

Multiscale Structural Patterns of Intertidal Salt Marsh Vegetation in Estuarine Wetlands and Its Interactions with Tidal Creeks

Abstract
1. Introduction
2. Materials and Methods
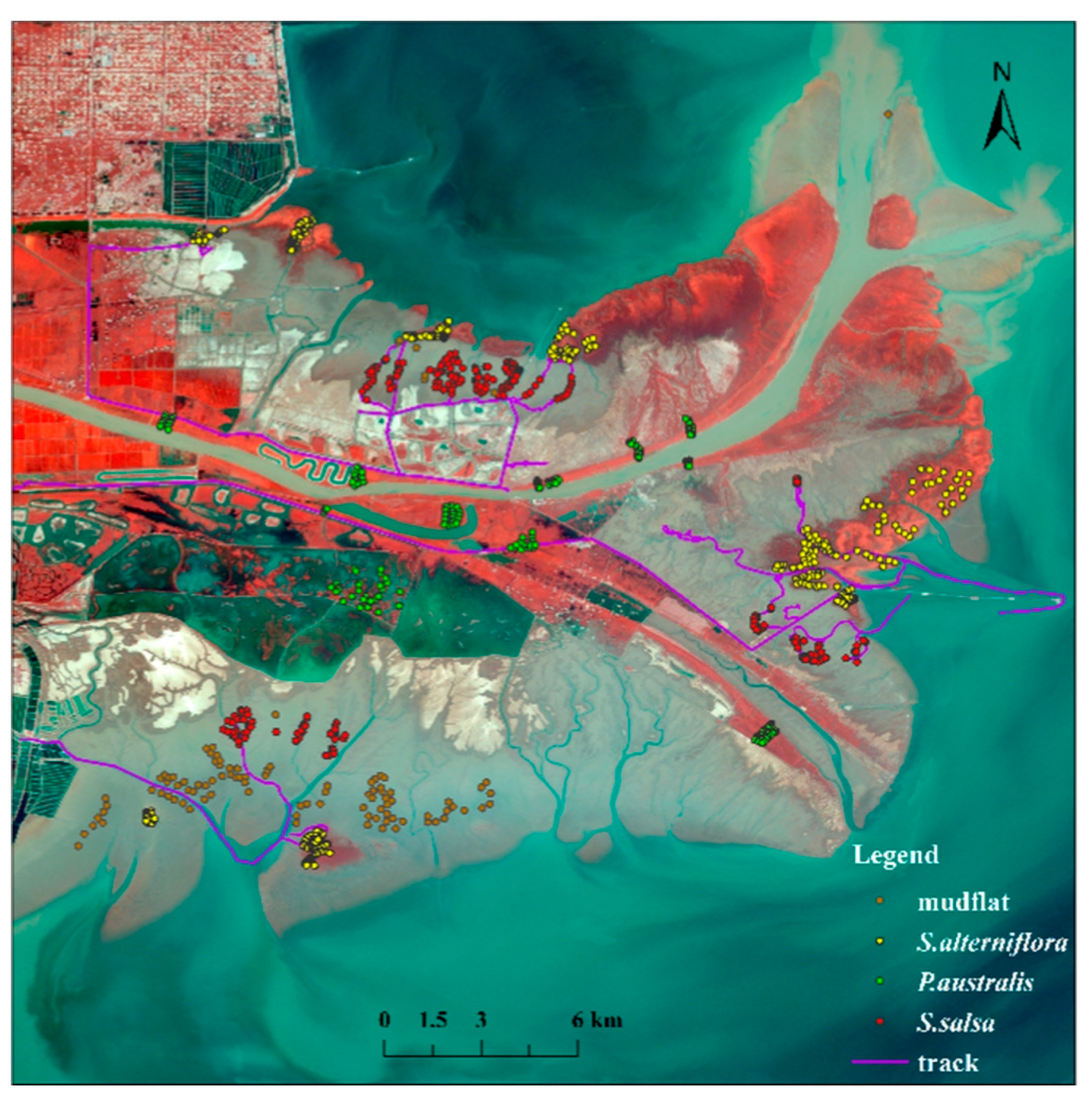
2.1. Study Area
2.2. Construction of the Annual Time Series Salt Marsh Vegetation Dataset
2.2.1. Development of the Multidimensional Feature Set
2.2.2. Construction of the Sample Sets
2.2.3. Extraction of Long-Term Time-Series Salt Marsh Vegetation Data Based on GEE
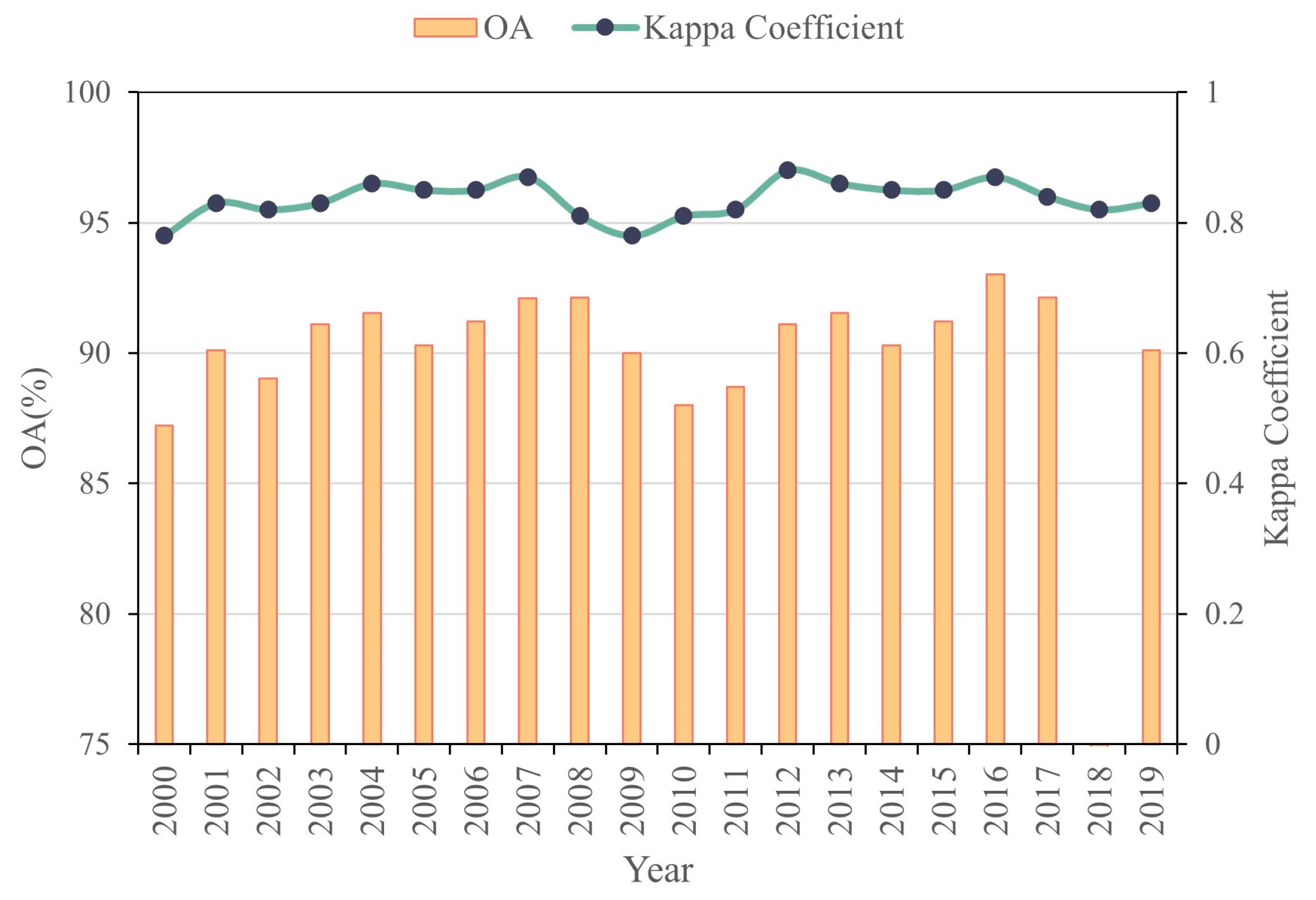
2.2.4. Accuracy Verification Method
2.3. Spatio-Temporal Succession Patterns of Salt Marsh Vegetation at the Landscape Scale
2.4. Spatio-Temporal Succession Patterns of Salt Marsh Vegetation at the Pixel Scale
2.5. Methods for Analyzing Coupled Interactions
2.5.1. Coupling Coefficients
2.5.2. GeoDetector Model
3. Results
3.1. Spatial Distribution of Salt Marsh Vegetation
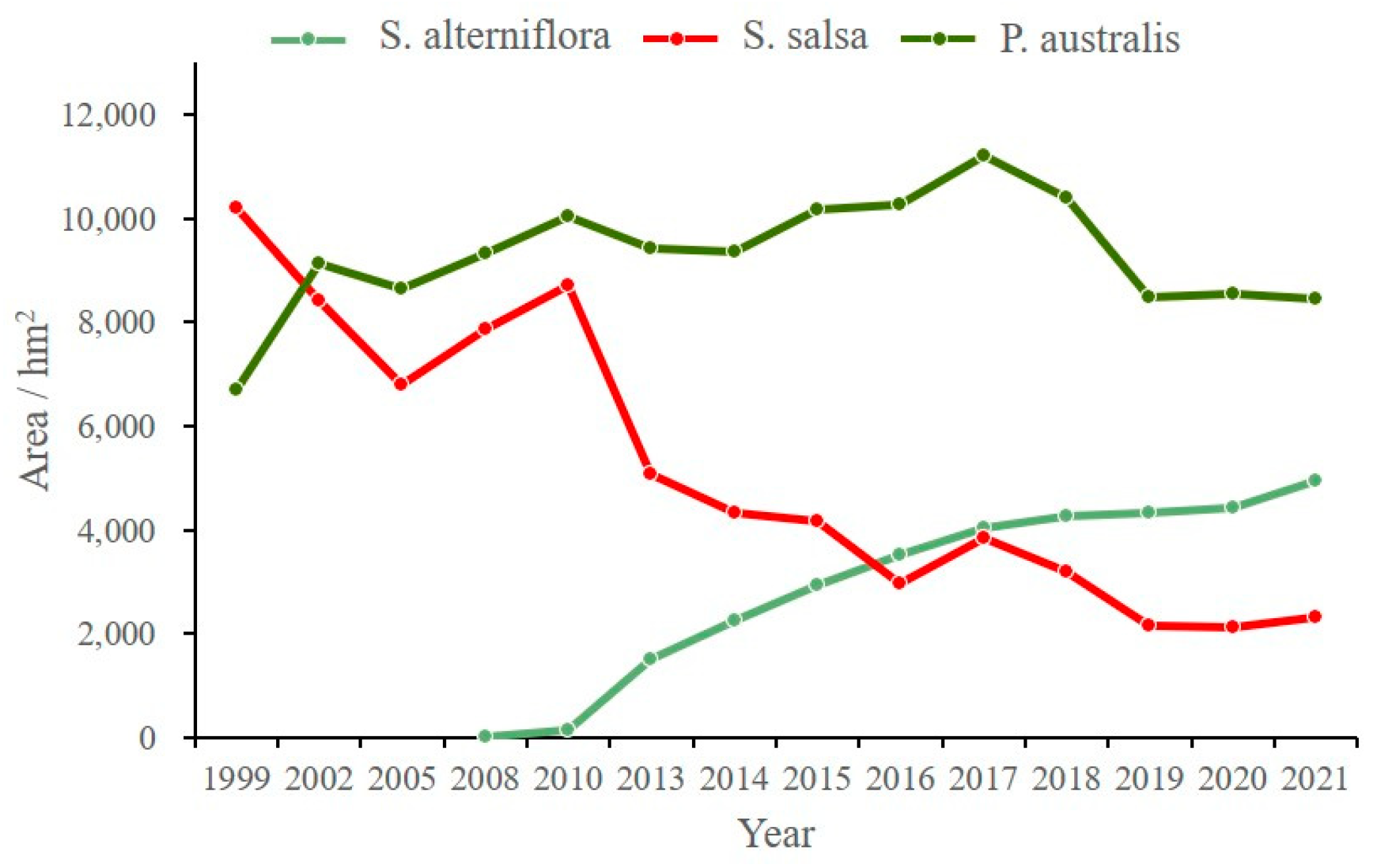
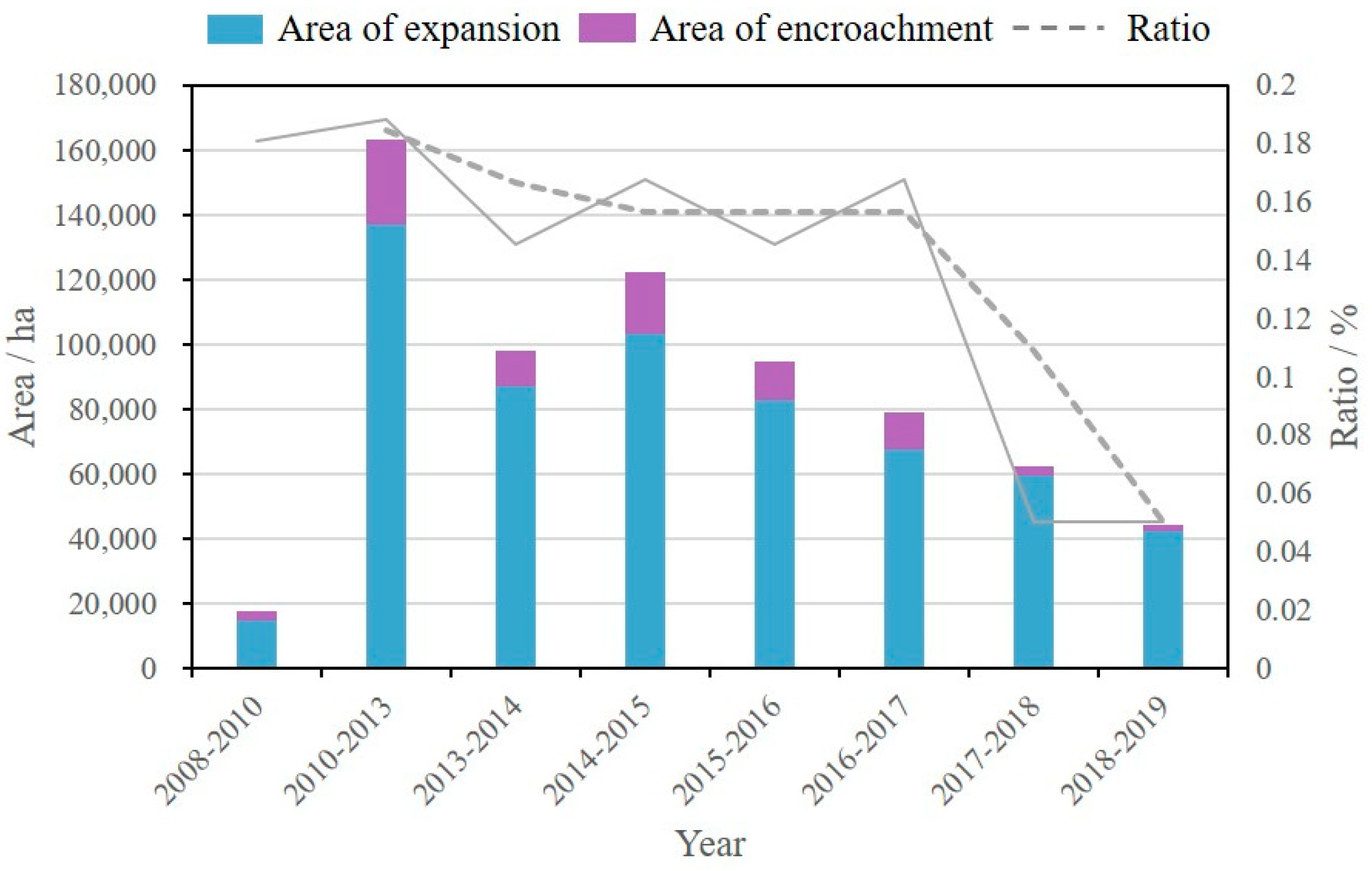
3.2. Evolution of Salt Marsh Vegetation at the Landscape Scale
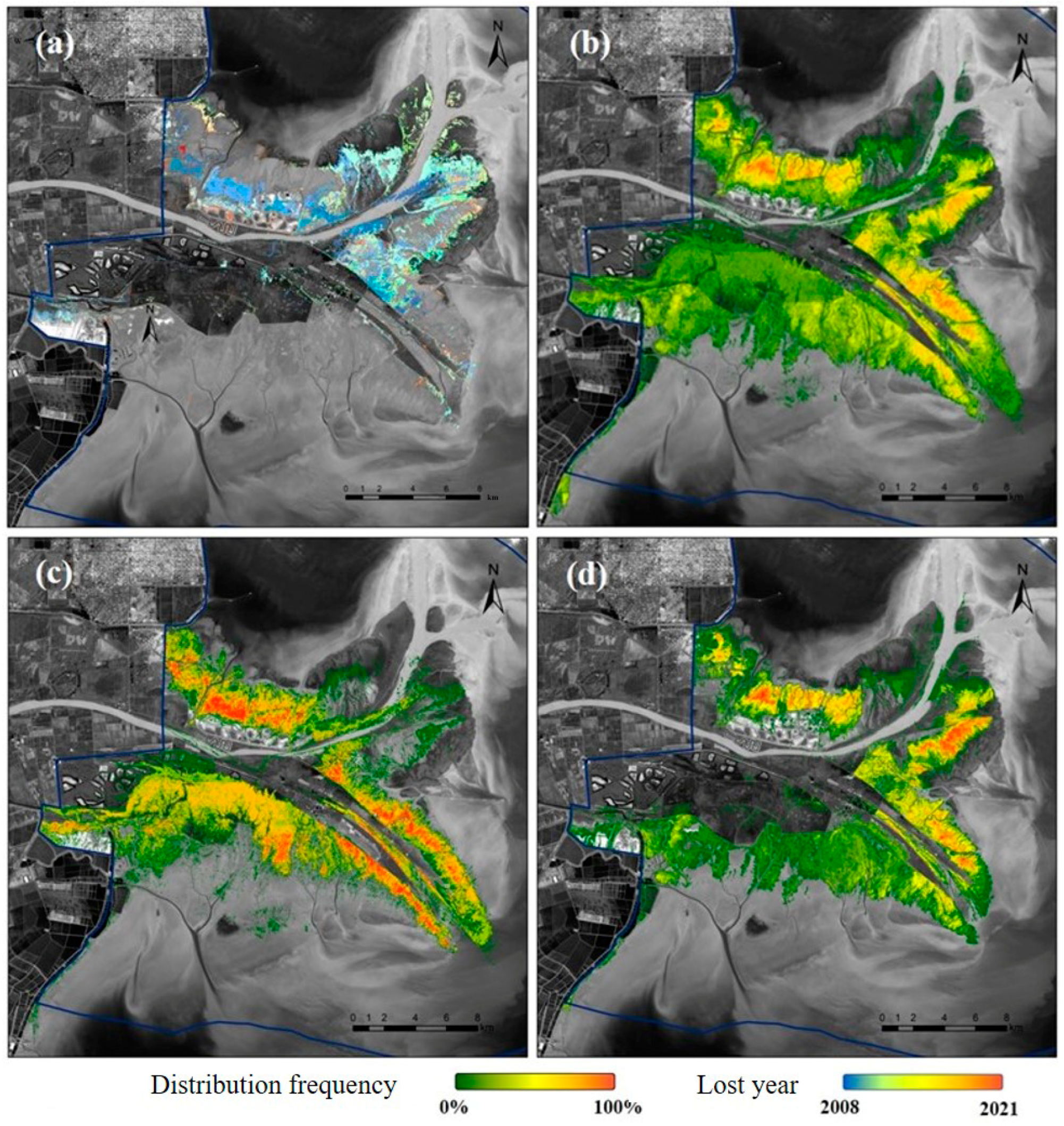
3.3. Evolution of Salt Marsh Vegetation at the Pixel Scale
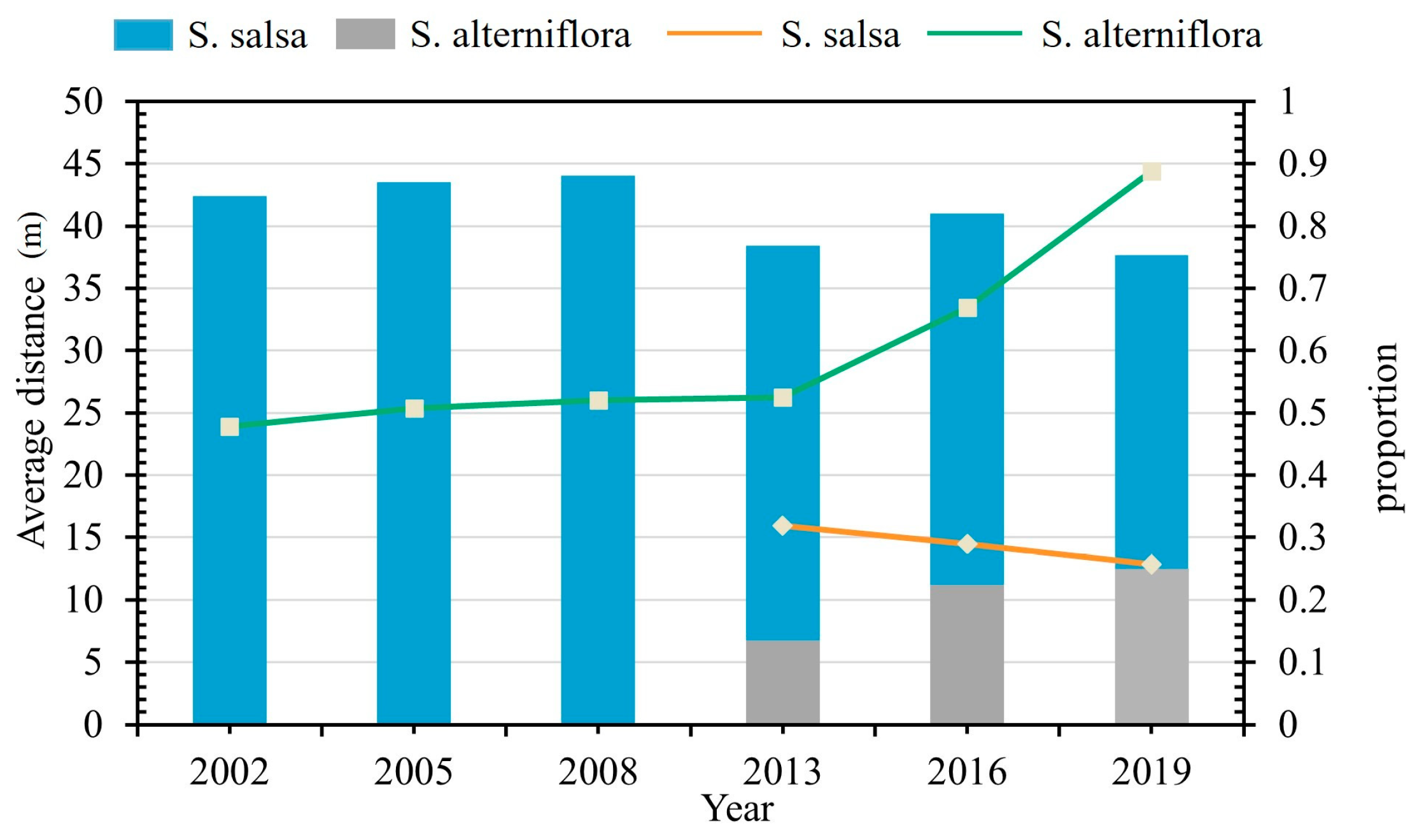
3.3.1. Variation in the State of Distribution of Native Vegetation
3.3.2. Occupation of Native Vegetation by S. alterniflora
3.4. Relationship Between Salt Marsh Vegetation and Tidal Creeks
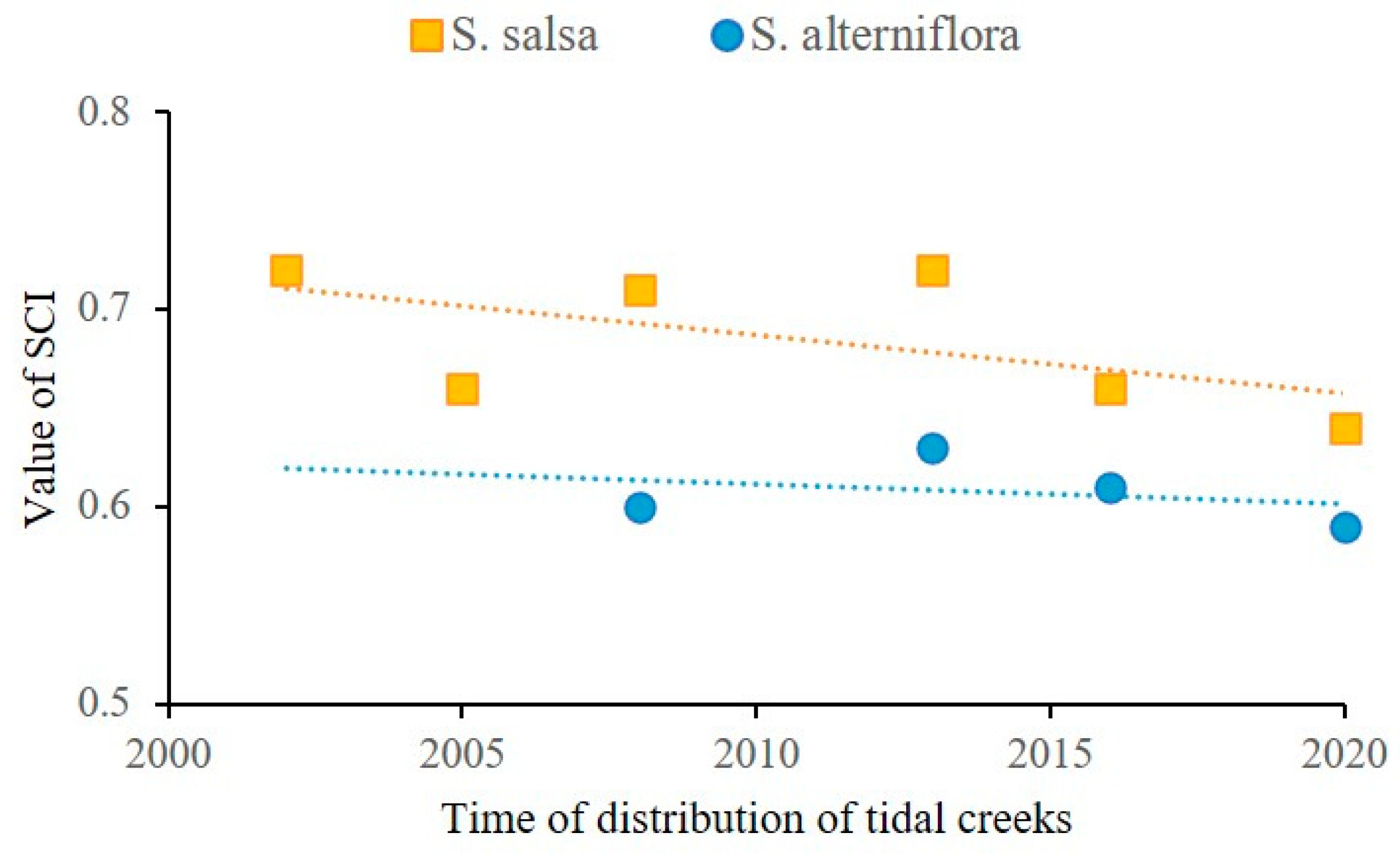
3.4.1. Interaction Studies Based on Coupling Coefficients
3.4.2. Interaction Studies Based on the GeoDetector Model
4. Discussion
4.1. Multiscale Structural Dynamics of Salt Marsh Vegetation
4.2. The Interaction of Salt Marsh Vegetation Coupled with Tidal Creeks
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| S. salsa | Suaeda salsa |
| S. alterniflora | Spartina alterniflora |
| P. australis | Phragmites australis |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Interpreting Signs | Description | Image Features (False Color) |
|---|---|---|
