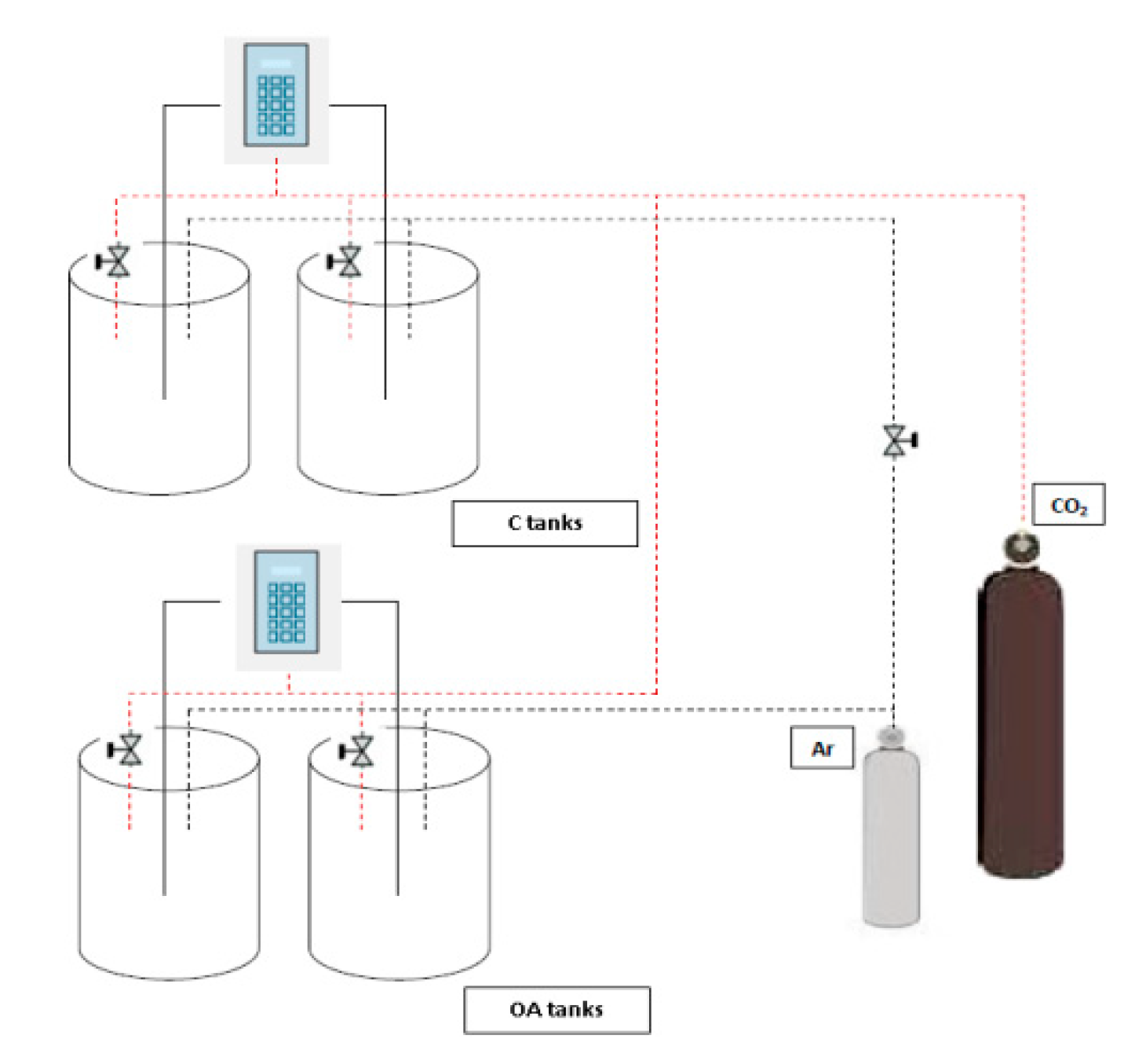
2. Materials and Methods
3. Results
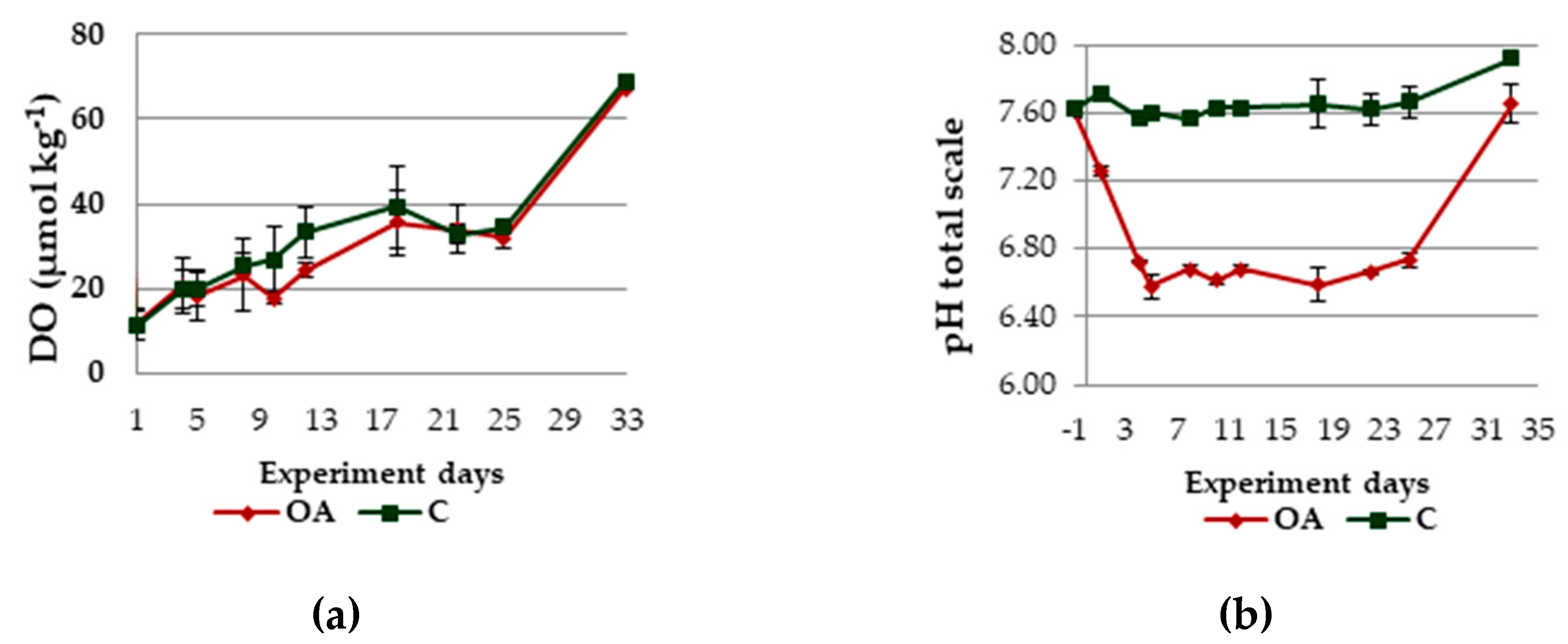
3.1. Physicochemical Parameters
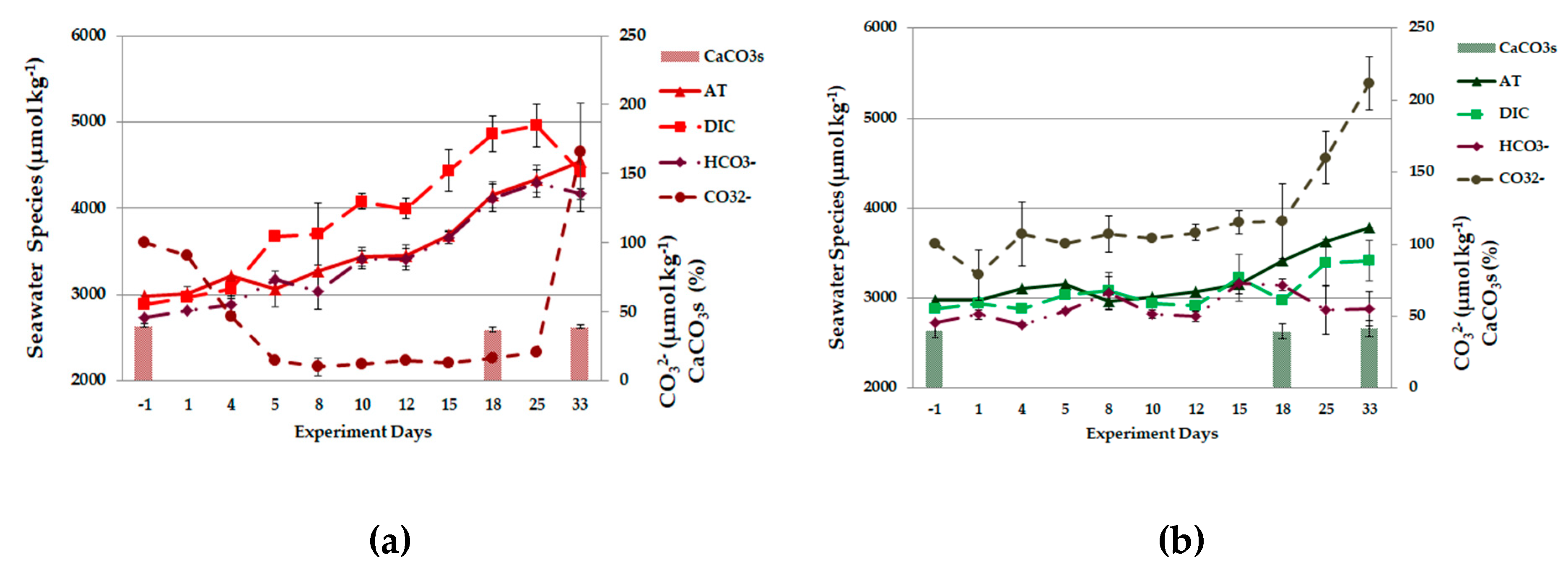
3.2. Carbonate System Parameters
3.2.1. Field Observations
3.2.2. Experiment Observations
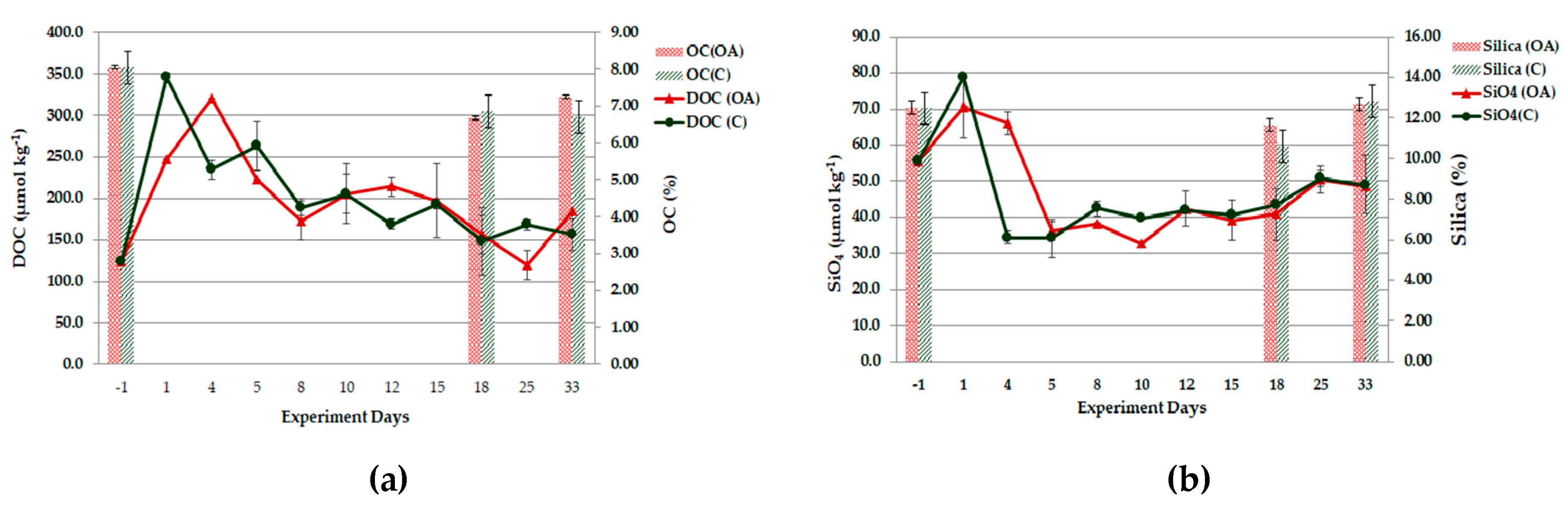
3.3. Nutrient Species and Carbon Analyses
3.3.1. Field Observations
3.3.2. Experiment Observations
3.4. Sediment Analyses
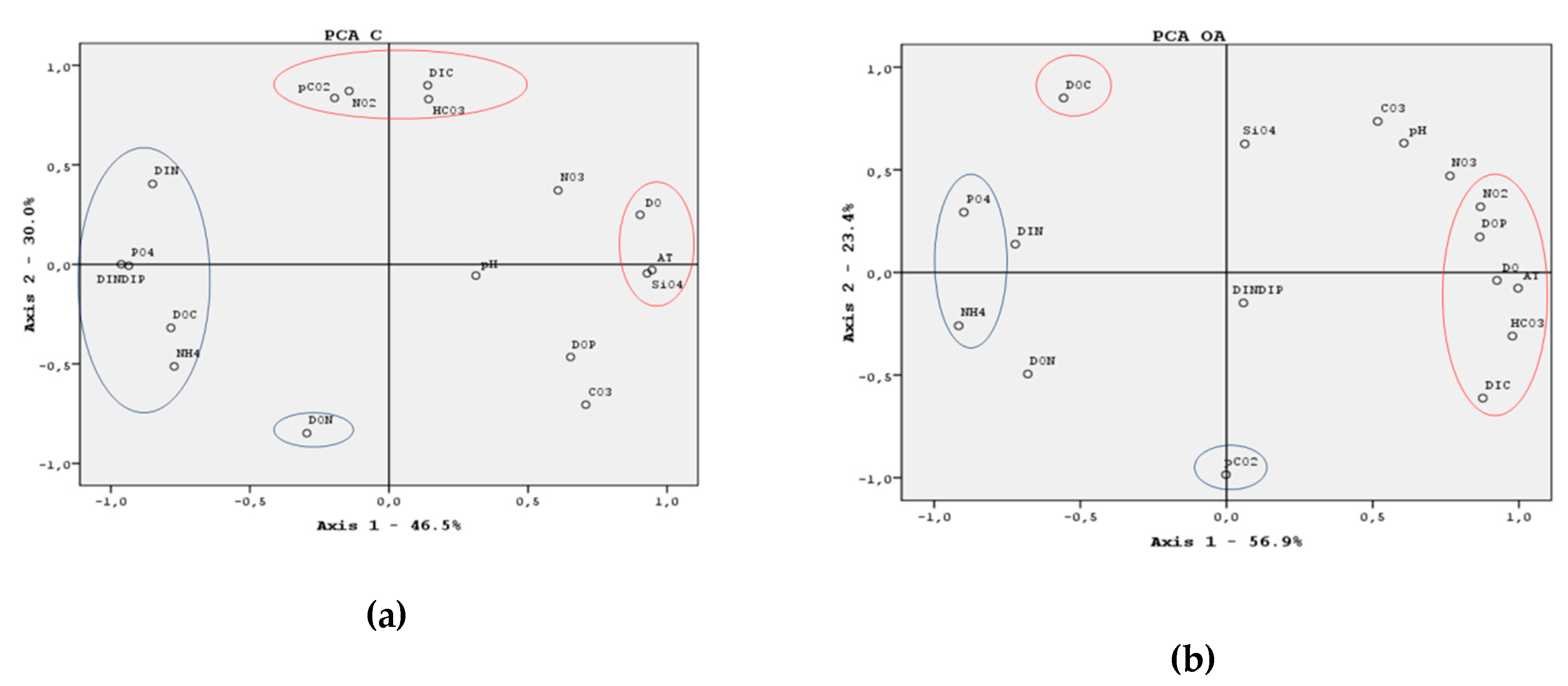
3.5. ANOVA Test and Principal Components Analysis (PCA)
4. Discussion
4.1. Carbonate System and Related Processes
4.2. Nutrients and Carbon—Evidence of Nitrification under Low DO
4.3. Implications and Interchanges of Intermittent Hypoxia/Anoxia and Coastal Acidification
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Diaz, R.; Rosenberg, R. Spreading Dead Zones and Consequences for Marine Ecosystems. Science 2008, 321, 926–929. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, A.; Peltzer, E.; Walz, P.; Brewer, P. Hypoxia by degrees:Establishing definitions for a changing ocean. Deep Sea Res. Part I Oceanogr. Res. Pap. 2011, 58, 1212–1226. [Google Scholar] [CrossRef]
- Ingrosso, G.; Giani, M.; Cibic, T.; Karuza, A.; Krajl, M.; Del Negro, P. Carbonate chemistry dynamics and biological processes along a river–sea gradient (Gulf of Trieste, northern Adriatic Sea). J. Mar. Syst. 2016, 155, 35–49. [Google Scholar] [CrossRef]
- Liu, J.; Weinbauer, M.G.; Maier, C.; Dai, M.; Gattuso, J.-P. Effect of ocean acidification on microbial diversity and on microbe-driven biogeoschemistry and ecosystem functioning. Aquat. Microb. Ecol. 2010, 61, 291–305. [Google Scholar] [CrossRef]
- Chan, F.; Boehm, A.B.; Barth, J.A.; Chornesky, E.A.; Dickson, A.G.; Feely, R.A.; Hales, B.; Hill, T.M.; Hofmann, G.; Ianson, D.; et al. The West Coast Ocean Acidification and Hypoxia Science Panel: Major Findings, Recommendations, and Actions; California Ocean Science Trust: Oakland, CA, USA, 2016; p. 40. [Google Scholar]
- Justic, D.; Rabalais, N.N.; Turner, R.E. Effects of climate change on hypoxia in coastal waters: A doubled CO2 scenario for the northern Gulf of Mexico. Limnol. Oceanogr. 1996, 41, 992–1003. [Google Scholar] [CrossRef]
- Cantoni, C.; Luchetta, A.; Celio, M.; Cozzi, S.; Raicich, F. Carbonate system variability in the Gulf of Trieste (North Adriatic Sea). Estuar. Coast. Shelf Sci. 2012, 115, 51–62. [Google Scholar] [CrossRef]
- Gobler, C.J.; DePasquale, E.L.; Griffith, A.W.; Baumann, H. Hypoxia and Acidification Have Additive and Synergistic Negative Effects on the Growth, Survival, and Metamorphosis of Early Life Stage Bivalves. PLoS ONE 2014, 9, e83648. [Google Scholar] [CrossRef]
