The Biotechnological Potential of Secondary Metabolites from Marine Bacteria

Abstract
1. Introduction
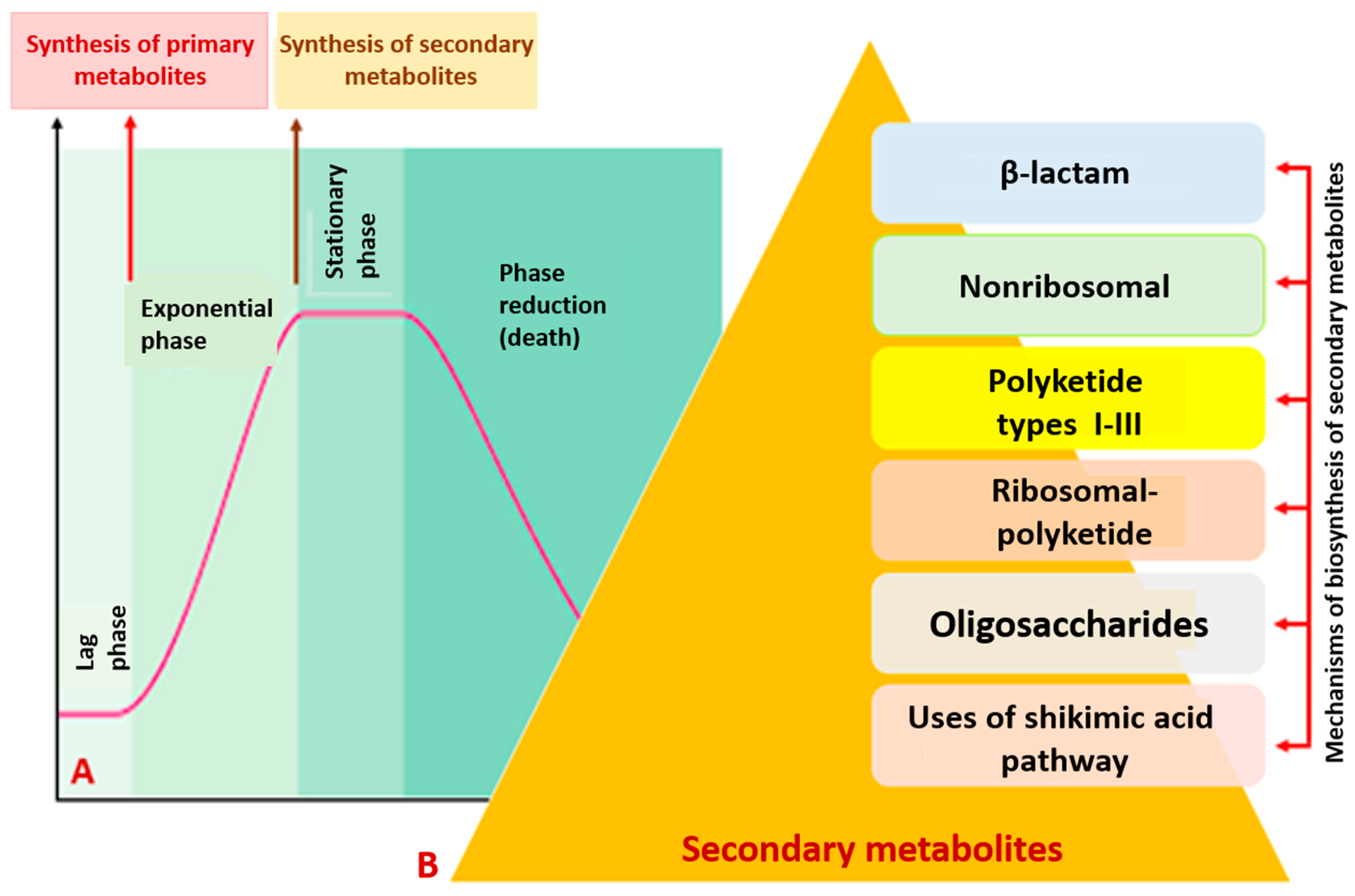
2. Bacterial Metabolites
Secondary Metabolites of Bacteria
3. Antimicrobial Substances of Marine Microorganisms
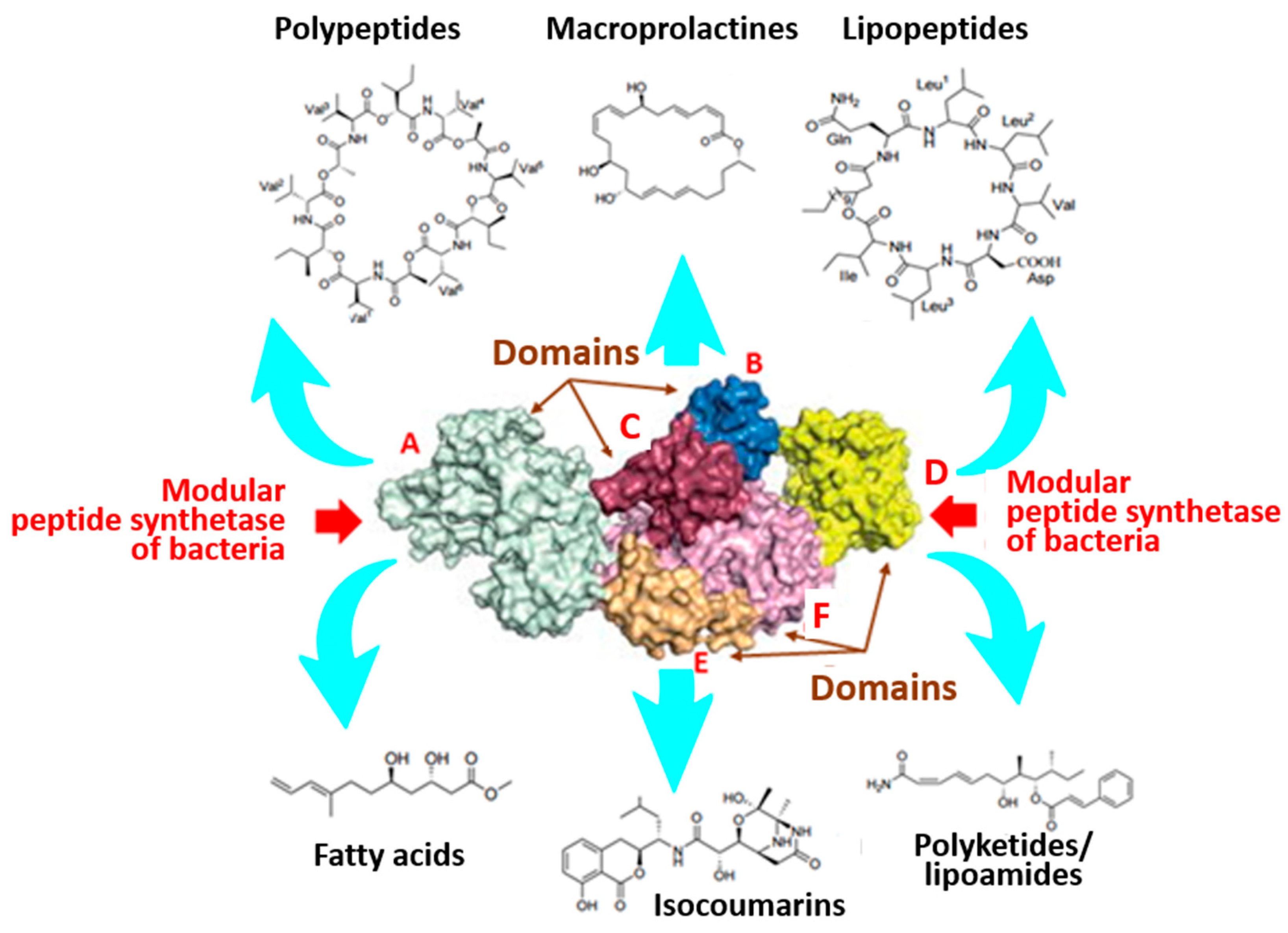
4. Marine Bacteria—An Inexhaustible Source of Non-Ribosomal Synthesized Secondary Metabolites
