
The Impact of Ekman Pumping and Transport on Dosidicus gigas (Jumbo Flying Squid) Fishing Ground by Chinese Jiggers off the Coast of Peru



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Fisheries and Environmental Data
2.2. Data Processing and Analysis
2.2.1. The Calculation of Ekman Transport and Pumping
2.2.2. Fishery Data Processing
2.2.3. Impact of Ekman Processes on the D. gigas Fishing Ground
2.2.4. The Impact of Ekman Processes on Chl-a Concentration
2.2.5. Generalized Additive Model (GAM)
2.2.6. The Impact of El Niño Events on Ekman Transport, Chlorophyll-a Concentrations, and D. gigas CPUE
3. Results
3.1. The Impact of Ekman Processes on the Abundance and Distribution of D. gigas
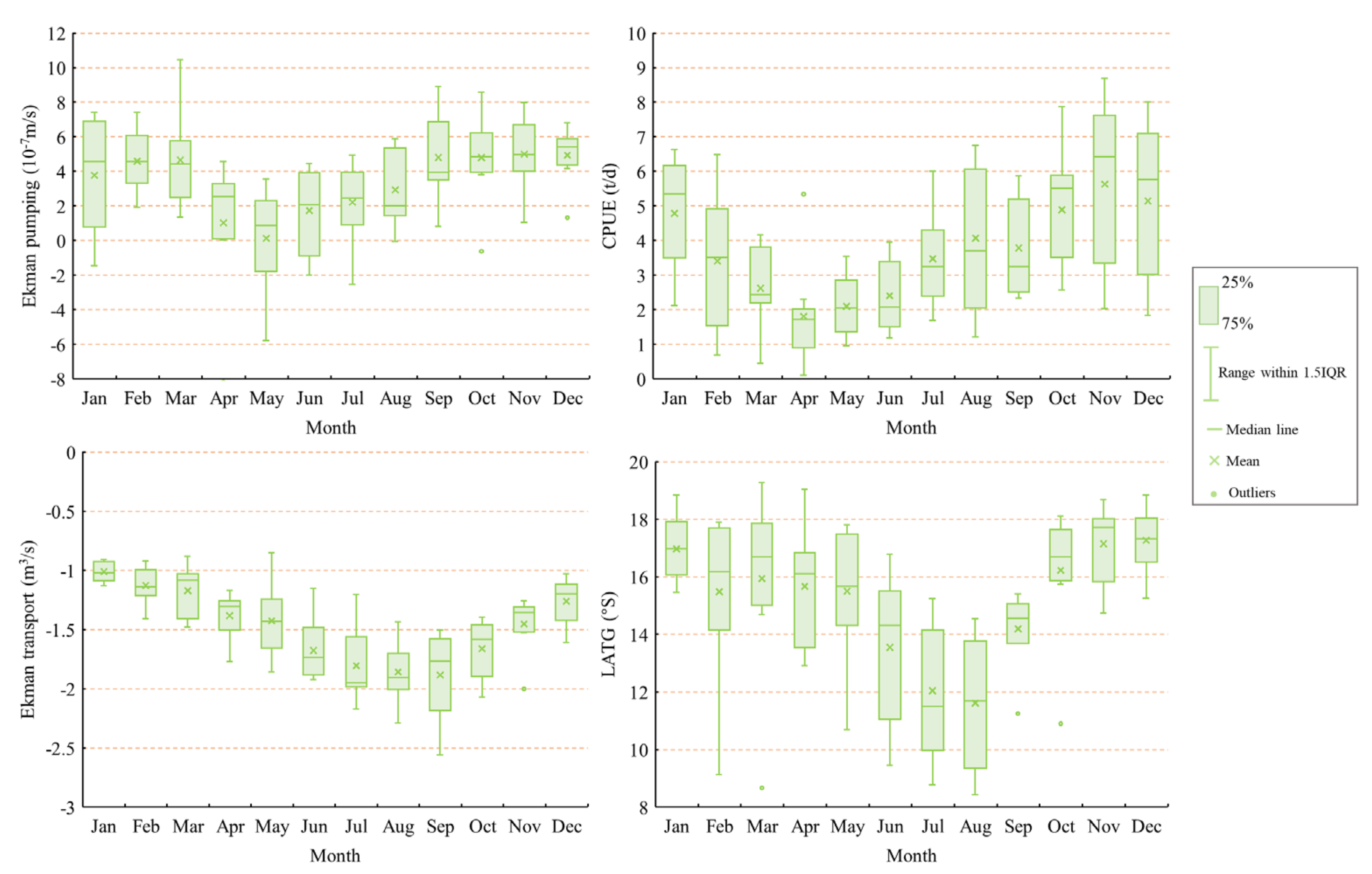
3.1.1. Monthly Variations in Ekman Pumping and Transport, and the CPUE and LATG of D. gigas
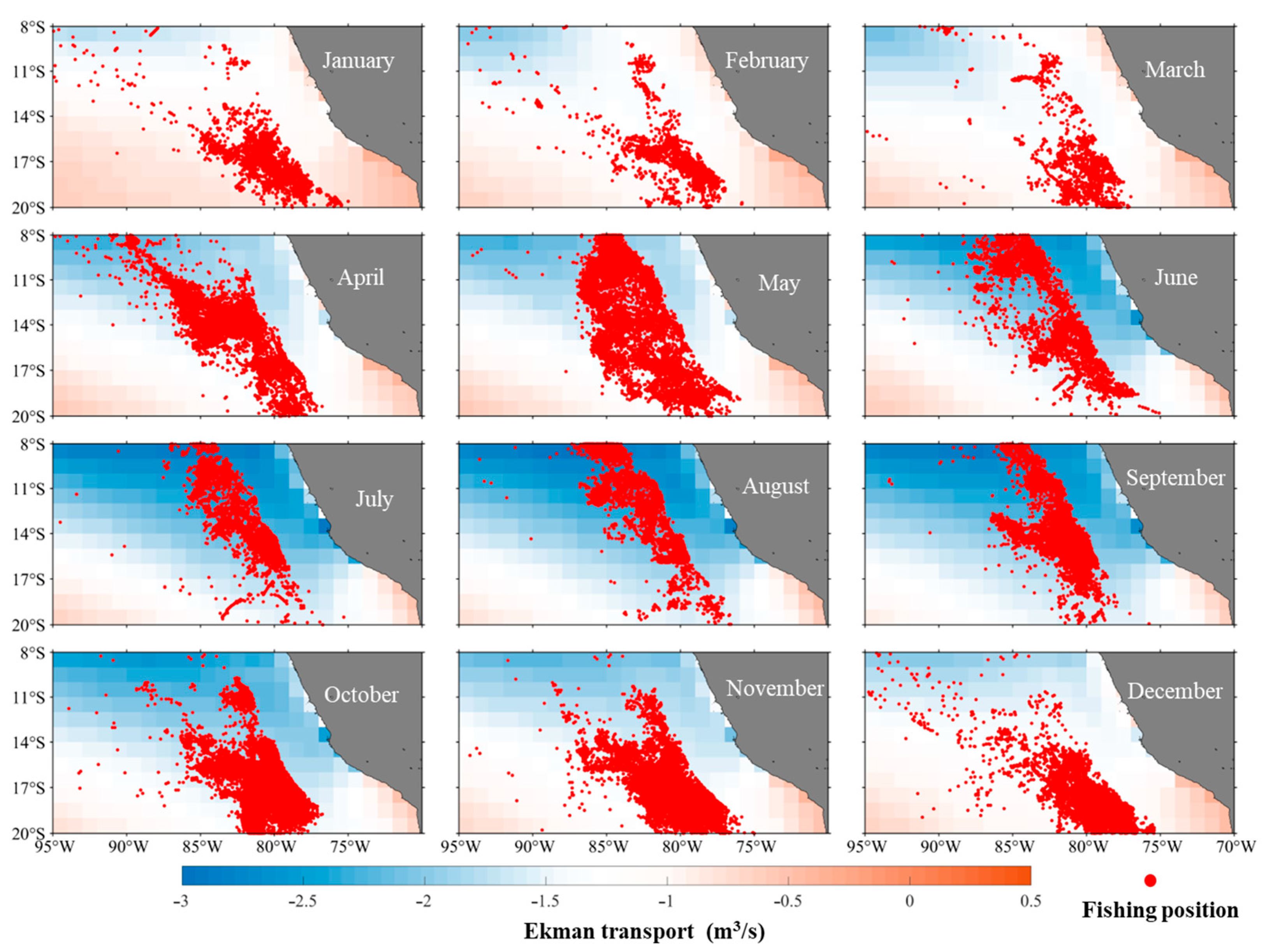
3.1.2. The Spatial Distribution Characteristics of Ekman Pumping
3.1.3. The Relationship Between the Spatial Distribution of Ekman Transport and D. gigas
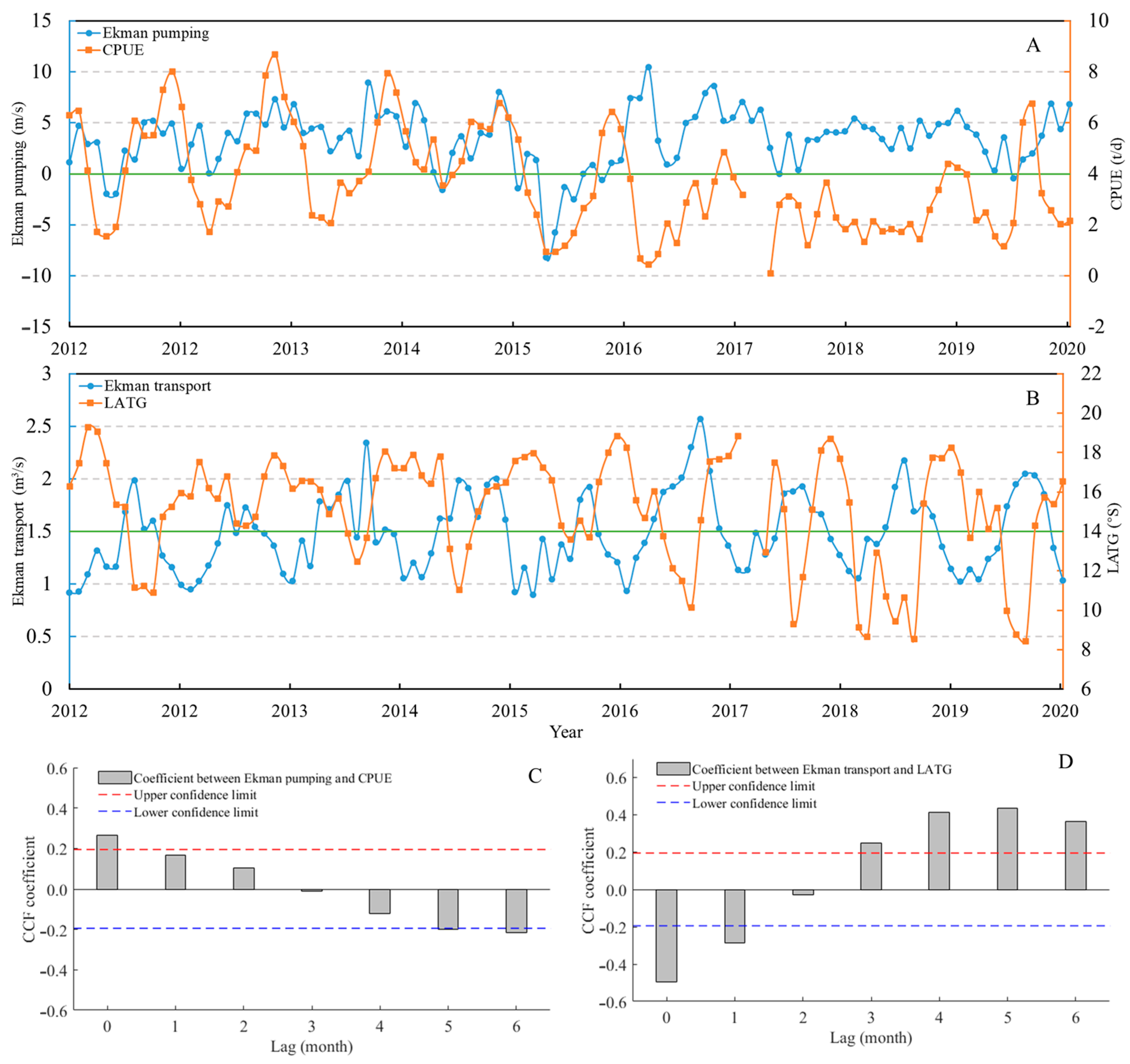
3.1.4. Annual Variation in Ekman Transport and Pumping, and the CPUE and LATG of D. gigas in 2012 to 2020
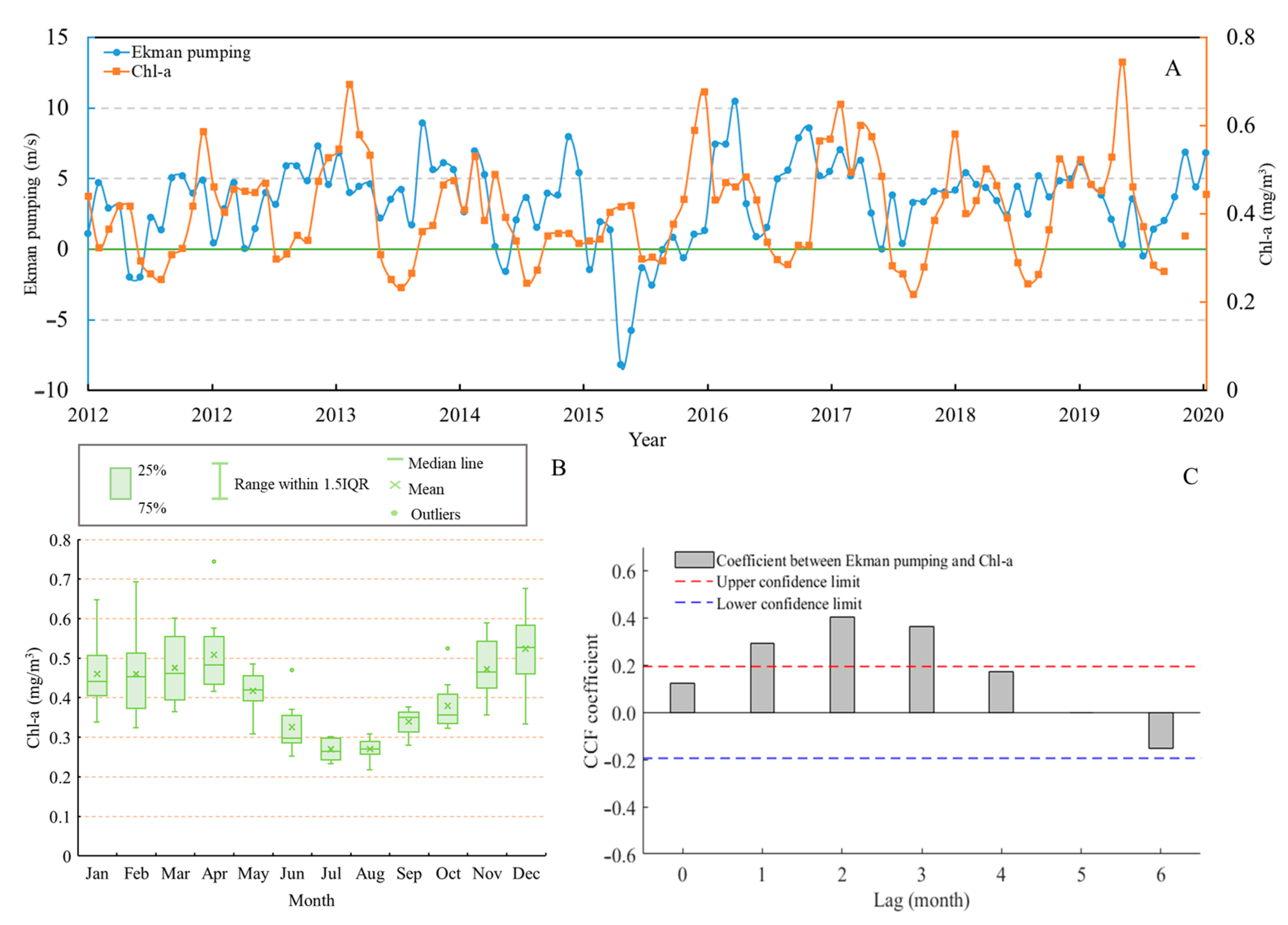
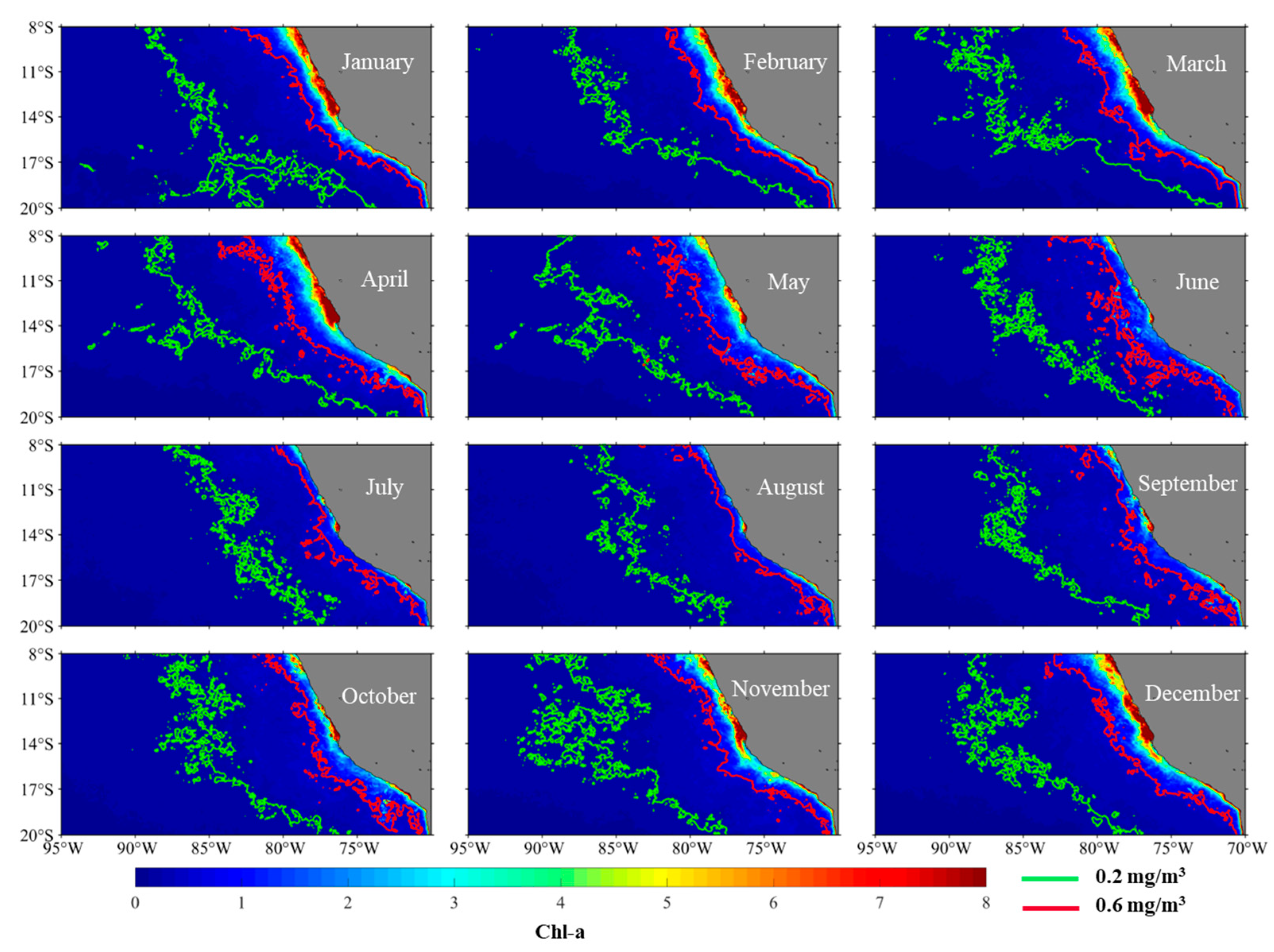
3.2. The Spatial Distribution Characteristics of Chl-a and Influence of Ekman Pumping on It
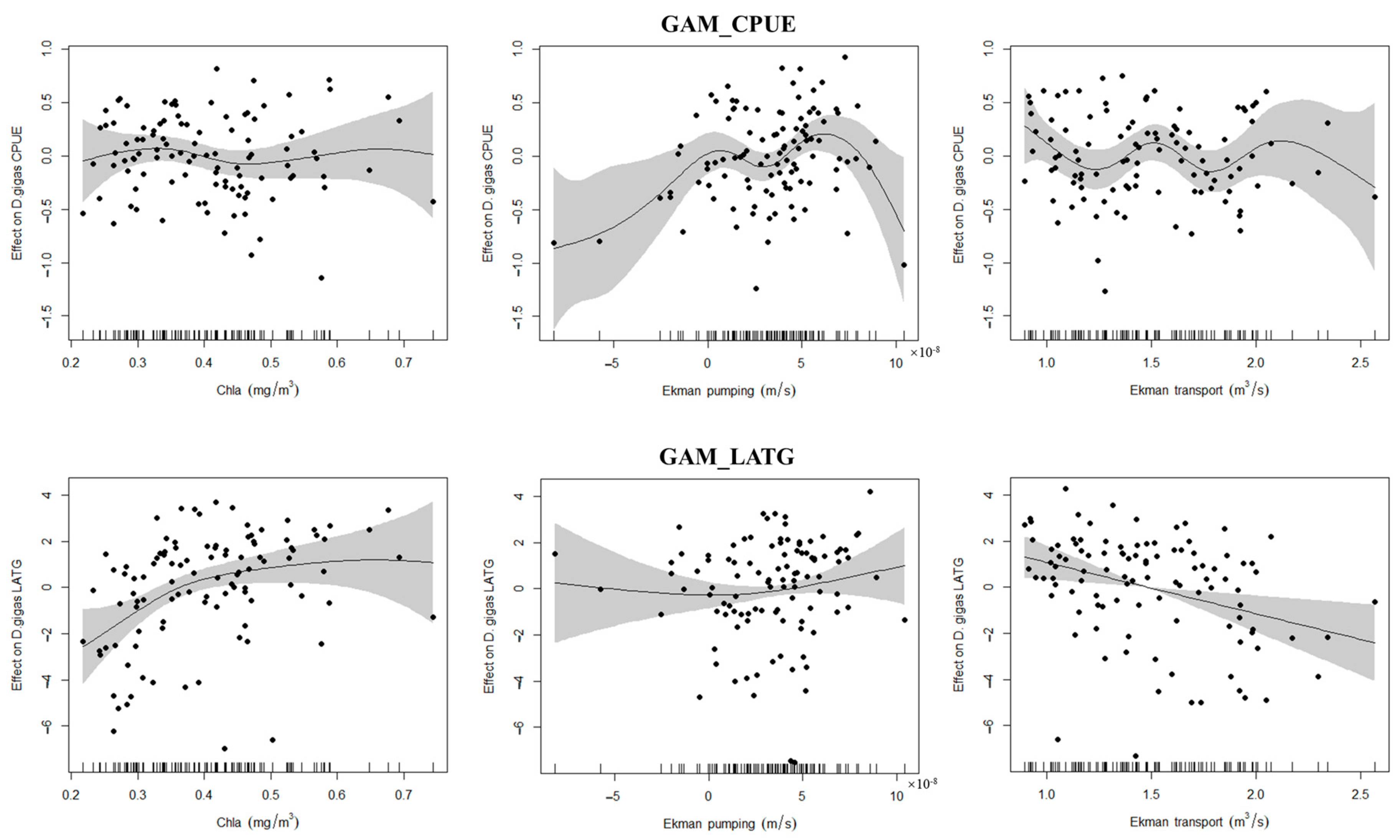
3.3. Relationship Between Ekman Pumping, Transport, and Chl-a on D. gigas CPUE and LATG
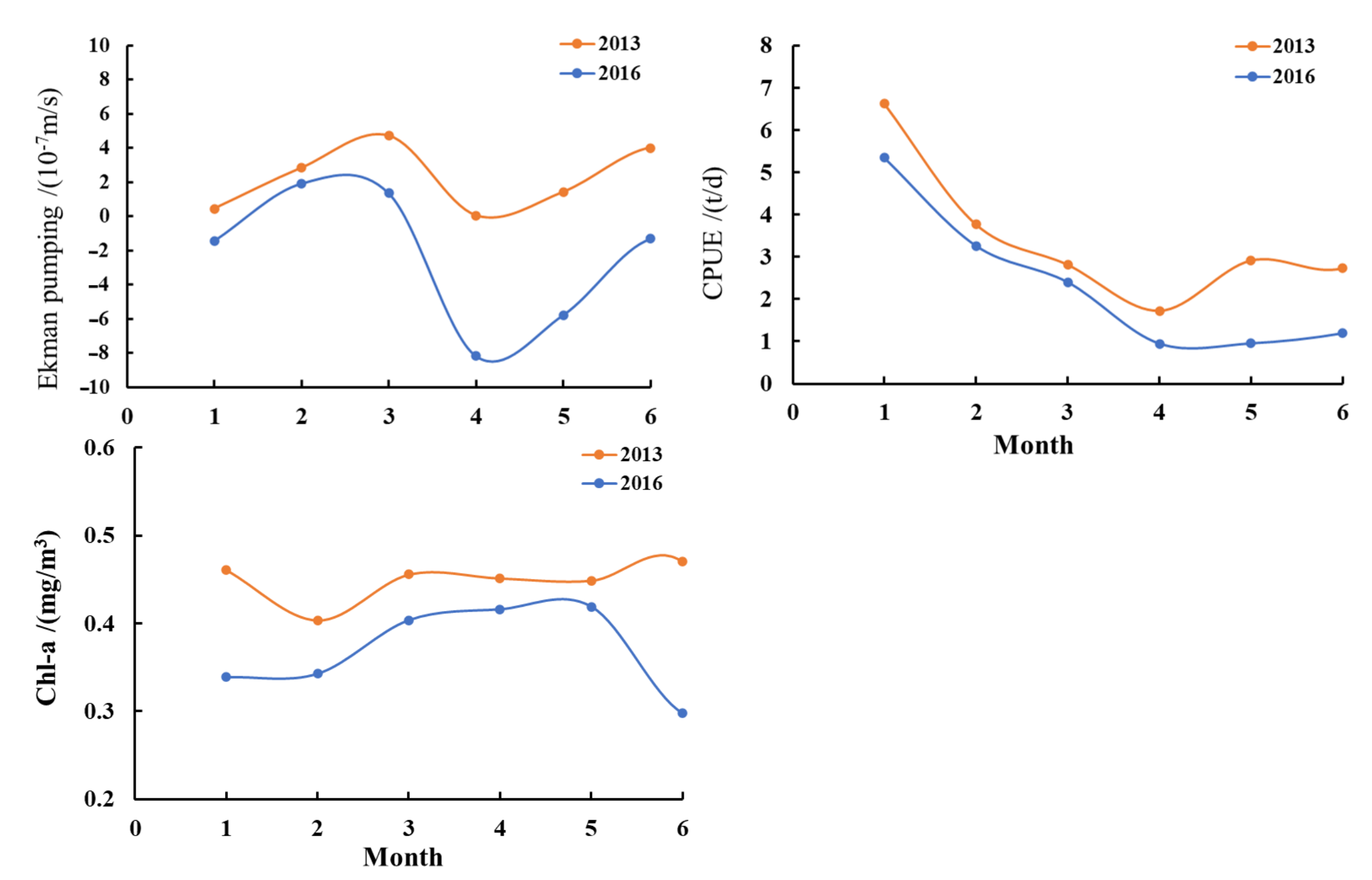
3.4. The Impact of El Niño on Ekman Pumping, Chl-a Concentration, and D. gigas CPUE
4. Discussion
4.1. Analysis of Ekman Processes and Their Effects on the Physical Marine Environment
4.2. The Effects of Ekman Processes on the D. gigas CPUE and LATG