- Melzner, F.; Thomsen, J.; Koeve, W.; Oschlies, A.; Gutowska, M.A.; Bange, H.W.; Hansen, H.P.; Kortzinger, A. Future ocean acidification will be amplified by hypoxia in coastal habitats. Mar. Biol. 2013, 160, 1875–1888. [Google Scholar] [CrossRef]
- Wallace, R.B.; Baumann, H.; Grear, J.S.; Aller, R.C.; Gobler, C.J. Coastal ocean acidification: The other eutrophication problem. Estuar. Coast. Shelf Sci. 2014, 148, 1–13. [Google Scholar] [CrossRef]
- Hofmann, G.E.; Smith, J.E.; Johnson, K.S.; Send, U.; Levin, L.A.; Micheli, F.; Paytan, A.; Price, N.N.; Peterson, B.; Takeshita, Y.; et al. High frequency dynamics of ocean pH: A multi-ecosystem comparison. PLoS ONE 2011, 6, e28983. [Google Scholar] [CrossRef]
- Waldbusser, G.G.; Salisbury, J.E. Ocean Acidification in the Coastal Zone from an Organism’s Perspective: Multiple System Parameters, Frequency Domains, and Habitats. Annu. Rev. Mar. Sci. 2014, 6, 221–247. [Google Scholar] [CrossRef] [PubMed]
- Duarte, C.M.; Hendriks, I.E.; Moore, T.S.; Olsen, Y.S.; Steckbauer, A.; Ramajo, L.; Carstensen, J.; Trotter, J.A.; McCulloch, M. Is Ocean Acidification an Open-Ocean Syndrome? Understanding Anthropogenic Impacts on Seawater pH. Estuaries Coast. 2013, 36, 221–236. [Google Scholar] [CrossRef]
- Kapsenberg, L.; Alliouane, S.; Gazeau, F.; Mousseau, L.; Gattuso, J.-P. Coastal ocean acidification and increasing total alkalinity in the northwestern Mediterranean Sea. Ocean Sci. 2017, 13, 411–426. [Google Scholar] [CrossRef]
- Cai, W.; Hu, X.; Huang, W.; Murrel, M.; Lehrter, J.; Lohrenz, S.; Chou, W.; Zhai, W.; Hollibaugh, J.; Wang, Y.; et al. Acidification of subsurface coastal waters enhanced by eutrophication. Nat. Geosci. 2011, 4, 766–770. [Google Scholar] [CrossRef]
- Carstensen, J.; Duarte, C.M. Drivers of pH Variability in Coastal Ecosystems. Environ. Sci. Technol. 2019, 53, 4020–4029. [Google Scholar] [CrossRef]
- Provoost, P.; Van Heuven, S.; Soetaert, K.; Laane, R.W.P.M.; Middelburg, J.J. Seasonal and long-term changes in pH in the Dutch coastal zone. Biogeosciences 2010, 7, 3869–3878. [Google Scholar] [CrossRef]
- Borges, A.V.; Gypens, N. Carbonate chemistry in the coastal zone responds more strongly to eutrophication than to ocean acidification. Limnol. Oceanogr. 2010, 55, 346–353. [Google Scholar] [CrossRef]
