Abstract
The tropical coral Siderasterea radians is typically found in mangrove areas, seagrass beds, and environments tolerated by few other scleractinians in the Caribbean and South Florida. Siderastrea radians experience bleaching stress in Buttonwood Sound in Florida Bay during April–May, a time when the seawater is below bleaching temperatures. Stress first appears as a pink coloration due to the presence of fluorescent pigments protecting the coral tissue and algal symbionts from exposure to excessive sunlight/UV light. Surveys of S. radians in areas frequented by parrotfish showed that up to 38.5% of the corals were pink, with about a quarter of all the corals having bite marks. In areas not frequented by parrotfish, only 3.2% of the corals were pink, and none had bite marks. A seasonal analysis in parrotfish-frequented seawater showed S. radians contained among the lowest tissue biomass and density of symbiotic algae during April–May 2010–2017, often looking pink. It is hypothesized that seawater temperature is not the cause of bleaching in Buttonwood Sound during April–May but rather predation by the rainbow parrotfish Scarus guacamaia.
1. Introduction
The coral Siderastrea radians is commonly found in backreef areas, such as seagrasses and mangroves, and is seen in shallow environments tolerated by few other scleractinians in the Florida Keys, the Bahamas, or in the Caribbean [1,2]. It is typically less than 10 cm in diameter, and the polyps are retracted into corallites during the daytime but come out at night to feed on demersal plankton [1]. S. radians tolerate high temperatures and a wide range of salinities [2,3]. It occurs in shallow water, usually 0–3 m deep, in areas where hard substrates are exposed [4]. Brooded planulae larvae settle on the calcium carbonate benthos, infrequently on a small rock or shell that can be overturned in windy conditions. The thick-tissued colonies of Siderastrea sp. are resistant to bleaching, becoming a pinkish color when they do bleach [5]. Siderastrea radians in the Florida Keys are found with Symbiodiniaceae of the genus Breviolum sp. (B5a) [6]; there have been no other species of symbiont dinoflagellates described in Florida [7,8].
Siderastrea stellata is a sister species found in Brazil that loses symbionts (bleaches) in March–June, which is during late summer, often turning a pink-blue color [9]. This phenomenon is a pigmentation response of the host to a variety of stressors, and the pink-blue coloration occurs due to the presence of pocilloporin fluorescent pigments in the underlying tissue, especially when the Symbiodiniaceae are less dense in the host [10,11,12]. These animal pigments act as a photo-protectant to animal tissues and the remaining symbionts therein, reducing exposure to excessive sunlight and especially UV light. Once symbiont densities decrease, the pocilloporins become visible, and mortality of part or all of the colony may occur. When stressful conditions are reduced, the symbionts grow back, and the pink-blue colonies return to a normal brown color.