4.3. Effects of Upwelling on Other Marine Organisms
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seo, H.; O’Neill, L.W.; Bourassa, M.A.; Czaja, A.; Drushka, K.; Edson, J.B.; Fox-Kemper, B.; Frenger, I.; Gille, S.T.; Kirtman, B.P.; et al. Ocean mesoscale and frontal-scale ocean–atmosphere interactions and influence on large-scale climate: A review. J. Clim. 2023, 36, 1981–2013. [Google Scholar] [CrossRef]
- Mahadevan, A. The impact of submesoscale physics on primary productivity of plankton. Annu. Rev. Mar. Sci. 2016, 8, 161–184. [Google Scholar] [CrossRef]
- McGillicuddy, D.J. Mechanisms of physical-biological-biogeochemical interaction at the oceanic mesoscale. Annu. Rev. Mar. Sci. 2016, 8, 125–159. [Google Scholar] [CrossRef]
- McGillicuddy, D.J.; Anderson, L.A.; Bates, N.R.; Bibby, T.; Buesseler, K.O.; Carlson, C.A.; Davis, C.S.; Falkowski, P.G.; Ewart, C.; Goldthwait, S.A.; et al. Eddy/Wind Interactions Stimulate Extraordinary Mid-Ocean Plankton Blooms. Deep-Sea Res. 2003, 1, 829. [Google Scholar] [CrossRef]
- Muñoz, R.; Vergara, O.A.; Figueroa, P.A.; Mardones, P.; Sobarzo, M.; Saldías, G.S. On the phenology of coastal upwelling off central-southern Chile. Dyn. Atmos. Ocean. 2023, 104, 101405. [Google Scholar] [CrossRef]
- Rossi, V.; Lopez, C.; Hernández-García, E.; Sudre, J.; Garçon, V.; Morel, Y. Surface mixing and biological activity in the four Eastern Boundary Upwelling Systems. Nonlinear Process. Geophys. 2009, 16, 557–568. [Google Scholar] [CrossRef]
- Chang, P.; Xu, G.; Kurian, J.; Small, R.; Danabasoglu, G.; Yeager, S.G.; Castruccio, F.S.; Zhang, Q.; Rosenbloom, N.A.; Chapman, P. Uncertain future of sustainable fisheries environment in eastern boundary upwelling zones under climate change. Commun. Earth Environ. 2023, 4, 19. [Google Scholar] [CrossRef]
