Abstract
Mucomics is the study of mucus and its biochemical properties. This discipline has gained increasing attention due to the critical roles mucus plays in protection, adhesion, and communication across species. Ethical restrictions on vertebrate research have driven the interest in invertebrate models such as mollusks. Mollusks produce large amounts of mucus which has several functions, including immune defense, digestion, and environmental adaptation. Mollusks—terrestrial, freshwater, or marine—are valuable models for investigating mucus composition and its responses to environmental stressors, including heavy metal contamination. Histochemical and glycomic techniques have revealed variations in mucin glycosylation patterns that influence mucus functionality, such as its viscoelastic and adhesive properties. Bivalves and gastropods, widely used as bioindicators and generally not subject to regulatory constraints in experimental use, accumulate pollutants in their mucus, reflecting environmental health. Investigative techniques such as lectin histochemistry, proteomic, and glycomic analyses provide insights into the impact of contaminants on mucus composition. Further research on molluscan mucins can enhance understanding of their physiological roles, environmental interactions, and potential biomedical applications. By integrating molecular and histochemical approaches, mucomic studies offer a comprehensive perspective on mucus function, advancing both ecological monitoring and biotechnological applications.
1. Introduction
The study of mucus and its biochemical properties, collectively referred to as mucomics, has gained increasing interest due to the biological significance of mucus in protection, adhesion, and communication in many species [1].
In recent decades, histochemical and glycohistochemical studies have characterized the mucins of various organisms, also vertebrates [2,3,4,5], with the aim of elucidating the biological and functional roles of mucus across different taxa and identifying potential similarities with humans. Nevertheless, the pursuit of experimental and applied models must necessarily consider both ethical constraints and housing requirements.
The European Parliament’s Directive 2010/63/EU on the protection of animals used for scientific purposes, implemented in Italy through legislative decree D.Lgs. 26/2014, has strongly restricted animal experimentation to promote animal welfare and minimize suffering. This legislation applies to (1) live non-human vertebrates, including larval forms capable of independent feeding and mammalian fetuses in the final third of their development, and (2) cephalopod mollusks.
As a result of these regulations, the use of animals such as fish (e.g., zebrafish, Danio rerio), amphibians (e.g., Xenopus laevis), mice, and rats in scientific research is highly controlled. These animals must be registered, housed in isolated enclosures at research facilities, and cared for by trained technicians responsible for maintaining their health and hygiene. Consequently, obtaining permits for the housing and use of these animals is challenging and requires dedicated facilities, qualified personnel, and adherence to stringent guidelines.
To address these challenges while upholding the principles of animal welfare, there is a growing emphasis on identifying invertebrates as alternative experimental models. Many invertebrate taxa exhibit morphological or functional traits that make them suitable for such purposes. Additionally, adherence to animal welfare standards is ensured through the development of tailored sedation protocols for different invertebrate groups [6,7].
Mollusks, with their diverse mucus secretions [8,9,10] provide a compelling model for investigating the molecular, histochemical, and lectin-based characterization of glycoconjugates. This review discusses the potential of mollusks in mucomic research, emphasizing histochemical and lectin-based studies as exemplified by recent research on various species.
Mollusks are promising experimental models for the study of mucomics. These organisms produce a variety of glycans in their mucus, which play crucial roles in processes such as locomotion, immune defense, and adhesion to surfaces [11,12,13,14]. This study applies an integrated “mucomic” approach, combining proteomics, glycomics, and materials science to understand the relationships between mucus structure and function.
Within the diverse phylum Mollusca, three classes of notable interest—both commercial and scientific—play a key role in animal experimentation and environment monitoring [15]. Among gastropods, terrestrial species such as Cornu aspersum [8] and Eobania vermiculata [16] stand out, while marine Gastropoda include the Aplysiida [17] and Patellida [18] orders. In fresh water, a gastropod like Lymnaea stagnalis [19] serves as an example. These species produce significant amounts of mucus which not only aids in locomotion and substrate adhesion but also serves functions such as immune defense and osmoregulation [1]. There are also potentialities of mucus that still need to be explored, such as the regulation of mitochondrial apoptosis [20]. Due to their substantial mucus production, combined with their ease of maintenance and reproduction under laboratory conditions, these gastropods are excellent candidates for research applications.
Regarding bivalves, families such as Mytilidae, Ostreidae, and Veneridae are widely used as environmental bioindicators [21,22,23,24] due to their sensitivity to chemical changes in water, exemplified by programs like Mussel Watch [25,26]. In these species, mucus is produced primarily for feeding purposes and to protect their gills—key filtering organs—from organic and inorganic particulates that could cause mechanical damage; additionally, mucus plays a role in defense against pathogens and environmental contaminants [1,27,28].
The study of cephalopod mucus, particularly octopuses and cuttlefish, also arouses interest. Even though there are studies on the histochemistry of their mucus [29], using these species in basic research and experimentation entails navigating ethical and legislative challenges in some regions, owing to their complex biology and behavior [30]. According to the European Directive 2010/63/UE, it is easier to obtain permits for research on their paralarvae [31].
While it has been observed in cephalopods that the mucus produced by the mantle serves several functions—including protection, osmotic regulation, and forming a barrier to the external environment—the secretory cells also increase buoyancy and reduce friction during swimming, making movement more efficient [32]. In the suckers, the presence of sulphated glycosaminoglycans helps in the adhesion process [29]. However, for conducting environmental monitoring studies and assessing mucin alterations after exposure to contaminants, we have opted for model organisms that are readily available, easily accommodated, and not subject to stringent constraints. Bivalves and gastropods, commonly exploited commercially and abundantly available, are cultivated in large quantities through aquaculture or heliciculture. This selection facilitates the execution of numerous experimental replicates, utilizing an appropriate number of individuals to yield statistically significant results, and encourages fellow researchers to adopt these animal models.
2. The Glycosylation of Mucins in Mollusks: Structures, Functions, and Environmental Impacts
Mucins are high-molecular-weight epithelial glycoproteins with a high content of O-linked oligosaccharides that are synthesized, stored, and secreted by epithelial mucosal cells, particularly goblet cells, and are characterized by a protein backbone with a variable number of repeated amino acid sequences [33]. These sequences are particularly rich in serine/threonine, which are the attachment sites for O-glycans. There are two main classes of mucins: secreted mucins, which are components of mucus, and membrane-bound mucins. The evolutionary role of mucins in mollusks also highlights how these proteins have adapted to different ecological niches and environmental challenges over time. For example, the adhesive mucus of C. aspersum contains proteins that form strong, flexible bonds, allowing the snail to adhere to various surfaces. The lubricating mucus is designed to reduce friction and facilitate movement, while the protective mucus combines properties of both adhesive and lubricating mucus to form a robust barrier against environmental threats. This adaptability of mucins demonstrates their evolutionary significance in helping mollusks survive and thrive in diverse habitats [8]. The synthesis of mucin apoproteins is encoded by a series of genes which in humans are referred to as MUC (MUC1, MUC2, etc.), and they have homologues in many other species [34]. The mucin apoprotein undergoes significant post-translational modifications, particularly extensive glycosylation. The carbohydrate content represents a notable percentage, ranging from 50% to 90% of the total molecular weight. O-linked oligosaccharide chains prevail, but N-linked chains are also present at the terminal regions, and although they are quantitatively less represented, they are functionally very important. N-linked oligosaccharides tend to be complex and branched. The first sugar in the chain is N-acetylglucosamine (GlcNAc), which is linked to an asparagine in the polypeptide chain, part of a specific consensus sequence (Asn-Xaa-Ser/Thr) [8]. Other sugars are added to the initial GlcNAc until a complex branched chain is formed. N-glycosylation begins in the rough endoplasmic reticulum and is completed in the Golgi apparatus. All N-glycans contain a common pentasaccharide core, Man3GlcNAc2. Additional glycosidic residues are added to the pentasaccharide core in the Golgi. O-linked oligosaccharide chains generally contain an initial residue of GalNAc. There are at least eight different types of cores. They are typically less branched than N-linked oligosaccharides. O-glycosylation begins in the Golgi, and sugars are added one by one to the chain through the action of specific glycosyltransferases as the glycoproteins migrate from the Cis to the Trans Golgi [35]. This results in a high heterogeneity of oligosaccharide chains. Mucins are highly glycosylated proteins that constitute key components of the mucus secreted by molluscan epithelia. Davies and Hawkins [1] highlighted various roles of mucus, confirmed by other authors [11,12,13,36]. They play various roles, including filter feeding, digestion, respiration, maintenance of the mantle cavity, defense against predators and pathogens, adhesion or penetration into substrates, osmoregulation, biomineralization, and signaling through luminescence both within and between species.