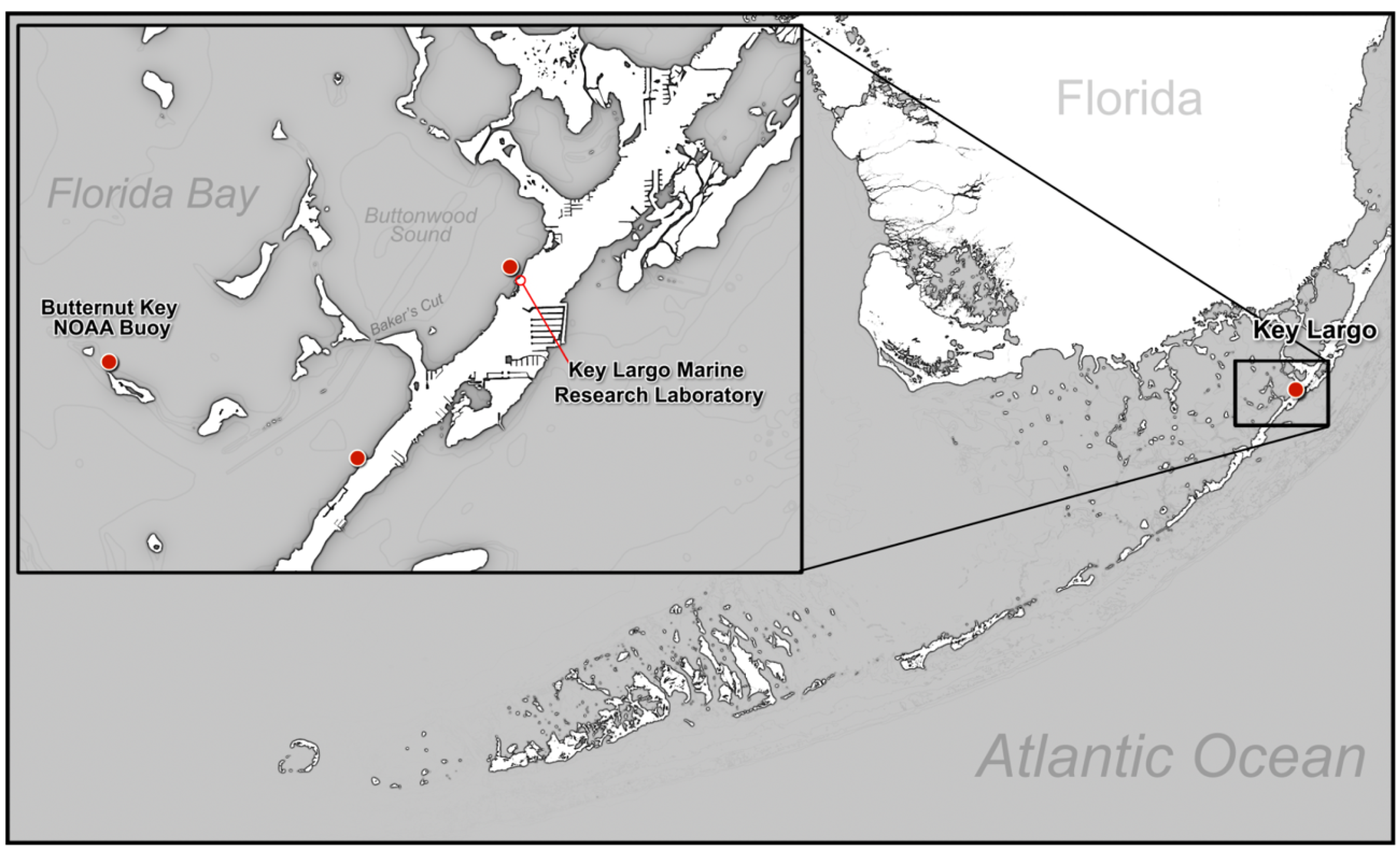
Many of the S. radians off the Key Largo Marine Research Laboratory (KLMRL) in Buttonwood Sound in Florida Bay (Figure 1) also appear pink seasonally (Figure 2B,C), but usually in the spring (April–May), several months before the warmest time of year (August–September). Many species of corals in Florida, the Bahamas, or the Caribbean experience warm-water bleaching at the end of summer (August–September), not during the spring [13]. It was first thought that the pink color of S. radians that arises in April–May was associated with the colonies becoming unattached and rolling on the bottom when the wind grew stronger, which is common in the normally calm waters of Florida Bay as cold fronts blow in periodically from the north between November–April.
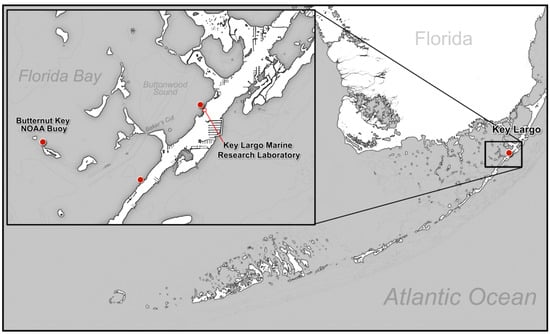
Figure 1.
Map of Buttonwood Sound, Florida Bay. Temperatures were taken from the Butternut Key NOAA Buoy (red dot). The sites of Surveys 1, 2, and 3 were conducted off Key Largo Marine Research Laboratory (red dot); Survey 4 was taken from a site south of Baker Cut (red dot).

Figure 2.
Normal coloration (brown) of Siderastrea radians (A), pink-colored coral (B), pink coral with bite marks on the coral, plus partial mortality on the right and top sides (C), and a rainbow parrotfish Scarus guacamaia in Buttonwood Sound (D). Bar = 1 cm.
Another disturbance is by fish-predation. Most fish in Florida Bay are not corallivores but eat smaller invertebrates or are herbivores. However, for at least 15 years, a population of the rainbow parrotfish Scarus guacamaia has made a seasonal home in the waters off Buttonwood Sound, taking up residence sometime in April and disappearing in September–October each year (Figure 2D). Scarus guacamaia is a coral reef herbivore with a presence in the northeastern part of Florida Bay, requiring both mangrove and coral reef habitats to complete its life cycle [14]. Scarus guacamaia is known to eat corals [15] in the Florida Keys (Figure 2C, [16]), effectively grinding coral with their teeth [17], which are fused to form strong beak-like jaws [18]. Parrotfish have been documented to take 8.1% of their bites on coral in Brazil [19]. The stoplight parrotfish Sparisoma viride has been observed to eat the coral Porites divaricata when they were experimentally moved from seagrass habitats and placed on the reef [20]. Such corallivory has been considered one of the determinants of the distribution of the corals’ Porites astreoides, P. furcata, and P. divaricata on Caribbean reefs [20,21,22].
The hypothesis of why a significant portion of S. radians in Buttonwood Sound (Florida Bay) appear pink and bleached, with a low density of symbionts and tissue biomass in April–May, is not because of high water temperatures but rather predation by the rainbow parrotfish Scarus guacamaia.