4.1. Cyclic Lipopeptides (cLPs)
4.2. Polyketides/Lipoamides
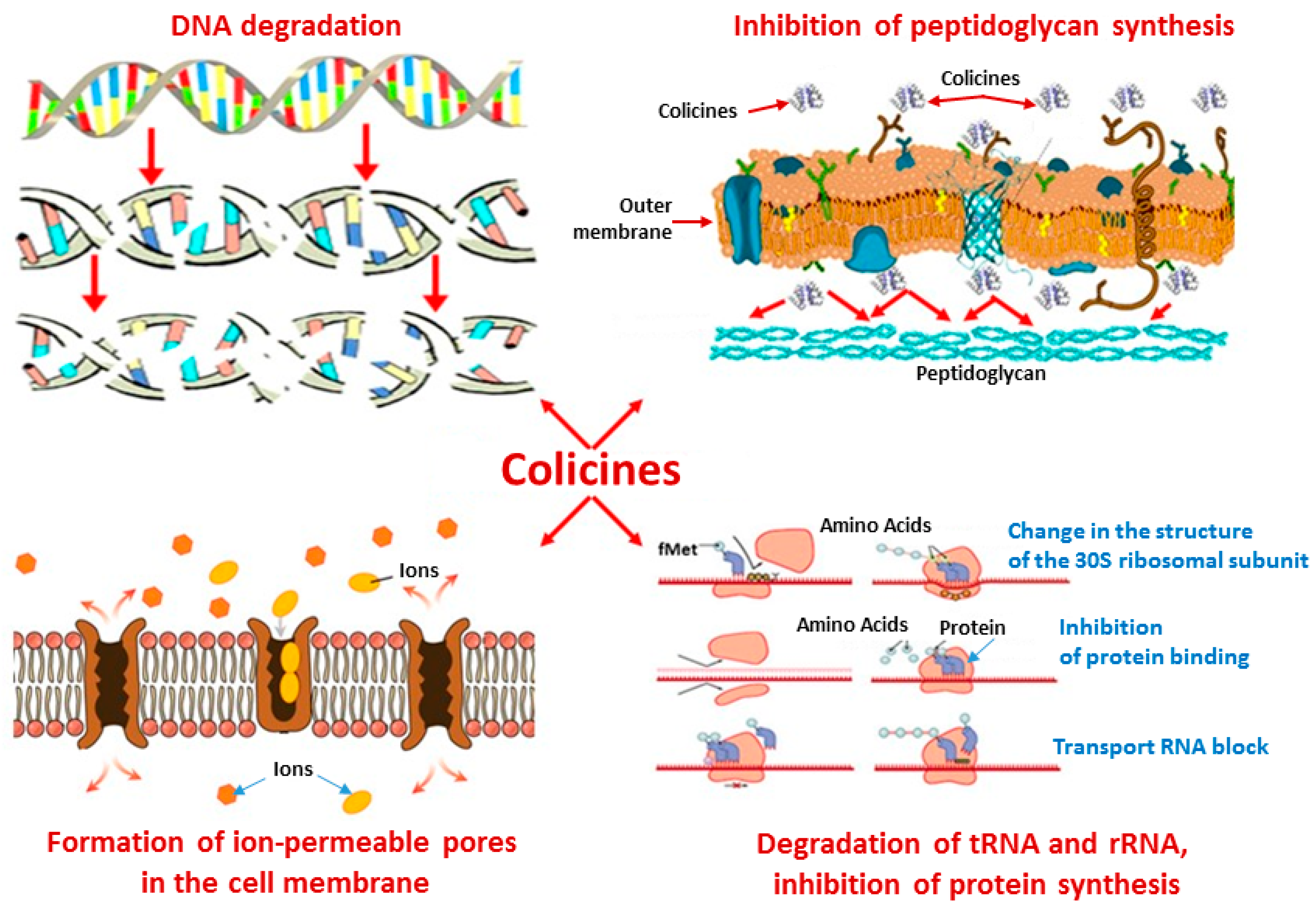
5. Mechanisms of Antimicrobial Action of Antibacterial Peptides
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization (WHO). Antimicrobial Resistance. 2015. Available online: http://www.who.int (accessed on 23 February 2019).