Filter Feeding: Mucus aids in the transport of food particles from the gills to the mouth in bivalves, and it is involved in the sorting and rejection of particles based on their nutritional value. Lectins in the mucus can mediate particle selection by binding to specific carbohydrates on the surface of microalgae, facilitating the ingestion of preferred particles and rejection of others.
Digestion: Mucus in the digestive tract helps in the lubrication and transport of food particles, and it may contain enzymes that aid in the breakdown of food.
Respiration: The mucus covering the gills can trap and remove particles that might otherwise clog the gill filaments, ensuring efficient gas exchange.
Maintenance of the mantle cavity: Mucus helps to keep the mantle cavity clean by trapping and expelling debris and pathogens.
Defense against predators and pathogens: Mucus can contain antimicrobial compounds such as lysozymes and lectins that help protect against infections. It can also make the mollusk slippery and difficult for predators to handle.
Adhesion or penetration into substrates: Mucus allows mollusks to adhere to surfaces, which is crucial for locomotion and maintaining position in their habitat. It can also aid in the penetration of substrates for burrowing species.
Osmoregulation: Mucus can help maintain the osmotic balance by acting as a barrier to water loss in terrestrial and intertidal species.
Biomineralization: Mucus plays a role in the formation and repair of shells by providing a medium for the deposition of calcium carbonate.
Signaling through luminescence: Some mollusks use mucus to produce bioluminescent signals for communication, which can be important for mating, deterring predators, or luring prey.
These functions highlight the versatility and importance of mucus in the survival and ecological interactions of marine mollusks. In addition to their various roles, mucins also influence interactions with aquatic microbiota, contributing to the overall health of mollusks and the stability of the ecosystem. Mucins found in pseudofeces contribute to the particulate organic matter present in coastal ecosystems [1]. In mucin structures, oligosaccharide chains are typically connected to the protein backbone via α-O-glycosidic bonds to serine or threonine residues. The variations in glycosylation and carbohydrate composition of these sugar chains have an impact on the viscoelastic, lubricating, and hydration characteristics of mucus, thereby influencing its functionality [37]. Comparative studies on mucins in various invertebrates and vertebrates could provide valuable insights, emphasizing both the similarities and differences in glycosylation patterns and functional roles. Despite their significance, research regarding the composition, distribution, and variability of mucins in molluscan secretions remains limited. Classical histochemical analyses suggest that the oligosaccharide chains in mucus can exhibit neutral or acidic properties, which result from the presence of carboxylated or sulfated residues [9,18,38,39,40,41,42].
However, in situ investigations on the composition of these oligosaccharide chains are even scarcer, indicating the presence of mannosylated, glycosaminoglycan-modified, galactosylated/galactosaminylated, and fucosylated residues, while the presence of sialic acid remains a matter of debate [9,18,39,40,41,42,43]. It has been noted that glycosylation varies across different types of secreting cells distributed in various anatomical regions of bivalves, including the mantle, foot, gills, and digestive gland [9,10,28,38,44,45]. Additionally, certain molluscan classes, such as gastropods and cephalopods, show variations in mucin composition within the same cell type due to physical factors (hydrodynamics, salinity, temperature, humidity) or specific biological activities (reproduction, growth, locomotion, feeding) [18,31,46]. Furthermore, changes in the qualitative and quantitative aspects of mucins have been linked to pollutants, indicating that such variations could serve as biomarkers [47,48]. Moreover, understanding how environmental changes, such as ocean acidification and global warming, impact mucin glycosylation and composition can help assess the overall health of mollusk populations. As filter-feeding organisms capable of filtering contaminants from the environment, bivalves play a vital role in water quality monitoring [49,50,51,52], and they are increasingly regarded as models for human health studies [53]. It is crucial to underscore the need for further research to explore the variability and function of mucins in various developmental stages of mollusks and across different species, as significant gaps remain in the current literature. Understanding the complexity of mucin glycosylation in mollusks not only provides fundamental insights into their biology but also contributes to environmental monitoring strategies and potential biomedical applications. Furthermore, the potential of mucins as biomarkers for environmental health cannot be overstated. They may serve as indicators of ecosystem integrity, particularly in the face of anthropogenic challenges such as pollution and climate change. Given that mollusks play a vital role as bioindicators in aquatic environments, understanding how mucin expression and composition are affected by environmental changes is crucial. Moreover, the investigation of mucin diversity can provide essential insights into the physiological adaptations of mollusks, thus enriching our understanding of their evolutionary pathways. In this regard, future research efforts should focus on integrating advanced glycomic technologies and in situ analyses to uncover the intricate mechanisms by which mucin variability impacts both molluscan physiology and their interactions with the environment. A comprehensive exploration of the functions and adaptations of mucins in mollusks promises to illuminate not only their biological significance but also their potential applications in environmental monitoring and biomedical research.
3. The Role of Mucins in Mollusks: Immune Protection, Digestion, and Environmental Adaptations
Mucomics, the study of mucins and their composition, provides an important window into the physiological processes in mollusks, particularly concerning immunity and digestion [8]. Mucins are highly glycosylated proteins that, as key components of mucus, perform multifunctional roles vital for the survival of these aquatic organisms. One of the main functions of mucins is their ability to act as physical barriers [54]. They protect mollusks from pathogens, pollutants, and other harmful agents present in the aquatic environment. Mucins create a protective layer that not only facilitates immune defense but also provides a means for the physical removal of foreign particles. Through adhesion and interaction mechanisms, mucins can trap and neutralize pathogens, preventing infections and diseases. This function is particularly important given the continuous exposure of mollusks to microorganisms and pollutants [55]. Furthermore, mucins play a significant role in modulating immune responses. The varied composition of mucins in response to environmental stimuli highlights how these organisms can adapt their defenses according to specific challenges [56]. For example, the group of mucins produced may change when exposed to new pathogens, suggesting a dynamic adaptation mechanism and an ability to activate the immune response in mollusks. From a digestive perspective, mucins are essential for lubricating food, facilitating the formation of a more manageable food bolus, and ensuring smooth passage through the digestive system. Due to their viscoelastic properties, mucins make food easier to handle and process, thereby improving the efficiency of the digestive process. Variations in mucin composition can also reflect the specific diet of mollusks, revealing how these animals adapt to different nutritional sources, from plant materials to plankton. Analyzing mucins and their variations also provides important insights into the health of molluscan ecotypes and environmental changes [57]. Mucins show significant diversity in relation to salinity, temperature, and other environmental conditions. These variations can serve as biomarkers to indicate environmental health and water quality, providing a useful method for monitoring the effects of pollution and environmental changes on biodiversity [58]. Recent studies have particularly highlighted the importance of mucin glycosylation, which can significantly influence the rheological properties of mucus [55]. Variations in glycosylation can have direct impacts on the lubricating and adhesive properties of mucus, crucial for the survival and functionality of mollusks in variable environments [59]. Understanding how different species of mollusks manipulate their mucins can also provide insights into addressing ecological and health-related challenges [60]. Finally, delving into the mechanisms of mucins in mollusks has not only ecological relevance but can also offer avenues for applications in medicine and biotechnology. Mucins could be harnessed for their potential in biomolecular contexts, opening new paths for research in human health and veterinary medicine. In summary, mucomics represents a promising field for uncovering the complexities of physiological processes in mollusks, contributing to a deeper understanding of their biology, ecology, and importance in aquatic ecosystems. Studying how mucins influence immunity and digestion can reveal vital information for both species conservation and advancements in biomedical technologies.