| Suaeda salsa | Scattered across coastal tidal flats; exhibits pinkish-red hues in false-color composites; typically shows low-density growth patterns |  |
| Phragmites australis | Forms continuous patches along riverbanks and waterfronts; distinct red coloration in false-color imagery; demonstrates linear distribution following hydrological features |  |
| Spartina alterniflora | Concentrated in intertidal zones; deep red tones in false-color representations; characterized by high-density canopy coverage |  |
References
- Gong, J.; Li, B.; Liu, C.; Li, P.; Hu, J.; Yang, G. Impact of salinity gradients on nitric oxide emissions and functional microbes in estuarine wetland sediments. Water Res. 2025, 273, 123046. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Chen, M.; Lan, X.; Li, G.; Wang, B.; Sun, D.Y.; Lin, X.B. Shifts of ammonia-oxidation process along salinity gradient in an estuarine wetland. Ecol. Indic. 2022, 145, 109655. [Google Scholar] [CrossRef]
- Wei, H.; Gao, D.; Liu, Y.; Lin, X. Sediment nitrate reduction processes in response to environmental gradients along an urban river-estuary-sea continuum. Sci. Total Environ. 2020, 718, 137185. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Li, B.; Hu, J.; Li, P.; Liu, Q.; Yang, G.; Liu, C. Driving force of tidal pulses on denitrifiers-dominated nitrogen oxide emissions from intertidal wetland sediments. Water Res. 2023, 247, 120770. [Google Scholar] [CrossRef]
- Douglas, E.J.; Lam-Gordillo, O.; Hailes, S.; Lohrer, A.; Cummings, V.J. Characterising intertidal sediment temperature gradients in estuarine systems. Estuar. Coast. Shelf Sci. 2024, 309, 108968. [Google Scholar] [CrossRef]
- Loi, L.T.; Phong, N.T.; Nguyen, L.T. Intertidal bare mudflat and wave attenuation: A case study in the Vietnamese Mekong Delta. Ecol. Eng. 2024, 206, 107320. [Google Scholar] [CrossRef]
- Marani, M.; Da Lio, C.; D’Alpaos, A. Vegetation engineers marsh morphology through multiple competing stable states. Proc. Natl. Acad. Sci. USA 2013, 110, 3259–3263. [Google Scholar] [CrossRef]