- Paraskevopoulou, V.; Zeri, C.; Kaberi, H.; Chalkiadaki, O.; Krasakopoulou, E.; Dassenakis, M.; Scoullos, M. Trace metal variability, background levels and pollution status assessment in line with the water framework and Marine Strategy Framework EU Directives in the waters of a heavily impacted Mediterranean Gulf. Mar. Pollut. Bull. 2014, 87, 323–337. [Google Scholar] [CrossRef]
- Pavlidou, A.; Kontoyiannis, H.; Anagnostou, C.; Siokou-Frangou, I.; Pagou, K.; Krasakopoulou, E.; Assimakopoulou, G.; Zervoudaki, S.; Zeri, C.; Chatzianestis, J.; et al. Biogeochemical Characteristics in the Elefsis Bay (Aegean Sea, Eastern Mediterranean) in Relation to Anoxia and Climate Changes. In Chemical Structure of Pelagic Redox Interfaces. The Handbook of Environmental Chemistry; Yakushev, E., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; Volume 22, pp. 161–201. [Google Scholar] [CrossRef]
- Ignatiades, L. Scaling the trophic status of the Aegean Sea, eastern Mediterranean. J. Sea Res. 2005, 54, 51–57. [Google Scholar] [CrossRef]
- Scoullos, M.; Rilley, J. Water circulation in the Gulf of Elefsis, Greece. Thalassia Jugosl. 1978, 14, 357–370. [Google Scholar]
- Scoullos, M. Chemical Studies of the Gulf of Elefsis, Greece. Ph.D. Thesis, Depterment of Oceanography, University of Liverpool, Liverpool, UK, 1979; p. 328. [Google Scholar]
- Scoullos, M.; Zeri, C.; Dassenakis, M.; Tsamaki, E. Experiments on the reversibility of transfer—Deposition processes of metal under intermittently anoxic conditions. Rapp. Comm. Int. Mer. Medit. 1986, 30, 122. [Google Scholar]
- Primpas, I.; Tsirtsis, G.; Karydis, M.; Kokkoris, G.D. Principal component analysis: Development of a multivariate index for assessing eutrophication according to the European water framework directive. Ecol. Indic. 2010, 10, 178–183. [Google Scholar] [CrossRef]
- Pavlidou, A.; Pagou, K.; Assimakopoulou, G.; Rousselaki, E. Evolution over the last 30 years of the trophic conditions in the Gulf of Elefsis. In Sustainable Mediterranean, Issue No 71. In Proceedings of the International Conference: Environmental Perspectives of the Gulf of Elefsis, A Mediterranean Case Study Where Science Meets the Society, Elefsis, Greece, 11–12 September 2015; pp. 20–22. [Google Scholar]
- Friligos, N. Some consequences of the decomposition of organic matter in the Elefsis bay, an anoxic basin. Mar. Pollut. Bull. 1982, 13, 103–106. [Google Scholar] [CrossRef]
- Kapetanaki, N.; Krasakopoulou, E.; Stathopoulou, E.; Pavlidou, A.; Zervoudaki, S.; Dassenakis, M.; Scoullos, M. Simulation of Coastal Processes affecting pH with Impacts on Carbon and Nutrient Biogeochemistry. Mediterr. Mar. Sci. 2018, 19, 290–304. [Google Scholar] [CrossRef]
