
Quantitative Real-Time Polymerase Chain Reaction (PCR) Assay for Rapid Monitoring of the Harmful Algal Bloom Species Cochlodinium polykrikoides

 ,
,  ,
,  , ,
, , 

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Culture of Cochlodinium polykrikoides
2.2. Design of Cochlodinium polykrikoides-Specific Primers
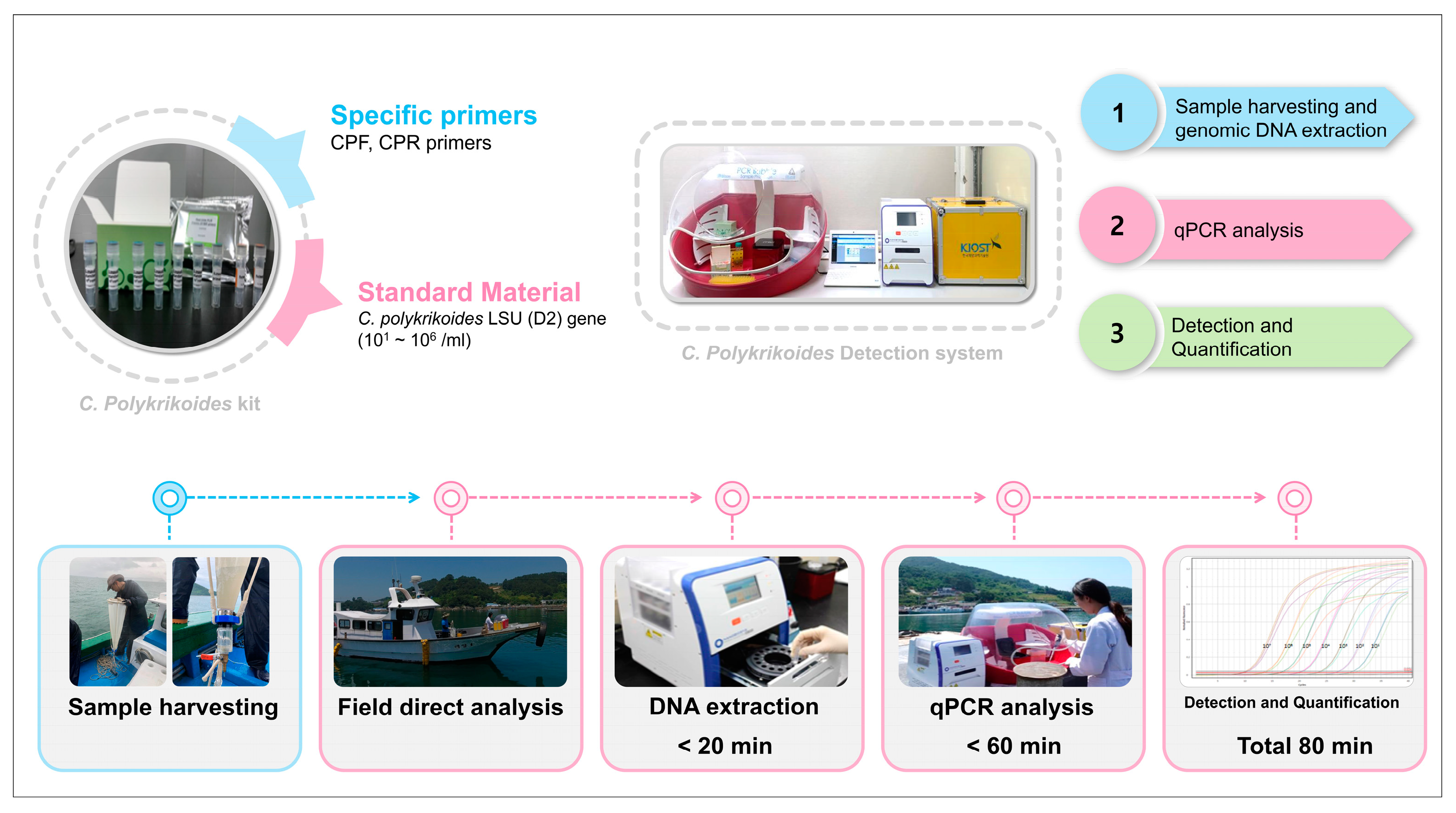
2.3. Development of LSU Standard Materials for Cochlodinium polykrikoides
2.4. Development of a Cochlodinium polykrikoides Detection System
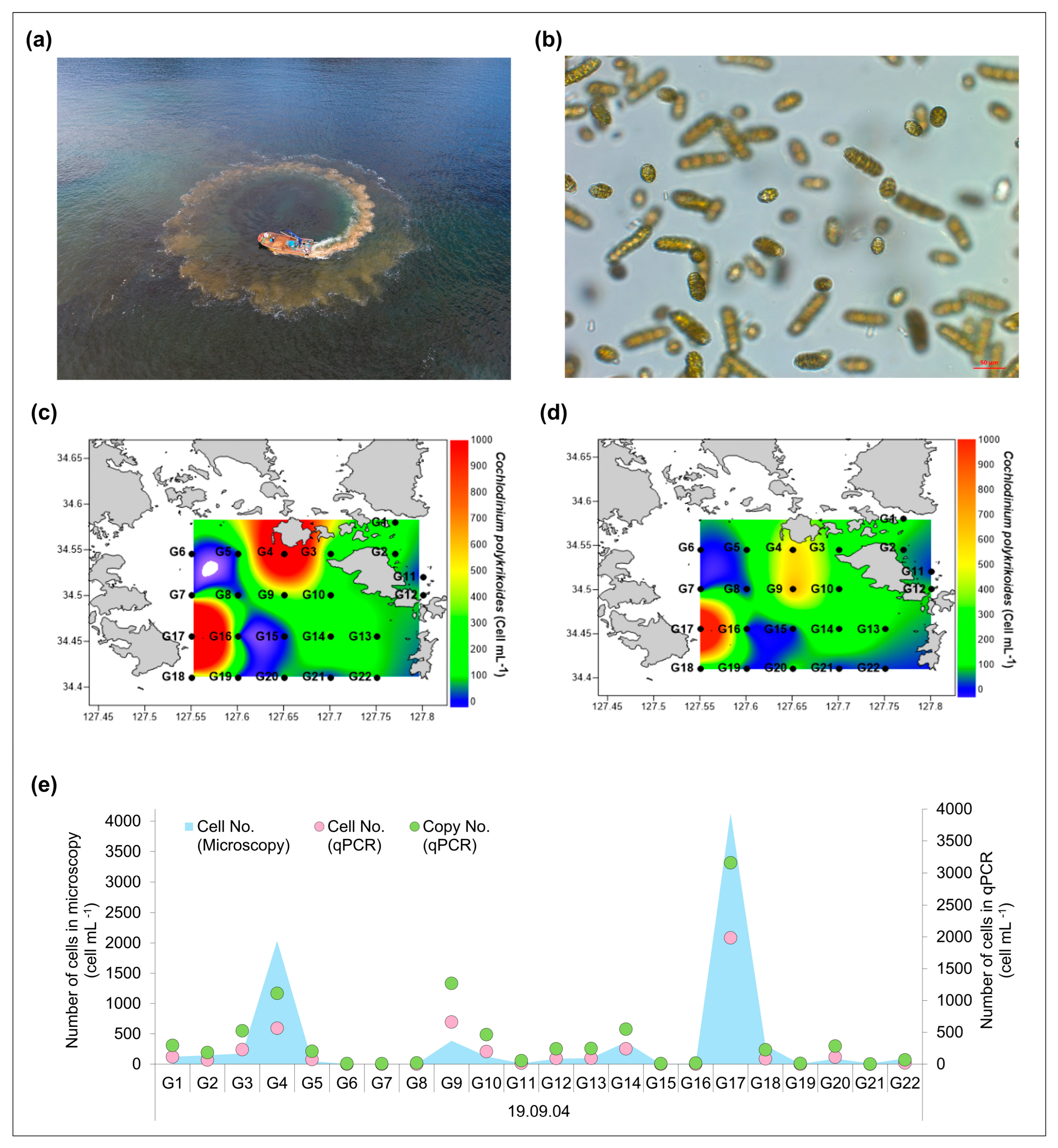
2.5. Field Application of the Cochlodinium polykrikoides Detection System
3. Results and Discussion
3.1. Evaluation of the Cochlodinium polykrikoides-Specific Primers
3.2. Verification Through Field Application
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ternon, E.; Pavaux, A.-S.; Marro, S.; Thomas, O.P.; Lemée, R. Allelopathic interactions between the benthic toxic dinoflagellate Ostreopsis cf. ovata and a co-occurring diatom. Harmful Algae 2018, 75, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Gobler, C.J. Climate change and harmful algal blooms: Insights and perspective. Harmful Algae 2020, 91, 101731. [Google Scholar] [CrossRef] [PubMed]
- Heil, C.A.; Muni-Morgan, A.L. Florida’s harmful algal bloom (HAB) problem: Escalating risks to human, environmental and economic health with climate change. Front Ecol. Evol. 2021, 9, 646080. [Google Scholar] [CrossRef]
- Lee, C.-K.; Park, T.-G.; Park, Y.-T.; Lim, W.-A. Monitoring and trends in harmful algal blooms and red tides in Korean coastal waters, with emphasis on Cochlodinium polykrikoides. Harmful Algae 2013, 30, S3–S14. [Google Scholar] [CrossRef]
- Park, T.G.; Lim, W.A.; Park, Y.T.; Lee, C.K.; Jeong, H.J. Economic impact, management and mitigation of red tides in Korea. Harmful Algae 2013, 30, S131–S143. [Google Scholar] [CrossRef]
- Lee, Y.-C.; Jin, E.; Jung, S.W.; Kim, Y.-M.; Chang, K.S.; Yang, J.-W.; Kim, S.-W.; Kim, Y.-O.; Shin, H.-J. Utilizing the algicidal activity of aminoclay as a practical treatment for toxic red tides. Sci. Rep. 2013, 3, 1292. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; Jeoung, G.; Kim, K.E.; Park, J.S.; Kang, D.; Baek, S.H.; Lee, C.Y.; Kim, H.; Cho, S.; Lee, T.-K.; et al. Co-variance between free-living bacteria and Cochlodinium polykrikoides (Dinophyta) harmful algal blooms, South Korea. Harmful Algae 2023, 122, 102371. [Google Scholar] [CrossRef]