- Dzimballa, S.; Willemsen, P.W.J.M.; Kitsikoudis, V.; Borsje, B.W.; Augustijn, D.C.M. Numerical modelling of biogeomorphological processes in salt marsh development: Do short-term vegetation dynamics influence long-term development. Geomorphology 2025, 471, 109534. [Google Scholar] [CrossRef]
- Xie, T.; Cui, B.; Bai, J.; Li, S.; Zhang, S. Rethinking the role of edaphic condition in halophyte vegetation degradation on salt marshes due to coastal defense structure. Phys. Chem. Earth Parts A/B/C 2018, 103, 81–90. [Google Scholar] [CrossRef]
- Murray, N.J.; Phinn, S.R.; Dewitt, M.; Ferrari, R.; Johnston, R.; Lyons, M.B.; Clinton, N.; Thau, D.; Fuller, R. The global distribution and trajectory of tidal flats. Nature 2019, 565, 222–225. [Google Scholar] [CrossRef]
- Song, S.; Wu, Z.; Wang, Y.; Cao, Z.; He, Z.; Su, Y. Mapping the Rapid Decline of the Intertidal Wetlands of China Over the Past Half Century Based on Remote Sensing. Front. Earth Sci. 2020, 8, 16. [Google Scholar] [CrossRef]
- Ouyang, Z.T.; Zhang, M.Q.; Xie, X.; Shen, Q.; Guo, H.; Zhao, B. A comparison of pixel-based and object-oriented approaches to VHR imagery for mapping saltmarsh plants. Ecol. Inform. 2011, 6, 136–146. [Google Scholar] [CrossRef]
- Kumar, L.; Sinha, P. Mapping salt-marsh land-cover vegetation using high-spatial and hyperspectral satellite data to assist wetland inventory. Mapp. Sci. Remote Sens. 2014, 51, 483–497. [Google Scholar] [CrossRef]
- Zeng, J.; Sun, Y.; Cao, P.; Wang, H. A phenology-based vegetation index classification (PVC) algorithm for coastal salt marshes using Landsat 8 images. Int. J. Appl. Earth Obs. Geoinf. 2022, 110, 102776. [Google Scholar] [CrossRef]
- Dong, D.; Wang, C.; Yan, J.; He, Q.; Zeng, J.; Wei, Z. Combing Sentinel-1 and Sentinel-2 image time series for invasive Spartina alterniflora mapping on Google Earth Engine: A case study in Zhangjiang Estuary. J. Appl. Remote Sens. 2020, 14, 044504. [Google Scholar]