4. Models Study
4.1. Gastropods
Terrestrial Gastropods: Helicidae such as the garden snail Cornu aspersum and the chocolate band snail Eobania vermiculata are prominent terrestrial gastropods extensively studied for their responses to environmental stressors, particularly aluminum exposure. Research on Cornu aspersum identified three types of mucus—protective, adhesive, and lubricating—each serving distinct mechanical and functional roles. Analytical approaches such as proteomic, glycomic, and biophysical methods, including advanced mass spectrometry and rheological assays, have characterized its macromolecular composition. These studies highlighted the high viscosity and elasticity of adhesive mucus compared to the simpler lubricating mucus, demonstrating its ecological significance [8].
For Eobania vermiculata, histochemical and lectin–histochemical techniques have been employed to analyze the effects of aluminum exposure on mucin secretion. Methods like Periodic Acid–Schiff reagent (PAS) and Alcian Blue staining revealed a reduction in glycosylation and the production of acidic glycans, impairing the viscosity and protective properties of mucus. Specific lectins, such as UEA I and WGA (detail in Table 1), detected fucosylated and GlcNAc-linked glycans, respectively, emphasizing the impact of environmental stressors on mucosal functionality [16,61]. These findings underscore the vulnerability of terrestrial gastropods to contaminants and their potential as models for studying the impact of pollutants on mucin structure and function.
Freshwater Gastropods: Freshwater species, such as the great pond snail (Lymnaea stagnalis), are key models for monitoring heavy metal pollution in aquatic environments. Lymnea stagnalis has demonstrated a high capacity for bioaccumulating cadmium (Cd) and zinc (Zn), reflecting site-specific contamination levels. This species is particularly valuable for assessing pollution dynamics in freshwater ecosystems, as its ability to sequester metals through mucus and other tissues provides critical insights into the effects of contaminants on molluscan physiology.
Other freshwater gastropods, such as river snails Viviparus spp., exhibit a dual feeding strategy—deposit feeding and suspension feeding—that allows them to reflect sediment-associated metal concentrations. This feeding behavior enhances their utility as bioindicators for sediment-bound pollutants, offering a comprehensive understanding of contamination in aquatic systems [62].
Marine gastropods, including the veined rapa whelk Rapana venosa and the moon snail Neverita didyma, exhibit significant bioaccumulation of Cd in coastal ecosystems. These species have been studied for their capacity to assess pollution levels in marine environments, particularly heavy metals. The high bioaccumulation potential of Rapana venosa makes it a valuable sentinel species for tracking cadmium contamination. Similarly, Neverita didyma has been employed to monitor cadmium levels, emphasizing the role of marine gastropods in evaluating the ecological impact of heavy metal pollution [62].
In the Mediterranean region, gastropods such as Phorcus turbinatus and Patella caerulea have been used to monitor copper (Cu) and Zn pollution due to their sensitivity to these metals. These species provide essential data on coastal metal contamination, contributing to efforts to manage and mitigate pollution in marine ecosystems [63].
4.2. Bivalves
Bivalves are a diverse group of mollusks characterized by their filter-feeding ability, which enables them to extract nutrients from suspended particles in the water. This feeding behavior also makes them effective bioindicators of environmental contamination, as they accumulate pollutants from their surroundings in their tissues and mucus. Across taxa, heavy metals such as Cd, lead (Pb), nickel (Ni), Cu, and Zn are the most studied contaminants. The mucus, enriched with glycans and proteins, serves a dual role as a protective barrier and a medium for trapping contaminants. For example, Ruditapes philippinarum efficiently traps organic pollutants like polycyclic aromatic hydrocarbons (PAHs), although prolonged exposure can cause oxidative stress, lipid peroxidation, and DNA damage [64,65].
Internally, the digestive glands and gills of bivalves act as primary sites for metal sequestration, with lysosomes in digestive cells playing a pivotal role in detoxification. These processes underscore the importance of mollusks as bioindicators for environmental contamination, providing valuable insights into pollutant dynamics and their ecological impact [66,67]. The mucus of bivalves plays critical roles in particle transport, pathogen defense, and environmental interactions. Its composition, enriched with glycans and proteins, varies under different stressors, reflecting changes in functionality.
Techniques such as histochemical staining and biochemical assays have provided insights into how environmental pollutants alter mucus composition and secretion.
Marine bivalves, including Mytilus galloprovincialis, Chamelea gallina, and Ruditapes philippinarum, are extensively studied for their responses to environmental contaminants. An example of the difference in their tissues and the different responses to histochemical treatments, which we will discuss later, can be seen in Figure 1.
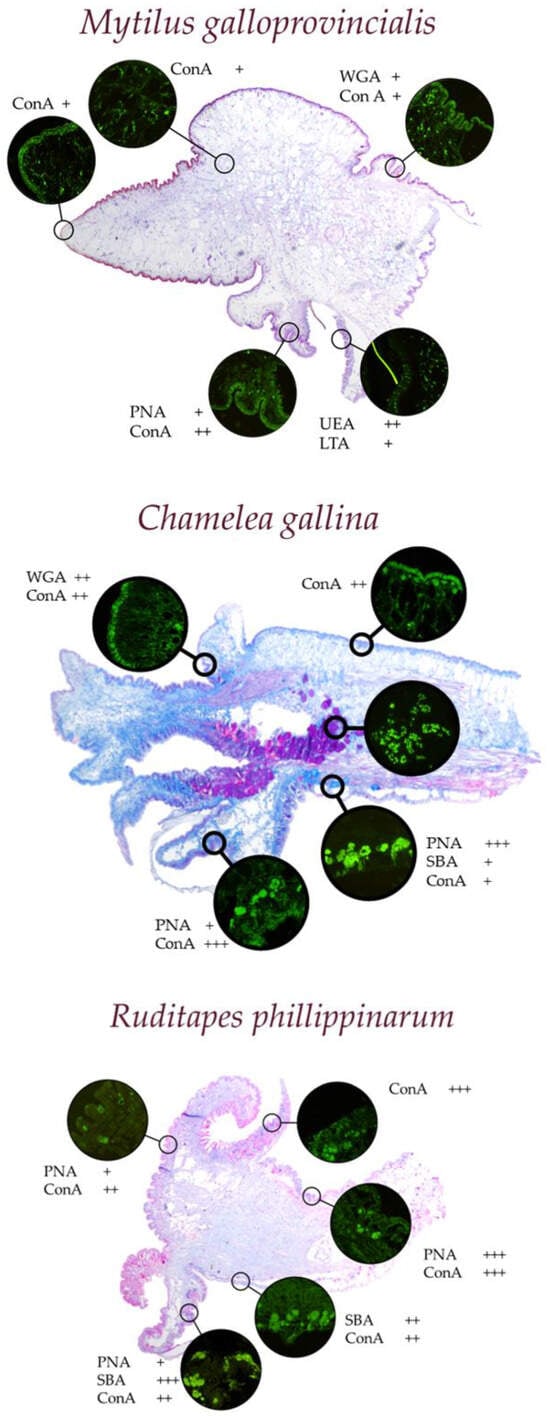
Figure 1.
Representative examples of different morphologies and responses to histological and histochemical staining in the mantle of various marine bivalves. In green on a dark field, the fluorescence response of the mucus-secreting cells present in the different folds of the mantle to the different lectins can be observed.