- Chavez, F.P.; Messié, M. A comparison of eastern boundary upwelling ecosystems. Prog. Oceanogr. 2009, 83, 80–96. [Google Scholar] [CrossRef]
- Bograd, S.J.; Jacox, M.G.; Hazen, E.L.; Lovecchio, E.; Montes, I.; Pozo Buil, M.; Rykaczewski, R.R. Climate change impacts on eastern boundary upwelling systems. Annu. Rev. Mar. Sci. 2023, 15, 303–328. [Google Scholar] [CrossRef]
- Fréon, P.; Barange, M.; Arístegui, J. Eastern boundary upwelling ecosystems: Integrative and comparative approaches. Prog. Oceanogr. 2009, 83, 1–14. [Google Scholar] [CrossRef]
- Rykaczewski, R.R.; Checkley, D.M. Influence of ocean winds on the pelagic ecosystem in upwelling regions. Proc. Natl. Acad. Sci. USA 2008, 105, 1965–1970. [Google Scholar] [CrossRef] [PubMed]
- Cury, P.; Bakun, A.; Crawford, R.J.M.; Jarre, A.; Quinones, R.A.; Shannon, L.J.; Verheye, H.M. Small pelagics in upwelling systems: Patterns of interaction and structural changes in “wasp-waist” ecosystems. ICES J. Mar. Sci. 2000, 57, 603–618. [Google Scholar] [CrossRef]
- Montecino, V.; Lange, C.B. The Humboldt Current System: Ecosystem components and processes, fisheries, and sediment studies. Prog. Oceanogr. 2009, 83, 65–79. [Google Scholar] [CrossRef]
- Gutiérrez, D.; Akester, M.; Naranjo, L. Productivity and sustainable management of the Humboldt current large marine ecosystem under climate change. Environ. Dev. 2016, 17, 126–144. [Google Scholar] [CrossRef]
- Garteizgogeascoa, M.; Kluger, L.C.; Gonzales, I.E.; Damonte, G.; Flitner, M. Contextualizing scenarios to explore social-ecological futures: A three step participatory case study for the humboldt current upwelling system. Front. Mar. Sci. 2020, 7, 557181. [Google Scholar] [CrossRef]