- Yi, W.; Wang, N.; Yu, H.; Jiang, Y.; Zhang, D.; Li, X.; Lv, L.; Xie, Z. An enhanced monitoring method for spatio-temporal dynamics of salt marsh vegetation using google earth engine. Estuar. Coast. Shelf Sci. 2024, 298, 108658. [Google Scholar] [CrossRef]
- Wang, D.; Bai, J.; Gu, C.; Gao, W.; Zhang, C.; Gong, Z.; Cui, B. Scale-dependent biogeomorphic feedbacks control the tidal marsh evolution under Spartina alterniflora invasion. Sci. Total Environ. 2021, 776, 146495. [Google Scholar] [CrossRef]
- Wan, D.; Yu, P.; Kong, L.; Zhang, J.; Chen, Y.H.; Zhao, D.; Liu, J. Effects of inland salt marsh wetland degradation on plant community characteristics and soil properties. Ecol. Indic. 2024, 159, 111582. [Google Scholar] [CrossRef]
- Song, J.; Liang, Z.; Li, X.; Wang, X.; Chu, X.J.; Zhao, M.L.; Zhang, X.; Li, P.; Song, W.; Huang, W.; et al. Precipitation changes alter plant dominant species and functional groups by changing soil salinity in a coastal salt marsh. J. Environ. Manag. 2024, 368, 122235. [Google Scholar] [CrossRef]
- Lefor, M.W.; Kennard, W.C.; Civco, D.L. Relationships of salt-marsh plant distributions to tidal levels in Connecticut, USA. Environ. Manag. 1987, 11, 61–68. [Google Scholar] [CrossRef]
- Hickey, D.; Bruce, E. Examining Tidal Inundation and Salt Marsh Vegetation Distribution Patterns using Spatial Analysis (Botany Bay, Australia). J. Coast. Res. 2010, 26, 94–102. [Google Scholar] [CrossRef]
- Mariotti, G.; Fagherazzi, S. A numerical model for the coupled long-term evolution of salt marshes and tidal flats. J. Geophys. Res. Earth Surf. 2010, 115, F01004. [Google Scholar] [CrossRef]
- Best, Ü.S.N.; Van der Wegen, M.; Dijkstra, J.; Willemsen, P.; Borsje, B.W.; Roelvink, D.J.A. Do salt marshes survive sea level rise? Modelling wave action, morphodynamics and vegetation dynamics. Environ. Model. Softw. 2018, 109, 152–166. [Google Scholar] [CrossRef]
- Mariotti, G. Beyond marsh drowning: The many faces of marsh loss (and gain). Adv. Water Resour. 2020, 144 Pt B, 103710. [Google Scholar] [CrossRef]