2. Materials and Methods
Water temperatures were monitored every hour from a NOAA buoy (BNKF1) off Butternut Key, on the outer mangrove islands near Baker Cut, just over 2 miles south of Buttonwood Sound. Temperatures greater than 32 °C, which are considered high enough to bleach corals, were plotted for the years 2010–2017.
Siderastrea radians were observed in May 2015 along a narrow band parallel to the shore (1–2.5 m in depth) in Buttonwood Sound (25.10217, −80.43868). Different sizes were measured (maximum diameter) to eliminate the ontogenetic differences between partially to wholly pink (the first ones seen, n = 12) and brown colonies (the first ones seen, n = 12). Following this, the first survey was conducted in May 2015 to assess the color and proportion of corals attached to the bottom. Based on these results, the following two surveys were conducted in two separate sections of Buttonwood Sound in May 2015. The color of the coral, the proportion attached to the substrate, the presence of algae growing on the skeleton (denoting partial mortality), and bite marks were noted. The last survey was conducted in the bay just to the south of Buttonwood Sound and Baker Cut during May 2015 (25.07596, −80.46416); the color of the coral, proportion of corals attached, presence of algae growing on the skeletons (denoting partial mortality), and whether the coral had any bite marks were observed.
Siderastrea radians were monitored seasonally in Buttonwood Sound (3–4 times per year) between August 2010 and June 2017. The ash-free dry weight (AFDW = biomass) of the tissue was determined by sampling six heads of S. radians each date, water-piking (freshwater) an area of the coral, freeze-drying the slurry, weighing an aliquot, burning the aliquots in a muffle furnace at 500 °C overnight to obtain the ash weight, subtracting the ash weight from the dry weight to obtain the AFDW, and finally dividing by the area water-picked [23]. The densities of symbiotic dinoflagellates were calculated from the replicate (n = 10) hemocytometer counts of homogenized water-picked slurry (in salt water) and surface area determinations by correlations between the weight and surface area of aluminum foil [23].
3. Results and Discussion
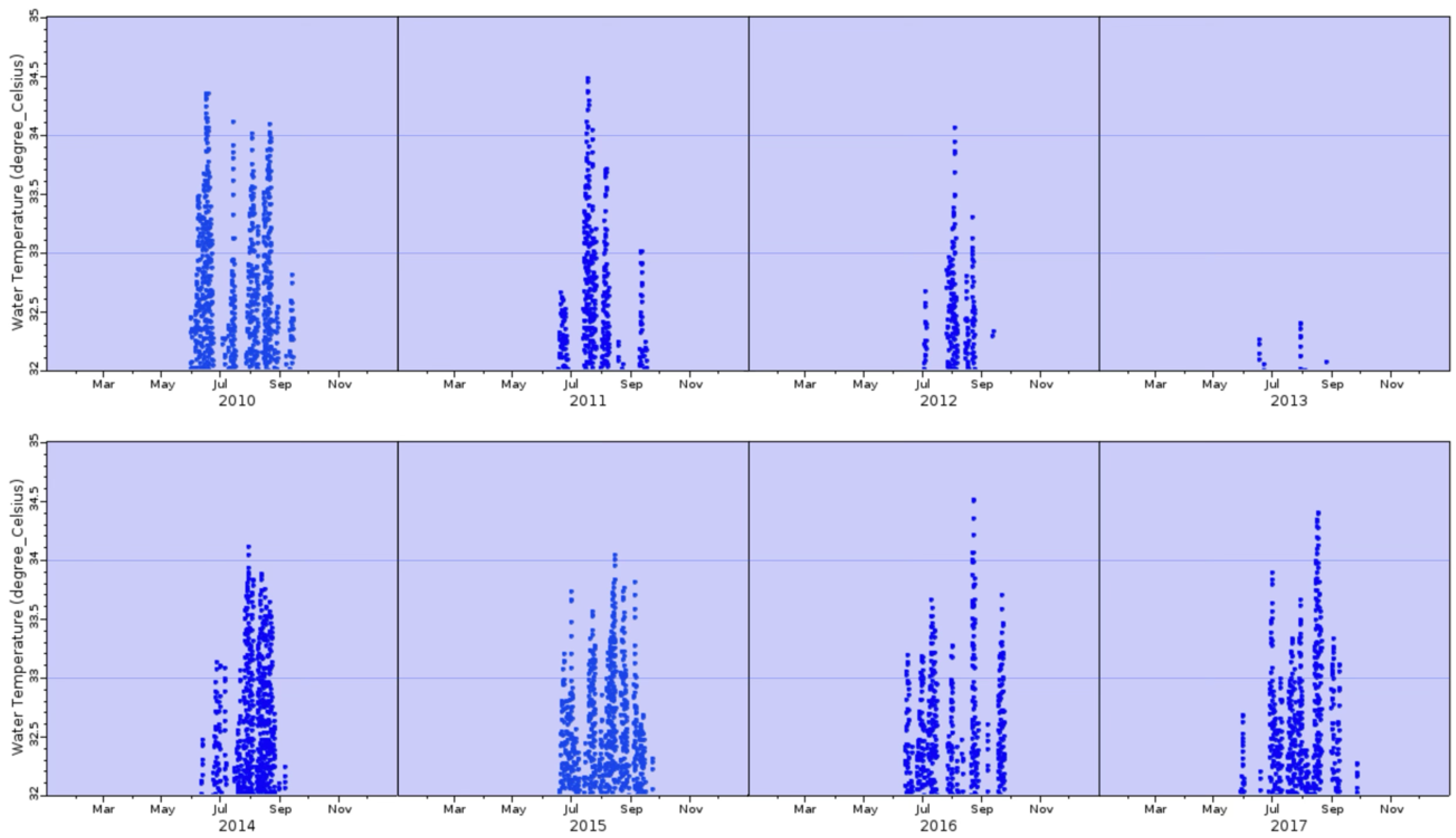
The water temperatures were monitored hourly from 2010 and 2017 from a NOAA buoy off Butternut Key, just over 2 miles south of Buttonwood Sound. Temperatures greater than 32 °C (denoting “bleaching” temperatures) were plotted every hour between 2010 and 2017 (Figure 3). There were only 20 h of points observed in the last 3 days of May 2017; no other periods of high temperature were observed in April–May (Figure 3).