- Chokshi, A.; Sifri, Z.; Cennimo, D.; Horng, H. Global Contributors to Antibiotic Resistance. J. Glob. Infect. Dis. 2019, 11, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.K.; Seo, C.H.; Park, Y. Marine Peptides and Their Anti-Infective Activities. Mar. Drugs 2015, 13, 618–654. [Google Scholar] [CrossRef] [PubMed]
- Mondol, M.; Shin, H.; Islam, M. Diversity of Secondary Metabolites from Marine Bacillus Species: Chemistry and Biological Activity. Mar. Drugs 2013, 11, 2846–2872. [Google Scholar] [CrossRef] [PubMed]
- Böhringer, N.; Fisch, K.M.; Schillo, D.; Bara, R.; Hertzer, C.; Grein, F.; Eisenbarth, J.H.; Kaligis, F.; Schneider, T.; Wägele, H.; et al. Antimicrobial Potential of Bacteria Associated with Marine Sea Slugs from North Sulawesi, Indonesia. Front. Microbiol. 2017, 8, 1092. [Google Scholar] [CrossRef] [PubMed]
- Andryukov, B.G.; Zaporozhets, T.S.; Besednova, N.N. Promising Strategies for Finding New Means of Combating Infectious Diseases. Antibiot. Chemother. 2018, 63, 44–55. [Google Scholar] [CrossRef]
- Andryukov, B.G.; Mikhaylov, V.V.; Besednova, N.N.; Zaporozhets, T.S.; Bynina, M.P.; Matosova, E.V. The Bacteriocinogenic Potential of Marine Microorganisms. Russ. J. Mar. Biol. 2018, 44, 433–441. [Google Scholar] [CrossRef]
- Gokulan, K.; Khare, S.; Cerniglia, C. Metabolic pathways. Production of Secondary Metabolites of Bacteria. Encycl. Food Microbiol. 2014, 561–569. [Google Scholar] [CrossRef]
- Wang, Y.-P.; Lei, Q.-Y. Metabolite sensing and signaling in cell metabolism. Signal Transduct. Target. Ther. 2018, 3, 30. [Google Scholar] [CrossRef]
- Pinu, F.R.; Villas-Boas, S.G.; Aggio, R. Analysis of Intracellular Metabolites from Microorganisms: Quenching and Extraction Protocols. Metabolites 2017, 7, 53. [Google Scholar] [CrossRef]
- Niu, G.Q.; Tan, H.R. Biosynthesis and regulation of secondary metabolites in microorganisms. Life Sci. 2013, 56, 581–583. [Google Scholar] [CrossRef][Green Version]
- Baral, B.; Akhgari, A.; Metsä-Ketelä, M. Activation of microbial secondary metabolic pathways: Avenues and challenges. Synth. Syst. Biotechnol. 2018, 3, 163–178. [Google Scholar] [CrossRef]
- Wright, G.D. Something old, something new: revisiting natural products in antibiotic drug discovery. Can. J. Microbiol. 2014, 60, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Tareq, F.S.; Kim, J.H.; Lee, M.A.; Lee, H.S.; Lee, Y.J.; Lee, J.S.; Shin, H.J. Ieodoglucomides A and B from a marine-derived bacterium Bacillus licheniformis. Org. Lett. 2012, 14, 1464–1467. [Google Scholar] [CrossRef] [PubMed]
- Robbins, T.; Liu, Y.C.; Cane, D.E.; Khosla, C. Structure and mechanism of assembly line polyketide synthases. Curr. Opin. Struct. Biol. 2016, 41, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Strieker, M.; Tanović, A.; Marahiel, M.A. Nonribosomal peptide synthetases: structures and dynamics. Curr. Opin. Struct. Biol. 2010, 20, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Gulick, A.M. Nonribosomal peptide synthetase biosynthetic clusters of ESKAPE pathogens. Nat. Prod. Rep. 2017, 34, 981–1009. [Google Scholar] [CrossRef] [PubMed]
- Alfermann, J.; Sun, X.; Mayerthaler, F.; Morrell, T.E.; Dehling, E.; Volkmann, G.; Komatsuzaki, T.; Yang, H.; Mootz, H.D. FRET monitoring of a nonribosomal peptide synthetase. Nat. Chem. Biol. 2017, 13, 1009–1015. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, A.; Naughton, L.M.; Montánchez, I.; Dobson, A.D.W.; Rai, D.K. Current Status and Future Prospects of Marine Natural Products (MNPs) as Antimicrobials. Mar. Drugs 2017, 15, 272. [Google Scholar] [CrossRef]
- Chen, D.; Qian, X. A Brief History of Bacteria: The Everlasting Game Between Humans and Bacteria; World Scientific Publishing Company: Singapore, 2017; p. 296. [Google Scholar]
- Mikhailov, V.V.; Pivkin, M.V. The Study of Marine Bacteria and Fungi; Some results and prospects of research; Bulletin of the Far Eastern Branch of the Russian Academy of Sciences: Vladivostok, Russia, 2014; Volume 173, pp. 149–156. [Google Scholar]
- Stonik, V.A.; Mikhailov, V.V. Prospects for the use of microorganisms of the marginal seas of the Far East and the Arctic for the search and practical application of natural bioactive substances. In Scientific and Technical Problems of the Development of the Arctic; Publishing Academic Book: Moscow, Russia, 2015; pp. 412–425. [Google Scholar]