- Hughes, Z.J. Tidal Channels on Tidal Flats and Marshes. In Principles of Tidal Sedimentology; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Sullivan, J.C.; Torres, R.; Garrett, A.; Blanton, J.; Alexander, C.; Robinson, M.; Moore, T.; Amft, J.; Hayes, D. Complexity in salt marsh circulation for a semienclosed basin. J. Geophys. Res. Earth Surf. 2014, 120, 1973–1989. [Google Scholar] [CrossRef]
- van de Vijsel, R.C.; van Belzen, J.; Boum, T.J.; van der Wal, D.; Borsje, B.W.; Temmerman, S.; Cornacchia, L.; Gourgue, O.; van de Koppel , J. Vegetation controls on channel network complexity in coastal wetlands. Nat. Commun. 2023, 14, 7158. [Google Scholar]
- Mallin, M.A.; Lewitus, A.J. The importance of tidal creek ecosystems. J. Exp. Mar. Biol. Ecol. 2004, 298, 145–149. [Google Scholar] [CrossRef]
- Schwarz, C.; Gourgue, O.; van Belzen, J.; Zhu, Z.; Bouma, T.J.; van de Koppel, J.; Ruessink, G.; Claude, N.; Temmerman, S. Self-organization of a biogeomorphic landscape controlled by plant life-history traits. Nat. Geoence 2018, 11, 672–677. [Google Scholar]
- van Iersel, W.; Straatsma, M.; Addink, E.; Middelkoop, H. Monitoring height and greenness of non-woody floodplain vegetation with UAV time series. ISPRS J. Photogramm. Remote Sens. 2018, 141, 112–123. [Google Scholar]
- Tan, L.; Ge, Z.; Fei, B.; Xie, L.; Li, Y.; Li, S.; Li, X.; Ysebaert, T. The roles of vegetation, tide and sediment in the variability of carbon in the salt marsh dominated tidal creeks. Estuar. Coast. Shelf Sci. 2020, 239, 106752. [Google Scholar] [CrossRef]
- Cui, L.; Ke, Y.; Min, Y.; Han, Y.; Zhang, M.; Zhou, D.M. Effects of tidal creeks on Spartina Alterniflora expansion: A perspective from multi-scale remote sensing. Ecol. Indic. 2024, 160, 111842. [Google Scholar] [CrossRef]
- Gao, Y.; Yi, Y.; Chen, K.; Xie, H. Simulation of suitable habitats for typical vegetation in the Yellow River Estuary based on complex hydrodynamic processes. Ecol. Indic. 2023, 154, 110623. [Google Scholar] [CrossRef]