- Albert, A.; Echevin, V.; Lévy, M.; Aumont, O. Impact of nearshore wind stress curl on coastal circulation and primary productivity in the Peru upwelling system. J. Geophys. Res. Ocean. 2010, 115(C12), C12033. [Google Scholar] [CrossRef]
- Halpern, D. Offshore Ekman transport and Ekman pumping off Peru during the 1997–1998 El Nino. Geophys. Res. Lett. 2002, 29, 19-1–19-4. [Google Scholar] [CrossRef]
- Pickett, M.H.; Paduan, J.D. Ekman transport and pumping in the California Current based on the US Navy’s high-resolution atmospheric model (COAMPS). J. Geophys. Res. Ocean. 2003, 108, 3327. [Google Scholar] [CrossRef]
- Wirasatriya, A.; Setiawan, J.; Sugianto, D.; Rosyadi, I.; Haryadi, H.; Winarso, G.; Setiawan, R.; Susanto, R. Ekman dynamics variability along the southern coast of Java revealed by satellite data. Int. J. Remote Sens. 2020, 41, 8475–8496. [Google Scholar] [CrossRef]
- Jacox, M.G.; Edwards, C.A.; Hazen, E.L.; Bograd, S.J. Coastal upwelling revisited: Ekman, Bakun, and improved upwelling indices for the US West Coast. J. Geophys. Res. Ocean. 2018, 123, 7332–7350. [Google Scholar] [CrossRef]
- Combes, V.C.F.D.L.E.; Chenillat, F.; Di Lorenzo, E.; Rivière, P.; Ohman, M.D.; Bograd, S.J. Cross-shore transport variability in the California Current: Ekman upwelling vs. eddy dynamics. Prog. Oceanogr. 2013, 109, 78–89. [Google Scholar] [CrossRef]
- Sachin, T.S.; Rasheed, K. Dynamics and forcing mechanisms of upwelling along the south eastern Arabian sea during south west monsoon. Reg. Stud. Mar. Sci. 2020, 40, 101519. [Google Scholar] [CrossRef]
- Kumar, J.; Ratheesh, S.; Agarwal, N.; Sharma, R. Study of upwelling and mixing process in the Somali coastal region using satellite and numerical model observations: A Lagrangian approach. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2024, 216, 105381. [Google Scholar] [CrossRef]