- Timmermans, M.L.; Paudel, Y.P.; Ross, A.C. Investigating the Biosynthesis of Natural Products from Marine Proteobacteria: A Survey of Molecules and Strategies. Mar. Drugs 2017, 15, 235. [Google Scholar] [CrossRef]
- Manivasagan, P.; Venkatesan, J.; Sivakumar, K.; Kim, S.K. Pharmaceutically active secondary metabolites of marine actinobacteria. Microbiol. Res. 2014, 169, 262–278. [Google Scholar] [CrossRef]
- Versluis, D.; Nijsse, B.; Naim, M.A.; Koehorst, J.J.; Wiese, J.; Imhoff, J.F.; Schaap, P.J.; van Passel, M.W.J.; Smidt, H.; Sipkema, D. Comparative Genomics Highlights Symbiotic Capacities and High Metabolic Flexibility of the Marine Genus Pseudovibrio. Genome Biol. Evol. 2018, 10, 125–142. [Google Scholar] [CrossRef]
- Yamashita, T.; Kuranaga, T.; Inoue, M. Solid-Phase Total Synthesis of Bogorol A: Stereocontrolled Construction of Thermodynamically Unfavored (E)-2-Amino-2-butenamide. Org. Lett. 2015, 17, 2170–2173. [Google Scholar] [CrossRef] [PubMed]
- Jang, C.H.; Park, H.; Cho, Y.B.; Choi, C.H. Effect of vancomycin-coated tympanostomy tubes on methicillin-resistant Staphylococcus aureus biofilm formation: In Vitro study. J. Laryngol. Otol. 2010, 124, 594–598. [Google Scholar] [CrossRef] [PubMed]
- Tuin, A.W.; Grotenbreg, G.M.; Spalburg, E.; de Neeling, A.J.; Mars-Groenendijk, R.H.; van der Marel, G.A.; Noort, D.; Overkleeft, H.S.; Overhand, M. Structural and biological evaluation of some loloatin C analogues. Bioorg. Med. Chem. 2009, 17, 6233–6240. [Google Scholar] [CrossRef] [PubMed]
- Rahman, H.; Austin, B.; Mitchell, W.J.; Morris, P.C.; Jamieson, D.J.; Adams, D.R.; Spragg, A.M.; Schweizer, M. Novel anti-infective compounds from marine bacteria. Mar. Drugs 2010, 8, 498–518. [Google Scholar] [CrossRef] [PubMed]
- Desjardine, K.; Pereira, A.; Wright, H.; Matainaho, T.; Kelly, M.; Andersen, R.J. Tauramamide, a lipopeptide antibiotic produced in culture by Brevibacillus laterosporus isolated from a marine habitat: Structure elucidation and synthesis. J. Nat. Prod. 2007, 70, 1850–1853. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, S.; Acharya, D.; Adholeya, A.; Barrow, C.J.; Deshmukh, S.K. Nonribosomal Peptides from Marine Microbes and Their Antimicrobial and Anticancer Potential. Front. Pharmacol. 2017, 8, 828. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.F.; Guo, Y.W. Bioactive natural products from Chinese marine flora and fauna. Acta Pharmacol. Sin. 2012, 33, 1159–1169. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yuan, J.; Zhao, M.; Li, R.; Huang, Q.; Rensing, C.; Raza, W.; Shen, Q. Antibacterial Compounds-Macrolactin Alters the Soil Bacterial Community and Abundance of the Gene Encoding PKS. Front. Microbiol. 2016, 7, 1904. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.W.; Kim, J.M.; Kwon, M.H.; Kim, D.H.; Kang, H.E. Pharmacokinetics of macrolactin A and 7-O-succinyl macrolactin A in mice. Xenobiotica 2014, 44, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Pettit, G.R.; Knight, J.C.; Herald, D.L.; Pettit, R.K.; Hogan, F.; Mukku, V.J.; Hamblin, J.S.; Dodson, M.J.; Chapuis, J.C. Antineoplastic agents. 570. Isolation and structure elucidation of bacillistatins 1 and 2 from a marine Bacillus silvestris. J. Nat. Prod. 2009, 72, 366–371. [Google Scholar] [CrossRef]
- Raimundo, I.; Silva, S.G.; Costa, R.; Keller-Costa, T. Bioactive Secondary Metabolites from Octocoral-Associated Microbes—New Chances for Blue Growth. Mar. Drugs 2018, 16, 485. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, K.; Degnes, K.F.; Zotchev, S.B. Isolation and characterization of the gene cluster for biosynthesis of the thiopeptide antibiotic TP-1161. Appl. Environ. Microbiol. 2010, 76, 7093–7101. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, K.; Degnes, K.F.; Kemmler, M.; Bredholt, H.; Fjaervik, E.; Klinkenberg, G.; Sletta, H.; Ellingsen, T.E.; Zotchev, S.B. Production of a new thiopeptide antibiotic, TP-1161, by a marine Nocardiopsis species. Appl. Environ. Microbiol. 2010, 76, 4969–4976. [Google Scholar] [CrossRef] [PubMed]