- Dezfooli, F.P.; Zoej, M.J.V.; Mansourian, A.; Youssefi, F.; Pirasteh, S. GEE-Based Environmental Monitoring and Phenology Correlation Investigation Using Support Vector Regression. Remote Sens. Appl. Soc. Environ. 2024, 37, 101445. [Google Scholar] [CrossRef]
- Deng, X.; Liu, Q.; Deng, Y.; Mahadevan, S. An improved method to construct basic probability assignment based on the confusion matrix for classification problem. Inf. Sci. 2016, 340, 250–261. [Google Scholar] [CrossRef]
- Xue, S.; Ma, B.; Wang, C.; Li, Z. Identifying key landscape pattern indices influencing the NPP: A case study of the upper and middle reaches of the Yellow River. Ecol. Model. 2023, 484, 110457. [Google Scholar] [CrossRef]
- Lan, S.; Zhang, Y.; Gao, T.; Tong, F.; Tian, Z.; Zhang, H.; Li, M.; Mustafa, N.S. UAV remote sensing monitoring of winter wheat tiller number based on vegetation pixel extraction and mixed-features selection. Int. J. Appl. Earth Obs. Geoinf. 2024, 131, 103940. [Google Scholar] [CrossRef]
- Wang, X.; Yang, J. A privacy image encryption algorithm based on piecewise coupled map lattice with multi dynamic coupling coefficient. Inf. Sci. 2021, 569, 217–240. [Google Scholar] [CrossRef]
- Aschonitis, V.G.; Gaglio, M.; Castaldelli, G.; Fano, E.A. Criticism on elasticity-sensitivity coefficient for assessing the robustness and sensitivity of ecosystem services values. Ecosyst. Serv. 2016, 20, 66–68. [Google Scholar] [CrossRef]
- Hou, W.; Liang, S.; Sun, Z.; Ma, Q.; Hu, X.; Zhang, R. Depositional dynamics and vegetation succession in self-organizing processes of deltaic marshes. Sci. Total Environ. 2024, 912, 169402. [Google Scholar] [CrossRef]
- Zhu, Y.; Ling, G.H.T. Driving forces and prediction of urban open spaces morphology: The case of Shanghai, China using geodetector and CA-Markov model. Ecol. Inform. 2024, 82, 102763. [Google Scholar] [CrossRef]
- Kong, D.; Miao, C.; Zheng, H.; Gong, J. Dynamic evolution characteristics of the Yellow River Delta in response to estuary diversion and a water-sediment regulation scheme. J. Hydrol. 2023, 627, 130447. [Google Scholar] [CrossRef]