- Dickson, A.G.; Sabine, C.L.; Christian, J.R. (Eds.) Guide to Best Practices for Ocean CO2 Measurements; PICES Special Publication 3: Darby, PA, USA, 2007; p. 191. [Google Scholar]
- Caldeira, K.; Wickett, M. Ocean Model predictions of chemistry changes from carbon dioxide emissions to the atmosphere and ocean. J. Geophys. Res. Oceans 2005, 110, 1–12. [Google Scholar] [CrossRef]
- Pierrot, D.; Lewis, E.; Wallace, D.W.R. MS Excel Program Developed for CO2 System Calculations. ORNL/CDIAC-105a. In Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory; U.S. Department of Energy: Oak Ridge, TN, USA, 2006. [Google Scholar] [CrossRef]
- Beman, M.J.; Chow, C.-E.; King, A.L.; Feng, Y.; Fuhrman, J.A.; Andersson, A.; Bates, N.R.; Popp, B.N.; Hutchins, D.A. Global declines in oceanic nitrification rates as a consequence of ocean acidification. Proc. Natl. Acad. Sci. USA 2011, 108, 208–213. [Google Scholar] [CrossRef]
- Lueker, T.J.; Dickson, A.G.; Keeling, C.D. Ocean pCO2 calculated from dissolved inorganic carbon, alkalinity, and equations for K1 and K2: Validation based on laboratory measurements of CO2 in gas and seawater at equilibrium. Mar. Chem. 2000, 70, 105–119. [Google Scholar] [CrossRef]
- Perez, F.F.; Fraga, F. Association constant of fluoride and hydrogen ions in seawater. Mar. Chem. 1987, 21, 161–168. [Google Scholar] [CrossRef]
- Dickson, A.G. Standard potential of the reaction: AgCI(s) + 1/2H2(g) = Ag(s) + HCI(aq), and the standard acidity constant of the ion HSO4− in synthetic sea water from 273.15 to 318.15 K. J. Chem. Thermodyn. 1990, 22, 113–127. [Google Scholar] [CrossRef]
- Lee, K.; Tae-Wook, K.; Byrne, R.H.; Millero, F.J.; Feely, R.A.; Liu, Y.-M. The universal ratio of the boron to chlorinity for the North Pacific and North Atlantic oceans. Geochim. Cosmochim. Acta 2010, 74, 1801–1811. [Google Scholar] [CrossRef]
- Karageorgis, A.P.; Katsanevakis, S.; Kaberi, H. Use of enrichment factors for the assessment of heavy metal contamination in the sediments of Koumoundourou Lake, Greece. Water Air Soil Pollut. 2009, 204, 243–258. [Google Scholar] [CrossRef]
- Riebesell, U.; Fabry, V.J.; Hansson, L.; Gattuso, J.-P. Guide to Best Practices for Ocean Acidification Research and Data Reporting; Publications Office of the European Union: Luxembourg, 2010; p. 259. [Google Scholar]
- Frew, N.M. The role of organic films in air-sea gas exchange. In The Sea Surface and Global Change; Liss, P.S., Duce, R.A., Eds.; Cambridge University Press: Cambridge, UK, 1997; pp. 121–172. [Google Scholar]