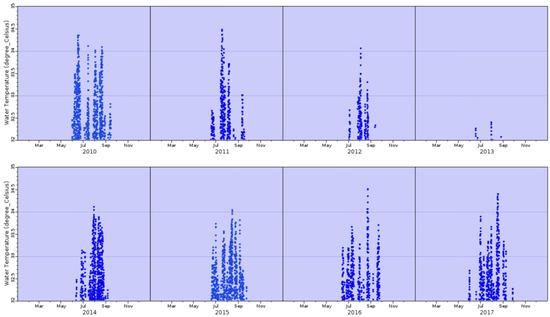
Figure 3.
Bleaching temperatures (above 32 °C) of seawater off Butternut Key, Florida Bay, for the years 2010–2017. Data were taken every hour.
The diameters of the first 12 brown (all brown) and 12 pink (pink visible) Siderastrea radians (Figure 2A–C) encountered in Buttonwood Sound 2015 were compared to see if there were any ontogenetic changes that might be a factor in coral color. The mean diameter of the brown corals (7.41 ± 3.99 cm, s.d., n = 12) was not significantly different from the pink corals (7.66 ± 2.71 cm, s.d., n = 12), suggesting that no developmental growth characteristics, nor maturation processes were involved in the coloration of the corals.
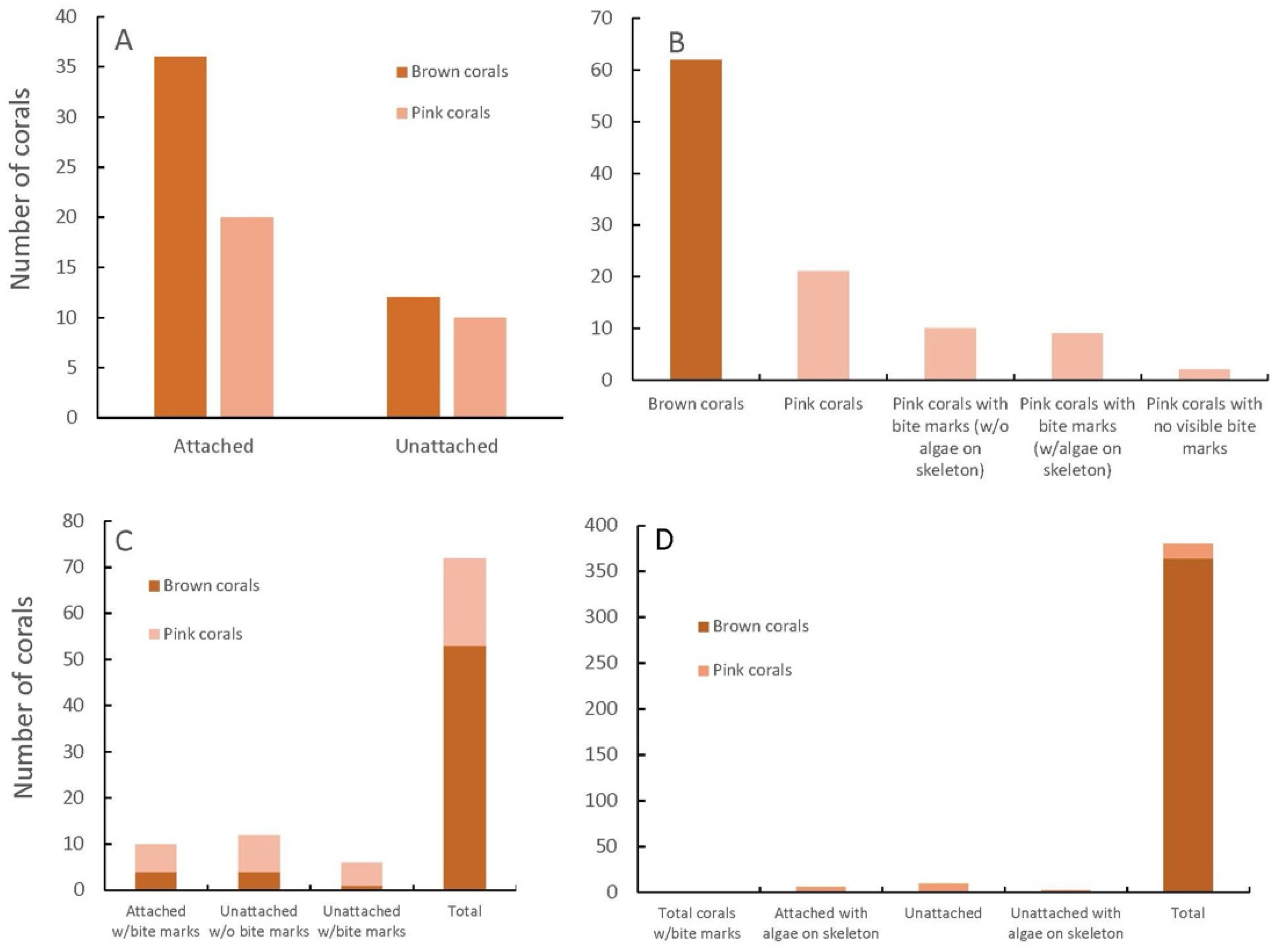
The first survey (Figure 4A) was conducted on S. radians in Buttonwood Sound in 2015 to see what color the corals were when they were attached to the bottom and whether they turned pink when they became unattached. A total of 48 corals were brown (61.5%), with 12 unattached, whereas 30 corals were pink (38.5%), with 10 unattached (Figure 4A). Since there were 20 pink corals that were attached, it was decided that being unattached and rolling around on the bottom could not be the main reason why the corals were pink.
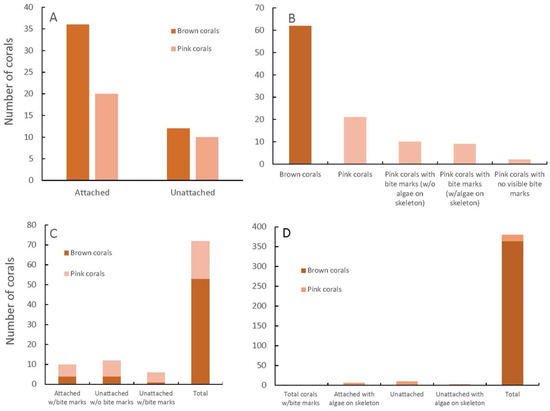
Figure 4.
Attached and non-attached pink and brown corals (Siderastrea radians) (A), pink and brown corals, pink with/without bite marks, and algae on the skeleton (=dead portions) (B), attached and unattached brown or pink corals with and without bite marks (C), attached or unattached brown or pink corals with or without algae on the skeleton, all with no bite marks (D).