- Romero, J.; Feijoo, C.G.; Navarrete, P. Antibiotics in Aquaculture—Use, Abuse and Alternatives. Health and Environment in Aquaculture; Carvalho, E., Ed.; InTech Europe: Rijeka, Croatia, 2012; 200p. [Google Scholar]
- Galinier, R.; Roger, E.; Sautiere, P.E.; Aumelas, A.; Banaigs, B.; Mitta, G. Halocyntin and papillosin, two new antimicrobial peptides isolated from hemocytes of the solitary tunicate, Halocynthia papillosa. J. Pept. Sci. 2009, 15, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Kojima, H.; Shinohara, R.; Itonori, S.; Ito, M. Characterization of a Novel Rhamnose-containing Acidic Glycosphingolipid from the Ascidian Halocynthia aurantium. J. Oleo Sci. 2017, 66, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Slightom, R.N.; Buchan, A. Surface colonization by marine roseobacters: Integrating genotype and phenotype. Appl. Environ. Microbiol. 2009, 75, 6027–6037. [Google Scholar] [CrossRef] [PubMed]
- Cude, W.N.; Mooney, J.; Tavanaei, A.A.; Hadden, M.K.; Frank, A.M.; Gulvik, C.A.; May, A.L.; Buchan, A. Production of the antimicrobial secondary metabolite indigoidine contributes to competitive surface colonization by the marine roseobacter Phaeobacter sp. strain Y4I. Appl. Environ. Microbiol. 2012, 78, 4771–4780. [Google Scholar] [CrossRef] [PubMed]
- Oku, N.; Kawabata, K.; Adachi, K.; Katsuta, A.; Shizuri, Y. Unnarmicins A and C, new antibacterial depsipeptides produced by marine bacterium Photobacterium sp. MBIC06485. J. Antibiot. 2008, 61, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Oku, N.; Adachi, K.; Matsuda, S.; Kasai, H.; Takatsaki, A.; Shizuri, Y. Ariakemicins A and B, novel polyketide-peptide antibiotics from a marine gliding bacterium of the genus Rapidithrix. Org. Lett. 2008, 10, 2481–2484. [Google Scholar] [CrossRef] [PubMed]
- Fotie, J.; Morgan, R.E. Depsipeptides from microorganisms: A new class of antimalarials. Mini Rev. Med. Chem. 2008, 8, 1088–1094. [Google Scholar] [CrossRef] [PubMed]
- Machado, H.; Månsson, M.; Gram, L. Draft Genome Sequence of Photobacterium halotolerans S2753, Producer of Bioactive Secondary Metabolites. Genome Announc. 2014, 2, e00535-14. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Mishra, V. Marine peptides act as novel chemotherapeutic agent. J. Microbiol. Exp. 2018, 6, 267–270. [Google Scholar]
- Rungprom, W.; Siwu, E.R.O.; Lambert, L.K.; Dechsakulwatana, C.; Barden, M.C.; Kokpol, U.; Blanchfield, J.T.; Kita, M.; Garson, M.J. Cyclic tetrapeptides from marine bacteria associated with the seaweed Diginea sp. and the sponge Halisarca ectofibrosa. Tetrahedron 2008, 64, 3147–3152. [Google Scholar] [CrossRef]
- Li, D.; Carr, G.; Zhang, Y.; Williams, D.E.; Amlani, A.; Bottriell, H.; Mui, A.L.; Andersen, R.J. Turnagainolides A and B, cyclic depsipeptides produced in culture by a Bacillus sp.: isolation, structure elucidation, and synthesis. J. Nat. Prod. 2011, 74, 1093–1099. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Phelan, V.; Ntai, I.; Farnet, C.M.; Zazopoulos, E.; Bachmann, B.O. Benzodiazepine biosynthesis in Streptomyces refuineus. Chem. Biol. 2007, 14, 691–701. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.M.; Kim, Y.B.; Choi, S.; Lee, Y.; Shin, S.G.; Unno, T.; Kim, Y.M. Prevalence of antibiotic resistance genes from effluent of coastal aquaculture, South Korea. Environ. Pollut. 2018, 233, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Yukiya, A. (Ed.) IAEA Bulletin: Protecting Our Marine Environment; IAEA Bulletin: Vienna, Austria, 2013. [Google Scholar]
- Chen, E.; Chen, Q.; Chen, S.; Xu, B.; Ju, J.; Wang, H. Mathermycin, Lantibiotic marine Actinomycete Marinactinospora thermotolerans SCSIO 00652. Appl. Environ. Microbiol. 2017, 83, e00926-17. [Google Scholar] [CrossRef]
- Karvonen, A.; Rintamäki, P.; Jokela, J.; Valtonen, E.T. Increasing water temperature and disease risks in aquatic systems: Climate change increases the risk of some, but not all, diseases. Int. J. Parasitol. 2010, 40, 1483–1488. [Google Scholar] [CrossRef]
- Das, S.; Ward, L.R.; Burke, C. Prospects of using marine actinobacteria as probiotics in aquaculture. Appl. Microbiol. Biotechnol. 2008, 81, 419–429. [Google Scholar] [CrossRef]
- Kim, S.K.; Bhatnagar, I.; Kang, K.H. Development of marine probiotics: Prospects and approach. Adv. Food Nutr. Res. 2012, 65, 353–362. [Google Scholar]