- Simanjuntak, F.; Lin, T.H. Monsoon effects on chlorophyll-a, sea surface temperature, and ekman dynamics variability along the southern coast of lesser Sunda islands and its relation to ENSO and IOD based on satellite observations. Remote Sens. 2022, 14, 1682. [Google Scholar] [CrossRef]
- Belmadani, A.; Echevin, V.; Codron, F.; Takahashi, K.; Junquas, C. What dynamics drive future wind scenarios for coastal upwelling off Peru and Chile? Clim. Dyn. 2014, 43, 1893–1914. [Google Scholar] [CrossRef]
- Wu, X.C.; Jin, P.C.; Zhang, Y.; Yu, W. Spatial Distribution and Abundance of a Pelagic Squid during the Evolution of Eddies in the Southeast Pacific Ocean. J. Mar. Sci. Eng. 2024, 12, 1015. [Google Scholar] [CrossRef]
- Nigmatullin, C.M.; Nesis, K.N.; Arkhipkin, A.I. A review of the biology of the jumbo squid Dosidicus gigas (Cephalopoda: Ommastrephidae). Fish. Res. 2001, 54, 9–19. [Google Scholar] [CrossRef]
- Yu, W.; Chen, X.J. Ocean warming-induced range-shifting of potential habitat for jumbo flying squid Dosidicus gigas in the southeast pacific ocean off peru. Fish. Res. 2018, 204, 137–146. [Google Scholar] [CrossRef]
- Yu, W.; Yi, Q.; Chen, X.J.; Chen, Y. Modelling the effects of climate variability on habitat suitability of jumbo flying squid, Dosidicus gigas, in the Southeast Pacific Ocean off Peru. ICES J. Mar. Sci. 2016, 73, 239–249. [Google Scholar] [CrossRef]
- Fang, X.N.; Zhang, Y.; Yu, W.; Chen, X.J. Geographical distribution variations of Humboldt squid habitat in the Eastern Pacific Ocean. Ecosyst. Health Sustain. 2023, 9, 0010. [Google Scholar] [CrossRef]
- Anderson, C.I.H.; Rodhouse, P.G. Life cycles, oceanography and variability: Ommastrephid squid in variable oceanographic environments. Fish. Res. 2001, 54, 133–143. [Google Scholar] [CrossRef]