A second survey (Figure 4B) was conducted on brown and pink S. radians from Buttonwood Sound in 2015, noting the bite marks of parrotfish and whether the corals had any dead areas (=algae growing) on them (Figure 2C). This survey found 62 brown coral colonies, all with no bites (74.7%), and 21 pink coral colonies (25.3%), with 19 of the pink colonies with bite marks and 9 of them with algae growing on the skeleton (Figure 4B). This survey pointed clearly towards parrotfish bites being correlated with the pink color and partial mortality of the colony.
The third survey (Figure 4C) was conducted the next day, just north of the second survey, and bite marks in attached and unattached brown and pink S. radians in Buttonwood Sound were observed. There were 53 colonies (73.6%) that were brown, with 4 attached and 1 unattached with bite marks, whereas 19 colonies (26.4%) were pink, with 6 attached and 3 unattached that had bite marks (Figure 4C). This survey suggested that parrotfish either did not have a preference for attached or unattached corals, and/or their nipping at the corals perhaps dislodged the coral from the substrate. The 10 unattached pink corals without bite marks could have rolled around on the substrate, thus giving rise to the pink color.
A fourth survey (Figure 4D) was conducted in 2015 on S. radians in Florida Bay just south of Buttonwood Sound (south of Baker Cut). No parrotfish have ever been seen during several surveys conducted in 2010–2023. All told, 364 brown (96.8%) and 12 pink (3.2%) corals were found, with none of these having bite marks (Figure 4D). Ten (10) of the pink corals were unattached (Figure 4D), suggesting that rolling around on the substrate is a form of stress and a way that S. radians can obtain the pink color. It was not possible to determine whether a coral was knocked free of its attachment by fish.