- Wang, H.; Zhou, Y.; Wu, J.; Wang, C.X.; Zhang, R.; Xiong, X.; Xu, C. Human activities dominate a staged degradation pattern of coastal tidal wetlands in Jiangsu province, China. Ecol. Indic. 2023, 154, 110579. [Google Scholar] [CrossRef]
- Cui, B.; He, Q.; Zhao, X. Ecological thresholds of Suaeda salsa to the environmental gradients of water table depth and soil salinity. Acta Ecol. Sin. 2008, 28, 1408–1418. [Google Scholar] [CrossRef]
- Ma, X.; Qiu, D.; Wang, F.; Jiang, X.; Sui, H.C.; Liu, Z.; Yu, S.; Ning, Z.; Gao, F.; Bai, J.H.; et al. Tolerance between non-resource stress and an invader determines competition intensity and importance in an invaded estuary. Sci. Total Environ. 2020, 724, 138225. [Google Scholar] [CrossRef] [PubMed]
- Ning, Z.; Li, D.; Chen, C.; Xie, C.J.; Chen, G.; Xie, T.; Wang, Q.; Bai, J.H.; Cui, B.S. The importance of structural and functional characteristics of tidal channels to smooth cordgrass invasion in the Yellow River Delta, China: Implications for coastal wetland management. J. Environ. Manag. 2023, 342, 118297. [Google Scholar] [CrossRef]
- Zhang, C.; Gong, Z.; Qiu, H.; Zhang, Y.; Zhou, D. Mapping typical salt-marsh species in the yellow river delta wetland supported by temporal-spatial-spectral multidimensional features. Sci. Total Environ. 2021, 783, 147061. [Google Scholar] [CrossRef]



| Feature | Source Images | Abbreviation | Description | Time Scale |
|---|---|---|---|---|
| Spectral features | Sentinel-2, Landsat | B | Spectral reflectance | Landsat: growing seasons in 2000–2015 |
| NDWI | (G − NIR)/(G + NIR) | |||
| NDVI | (NIR − R)/(NIR + R) | |||
| RDVI | Sentinel-2: growing seasons in 2016–2021 | |||
| DVI | NIR − R | |||
| MASVI | ||||
| RVI | NIR/R | |||
| Sentinel-2 | NDVIre1 | (B8 − B5)/(B8 + B5) | ||
| NDVIre2 | (B8 − B6)/(B8 + B6) | |||
| NDVIre3 | (B8 − B7)/(B8 + B7) | |||
| Spatial features | Sentinel-2, Landsat | ASM | angular second-order moment | Growing seasons in 2000–2021 |
| Cor | Correlation | |||
| Con | Contrast | |||
| Ent | entropy | |||
| Var | variance |