The Mediterranean mussel Mytilus galloprovincialis is a recognized bioindicator and is widely used for biomonitoring programs such as “Mussel Watch”. Its sessile nature and high bioaccumulation capacity make it ideal for studying heavy metals, including arsenic (As), Pb, chromium (Cr), manganese (Mn), mercury (Hg), and Zn [66,68,69]. Similarly, the brown mussel Perna perna and oysters such as Crassostrea angulata and Crassostrea virginica have been employed in laboratory and field studies to evaluate the uptake, loss, and biological effects of these metals. In studies along the Chinese Bohai Sea, Crassostrea talienwhanensis was found to have a particularly high capacity for bioaccumulation of Cu and Zn compared to other species, while Rapana venosa showed significant cadmium bioaccumulation [65,70]. Notably, the Manila clam Ruditapes philippinarum exhibited the highest Ni content among studied species, emphasizing its utility in assessing Ni contamination [65]. Advanced techniques such as immunohistochemistry, in situ hybridization, and lectin arrays have been used to map gene expression and glycosylation patterns, while ICP-AES quantified seasonal bioaccumulation of trace metals, distinguishing anthropogenic from natural sources [71]. Studies have also demonstrated the role of Mytilus galloprovincialis mucus in mitigating metal toxicity, particularly for Zn and iron (Fe), underscoring its ecological relevance [72].
The Venus striped clam Chamelea gallina has been studied in normal conditions [73] and for pollutant-induced alterations in mucin secretion. Histochemical and lectin–histochemical analyses revealed reductions in glycan diversity, acidity, and viscosity under aluminum exposure. These changes impair the protective and pathogen defense functions of mucus, highlighting the vulnerability of Chamelea gallina to aluminum (Al) pollutants [21].
Ruditapes philippinarum has shown a significant capacity for bioaccumulating polycyclic aromatic hydrocarbons (PAHs) and heavy metals like Cd and Ni. Biochemical assays and molecular markers, including oxidative stress indicators like DNA strand breaks and lipid peroxidation, have been used to detail the impacts of these contaminants. High-performance liquid chromatography (HPLC) has been instrumental in elucidating bioaccumulation pathways and tissue-specific detoxification mechanisms [65,74]. Ruditapes philippinarum has recently also been tested for contamination by rare earth elements such as lanthanum (La) [61].
Freshwater bivalves, such as Dreissena polymorpha (zebra mussel) and Pletholophus swinhoei, play a crucial role in pollution monitoring in riverine and lacustrine ecosystems.
The former has been widely used to assess trace metal pollution in freshwater systems like the Po River in Italy. Studies revealed increasing concentrations of Cd, Pb, Cr, and Ni in mussel tissues over time, reflecting the intensification of pollution in aquatic ecosystems. Stable Cu and Zn levels, as well as consistently low Hg levels, highlight the potential of Dreissena polymorpha as a reliable bioindicator for specific contaminants [67].
Research on Pletholophus swinhoei has focused on site-specific variations in heavy metal concentrations, including As, Ba, Cr, and Ni. These findings provide valuable data for evaluating localized pollution impacts and guiding environmental management strategies [67].
In bivalves, Chamelea gallina mucus [21] serves a critical role in particle transport and pathogen defense. Using Alcian Blue staining and lectins like SBA and PNA, researchers identified significant reductions in glycan diversity and acidity under aluminum exposure, leading to functional impairments. Mytilus galloprovincialis, a widely studied bioindicator, employs mucus to trap and detoxify metals such as zinc and iron. Studies by Miglioli et al. [72] utilized immunohistochemistry, in situ hybridization, and lectin arrays to map gene expression and glycosylation patterns. The seasonal bioaccumulation of trace metals was quantified through ICP-AES, demonstrating the ecological role of mucus in mitigating metal toxicity. Ruditapes philippinarum, as studied by Li et al. [74], uses mucus to trap polycyclic aromatic hydrocarbons (PAHs). Histological and biochemical analyses, including PAS staining and oxidative stress biomarkers such as DNA strand breaks and lipid peroxidation, revealed that mucus entraps contaminants but is affected by oxidative damage due to PAH exposure.
The functional properties of molluscan mucus extend beyond the individual organism, influencing ecosystem dynamics. According to Davies and Hawkins [1], mucus production can consume up to 70% of a mollusk’s energy budget, underscoring its ecological importance. Mucus contributes to organic particulate matter, supports locomotion, and forms barriers against environmental threats. However, as highlighted across these studies, environmental pollutants like aluminum and PAHs can disrupt mucus composition and functionality, with significant implications for molluscan health and ecosystem stability. The integration of advanced techniques, including mass spectrometry, histochemical staining, and lectin arrays, has been instrumental in elucidating the complex structure-function relationships of mucus and its response to environmental stressors.
5. Investigative Techniques and Contaminants
Across mollusks, a range of investigative techniques has been applied to study mucus composition and pollutant bioaccumulation. Proteomic and glycomic analyses, including mass spectrometry and glycan profiling, have been essential in characterizing mucus properties in terrestrial species [75]. Histochemical staining techniques, such as PAS and Alcian Blue and High Iron Diamine, combined with lectin–histochemical methods, have been widely used to investigate mucin glycosylation patterns and their alterations under environmental stressors like aluminum [21,61]. Lectins are proteins useful for recognizing specific carbohydrate structures, and for this reason, they are widely used to perform these glycohistochemical studies. As for the advantages of histochemical methods, they allow for the in situ localization of mucus residues, considering the involvement of multiple cell types in mucus production. Understanding which cells participate in mucus production and the internalization of contaminants outweighs the disadvantages, such as the extended timelines and high costs associated with purchasing lectins. The origin of the most commonly used lectins is listed in Table 1.

Table 1.
Lectins employed with their diluting buffers, working dilutions, and inhibiting sugars. Binding specificities: PNA: Galβ1,3GalNAc; SBA: GalNAc; WGA: (GlcNAcβ1,4)n; LTA: L-Fucα1,6GlcNAc; L-Fucα1,2Galβ1,4(L-Fuc1,3)GlcNAcβ1,6R; UEA I: Fucα1,2; AAL: Fucα1,6GlcNAc-βNAsn; Fucα1,3,Fucα1,4; ConA: D-Man, D-Glc. [76,77,78,79,80,81,82,83].
Table 1.
Lectins employed with their diluting buffers, working dilutions, and inhibiting sugars. Binding specificities: PNA: Galβ1,3GalNAc; SBA: GalNAc; WGA: (GlcNAcβ1,4)n; LTA: L-Fucα1,6GlcNAc; L-Fucα1,2Galβ1,4(L-Fuc1,3)GlcNAcβ1,6R; UEA I: Fucα1,2; AAL: Fucα1,6GlcNAc-βNAsn; Fucα1,3,Fucα1,4; ConA: D-Man, D-Glc. [76,77,78,79,80,81,82,83].
| Lectin | Origin | Buffer | Dilution | Inhibitory Sugar Conc. |
|---|---|---|---|---|
| PNA | Arachis hypogaea | Hepes | 10 mg/mL | 0.2 M Gal |
| SBA | Glycine max | Hepes | 20 mg/mL | 0.2 M GalNAc |
| WGA | Triticum vulgaris | Hepes | 20 mg/mL | 0.5 M GlcNAc |
| LTA | Tetragonolobus purpureus | Hepes | 20 mg/mL | 0.2 M L-Fuc |
| UEA I | Ulex europaeus | Hepes | 10 mg/mL | 0.2 M L-Fuc |
| AAL | Aleuria aurantia | Hepes | 10 mg/mL | 0.2 M L-Fuc |
| ConA | Canavalia ensiformis | Hepes | 20 mg/mL | 0.1 M MαM |
For aquatic gastropods, biochemical assays and tissue analysis methods have been employed to assess bioaccumulated heavy metals, including Cd, Zn, and Cu [84]. These studies utilize advanced techniques such as HPLC (high-performance liquid chromatography), ICP-AES, and ICP-MS to quantify contaminant levels and evaluate their effects on molluscan health and environmental interactions [85,86].