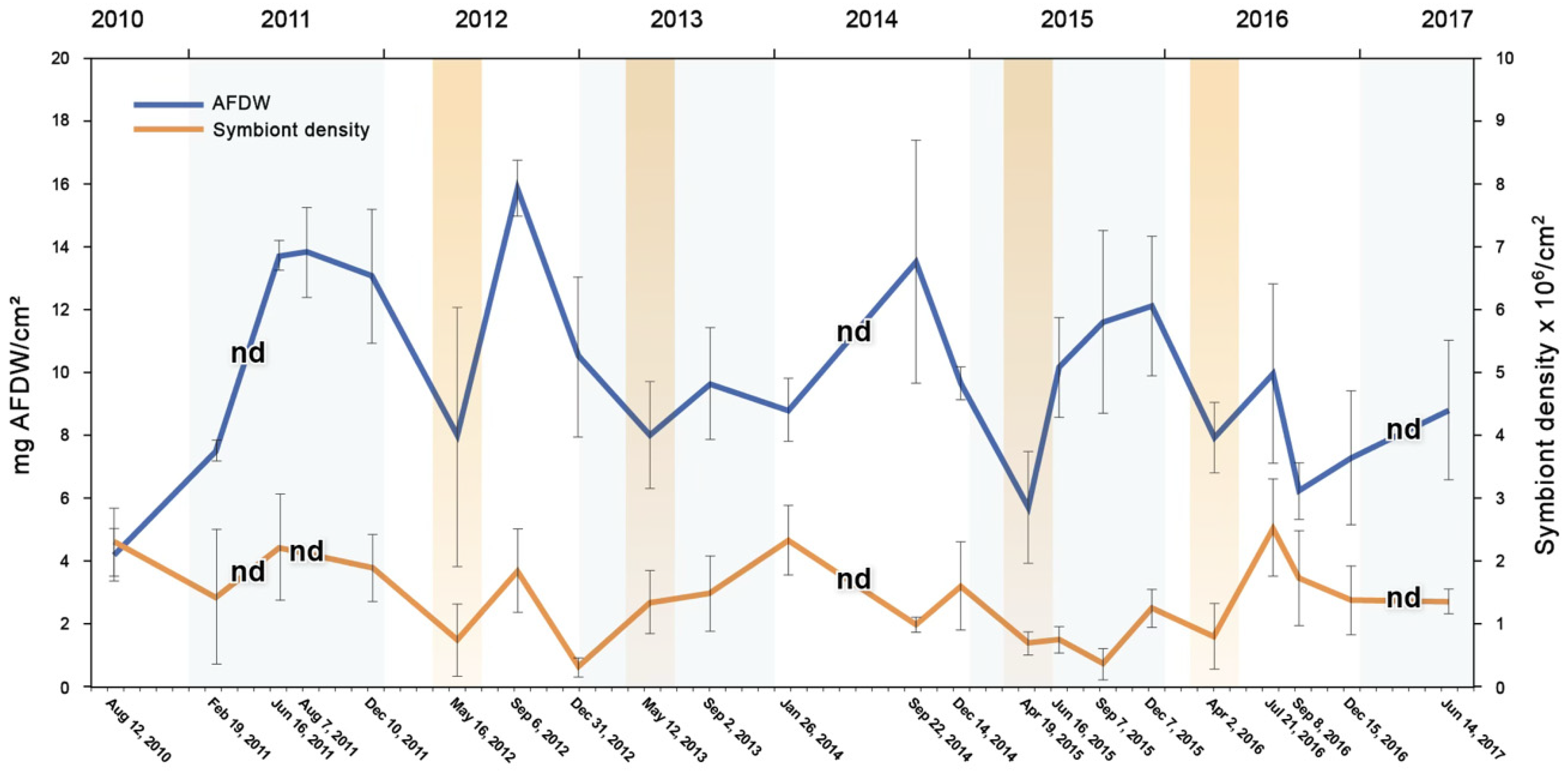
The seasonal analysis of the symbiont densities and biomass (=AFDW) of S. radians was determined from August 2010 through June 2017 (Figure 5). There were four years when the April–May periods were sampled (colored bars) and three years when there were no data taken during these months (nd). The AFDW and symbiont density would normally increase during the spring [23] but had decreased those four years, suggesting that something was stressing the S. radians.
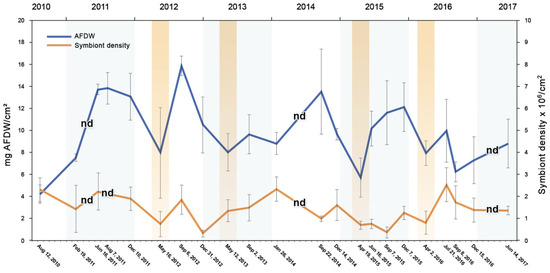
Figure 5.
Biomass (mg ash-free dry weight/cm2, blue) and density of symbiotic algae (millions/cm2, orange) in the coral Siderastrea radians between August 2010 and June 2017 (dates and year are listed). Colored bars = samples taken in April–May, and nd = no data taken in April–May (and 7 August 2011 symbiont density).