- Wu, X.C.; Jin, P.C.; Zhang, Y.; Yu, W. Changing Humboldt Squid Abundance and Distribution at Different Stages of Oceanic Mesoscale Eddies. J. Mar. Sci. Eng. 2024, 12, 626. [Google Scholar] [CrossRef]
- Jin, P.C.; Fan, J.T.; Wu, X.C.; Du, Y.L.; Yu, W. Jumbo flying squid distribution within anticyclonic eddies under anomalous climatic conditions. Ecosyst. Health Sustain. 2024, 10, 0177. [Google Scholar] [CrossRef]
- Fang, X.N.; Yu, W.; Chen, X.J.; Zhang, Y. Response of abundance and distribution of Humboldt squid (Dosidicus gigas) to short-Lived eddies in the eastern equatorial Pacific Ocean from April to June 2017. Front. Mar. Sci. 2021, 8, 721291. [Google Scholar] [CrossRef]
- Tiedemann, M.; Brehmer, P. Larval fish assemblages across an upwelling front: Indication for active and passive retention. Estuar. Coast. Shelf Sci. 2017, 187, 118–133. [Google Scholar] [CrossRef]
- Sato, M.; Barth, J.A.; Benoit-Bird, K.J.; Pierce, S.D.; Cowles, T.J.; Brodeur, R.D.; Peterson, W.T. Coastal upwelling fronts as a boundary for planktivorous fish distributions. Mar. Ecol. Prog. Ser. 2018, 595, 171–186. [Google Scholar] [CrossRef]
- Reese, D.C.; O’Malley, R.T.; Brodeur, R.D.; Churnside, J.H. Epipelagic fish distributions in relation to thermal fronts in a coastal upwelling system using high-resolution remote-sensing techniques. ICES J. Mar. Sci. 2011, 68, 1865–1874. [Google Scholar] [CrossRef]
- Subarna, D. The effect of monsoon variability on fish landing in the Sadeng Fishing Port of Yogyakarta, Indonesia. IOP Conf. Ser. Earth Environ. Sci. 2018, 139, 012027. [Google Scholar] [CrossRef]
- Villanoy, C.L.; Cabrera, O.C.; Yniguez, A.; Camoying, M.; de Guzman, A.; David, L.T.; Flament, P. Monsoon-driven coastal upwelling off Zamboanga Peninsula, Philippines. Oceanography 2011, 24, 156–165. [Google Scholar] [CrossRef]