6. Contaminants and Mucus Functionality
Across taxa, heavy metals such as Cd, Pb, Ni, Cu, and Zn are the most studied contaminants. Gastropods like Rapana venosa and bivalves such as Mytilus galloprovincialis exhibit significant alterations in mucus production and composition when exposed to these pollutants. The mucus, enriched with glycans and proteins, serves a dual role as a protective barrier and a medium for trapping contaminants. For example, Ruditapes philippinarum efficiently traps organic pollutants like polycyclic aromatic hydrocarbons (PAHs), although prolonged exposure can cause oxidative stress, lipid peroxidation, and DNA damage [64,65].
Internally, the digestive glands and gills of bivalves act as primary sites for metal sequestration [87,88], with lysosomes in digestive cells playing a pivotal role in detoxification [89]. These processes underscore the importance of mollusks as bioindicators for environmental contamination, providing valuable insights into pollutant dynamics and their ecological impact [66,67,90,91].
7. Conclusions
Mollusks are excellent environmental indicators, as demonstrated using many bivalves in international monitoring programs. We recognize that the use of mollusks as models for translational research towards vertebrates and humans has some significant limitations that deserve discussion. In particular, one critical aspect concerns the biochemical composition of mollusks. It is known that these organisms possess a lower amount, or no amount—this aspect has been debated—of sialic acid compared to vertebrates, which can influence the cellular interactions and biological processes we study. This difference may limit the generalization of results obtained from mollusks to vertebrate physiology.
Moreover, the oligosaccharide chains present in mollusks are structurally different and do not always correspond to those of vertebrates. For example, the linkages and branching of these chains can vary considerably, affecting their functionality and interactions with proteins. This is a crucial element to consider, given the importance of oligosaccharides in cellular recognition and signaling. We are aware that such differences could impact the interpretation of results obtained from our studies using these models. However, we believe that, despite these limitations, mollusks can provide valuable and supplementary information, especially in areas where their uniqueness can offer research insights not obtainable with vertebrate models.
Another aspect to consider during the study and experimental utilization of mollusks pertains to their housing requirements. Aquatic organisms necessitate optimal acclimatization and housing conditions that prevent mortality among the specimens. The overpopulation of tanks is a critical factor to be considered. Additionally, it is necessary to ensure that microalgae or an appropriate food source is available and is supplied throughout the acclimatization and experimental period. Furthermore, the limited knowledge of many species is limited only to the morphology of the shells, neglecting anatomical and histological aspects. Before starting an experimental phase with a new model, it would be advisable to study this model under natural conditions, analyzing different populations. This is relatively easy in the case of stocks from fisheries or shellfish farming but can be more complex and expensive for species that are not commercially important.
A mucomic approach to the study of mucus provides important information on its ecological, physiological and structural significance. Advances in histochemical, proteomic and glycomic techniques have allowed a more detailed characterization of mucins, shedding light on their physiological functions and responses to environmental stresses. However, significant gaps remain in our understanding of the variability of mucins among species, life stages, and environmental conditions.
Future research should aim to deepen our understanding of how pollutants interact with mucins, develop predictive models for assessing pollution, and explore the biotechnological potential of molluscan mucins. Additionally, investigating the impact of climate change on molluscan mucus function will be crucial, as it may alter the role of these organisms in their ecosystems and affect their utility in environmental monitoring. Expanding mucomic studies to include a broader range of mollusk species will be critical in refining our understanding of this vital biological system. Such comparative approaches can uncover unique adaptations and responses, enhancing our ability to use mollusks as sentinel organisms for ecosystem health and human environmental risk assessments. This comprehensive understanding will not only contribute to environmental science but also offer potential innovations in biotechnology, medicine, and climate change resilience strategies. Integrating interdisciplinary approaches will strengthen our knowledge, paving the way for improved environmental management and protection efforts.
Author Contributions
Conceptualization, D.M., M.M. and M.V.G.; writing—original draft preparation, D.M., D.S. and M.V.G.; writing—review and editing, C.D.B., D.S. and M.M.; supervision, D.M., M.M. and M.V.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
We are grateful to Michele Civiello for his technical assistance in preparing this review.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Davies, M.S.; Hawkins, S. Mucus from marine molluscs. In Advances in Marine Biology; Elsevier: Amsterdam, The Netherlands, 1998; Volume 34, pp. 1–71. [Google Scholar]
- Carlucci, R.; Mentino, D.; Semeraro, D.; Ricci, P.; Sion, L.; Scillitani, G. Comparative histochemical analysis of intestinal glycoconjugates in the blunthead pufferfish Sphoeroides pachygaster and grey triggerfish Balistes capriscus (Teleostei: Tetraodontiformes). J. Fish. Biol. 2019, 94, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Gristina, M.; Bertrandino, S.; Cardone, F.; Mentino, D.; Corriero, G.; Scillitani, G. Skin filament recovery after clipping in Hippocampus guttulatus: Behavioural and histological aspects. Aquat. Biol. 2017, 26, 149–157. [Google Scholar] [CrossRef]
- Liquori, G.E.; Mastrodonato, M.; Zizza, S.; Ferri, D. Glycoconjugate histochemistry of the digestive tract of Triturus carnifex (Amphibia, Caudata). J. Mol. Histol. 2007, 38, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Carlucci, R.; Cipriano, G.; Fanizza, C.; Gerussi, T.; Maglietta, R.; Petrella, A.; Pietroluongo, G.; Ricci, P.; Semeraro, D.; Guglielmi, M.V.; et al. Glycopatterns of the foregut in the striped dolphin Stenella coeruleoalba, Meyen 1833 from the Mediterranean Sea. Mar. Mammal Sci. 2021, 38, 190–211. [Google Scholar] [CrossRef]