- Felnagle, E.A.; Jackson, E.E.; Chan, Y.A.; Podevels, A.M.; Berti, A.D.; McMahon, M.D.; Thomas, M.G. Nonribosomal Peptide Synthetases Involved in the Production of Medically Relevant Natural Products. Mol. Pharm. 2008, 5, 191–211. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.R.; Gulick, A.M. Structural Biology of Nonribosomal Peptide Synthetases. Methods Mol. Biol. 2016, 1401, 3–29. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Chaudhary, S.; Sareen, D. Nonribosomal peptide synthetases: Identifying the cryptic gene clusters and decoding the natural product. J. Biosci. 2017, 42, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Reimer, J.M.; Harb, I.; Ovchinnikova, O.G.; Jiang, J.; Whitfield, C.; Schmeing, T.M. Structural Insight into a Novel Formyltransferase and Evolution to a Nonribosomal Peptide Synthetase Tailoring Domain. ACS Chem Biol. 2018, 13, 3161–3172. [Google Scholar] [CrossRef] [PubMed]
- Klapper, M.; Braga, D.; Lackner, G.; Herbst, R.; Stallforth, P. Bacterial Alkaloid Biosynthesis: Structural Diversity via a Minimalistic Nonribosomal Peptide Synthetase. Cell Chem Biol. 2018, 25, 659–665.e9. [Google Scholar] [CrossRef]
- Bloudoff, K.; Schmeing, T.M. Structural and functional aspects of the nonribosomal peptide synthetase condensation domain superfamily: Discovery, dissection and diversity. Biochim. Biophys. Acta Proteins Proteom. 2017, 1865, 1587–1604. [Google Scholar] [CrossRef] [PubMed]
- Gratia, J.P. Andre Gratia: A forerunner in microbial and viral genetics. Genetics 2000, 156, 471–476. [Google Scholar] [PubMed]
- Reimer, J.M.; Haque, A.S.; Tarry, M.J.; Schmeing, T.M. Piecing together nonribosomal peptide synthesis. Curr. Opin. Struct. Biol. 2018, 49, 104–113. [Google Scholar] [CrossRef]
- Kitagaki, J.; Shi, G.; Miyauchi, S.; Murakami, S.; Yang, Y. Cyclic depsipeptides as potential cancer therapeutics. Anti Cancer Drugs 2015, 26, 259–271. [Google Scholar] [CrossRef]
- Offret, C.; Desriac, F.; Le Chevalier, P.; Mounier, J.; Jégou, C.; Fleury, Y. Spotlight on Antimicrobial Metabolites from the Marine Bacteria Pseudoalteromonas: Chemodiversity and Ecological Significance. Mar. Drugs 2016, 14, 129. [Google Scholar] [CrossRef]
- Giang, C.N.D.; Sebesvari, Z.; Renaud, F.; Rosendahl, I.; Minh, Q.H.; Amelung, W. Occurrence and Dissipation of the Antibiotics Sulfamethoxazole, Sulfadiazine, Trimethoprim, and Enrofloxacin in the Mekong Delta, Vietnam. PLoS ONE 2015, 10, e0131855. [Google Scholar]
- Das, S.; Ward, L.R.; Burke, C. Screening of marine Streptomyces spp. for potential use as probiotics in aquaculture. Aquaculture 2010, 305, 32–41. [Google Scholar] [CrossRef]
- Gao, X.Y.; Liu, Y.; Miao, L.L.; Li, E.W.; Hou, T.T.; Liu, Z.P. Mechanism of anti-Vibrio activity of marine probiotic strain Bacillus pumilus H2, and characterization of the active substance. AMB Express 2017, 7, 23. [Google Scholar] [CrossRef] [PubMed]
- Kers, J.A.; Sharp, R.E.; Defusco, A.W.; Park, J.H.; Xu, J.; Pulse, M.E.; Weiss, W.J.; Handfield, M. Mutacin 1140 Lantibiotic Variants Are Efficacious Against Clostridium Difficile Infection. Front. Microbiol. 2018, 9, 415. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, B.R.; Sahoo, P.K. Edwardsiellosis in fish: A brief review. J. Biosci. 2007, 32, 1331–1344. [Google Scholar] [CrossRef] [PubMed]
- Phelan, R.W.; Barret, M.; Cotter, P.D.; O’Connor, P.; Chen, R.; Morrissey, J.; Dobson, A.; O’Gara, F.; Barbosa, T. Subtilomycin: A new lantibiotic from Bacillus subtilis strain MMA7 isolated from the marine sponge Haliclona simulans. Mar. Drugs 2013, 11, 1878–1898. [Google Scholar] [CrossRef] [PubMed]
- Selvin, J.; Joseph, S.; Asha, K.R.; Manjusha, W.A.; Sangeetha, V.S.; Jayaseema, D.M.; Antony, M.C.; Denslin Vinitha, A.J. Antibacterial potential of antagonistic Streptomyces sp. isolated from marine sponge Dendrilla nigra. FEMS Microbiol. Ecol. 2004, 50, 117–122. [Google Scholar] [CrossRef]
- Rivetti, I.; Fraschetti, S.; Lionello, P.; Zambianchi, E.; Boero, F. Global Warming and Mass Mortalities of Benthic Invertebrates in the Mediterranean Sea. PLoS ONE 2014, 9, e115655. [Google Scholar] [CrossRef] [PubMed]