- Dickson, A.G. An exact definition of total alkalinity and a procedure for the estimation of alkalinity and total inorganic carbon from titration data. Deep Sea Res. Part A Oceanogr. Res. Pap. 1981, 28, 609–623. [Google Scholar] [CrossRef]
- Andersson, A.J.; Bates, N.R.; Mackenzie, F.T. Dissolution of Carbonate Sediments under Rising pCO2 and Ocean Acidification: Observations from Devil’s Hole, Bermuda. Aquat. Geochem. 2007, 13, 237–264. [Google Scholar] [CrossRef]
- Krumins, V.; Gehlen, M.; Arndt, S.; Van Cappellen, P.; Regnier, P. Dissolved inorganic carbon and alkalinity fluxes from coastal marine sediments: Model estimates for different shelf environments and sensitivity to global change. Biogeosciences 2013, 10, 371–398. [Google Scholar] [CrossRef]
- Chen, C.-T.A. Shelf-vs. dissolution-generated alkalinity above the chemical lysocline. Deep Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 5365–5375. [Google Scholar] [CrossRef]
- Chen, C.-T.A.; Liu, K.K.; Macdonald, R. Continental Margin Exchanges. In Ocean Biogeochemistry. Global Change—The IGBP Series; Fasham, M.J.R., Ed.; Springer: Berlin/Heidelberg, Germany, 2003; pp. 53–97. [Google Scholar] [CrossRef]
- Thomas, H.; Schiettecatte, L.-S.; Suykens, K.; Kone, Y.J.M.; Shadwick, E.H.; Prowe, A.E.E.; Bozec, Y.; de Baar, H.J.W.; Borges, A.V. Enhanced ocean carbon storage from anaerobic alkalinity generation in coastal sediments. Biogeosciences 2009, 6, 267–274. [Google Scholar] [CrossRef]
- Hu, X.; Cai, W.J. An assessment of ocean margin anaerobic processes on oceanic alkalinity budget. Glob. Biogeochem. Cycles 2011, 25, GB3003. [Google Scholar] [CrossRef]
- Hulth, S.; Aller, R.C.; Gilbert, F. Coupled anoxic nitrification/manganese reduction in marine sediments. Geochim. Cosmochim. Acta 1999, 63, 49–66. [Google Scholar] [CrossRef]
- Huesemann, M.H.; Skillman, A.D.; Crecelius, E.A. The inhibition of marine nitrification by ocean disposal of carbon dioxide. Mar. Pollut. Bull. 2002, 44, 142–148. [Google Scholar] [CrossRef]
- Widdicombe, S.; Dashfield, S.L.; McNeil, C.L.; Needham, H.R.; Beesly, A.; McEvoy, A.; Oxnevad, S.; Clarke, K.R.; Berge, J.A. Effects of CO2 induced seawater acidification on infaunal diversity and sediment nutrient fluxes. Mar. Ecol. Prog. Ser. 2009, 379, 59–75. [Google Scholar] [CrossRef]
- Scoullos, M. Trace Metals in a landlocked intermittently anoxic basin. In Trace Metals in Sea Water; Wong, C.S., Boyle, E., Bruland, K.W., Burton, J.D., Goldberg, E.D., Eds.; Plenum Press: New York, NY, USA, 1983; pp. 351–366. [Google Scholar]