- Ross, L.G.; Ross, B. Anaesthetic and Sedative Techniques for Aquatic Animals; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Wahltinez, S.J.; Harms, C.A.; Lewbart, G.A. Anesthesia and analgesia in invertebrates. In Anesthesia and Analgesia in Laboratory Animals; Melissa, C., Dyson, P.J., Lofgren, J., Nunamaker, E.A., Pang, D., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 647–671. [Google Scholar]
- Cerullo, A.R.; McDermott, M.B.; Pepi, L.E.; Liu, Z.-L.; Barry, D.; Zhang, S.; Yang, X.; Chen, X.; Azadi, P.; Holford, M.; et al. Comparative mucomic analysis of three functionally distinct Cornu aspersum Secretions. Nat. Commun. 2023, 14, 5361. [Google Scholar] [CrossRef] [PubMed]
- Calabro, C.; Albanese, M.P.; Martella, S.; Licata, P.; Lauriano, E.R.; Bertuccio, C.; Licata, A. Glycoconjugate histochemistry and nNOS immunolocalization in the mantle and foot epithelia of Tapes philippinarum (bivalve mollusc). Folia Histochem. Cytobiol. 2005, 43, 151–156. [Google Scholar]
- Eble, A.F. Chapter 4 Anatomy and histology of Mercenaria mercenaria. In Biology of the Hard Clam; Elsevier: Amsterdam, The Netherlands, 2001; pp. 117–220. [Google Scholar]
- Espinosa, E.P.; Perrigault, M.; Ward, J.E.; Shumway, S.E.; Allam, B. Lectins associated with the feeding organs of the oyster Crassostrea virginica can mediate particle selection. Biol. Bull. 2009, 217, 130–141. [Google Scholar] [CrossRef]
- Prezant, R.S. Form, Function and Phylogeny of Bivalve Mucins; Hong Kong University Press: Hong Kong, China, 1990. [Google Scholar]
- Bolognani Fantin, A.; Vigo, E. La mucinogenesi nei Molluschi. IV. Caratteristiche istochimiche dei tipi cellulari presenti nel piede e nel mantello di alcune specie di Gasteropodi. Riv. Istochem. Norm. Pat 1967, 13, 1–28. [Google Scholar]
- Smolowitz, R. Mollusca: Bivalvia. Invertebr. Hist. 2021, 163–183. [Google Scholar] [CrossRef]
- Gupta, S.K.; Singh, J. Evaluation of mollusc as sensitive indicator of heavy metal pollution in aquatic system: A review. IIOAB J. 2011, 2, 49–57. [Google Scholar]
- Cofone, R.; Carraturo, F.; Capriello, T.; Libralato, G.; Siciliano, A.; Del Giudice, C.; Maio, N.; Guida, M.; Ferrandino, I. Eobania vermiculata as a potential indicator of nitrate contamination in soil. Ecotoxicol. Environ. Saf. 2020, 204, 111082. [Google Scholar] [CrossRef]
- Lobo-da-Cunha, A.; Batista-Pinto, C. Stomach of Aplysia depilans (Mollusca, Opisthobranchia): A histochemical, ultrastructural, and cytochemical study. J. Morphol. 2003, 256, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Petraccioli, A.; Maio, N.; Guarino, F.M.; Scillitani, G. Seasonal variation in glycoconjugates of the pedal glandular system of the rayed Mediterranean limpet, Patella caerulea (Gastropoda: Patellidae). Zoology 2013, 116, 186–196. [Google Scholar] [CrossRef]
- Jugdaohsingh, R.; Campbell, M.M.; Thompson, R.P.; McCrohan, C.R.; White, K.N.; Powell, J.J. Mucus secretion by the freshwater snail Lymnaea stagnalis limits aluminum concentrations of the aqueous environment. Environ. Sci. Technol. 1998, 32, 2591–2595. [Google Scholar] [CrossRef]
- Colella, F.; Scillitani, G.; Pierri, C.L. Sweet as honey, bitter as bile: Mitochondriotoxic peptides and other therapeutic proteins isolated from animal tissues, for dealing with mitochondrial apoptosis. Toxicology 2021, 447, 152612. [Google Scholar] [CrossRef] [PubMed]
- Guglielmi, M.V.; Semeraro, D.; Ricci, P.; Mastrodonato, M.; Mentino, D.; Carlucci, R.; Mastrototaro, F.; Scillitani, G. First data on the effect of Aluminium intake in Chamelea gallina of exploited stocks in the southern Adriatic Sea (Central Mediterranean Sea). Reg. Stud. Mar. Sci. 2023, 63, 103025. [Google Scholar] [CrossRef]
- Perić, L.; Perusco, V.S.; Nerlović, V. Differential response of biomarkers in the native European flat oyster Ostrea edulis and the non-indigenous Pacific oyster Crassostrea gigas co-exposed to cadmium and copper. J. Exp. Mar. Biol. Ecol. 2020, 523, 151271. [Google Scholar] [CrossRef]
- Spada, L.; Annicchiarico, C.; Cardellicchio, N.; Giandomenico, S.; Di Leo, A. Heavy metals monitoring in mussels Mytilus galloprovincialis from the Apulian coasts (Southern Italy). Med. Mar. Sci. 2013, 14, 99–108. [Google Scholar] [CrossRef]
- Moschino, V.; Delaney, E.; Meneghetti, F.; Ros, L.D. Biomonitoring approach with mussel Mytilus galloprovincialis (Lmk) and clam Ruditapes philippinarum (Adams and Reeve, 1850) in the Lagoon of Venice. Environ. Monit. Assess. 2011, 177, 649–663. [Google Scholar] [CrossRef]
- Farrington, J.W.; Tripp, B.W.; Tanabe, S.; Subramanian, A.; Sericano, J.L.; Wade, T.L.; Knap, A.H.; Edward, D. Goldberg’s proposal of “the mussel watch”: Reflections after 40 years. Mar. Poll. Bull. 2016, 110, 501–510. [Google Scholar] [CrossRef]
- Goldberg, E.D.; Bowen, V.T.; Farrington, J.W.; Harvey, G.; Martin, J.H.; Parker, P.L.; Risebrough, R.W.; Robertson, W.; Schneider, E.; Gamble, E. The mussel watch. Environ. Cons. 1978, 5, 101–125. [Google Scholar] [CrossRef]
- Allam, B.; Espinosa, E.P. Mucosal immunity in mollusks. In Mucosal Health in Aquaculture; Elsevier: Amsterdam, The Netherlands, 2015; pp. 325–370. [Google Scholar]
- Beninger, P.; St-Jean, S. The role of mucus in particle processing by suspension-feeding marine bivalves: Unifying principles. Mar. Biol. 1997, 129, 389–397. [Google Scholar] [CrossRef]
- González-Costa, A.; Fernández-Gago, R.; Carid, S.; Molist, P. Mucus characterisation in the Octopus vulgaris skin throughout its life cycle. Anat. Histol. Embryol. 2020, 49, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Fiorito, G.; Affuso, A.; Basil, J.; Cole, A.; de Girolamo, P.; D’Angelo, L.; Dickel, L.; Gestal, C.; Grasso, F.; Kuba, M.; et al. Guidelines for the Care and Welfare of Cephalopods in Research—A consensus based on an initiative by CephRes, FELASA and the Boyd Group. Lab. Anim. 2015, 49 (Suppl. S2), 1–90. [Google Scholar] [CrossRef] [PubMed]
- Accogli, G.; Scillitani, G.; Mentino, D.; Desantis, S. Characterization of the skin mucus in the common octopus Octopus vulgaris (Cuvier) reared paralarvae. Eur. J. Histochem. EJH 2017, 61, 2518. [Google Scholar] [CrossRef]
- Lee, D.G.; Park, M.W.; Kim, B.H.; Kim, H.; Jeon, M.A.; Lee, J.S. Microanatomy and ultrastructure of outer mantle epidermis of the cuttlefish, Sepia esculenta (Cephalopoda: Sepiidae). Micron 2014, 58, 38–46. [Google Scholar] [CrossRef]
- Strous, G.J.; Dekker, J. Mucin-type glycoproteins. Crit. Rev. Biochem. Mol. Biol. 1992, 27, 57–92. [Google Scholar] [CrossRef]
- Rose, M.C.; Voynow, J.A. Respiratory tract mucin genes and mucin glycoproteins in health and disease. Physiol. Rev. 2006, 86, 245–278. [Google Scholar] [CrossRef]
- Stanley, P. Golgi glycosylation. Cold Spring Harb. Perspect. Biol. 2011, 3, a005199. [Google Scholar] [CrossRef]
- Jørgensen, C.B. Bivalve filter feeding revisited. Mar. Ecol. Prog. Ser. 1996, 142, 287–302. [Google Scholar] [CrossRef]