- Romanenko, L.A.; Uchino, M.; Kalinovskaya, N.I.; Mikhailov, V.V. Isolation, phylogenetic analysis and screening of marine mollusc-associated bacteria for antimicrobial, hemolytic and surface activities. Microbiol. Res. 2008, 163, 633–644. [Google Scholar] [CrossRef]
- Pettit, G.R.; Ye, Q.; Herald, D.L.; Hogan, F.; Pettit, R.K. Antineoplastic agents. 573. isolation and structure of papilistatin from the papilionid butterfly Byasa polyeuctes termessa. J Nat Prod. 2010, 26, 164–166. [Google Scholar] [CrossRef]
- Mansson, M.; Gram, L.; Larsen, T.O. Production of bioactive secondary metabolites by marine Vibrionaceae. Mar. Drugs 2011, 9, 1440–1468. [Google Scholar] [CrossRef] [PubMed]
- Jang, K.H.; Nam, S.J.; Locke, J.B.; Kauffman, C.A.; Beatty, D.S.; Paul, L.A.; Fenical, W. Anthracimycin, a potent anthrax antibiotic from a marine-derived actinomycete. Angew. Chem. Int. Ed. 2013, 52, 7822–7824. [Google Scholar] [CrossRef] [PubMed]
- Hammami, R.; Zouhir, A.; Ben Hamida, J.; Fliss, I. BACTIBASE: A new web-accessible database for bacteriocin characterization. BMC Microbiol. 2007, 7, 89. [Google Scholar] [CrossRef] [PubMed]
- De Jong, A.; van Heel, A.J.; Kok, J.; Kuipers, O.P. BAGEL2: Mining for bacteriocins in genomic data. Nucleic Acids Res. 2010, 38, W647–W651. [Google Scholar] [CrossRef] [PubMed]
- Van Heel, A.J.; de Jong, A.; Montalban-Lopez, M.; Kok, J.; Kuipers, O.P. BAGEL3: Automated identification of genes encoding bacteriocins and (non-)bactericidal posttranslationally modified peptides. Nucleic Acids Res. 2013, 41, W448–W453. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, X.; Wang, Z. APD3: the antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2015, 44, D1087–D1093. [Google Scholar] [CrossRef] [PubMed]
- Brahmachary, M.; Krishnan, S.P.; Koh, J.L.; Khan, A.M.; Seah, S.H.; Tan, T.W.; Brusic, V.; Bajic, V.B. ANTIMIC: A database of antimicrobial sequences. Nucleic Acids Res. 2004, 32, D586–D589. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.K.; Kaas, Q.; Chiche, L.; Craik, D.J. CyBase: A database of cyclic protein sequences and structures, with applications in protein discovery and engineering. Nucleic Acids Res. 2007, 36, D206–D210. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yin, T.; Xiao, X.; He, D.; Xue, Z.; Jiang, X.; Wang, Y. StraPep: A structure database of bioactive peptides. Database 2018, 2018, bay038. [Google Scholar] [CrossRef] [PubMed]
- Bibi, F.; Faheem, M.; Azhar, E.I.; Yasir, M.; Alvi, S.A.; Kamal, M.A.; Ullah, I.; Naseer, M.I. Bacteria From Marine Sponges: A Source of New Drugs. Curr. Drug Metab. 2017, 18, 11–15. [Google Scholar] [CrossRef]
- Ruocco, N.; Costantini, S.; Palumbo, F.; Costantini, M. Marine Sponges and Bacteria as Challenging Sources of Enzyme Inhibitors for Pharmacological Applications. Mar. Drugs 2017, 15, 173. [Google Scholar] [CrossRef] [PubMed]
- Dias, T.; Gaudêncio, S.P.; Pereira, F. A Computer-Driven Approach to Discover Natural Product Leads for Methicillin-Resistant Staphylococcus aureus Infection therapy. Mar. Drugs 2018, 17, 16. [Google Scholar] [CrossRef] [PubMed]
- Giordano, D.; Coppola, D.; Russo, R.; Verde, C.; Denaro, R.; Giuliano, L.; Lauro, F.M.; di Prisco, G. Marine Microbial Secondary Metabolites: Pathways, Evolution and Physiological Roles. Adv. Microb. Physiol. 2015, 66, 357–428. [Google Scholar] [CrossRef] [PubMed]
- Amin, S.A.; Parker, M.S.; Armbrust, E.V. Interactions between diatoms and bacteria. Microbiol. Mol. Biol. Rev. 2012, 76, 667–684. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, X.; Wang, J. Recent Advances in Antibacterial and Antiendotoxic Peptides or Proteins from MarineResources. Mar. Drugs 2018, 16, 57. [Google Scholar] [CrossRef]
- Pereira, F.; Aires-de-Sousa, J. Computational Methodologies in the Exploration of Marine Natural Product Leads. Mar. Drugs 2018, 16, 236. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Torres, V.; Encinar, J.A.; Herranz-López, M.; Pérez-Sánchez, A.; Galiano, V.; Barrajón-Catalán, E.; Micol, V. An Updated Review on Marine Anticancer Compounds: The Use of Virtual Screening for the Discovery of Small-Molecule Cancer Drugs. Molecules 2017, 22, 1037. [Google Scholar] [CrossRef]