- Kang, D.; Kim, B.K.; Jung, S.W.; Baek, S.H.; Choi, J.-Y.; Cho, H.-Y.; Lee, S.-J.; Kim, H. Development and application of an integrated system for the detection and prediction of harmful algal blooms in Korea. J. Mar. Sci. Eng. 2023, 11, 2207. [Google Scholar] [CrossRef]
- LeGresley, M.; McDermott, G. Counting Chamber Methods for Quantitative Phytoplankton Analysis-Haemocytometer, Palmer-Maloney Cell and Sedgewick-Rafter Cell. In United Nations Educational, Scientific and Cultural Organization (IOC Manuals and Guides); UNESCO: Paris, France, 2010; Volume 55, pp. 25–30. [Google Scholar]
- Antonella, P.; Luca, G. The quantitative real-time PCR applications in the monitoring of marine harmful algal bloom (HAB) species. Environ. Sci. Pollut. Res. 2013, 20, 6851–6862. [Google Scholar] [CrossRef] [PubMed]
- Karlson, B.; Cusack, C.; Bresnan, E. (Eds.) Microscopic and Molecular Methods for Quantitative Phytoplankton Analysis; UNESCO: Paris, France, 2010; p. 110. [Google Scholar]
- Medlin, L. Molecular tools for monitoring harmful algal blooms. Environ. Sci. Pollut. Res. Int. 2013, 20, 6683–6685. [Google Scholar] [CrossRef]
- Lee, H.-G.; Kim, H.M.; Min, J.; Park, C.; Jeong, H.J.; Lee, K.; Kim, K.Y. Quantification of the paralytic shellfish poisoning dinoflagellate Alexandrium species using a digital PCR. Harmful Algae 2020, 92, 101726. [Google Scholar] [CrossRef] [PubMed]
- Feist, S.M.; Lance, R.F. Genetic detection of freshwater harmful algal blooms: A review focused on the use of environmental DNA (eDNA) in Microcystis aeruginosa and Prymnesium parvum. Harmful Algae 2021, 110, 102124. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Y.; Lai, J.; Li, J.; Yu, K. Improvement and application of qPCR assay revealed new insight on early warning of Phaeocystis globosa bloom. Water Res. 2023, 229, 119439. [Google Scholar] [CrossRef]
- Kim, W.; Han, T.; Jung, S.; Kang, D. Analysis on the Optical Absorption Property of Sea Waters Dominated by Alexandrium affine in Coastal Waters off Tongyeong, 2017. Photogramm. Cartogr. 2019, 37, 563–570. [Google Scholar]
- Choi, J.K.; Min, J.E.; Noh, J.H.; Han, T.H.; Yoon, S.; Park, Y.J.; Park, J.H. Harmful algal bloom (HAB) in the East Sea identified by the Geostationary Ocean Color Imager (GOCI). Harmful Algae 2014, 39, 295–302. [Google Scholar] [CrossRef]