- Bansil, R.; Turner, B.S. The biology of mucus: Composition, synthesis and organization. Adv. Drug Deliv. Rev. 2018, 124, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Bolognani-Fantin, A.; Vigo, E.; Gervaso, M. La mucinogenesi nei Molluschi. VI. Caratteristiche dei tipi cellulari presenti nel piede e nel mantello di alcune specie di Lamellibranchi. Riv. Istochem. Norm. Patol. 1969, 15, 345–370. [Google Scholar]
- Ottaviani, E.; Fantin, A.M.B.; Bolognani, L. Muramic acid as a glycoconjugate component in Mollusca Gastropoda. Comp. Biochem. Physiol. Part B Comp. Biochem. 1990, 96, 627–632. [Google Scholar] [CrossRef]
- Albanese, M.P.; Calabro, C. Studio strutturale ed istochimico sul mantello e sul piede di natica millepunctata lmk (MOLL. GAST. PROS.). Riv. Idrobiol. 1996, 35, 87–99. [Google Scholar]
- Robledo, Y.; Madrid, J.F.; Leis, O.; Cajaraville, M.P. Analysis of the distribution of glycoconjugates in the digestive gland of the bivalve mollusc Mytilus galloprovincialis by conventional and lectin histochemistry. Cell Tissue Res. 1997, 288, 591–602. [Google Scholar] [CrossRef]
- Bravo Portela, I.; Martínez-Zorzano, V.S.; Molist-Pérez, I.; Molist Garcia, P. Ultrastructure and glycoconjugate pattern of the foot epithelium of the abalone Haliotis tuberculata (Linnaeus, 1758) (Gastropoda, Haliotidae). Sci. World J. 2012, 2012, 960159. [Google Scholar] [CrossRef]
- Ahn, H.-Y.; Sue, L.-F.; Ma, J.K.; Pinkstaff, C.A.; Pore, R.S.; Overman, D.O.; Malanga, C.J. Synthesis and secretion fo mucous glycoprotein by the gill of Mytilus edulis I. Histochemical and chromatographic analysis of [14C] glucosamine bioincorporation. Biochim. Biophys. Acta Gen. Subj. 1988, 966, 122–132. [Google Scholar] [CrossRef]
- Nogarol, L.R.; Brossi-Garcia, A.L.; de Oliveira David, J.A.; Fontanetti, C.S. Morphological and histochemical characterization of gill filaments of the brazilian endemic bivalve Diplodon expansus (Küster, 1856) (Mollusca, Bivalvia, Hyriidae). Microsc. Microanal. 2012, 18, 1450–1458. [Google Scholar] [CrossRef]
- Bilgin, M.; Uluturhan-Suzer, E. Assessment of trace metal concentrations and human health risk in clam (Tapes decussatus) and mussel (Mytilus galloprovincialis) from the Homa Lagoon (Eastern Aegean Sea). Environ. Sci. Pollut. Res. 2017, 24, 4174–4184. [Google Scholar] [CrossRef]
- Davies, M.; Hawkins, S.; Jones, H. Mucus production and physiological energetics in Patella vulgata L. J. Molluscan Stud. 1990, 56, 499–503. [Google Scholar] [CrossRef]
- Cutuli, M.A.; Guarnieri, A.; Pietrangelo, L.; Magnifico, I.; Venditti, N.; Recchia, L.; Mangano, K.; Nicoletti, F.; Di Marco, R.; Petronio, G. Potential mucosal irritation discrimination of surface disinfectants employed against SARS-CoV-2 by Limacus flavus slug mucosal irritation assay. Biomedicines 2021, 9, 424. [Google Scholar] [CrossRef]
- de Oliveira David, J.A.; Fontanetti, C.S. The Role of Mucus in Mytella falcata (Orbigny 1842) gills from polluted environments. Water Air Soil Pollut. 2009, 203, 261–266. [Google Scholar] [CrossRef]
- Goldberg, E.D. The mussel watch—A first step in global marine monitoring. Mar. Pollut. Bull. 1975, 6, 111. [Google Scholar] [CrossRef]
- Kádár, E.; Salánki, J.; Jugdaohsingh, R.; Powell, J.J.; McCrohan, C.R.; White, K.N. Avoidance responses to aluminium in the freshwater bivalve Anodonta cygnea. Aquat. Toxicol. 2001, 55, 137–148. [Google Scholar] [CrossRef]
- March, F.A.; Dwyer, F.J.; Augspurger, T.; Ingersoll, C.G.; Wang, N.; Mebane, C.A. An evaluation of freshwater mussel toxicity data in the derivation of water quality guidance and standards for copper. Environ. Toxicol. Chem. 2007, 26, 2066–2074. [Google Scholar] [CrossRef]
- Helmholz, H.; Ruhnau, C.; Pröfrock, D.; Erbslöh, H.-B.; Prange, A. Seasonal and annual variations in physiological and biochemical responses from transplanted marine bioindicator species Mytilus spp. during a long term field exposure experiment. Sci. Total Environ. 2016, 565, 626–636. [Google Scholar] [CrossRef]
- Robledo, J.A.F.; Yadavalli, R.; Allam, B.; Espinosa, E.P.; Gerdol, M.; Greco, S.; Stevick, R.J.; Gómez-Chiarri, M.; Zhang, Y.; Heil, C.A. From the raw bar to the bench: Bivalves as models for human health. Dev. Comp. Immunol. 2019, 92, 260–282. [Google Scholar] [CrossRef]
- Pinzon Martin, S.; Seeberger, P.H.; Varon Silva, D. Mucins and Pathogenic Mucin-Like Molecules Are Immunomodulators During Infection and Targets for Diagnostics and Vaccines. Front. Chem. 2019, 7, 710. [Google Scholar]
- Sheng, Y.H.; Hasnain, S.Z. Mucus and Mucins: The Underappreciated Host Defence System. Front. Cell Infect. Microbiol. 2022, 12, 856962. [Google Scholar] [CrossRef]
- Linden, S.K.; Sutton, P.; Karlsson, N.G.; Korolik, V.; McGuckin, M.A. Mucins in the mucosal barrier to infection. Mucosal Immunol. 2008, 1, 183–197. [Google Scholar] [CrossRef]
- Montagne, L.; Piel, C.; Lalles, J.P. Effect of diet on mucin kinetics and composition: Nutrition and health implications. Nutr. Rev. 2004, 62, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Bhagwat, P.V.; Ravindran, C.; Irudayarajan, L. Beneficial properties of mucus in coral adaptations and ecological interactions. Mar. Biol. 2024, 171, 46. [Google Scholar] [CrossRef]
- Hanson, R.; Hollingsworth, M. Functional Consequences of Differential O-glycosylation of MUC1, MUC4, and MUC16 (Downstream Effects on Signaling). Biomolecules 2016, 6, 34. [Google Scholar] [CrossRef] [PubMed]
- El-Gendy, K.S.; Gad, A.F.; Radwan, M.A. Physiological and behavioral responses of land molluscs as biomarkers for pollution impact assessment: A review. Environ. Res. 2021, 193, 110558. [Google Scholar] [CrossRef]
- Guglielmi, M.V.; Mastrodonato, M.; Semeraro, D.; Mentino, D.; Capriello, T.; La Pietra, A.; Giarra, A.; Scillitani, G.; Ferrandino, I. Aluminum exposure alters the pedal mucous secretions of the chocolate-band snail, Eobania vermiculata (Gastropoda: Helicidae). Microsc. Res. Tech. 2024, 87, 1453–1466. [Google Scholar] [CrossRef] [PubMed]
- De Gregori, I.; Pinochet, H.; Gras, N.; Muñoz, L. Variability of cadmium, copper and zinc levels in molluscs and associated sediments from Chile. Environ. Pollut. 1996, 92, 359–368. [Google Scholar] [CrossRef]
- Campanella, L.; Conti, M.E.; Cubadda, F.; Sucapane, C. Trace metals in seagrass, algae and molluscs from an uncontaminated area in the Mediterranean. Environ. Pollut. 2001, 111, 117–126. [Google Scholar] [CrossRef]
- Rainbow, P.; Wolowicz, M.; Fialkowski, W.; Smith, B.; Sokolowski, A. Biomonitoring of trace metals in the Gulf of Gdansk, using mussels (Mytilus trossulus) and barnacles (Balanus improvisus). Water Res. 2000, 34, 1823–1829. [Google Scholar] [CrossRef]
- Lee, C.S.-L.; Li, X.; Shi, W.; Cheung, S.C.-N.; Thornton, I. Metal contamination in urban, suburban, and country park soils of Hong Kong: A study based on GIS and multivariate statistics. Sci. Total Environ. 2006, 356, 45–61. [Google Scholar] [CrossRef]