- Kapetanaki, N.; Stathopoulou, E.; Karavoltsos, S.; Katsouras, G.; Krasakopoulou, E.; Dassenakis, E.; Scoullos, M. Biogeochemical processes under anoxic conditions affecting Arsenic (As) distribution in Elefsis Gulf: Possible alterations due to simulated ocean acidification conditions—A preliminary assessment. In Sustainable Mediterranean, Issue No 71. In Proceedings of the International Conference: Environmental Perspectives of the Gulf of Elefsis, A Mediterranean Case Study Where SCIENCE Meets the Society, Elefsis, Greece, 11–12 September 2015; pp. 74–78. [Google Scholar]
- Bartlett, R.; Mortimer, R.J.G.; Morris, K. Anoxic nitrification: Evidence from Humber Estuary sediments (UK). Chem. Geol. 2008, 250, 29–39. [Google Scholar] [CrossRef]
- Louis, J.; Guieu, C.; Gazeau, F. Nutrient dynamics under different ocean acidification scenarios in a low nutrient low chlorophyll system: The Northwestern Mediterranean Sea. Estuar. Coast. Shelf Sci. 2016, 186, 30–44. [Google Scholar] [CrossRef]
- Hutchins, D.A.; Mulholland, M.R.; Fu, F. Nutrient Cycles and Marine Microbes in a CO₂-Enriched Ocean. Oceanography 2009, 22, 128–145. [Google Scholar] [CrossRef]
- Tanaka, T.; Thingstad, T.F.; Løvdal, T.; Grossart, H.-P.; Larsen, A.; Allgaier, M.; Meyerhöfer, M.; Schulz, K.G.; Wohlers, J.; Zöllner, E.; et al. Availability of phosphate for phytoplankton and bacteria and of glucose for bacteria at different pCO2 levels in a mesocosm study. Biogeosciences 2008, 5, 669–678. [Google Scholar] [CrossRef]
- Gunnars, A.; Blomqvist, S.; Johansson, P.; Andersson, C. Formation of Fe(III) oxyhydroxide colloids in freshwater and brackish seawater, with incorporation of phosphate and calcium. Geochim. Cosmochim. Acta 2002, 66, 745–758. [Google Scholar] [CrossRef]
- Skoog, A.C.; Arias-Esquivel, V.A. The effect of induced anoxia and reoxygenation on benthic fluxes of organic carbon, phosphate, iron, and manganese. Sci. Total Environ. 2009, 407, 6085–6092. [Google Scholar] [CrossRef]
- Rousselaki, E.; Pavlidou, A.; Michalopoulos, P.; Dassenakis, M.; Scoullos, M. Nutrient benthic fluxes and porewater concentrations in the Gulf of Elefsis. In Sustainable Mediterranean, Issue No 71. In Proceedings of the International Conference: Environmental Perspectives of the Gulf of Elefsis, A Mediterranean Case Study Where Science Meets the Society, Elefsis, Greece, 11–12 September 2015; pp. 36–39. [Google Scholar]
- NASA—Global Climate Change. Available online: https://climate.nasa.gov/ (accessed on 21 November 2018).
- González-Dávila, M.; Santana-Casiano, J.M.; Petihakis, G.; Ntoumas, M.; Suárez de Tangil, M.; Krasakopoulou, E. Seasonal pH variability in the Saronikos gulf: A year-study using a new photometric pH sensor. J. Mar. Syst. 2016, 162, 37–46. [Google Scholar] [CrossRef]