The pink color of the S. radians indicates bleaching, which can be seen in the biomass and density of the symbionts during the April–May seasonal samples (Figure 5). The seasonal analyses showed that the lowest AFDW and density of the symbionts tended to be in the four years when the samples were taken in April–May, which is not when the seawater temperatures are the highest [13]. Though the analysis does not show cause and effect, a maximum of a third to a half of the biomass and about half of the symbiont density disappeared in April–May (Figure 5). It is not known why S. radians do not look pink during June–July, though warmer water during the summer (Figure 3) may be associated with greater growth rates, masking any potential predation by the parrotfish (Figure 2C,D). The only time S. radians showed a warm-water bleaching response (low symbiont density) between 2010–2017 was between December 2012 and September 2015 (Figure 5).
Coralivorous parrotfish use their sharp parrot-like teeth to bite corals, presumably to access the symbionts and/or coral tissue. Parrotfish caused >13% of Porites astreoides to exhibit partial or total mortality in Belize, some with >50% tissue loss [22]. Corals have one or more potential barriers that inhibit predation by parrotfish, such as potent nematocysts (stinging organelle) or recessed polyps [24]. S. radians apparently have ineffective nematocysts, and the skeletal portion of the polyps can be penetrated by the parrotfish. Scarus guacamaia is known to eat sister species Siderastrea siderea on the outer reefs of the Florida Keys [16] and has been observed to pick on corals and algae in Buttonwood Sound in this study. It is thought that the earliest populations of S. guacamaia in Buttonwood Sound were present when this study began, though not in the enormous (>50) numbers that have been seen in the past several years. The data presented here suggest that this parrotfish could feed on up to 22.9% of the S. radians in Buttonwood Sound (Table 1, Figure 4B), with 12.5% (Figure 4B) showing partial (Figure 2C) to near-total mortality.

Table 1.
The percent of pink corals, with and without bite marks.
Siderastrea radians are common in seagrass, mangroves, and shallow disturbed sites in the Florida Keys. It is quite possible that predation of this species, especially by parrotfish, is responsible for their distribution. Littler et al. [21], Miller and Hay [20], and Rotjan and Lewis [22] have documented parrotfish predation as a determinant for the distribution of Porites astreoides, P. furcata, and P. divaricata.
4. Conclusions
Predation by the rainbow parrotfish Scarus guacamaia is thought to be responsible for much of the seasonal stress on the coral S. radians in Buttonwood Sound, resulting in low symbiont density and tissue biomass during April–May. The coral produces pink coloration during these months due to lower symbiont density. Surveys in areas frequented by parrotfish showed that up to 38.5% of the corals were pink, with 23% demonstrating bite marks. In areas not frequented by parrotfish, only 3.2% of the corals were pink, and none had bite marks. The seasonal analysis in parrotfish-frequented water showed S. radians contained among the lowest biomass and density of symbiotic algae during April–May 2010–2017, often looking pink.
Funding
This research received no external funding.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Acknowledgments
Melantha Becol and Colton Fowlkes contributed data on the S. radians while at the Key Largo Marine Research Laboratory (KLMRL) on Buttonwood Sound. Nathan Jess improved the figures. Paul and Cathryn Canty took the picture of the rainbow parrotfish at KLMRL. The Florida Keys National Marine Sanctuary kindly provided permits (2008-038-A1, 2011-093, 2014-117) for the collection of S. radians. The manuscript benefitted from reading and comments by Henry Laird and three reviewers.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Lirman, D.; Orlando, B.; Macia, S.; Manzello, D.; Kaufman, L.; Biber, P.; Hones, T. Coral communities of Biscayne Bay, Florida and adjacent offshore areas: Diversity, abundance, distribution, and environmental correlates. Aquatic. Conserv. Mar. Freshw. Ecosyst. 2003, 13, 121–135. [Google Scholar] [CrossRef]
- Lirman, D.; Manzello, D. Patterns of resistance and resilience of the stress-tolerant coral Siderastrea radians (Pallas) to sub-optimal salinity and sediment burial. JEMBE 2009, 369, 72–79. [Google Scholar] [CrossRef]