| LPI | Formula | Description |
|---|---|---|
| Proportion of landscape area occupied by patches | Area characterizing patch type as a percentage of total landscape area | |
| Patch density | Edge length between a landscape element patch and its adjacent heterogeneous patches | |
| Fragmentation index | Changes in the shape of landscape types | |
| Edge density | Complexity of patch shapes and patch edges in the landscape | |
| Patch shape index | Extent to which topographic and hydrologic conditions affect this type of patch | |
| Subdimensional index | Aggregated or dispersed state of patches in the landscape | |
| Scattering and juxtaposition index | Connectivity within landscape type patches | |
| Cohesion index | Degree of dispersion and aggregation within the same landscape type | |
| Aggregation index | Degree of landscape fragmentation and spatial heterogeneity |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Yan, J.; Bian, Z.; Gong, Z.; Zhu, D. Multiscale Structural Patterns of Intertidal Salt Marsh Vegetation in Estuarine Wetlands and Its Interactions with Tidal Creeks. J. Mar. Sci. Eng. 2025, 13, 946. https://doi.org/10.3390/jmse13050946
Hu J, Yan J, Bian Z, Gong Z, Zhu D. Multiscale Structural Patterns of Intertidal Salt Marsh Vegetation in Estuarine Wetlands and Its Interactions with Tidal Creeks. Journal of Marine Science and Engineering. 2025; 13(5):946. https://doi.org/10.3390/jmse13050946
Chicago/Turabian StyleHu, Jianfang, Jiapan Yan, Zhenbang Bian, Zhaoning Gong, and Duowen Zhu. 2025. "Multiscale Structural Patterns of Intertidal Salt Marsh Vegetation in Estuarine Wetlands and Its Interactions with Tidal Creeks" Journal of Marine Science and Engineering 13, no. 5: 946. https://doi.org/10.3390/jmse13050946
APA StyleHu, J., Yan, J., Bian, Z., Gong, Z., & Zhu, D. (2025). Multiscale Structural Patterns of Intertidal Salt Marsh Vegetation in Estuarine Wetlands and Its Interactions with Tidal Creeks. Journal of Marine Science and Engineering, 13(5), 946. https://doi.org/10.3390/jmse13050946