- Nguyen, T.V.; Alfaro, A.C.; Young, T.; Green, S.; Zarate, E.; Merien, F. Itaconic acid inhibits growth of a pathogenic marine Vibrio strain: A metabolomics approach. Sci. Rep. 2019, 9, 5937. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Primary Metabolites | Secondary Metabolites |
|---|---|
| Small-sized molecules | Small-sized molecules |
| Produce several intermediate and terminal product | Participate in the synthesis of new compounds and a multitude of molecules |
| Terminal products involved in the synthesis of macromolecules, coenzyme | Not vitally important for cell growth |
| Important for cell growth and viability | Have unusual chemical structures |
| Have a simple chemical structure | Terminal products are used as antibacterial agents |
| Synthesized during the lag phase of bacterial growth | Synthesized at the beginning of stationary phase of bacterial growth |
| Used in food and feed industry | Used in medicine, cosmetics and agriculture as preservatives |
| Provide energy reserve for communication of cells | Protect bacteria during the period of adverse conditions |
| Main source of energy for cellular metabolism and life support | Participate in intercellular communication, cell protection and competition for food and space |
| Metabolite | Producer | Inhibiting | Active Concentration | References |
|---|---|---|---|---|
| Bogorol A | Bacillus sp. | Methicillin-resistant Staphylococcus aureus (MRSA) | 2 µg/mL (MIC) | [26,27] |
| Loloatin B | Bacillus sp. | Methicillin-resistant S. aureus (MRSA), vancomycin-resistant Enterococcus faecium (VRE) | 1–2 µg/mL (MIC) | [28,29] |
| Tauramamide | Brevibacillus laterosporus | Enterococcus sp. | 0.1 μg/mL (MIC) | [30,31] |
| Halobacillin | Bacillus sp. CND-914 | S. aureus, P. vulgaris and E. faecalis. Human HCT-116 cancer cells | 0.98 µg/mL (IC50) | [4,32] |
| Macrolactin S | Bacillus amyloliquefaciens | Escherichia coli, S. aureus | 0.1– 0.3 µg/mL (MIC) | [33,34] |
| Macrolactin V | B. amyloliquefaciens | E. coli, Bacillus subtilis, S. aureus | 0.1 µg/mL (MIC) | [33,34] |
| Bacillistatins | Bacillus silvestris | Streptococcus pneumonia | 0.5–2 μg/mL (GI50) | [35] |
| Triopeptid TP-1161 | Nocardiopsis sp. | Vancomycin-resistant Enterococcus faecium (VRE) | 1.0 μg/mL (MIC) | [36,37,38,39] |
| Halocintin | Halocynthia papillosa | Micrococcus luteus, Bacillus megaterium, Aerococcus viridans, S. aureus, Enterococcus faecalis | 0.39–50 μM (MBC) | [40,41] |
| Indigoidin | Phaeobacter sp. | Vibrio fischeri | n/d | [42,43] |
| Unnarrmicins A, C | Photobacterium sp. | Pseudovibrio sp. | 7–18 μg/disk | [44,45] |
| Ngercheumicins A–D | Photobacterium sp. | Gram (-) bacteria | n/d | [31,46] |
| Solonamidin A | Photobacterium sp. | S. aureus, methicillin-resistant S. aureus (MRSA) | n/d | [3,47] |
| Cyclo-peptides | Pseudomonas sp. | S. aureus, M. luteus, B. subtilis, E. coli, Vibrio anguillarum | n/d | [3,4,48,49] |
| Ariakemicins A, B | Rapidithrix sp. | Brevibacterium sp., S. aureus, B. subtilis | 0.46–80 μg/mL (MIC) | [44,45] |
| Turnagainolides A, B | Bacillus sp. RJA 2194 | MRSA, VRE, and penicillin-resistant Streptococcus pneumoniae | 1–2 μg/mL (MIC) | [50] |
| Anthramycin | Streptomyces sp. | Bacillus anthracis, E. faecalis, S. pneumonia, S. aureus, MSSA, MRSA, S. aureus (VRE) | 0.03125–0.25 μg/mL (MIC) | [51,52] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andryukov, B.; Mikhailov, V.; Besednova, N. The Biotechnological Potential of Secondary Metabolites from Marine Bacteria. J. Mar. Sci. Eng. 2019, 7, 176. https://doi.org/10.3390/jmse7060176
Andryukov B, Mikhailov V, Besednova N. The Biotechnological Potential of Secondary Metabolites from Marine Bacteria. Journal of Marine Science and Engineering. 2019; 7(6):176. https://doi.org/10.3390/jmse7060176
Chicago/Turabian StyleAndryukov, Boris, Valery Mikhailov, and Nataly Besednova. 2019. "The Biotechnological Potential of Secondary Metabolites from Marine Bacteria" Journal of Marine Science and Engineering 7, no. 6: 176. https://doi.org/10.3390/jmse7060176
APA StyleAndryukov, B., Mikhailov, V., & Besednova, N. (2019). The Biotechnological Potential of Secondary Metabolites from Marine Bacteria. Journal of Marine Science and Engineering, 7(6), 176. https://doi.org/10.3390/jmse7060176