- Wagner, D.E.; Kramer, P.; van Woesik, R. Species composition, habitat, and water quality influence coral bleaching in southern Florida. J. Exp. Mar. Biol. Ecol. 2010, 408, 65–78. [Google Scholar] [CrossRef]
- Fenner, D.P. Species distinctions among several Caribbean stony corals. Bull. Mar. Sci. 1993, 53, 1099–1116. [Google Scholar]
- Sassi, R.; Sassi, C.F.C.; Gorlach-Lira, K.; Fitt, W.K. Pigmentation changes in Siderastrea spp. during bleaching events in the costal reefs of northeastern Brazil. Lat. Am. J. Aquat. Res. 2015, 43, 176–185. [Google Scholar] [CrossRef]
- LaJuenesse, T.C.; Parkinson, J.V.; Gabrielson, P.W.; Jeong, H.J.; Reimer, J.D.; Voolstra, C.R.; Santos, S.R. Systematic revision of Symbiodiniaceae highlights the antiquity and diversity of coral endosymbionts. Curr. Biol. 2018, 28, 2570–2580. [Google Scholar] [CrossRef]
- Thornhill, D.J.; Fitt, W.K.; Schmidt, G.W. Highly stable symbioses among western Atlantic brooding corals. Coral Reefs 2006, 25, 515–519. [Google Scholar] [CrossRef]
- Kemp, D.W.; Kempf, S.C.; Fitt, W.K. The weight of it all: Symbiotic dinoflagellates in Caribbean reef-building corals. Mar. Biol. 2020, 167, 121–126. [Google Scholar] [CrossRef]
- Sassi, C.F.C.; Sassi, R.; Gorlach-Lira, K.; De Cassia, R.; De Lima, P. Pigmentation patterns of Siderastrea stellata Verrill, 1868 (Cnidaria, Scleractinia) from coastal reefs in Northeastern Brazil and its relation with zooxanthellae and other microsymbionts. Pan-Am. J. Aquatic. Sci. 2014, 9, 207–222. [Google Scholar]
- Dove, S.G.; Hoegh-Guldberg, O.; Ranganathan, S. Major colour patterns of reef-building corals are due to a family of GFP-like proteins. Coral Reefs 2001, 19, 197–204. [Google Scholar] [CrossRef]
- Bongiorni, L.; Rinkevich, B. The pink-blue spot syndrome in Acropora eurystoma (Eilat, Red Sea): A possible marker of stress? Zoology 2005, 108, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Casareto, B.E.; Yucharoen, M.; Dohra, H.; Suzuki, Y. Coesistence of nonfluorescent chromaoproteins and fluorescent proteins in massive Porites spp. corals manifesting a pink pigmmentation response. Front. Physiol. 2024, 15, 1339907. [Google Scholar] [CrossRef] [PubMed]
- Spalding, M.D.; Brown, B.E. Warm-water coral reefs and climate change. Science 2015, 350, 769–771. [Google Scholar] [CrossRef] [PubMed]
- Machemer, E.G.P.; Walter, J.F., III; Serafy, J.E.; Kerstetter, D.W. Importance of mangrove shorelines for rainbow parrotfish Scarus guacamaia: Habitat suitability modeling in a subtropical bay. Aquat Biol. 2012, 15, 67–98. [Google Scholar] [CrossRef]
- Frydl, P. The effect of parrotfish (Scaridae) on coral in Barbados, WI. Int. Revue Ges. Hydrobiol. 1979, 64, 737–748. [Google Scholar]
- Burkepile, D.E.; Adam, T.C.; Roycroft, M.; Ladd, M.C.; Munsterman, K.S.; Ruttenberg, B.I. Species-specific patterns in corallivory and spongivory among Caribbean parrotfishes. Coral Reefs 2019, 38, 417–423. [Google Scholar] [CrossRef]
- Gobalet, K.W. Morphology of the parrotfish pharyngeal jaw apparatus. Am. Zool. 1989, 29, 319–331. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Choat, H. A functional analysis of grazing in parrotfishes (family Scaridae): The ecological implications. Environ. Biol. Fishes 1990, 28, 189–214. [Google Scholar] [CrossRef]
- Francini-Filgo, R.B.; Moura, R.; Ferreira, C.M.; Coni, E.O.C. Live coral predation by parrotfishes (Percifioirmes: Scaridae) in the Abrolhos Bank, eastern Brazil, with comments on the classification of species into functional groups. Neotrop. Ichthyol. 2008, 6, 191–200. [Google Scholar] [CrossRef]
- Miller, M.W.; Hay, M.E. Effects of fish predation and seaweed competition on survival and growth of corals. Oecologia 1998, 113, 231–238. [Google Scholar] [CrossRef]
- Littler, M.M.; Taylor, P.R.; Littler, D.S. Complex interactions in the control of coral zonation on a Caribbean reef flat. Oecologia 1989, 80, 331–340. [Google Scholar] [PubMed]
- Rotjan, R.D.; Lewis, S.M. Selective predation by parrotfishes on the reef coral Porites astreoides. J. Exp. Mar. Biol. Ecol. 2005, 305, 193–201. [Google Scholar]
- Fitt, W.K.; McFarland, F.K.; Warner, M.E.; Chilcoat, G.C. Seasonal cycles of tissue biomass and zooxanthellae densities in Caribbean reef corals. Limnol. Ocean. 2000, 45, 677–685. [Google Scholar]
- Rotjan, R.D.; Dimond, J.L. Discriminating causes from consequences of persistent parrotfish corallivory. J. Exp. Mar. Biol. Ecol. 2010, 390, 188–195. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).