- Wagner, A.; Boman, J. Biomonitoring of trace elements in muscle and liver tissue of freshwater fish. Spectroch Acta Part B At. Spectrosc. 2003, 58, 2215–2226. [Google Scholar] [CrossRef]
- Camusso, M.; Balestrini, R.; Muriano, F.; Mariani, M. Use of freshwater mussel Dreissena polymorpha to assess trace metal pollution in the lower river Po (Italy). Chemosphere 1994, 29, 729–745. [Google Scholar] [CrossRef]
- Di Leo, A.; Cardellicchio, N.; Giandomenico, S.; Spada, L. Mercury and methylmercury contamination in Mytilus galloprovincialis from Taranto Gulf (Ionian Sea, Southern Italy): Risk evaluation for consumers. Food Chem. Toxicol. 2010, 48, 3131–3136. [Google Scholar] [CrossRef]
- Idris, A.M.; Eltayeb, M.; Potgieter-Vermaak, S.S.; Van Grieken, R.; Potgieter, J. Assessment of heavy metals pollution in Sudanese harbours along the Red Sea Coast. Microchem. J. 2007, 87, 104–112. [Google Scholar] [CrossRef]
- Sokolova, I.; Ringwood, A.; Johnson, C. Tissue-specific accumulation of cadmium in subcellular compartments of eastern oysters Crassostrea virginica Gmelin (Bivalvia: Ostreidae). Aquatic Toxicol. 2005, 74, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Chouikh, N.-E.; Alahyane, H.; Mounir, A.; El Hachimi, Y.; Cheggour, M. Trace metal bioaccumulation in Mytilus galloprovincialis from Essaouira protected coastal area (Atlantic coast of Morocco): Implications for marine ecosystem and human health. Mar. Pollut. Bull. 2024, 209, 117126. [Google Scholar] [CrossRef] [PubMed]
- Miglioli, A.; Tredez, M.; Boosten, M.; Sant, C.; Carvalho, J.E.; Dru, P.; Canesi, L.; Schubert, M.; Dumollard, R. The Mediterranean mussel Mytilus galloprovincialis: A novel model for developmental studies in mollusks. Development 2024, 151, dev202256. [Google Scholar] [CrossRef]
- Guglielmi, M.V.; Semeraro, D.; Mentino, D.; Mastrodonato, M.; Mastrototaro, F.; Scillitani, G. Season- and sex-related variation in mucin secretions of the striped Venus clam, Chamelea gallina (Linnaeus, 1758) (Bivalvia: Veneridae). Eur. Zool. J. 2023, 90, 252–269. [Google Scholar] [CrossRef]
- Li, D.; Liu, T.; Pan, L.; Hu, F.; Jin, Q. Bioaccumulation and oxidative damage of polycyclic aromatic hydrocarbon mixtures in Manila clam Ruditapes philippinarum. Ecotoxicol. Environ. Saf. 2020, 197, 110558. [Google Scholar] [CrossRef]
- Liegertova, M.; Maly, J. Gastropod Mucus: Interdisciplinary Perspectives on Biological Activities, Applications, and Strategic Priorities. ACS Biomater. Sci. Eng. 2023, 9, 5567–5579. [Google Scholar] [CrossRef]
- Lotan, R.; Sharon, N. Peanut (Arachis hypogaea) agglutinin. Methods Enzymol. 1978, 50, 361–367. [Google Scholar]
- Bhattacharyya, L.; Haraldsson, M.; Brewer, C.F. Precipitation of galactose-specific lectins by complex-type oligosaccharides and glycopeptides: Studies with lectins from Ricinus communis (agglutinin I), Erythrina indica, Erythrina arborescens, Abrus precatorius (agglutinin), and Glycine max (soybean). Biochemistry 1988, 27, 1034–1041. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, J.T.; Morris, A.; Dexter, T.M. Identification of two binding sites for wheat-germ agglutinin on polylactosamine-type oligosaccharides. Biochem. J. 1985, 231, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Debray, H.; Decout, D.; Strecker, G.; Spik, G.; Montreuil, J. Specificity of twelve lectins towards oligosaccharides and glycopeptides related to N-glycosylproteins. Eur. J. Biochem. 1981, 117, 41–55. [Google Scholar] [CrossRef] [PubMed]
- Sughii, S.; Kabat, E.A.; Baer, H.H. Further immunochemical studies on the combining sites of Lotus tetragonolobus and Ulex europaeus I and II lectins. Carbohydr. Res. 1982, 99, 99–101. [Google Scholar] [CrossRef]
- Chandrasekaran, E.V.; Xue, J.; Xia, J.; Khaja, S.D.; Piskorz, C.F.; Locke, R.D.; Neelamegham, S.; Matta, K.L. Novel interactions of complex carbohydrates with peanut (PNA), Ricinus communis (RCA-I), Sambucus nigra (SNA-I) and wheat germ (WGA) agglutinins as revealed by the binding specificities of these lectins towards mucin core-2 O-linked and N-linked glycans and related structures. Glycoconj. J. 2016, 33, 819–836. [Google Scholar]
- Debray, H.; Montreuil, J. Aleuria aurantia agglutinin. A new isolation procedure and further study of its specificity towards various glycopeptides and oligosaccharides. Carbohydr. Res. 1989, 185, 15–26. [Google Scholar] [CrossRef]
- Finne, J.; Krusius, T. Preparation and fractionation of glycopeptides. Methods Enzymol. 1982, 83, 269–277. [Google Scholar]
- Menon, M.; Mohanraj, R.; Vb, J.; Prasath Rv, A. Bioaccumulation of heavy metals in a gastropod species at the Kole wetland agroecosystem, a Ramsar site. J. Environ. Manag. 2023, 329, 117027. [Google Scholar] [CrossRef]
- Conti, M.E.; Tudino, M.; Muse, J.O.; Cecchetti, G. Biomonitoring of heavy metals and their species in the marine environment: The contribution of atomic absorption spectroscopy and inductively coupled plasma spectroscopy. Res. Trends Appl. Spectrosc. 2002, 4, 295–324. [Google Scholar]
- Dallinger, R.; Zerbe, O.; Baumann, C.; Egger, B.; Capdevila, M.; Palacios, O.; Albalat, R.; Calatayud, S.; Ladurner, P.; Schlick-Steiner, B.C.; et al. Metallomics reveals a persisting impact of cadmium on the evolution of metal-selective snail metallothioneins. Metallomics 2020, 12, 702–720. [Google Scholar] [CrossRef]
- Jebali, J.; Chouba, L.; Banni, M.; Boussetta, H. Comparative study of the bioaccumulation and elimination of trace metals (Cd, Pb, Zn, Mn and Fe) in the digestive gland, gills and muscle of bivalve Pinna nobilis during a field transplant experiment. J. Trace Elem. Med. Biol. 2014, 28, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Tanhan, P.; Imsilp, K.; Lansubsakul, N.; Tantiwisawaruji, S.; Thong-Asa, W. Oxidative response to accumulation of trace metals in tissue of two bivalves, the Asian green mussel Perna viridis and the blood cockle Tegillarca granosa, living in Pattani Bay, Thailand. J. Aquat. Anim. Health 2024, 36, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Podgurskaya, O.V.; Kavun, V.Y.; Lukyanova, O.N. Heavy Metal Accumulation and Distribution in Organs of the Mussel Crenomytilus grayanus from Upwelling Areas of the Sea of Okhotsk and the Sea of Japan. Russ. J. Mar. Biol. 2004, 30, 188–195. [Google Scholar] [CrossRef]
- Chahouri, A.; Yacoubi, B.; Moukrim, A.; Banaoui, A. Bivalve molluscs as bioindicators of multiple stressors in the marine environment: Recent advances. Contin. Shelf Res. 2023, 264, 105056. [Google Scholar] [CrossRef]
- De Roma, A.; Neola, B.; Serpe, F.P.; Sansone, D.; Picazio, G.; Cerino, P.; Esposito, M. Land snails (Helix aspersa) as bioindicators of trace element contamination in Campania (Italy). Open Access Libr. J. 2017, 4, 1–12. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).