- Mentzafou, A.; Dimitriou, E.; Zogaris, S. Integrated ecological assessment and restoration planning in a heavily modified peri-urban Mediterranean lagoon. Environ. Earth Sci. 2016, 75, 983. [Google Scholar] [CrossRef]
- Northcott, D.; Sevadjian, J.; Sancho-Gallegos, D.A.; Wahl, C.; Friederich, J.; Chavez, F.P. Impacts of urban carbon dioxide emissions on sea-air flux and ocean acidification in nearshore waters. PLoS ONE 2019, 14, e0214403. [Google Scholar] [CrossRef]
- Hassoun, A.; Fakhri, M.; Raad, N.; Abboud-Abi Saab, M.; Gemayel, E.; De Carlo, E.H. The carbonate system of the Eastern-most Mediterranean Sea, Levantine Sub-basin: Variations and drivers. Deep Sea Res. Part II Top. Stud. Oceanogr. 2019, 164, 54–73. [Google Scholar] [CrossRef]
- Wolf-Gladrow, D.A.; Zeebe, R.E.; Klaas, C.; Kortzinger, A.; Dickson, A.G. Total alkalinity: The explicit conservative expression and its application to biogeochemical processes. Mar. Chem. 2007, 106, 287–300. [Google Scholar] [CrossRef]
- Zervoudaki, S.; Frangoulis, C.; Giannoudi, L.; Krasakopoulou, E. Effects of low pH and raised temperature on egg production, hatching and metabolic rates of a Mediterranean copepod species (Acartia clausi) under oligotrophic conditions. Mediterr. Mar. Sci. 2014, 15, 74–83. [Google Scholar] [CrossRef][Green Version]
- Wimart-Rousseau, C.; Lajaunie-Salla, K.; Marrec, P.; Wagener, T.; Raimbault, P.; Lagadec, V.; Lafont, M.; Garcia, N.; Diaz, F.; Pinazo, C.; et al. Temporal variability of the carbonate system and air-sea CO2 exchanges in a Mediterranean human-impacted coastal site. Estuar. Coast. Shelf Sci. 2020, 236, 106641. [Google Scholar] [CrossRef]
- Kulinski, K.; Schneider, B.; Hammer, K.; Machulik, U.; Schulz-Bull, D. The influence of dissolved organic matter on the acid–base system of the Baltic Sea. J. Mar. Syst. 2014, 132, 106–115. [Google Scholar] [CrossRef]
- Howes, E.L.; Stemmann, L.; Assailly, C.; Irisson, J.O.; Dima, M.; Bijma, J.; Gattuso, J.-P. Pteropod time series from the North Western Mediterranean (1967–2003): Impacts of pH and climate variability. Mar. Ecol. Prog. Ser. 2015, 531, 193–206. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Field/ Condition | pHT | AT | pCO2 | CO2 | HCO3− | CO32− | DIC | Ωar. | Ωca | |
|---|---|---|---|---|---|---|---|---|---|---|
| Field | 2 m | 8.03 | 2974.0 | 522.6 | 13.7 | 2237.3 | 306.8 | 2557.8 | 4.80 | 7.19 |
| 10 m | 8.05 | 2990.9 | 504.2 | 15.1 | 2334.4 | 274.1 | 2623.6 | 4.18 | 6.38 | |
| 20 m | 7.76 | 2978.6 | 1089.0 | 36.0 | 2644.4 | 139.7 | 2820.1 | 2.11 | 3.24 | |