- Menon, N.N.; Sankar, S.; Smitha, A.; George, G.; Shalin, S.; Sathyendranath, S.; Platt, T. Satellite chlorophyll concentration as an aid to understanding the dynamics of Indian oil sardine in the southeastern Arabian Sea. Mar. Ecol. Prog. Ser. 2019, 617, 137–147. [Google Scholar] [CrossRef]
- Van Der Sleen, P.; Rykaczewski, R.R.; Turley, B.D.; Sydeman, W.J.; García-Reyes, M.; Bograd, S.J.; Van Der Lingen, C.D.; Coetzee, J.C.; Lamont, T.; Black, B.A. Non-stationary responses in anchovy (Engraulis encrasicolus) recruitment to coastal upwelling in the Southern Benguela. Mar. Ecol. Prog. Ser. 2018, 596, 155–164. [Google Scholar] [CrossRef]
- Sonderblohm, C.P.; Pereira, J.S.; Erzini, K. Environmental and fishery-driven dynamics of the common octopus (Octopus vulgaris) based on time-series analyses from leeward Algarve, southern Portugal. ICES J. Mar. Sci. 2014, 71, 2231–2241. [Google Scholar] [CrossRef]
- Echevin, V.; Puillat, I.; Grados, C.; Dewitte, B. Seasonal and mesoscale variability in the Peru upwelling system from in situ data during the years 2000 to 2004. Gayana 2004, 68, 167–173. [Google Scholar] [CrossRef]
- Croquette, M.; Eldin, G.; Echevin, V. On the contributions of Ekman transport and pumping to the dynamics of coastal upwelling in the South-East Pacific. Gayana 2004, 68, 136–141. [Google Scholar] [CrossRef]
- Aravena, G.; Broitman, B.; Stenseth, N.C. Twelve years of change in coastal upwelling along the central-northern coast of Chile: Spatially heterogeneous responses to climatic variability. PLoS ONE 2014, 9, e90276. [Google Scholar] [CrossRef]
- Escribano, R.; Hidalgo, P.; Fuentes, M.; Donoso, K. Zooplankton time series in the coastal zone off Chile: Variation in upwelling and responses of the copepod community. Prog. Oceanogr. 2012, 97, 174–186. [Google Scholar] [CrossRef]