- Dizaji, S.Z.; Fariman, G.A.; Zahedi, M.M. Pigment content analysis in two HAB forming dinoflagellate species during the growth period. J. Appl. Phycol. 2021, 33, 807–817. [Google Scholar] [CrossRef]
- Medlin, L.K.; Orozco, J. Molecular techniques for the detection of organisms in aquatic environments, with emphasis on harmful algal bloom species. Sensors 2017, 17, 1184. [Google Scholar] [CrossRef]
- Hernández-López, E.L.; Gasperin, J.; Bernáldez-Sarabia, J.; Licea-Navarro, A.F.; Guerrero, A.; Lizárraga-Partida, M.L. Detection of Alcanivorax spp., Cycloclasticus spp., and Methanomicrobiales in water column and sediment samples in the Gulf of Mexico by qPCR. Environ. Sci. Pollut. Res. Int. 2019, 26, 35131–35139. [Google Scholar] [CrossRef]
- Engesmo, A.; Strand, D.; Gran-Stadniczeñko, S.; Edvardsen, B.; Medlin, L.K.; Eikrem, W. Development of a qPCR assay to detect and quantify ichthyotoxic flagellates along the Norwegian coast, and the first Norwegian record of Fibrocapsa japonica (Raphidophyceae). Harmful Algae 2018, 75, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Hatfield, R.G.; Bean, T.; Turner, A.D.; Lees, D.N.; Lowther, J.; Lewis, A.; Baker-Austin, C. Development of a TaqMan qPCR assay for detection of Alexandrium spp. and application to harmful algal bloom monitoring. Toxicon X 2019, 2, 100011. [Google Scholar] [CrossRef]
- Jung, S.W.; Joo, H.M.; Park, J.S.; Lee, J.H. Development of a rapid and effective method for preparing delicate dinoflagellates for scanning electron microscopy. J. Appl. Phycol. 2010, 22, 313–317. [Google Scholar] [CrossRef]
- Joanes, D.N.; Gill, C.A. Comparing measures of sample skewness and kurtosis Soc Ser D-Stat 47(1). J. R. Stat. Soc. Ser. D 1998, 47, 183–189. [Google Scholar] [CrossRef]
- Dhar, S.S.; Chakraborty, B.; Chaudhuri, P. Comparison of multivariate distributions using quantile–quantile plots and related tests. Bernoulli 2014, 20, 1484–1506. [Google Scholar] [CrossRef]
- Jung, S.W.; Kang, D.; Kim, H.-J.; Shin, H.H.; Park, J.S.; Park, S.Y.; Lee, T.-K. Mapping distribution of cysts of recent dinoflagellate and Cochlodinium polykrikoides using next-generation sequencing and morphological approaches in South Sea, Korea. Sci. Rep. 2018, 8, 7011. [Google Scholar] [CrossRef] [PubMed]