| 32 m | 7.62 | 2974.9 | 1538.4 | 52.3 | 2726.1 | 100.5 | 2879.0 | 1.51 | 2.33 | |
| OA | mean | 6.65 | 3622.2 | 18,867.8 | 641.1 | 3586.1 | 15.1 | 4242.4 | 0.22 | 0.34 |
| St. dev. | 0.06 | 475.4 | 2810.3 | 94.9 | 469.5 | 2.7 | 523.8 | 0.05 | 0.08 | |
| max | 6.73 | 4340.9 | 22,525.8 | 766.3 | 4292.3 | 20.3 | 4956.9 | 0.31 | 0.47 | |
| min | 6.58 | 3062.4 | 14,103.0 | 479.8 | 3039.1 | 12.4 | 3667.5 | 0,15 | 0.24 | |
| C | mean | 7.62 | 3143.18 | 1660.43 | 56.49 | 2893.69 | 107.06 | 3057.23 | 1.61 | 2.49 |
| St. dev. | 0.03 | 107.34 | 117.34 | 3.99 | 162.29 | 12.67 | 176.14 | 0.19 | 0.29 | |
| max | 7.66 | 3308.52 | 1812.77 | 61.67 | 3201.87 | 132.53 | 3394.52 | 2.00 | 3.08 | |
| min | 7.58 | 3064.52 | 1525.92 | 51.91 | 2759.01 | 98.25 | 2914.68 | 1.48 | 2.29 |
| Parameter | F | p | Parameter | F | p |
|---|---|---|---|---|---|
| DO | 0.015 | 0.903 | NO3− | 0.141 | 0.712 |
| AT | 3.910 | 0.066 | NO2− | 4.078 | 0.061 |
| HCO3− | 13.001 | 0.002 | NH4+ | 2.841 | 0.111 |
| CO32− | 11.097 | 0.004 | DON | 7.662 | 0.014 |
| DIC | 21.139 | 0.000 | PO43− | 18.981 | 0.000 |
| Ωar. | 11.097 | 0.004 | DOP | 1.402 | 0.254 |
| Ωcalc. | 11.097 | 0.004 | SiO44− | 0.126 | 0.728 |
| DOC | 1.423 | 0.250 | DIN:DIP | 0.823 | 0.378 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kapetanaki, N.; Krasakopoulou, E.; Stathopoulou, E.; Dassenakis, M.; Scoullos, M. Severe Coastal Hypoxia Interchange with Ocean Acidification: An Experimental Perturbation Study on Carbon and Nutrient Biogeochemistry. J. Mar. Sci. Eng. 2020, 8, 462. https://doi.org/10.3390/jmse8060462
Kapetanaki N, Krasakopoulou E, Stathopoulou E, Dassenakis M, Scoullos M. Severe Coastal Hypoxia Interchange with Ocean Acidification: An Experimental Perturbation Study on Carbon and Nutrient Biogeochemistry. Journal of Marine Science and Engineering. 2020; 8(6):462. https://doi.org/10.3390/jmse8060462
Chicago/Turabian StyleKapetanaki, Natalia, Evangelia Krasakopoulou, Eleni Stathopoulou, Manos Dassenakis, and Michael Scoullos. 2020. "Severe Coastal Hypoxia Interchange with Ocean Acidification: An Experimental Perturbation Study on Carbon and Nutrient Biogeochemistry" Journal of Marine Science and Engineering 8, no. 6: 462. https://doi.org/10.3390/jmse8060462
APA StyleKapetanaki, N., Krasakopoulou, E., Stathopoulou, E., Dassenakis, M., & Scoullos, M. (2020). Severe Coastal Hypoxia Interchange with Ocean Acidification: An Experimental Perturbation Study on Carbon and Nutrient Biogeochemistry. Journal of Marine Science and Engineering, 8(6), 462. https://doi.org/10.3390/jmse8060462