- Oerder, V.; Colas, F.; Echevin, V.; Codron, F.; Tam, J.; Belmadani, A. Peru-chile upwelling dynamics under climate change. J. Geophys. Res. Ocean. 2015, 120, 1152–1172. [Google Scholar] [CrossRef]
- Ramajo, L.; Valladares, M.; Astudillo, O.; Fernández, C.; Rodríguez-Navarro, A.B.; Watt-Arévalo, P.; Núñez, M.; Grenier, C.; Román, R.; Aguayo, P.; et al. Upwelling intensity modulates the fitness and physiological performance of coastal species: Implications for the aquaculture of the scallop Argopecten purpuratus in the Humboldt Current System. Sci. Total Environ. 2020, 745, 140949. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, X.; Zhang, X.; Chen, X.; Yu, W. The Impact of Ekman Pumping and Transport on Dosidicus gigas (Jumbo Flying Squid) Fishing Ground by Chinese Jiggers off the Coast of Peru. J. Mar. Sci. Eng. 2025, 13, 280. https://doi.org/10.3390/jmse13020280
Fang X, Zhang X, Chen X, Yu W. The Impact of Ekman Pumping and Transport on Dosidicus gigas (Jumbo Flying Squid) Fishing Ground by Chinese Jiggers off the Coast of Peru. Journal of Marine Science and Engineering. 2025; 13(2):280. https://doi.org/10.3390/jmse13020280
Chicago/Turabian StyleFang, Xingnan, Xin Zhang, Xinjun Chen, and Wei Yu. 2025. "The Impact of Ekman Pumping and Transport on Dosidicus gigas (Jumbo Flying Squid) Fishing Ground by Chinese Jiggers off the Coast of Peru" Journal of Marine Science and Engineering 13, no. 2: 280. https://doi.org/10.3390/jmse13020280
APA StyleFang, X., Zhang, X., Chen, X., & Yu, W. (2025). The Impact of Ekman Pumping and Transport on Dosidicus gigas (Jumbo Flying Squid) Fishing Ground by Chinese Jiggers off the Coast of Peru. Journal of Marine Science and Engineering, 13(2), 280. https://doi.org/10.3390/jmse13020280