- Park, T.G.; Kim, J.J.; Kim, W.J.; Won, K.M. Development of real-time RT-PCR for detecting viable Cochlodinium polykrikoides (Dinophyceae) cysts in sediment. Harmful Algae 2016, 60, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Litaker, R.W.; Vandersea, M.W.; Kibler, S.R.; Reece, K.S.; Stokes, N.A.; Steidinger, K.A.; Millie, D.F.; Bendis, B.J.; Pigg, R.J.; Tester, P.A. Identification of Pfiesteria piscicida (Dinophyceae) and Pfiesteria-like organisms using internal transcribed spacer-specific pcr assays 1. J. Phycol. 2003, 39, 754–761. [Google Scholar] [CrossRef]
- Sonnenberg, R.; Nolte, A.W.; Tautz, D. An evaluation of LSU rDNA D1-D2 sequences for their use in species identification. Front. Zool. 2007, 4, 6. [Google Scholar] [CrossRef] [PubMed]
- Park, T.G.; Park, Y.T.; Lee, Y. Development of a SYTO9 based real-time PCR probe for detection and quantification of toxic dinoflagellate Karlodinium veneficum (Dinophyceae) in environmental samples. Phycologia 2009, 48, 32–43. [Google Scholar] [CrossRef]
- Handy, S.M.; Demir, E.; Hutchins, D.A.; Portune, K.J.; Whereat, E.B.; Hare, C.E.; Rose, J.M.; Warner, M.; Farestad, M.; Cary, S.C.; et al. Using quantitative real-time PCR to study competition and community dynamics among Delaware Inland Bays harmful algae in field and laboratory studies. Harmful Algae 2008, 7, 599–613. [Google Scholar] [CrossRef]
- Kim, C.-J.; Kim, H.-G.; Kim, C.-H.; Oh, H.-M. Life cycle of the ichthyotoxic dinoflagellate Cochlodinium polykrikoides in Korean coastal waters. Harmful Algae 2007, 6, 104–111. [Google Scholar] [CrossRef]
- Shin, H.H.; Li, Z.; Yoon, Y.H.; Oh, S.J.; Lim, W.A. Formation and germination of temporary cysts of Cochlodinium polykrikoides Margalef (Dinophyceae) and their ecological role in dense blooms. Harmful Algae 2017, 66, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wang, S.; Du, X.; He, Q.; Liu, Y.; Wang, Z.; Feng, K.; Li, Y.; Deng, Y. ddPCR surpasses classical qPCR technology in quantitating bacteria and fungi in the environment. Mol. Ecol. Resour. 2022, 22, 2587–2598. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.-K.; Lin, Z.-R.; Zhang, Q.-C.; Kong, F.-Z.; Cen, J.-Y.; Zeng, Y.-L.; Yu, R.-C. Detection of bloom-forming dinoflagellates Karenia mikimotoi and Prorocentrum donghaiense using qPCR assays. J. Appl. Phycol. 2022, 34, 1483–1496. [Google Scholar] [CrossRef]
- Gałuszka, A.; Migaszewski, Z.M.; Namieśnik, J. Moving your laboratories to the field–Advantages and limitations of the use of field portable instruments in environmental sample analysis. Environ. Res. 2015, 140, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.G.; Kim, H.M.; Min, J.; Kim, K.; Park, M.G.; Jeong, H.J.; Kim, K.Y. An advanced tool, droplet digital PCR (ddPCR), for absolute quantification of the red-tide dinoflagellate, Cochlodinium polykrikoides Margalef (Dinophyceae). Algae 2017, 32, 189–197. [Google Scholar] [CrossRef]
- Zowawi, H.M.; Alenazi, T.H.; AlOmaim, W.S.; Wazzan, A.; Alsufayan, A.; Hasanain, R.A.; Aldibasi, O.S.; Althawadi, S.; Altamimi, S.A.; Mutabagani, M.; et al. Portable RT-PCR system: A rapid and scalable diagnostic tool for COVID-19 testing. J. Clin. Microbiol. 2021, 59, 10.1128. [Google Scholar] [CrossRef]
- Spikmans, V. The evolution of environmental forensics: From laboratory to field analysis. WIREs Forensic Sci. 2019, 1, e1334. [Google Scholar] [CrossRef]
- Lepej, S.Z.; Poljak, M. Portable molecular diagnostic instruments in microbiology: Current status. Clin. Microbiol. Infect. 2020, 26, 411–420. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.-J.; Kim, H.-J.; Park, J.S.; Kang, D.; Cho, S.; Kim, H.; Baek, S.H.; Park, J.J.C.; Han, J.; Kim, K.E.; et al. Quantitative Real-Time Polymerase Chain Reaction (PCR) Assay for Rapid Monitoring of the Harmful Algal Bloom Species Cochlodinium polykrikoides. J. Mar. Sci. Eng. 2025, 13, 277. https://doi.org/10.3390/jmse13020277
Kim M-J, Kim H-J, Park JS, Kang D, Cho S, Kim H, Baek SH, Park JJC, Han J, Kim KE, et al. Quantitative Real-Time Polymerase Chain Reaction (PCR) Assay for Rapid Monitoring of the Harmful Algal Bloom Species Cochlodinium polykrikoides. Journal of Marine Science and Engineering. 2025; 13(2):277. https://doi.org/10.3390/jmse13020277
Chicago/Turabian StyleKim, Min-Jeong, Hyun-Jung Kim, Joon Sang Park, Donhyug Kang, Sungho Cho, Hansoo Kim, Seung Ho Baek, Jordan Jun Chul Park, Jeonghoon Han, Kang Eun Kim, and et al. 2025. "Quantitative Real-Time Polymerase Chain Reaction (PCR) Assay for Rapid Monitoring of the Harmful Algal Bloom Species Cochlodinium polykrikoides" Journal of Marine Science and Engineering 13, no. 2: 277. https://doi.org/10.3390/jmse13020277
APA StyleKim, M.-J., Kim, H.-J., Park, J. S., Kang, D., Cho, S., Kim, H., Baek, S. H., Park, J. J. C., Han, J., Kim, K. E., & Jung, S. W. (2025). Quantitative Real-Time Polymerase Chain Reaction (PCR) Assay for Rapid Monitoring of the Harmful Algal Bloom Species Cochlodinium polykrikoides. Journal of Marine Science and Engineering, 13(2), 277. https://doi.org/10.3390/jmse13020277