
Numerical Study on the Swimming and Energy Self-Sufficiency of Multi-Joint Robotic Fish


Abstract
1. Introduction
2. Methodology and Validation
2.1. SPH Fundamental
2.2. Governing Equations
2.3. Project Chrono and Coupling with DualSPHysics
2.4. Model and Evaluation of Multi-Articular Robotic Fish
2.4.1. The Model of Eel-like Locomotion
2.4.2. Energy Costs and Efficiency of Swimming
2.4.3. PTO System for the Energy-Harvesting Model
2.4.4. Energy Harvesting and Efficiency
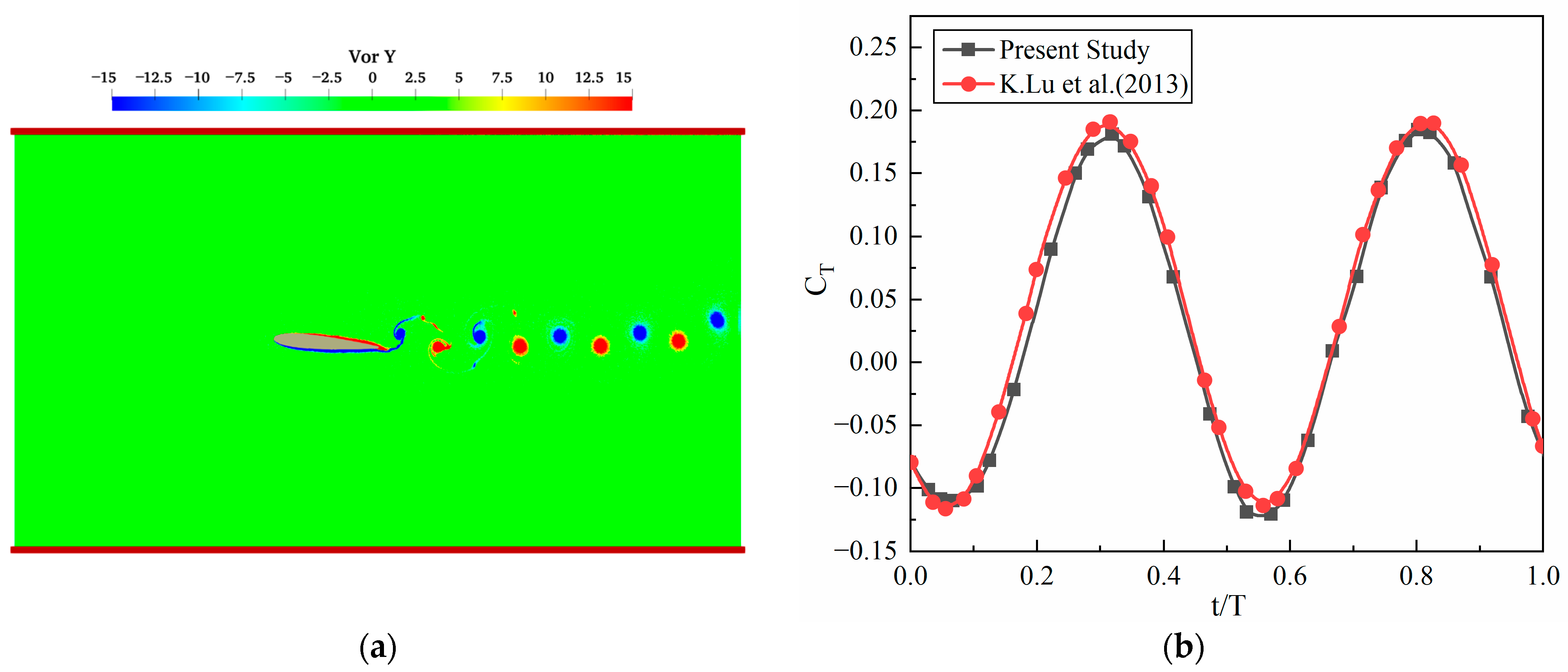
2.5. Numerical Model Validation
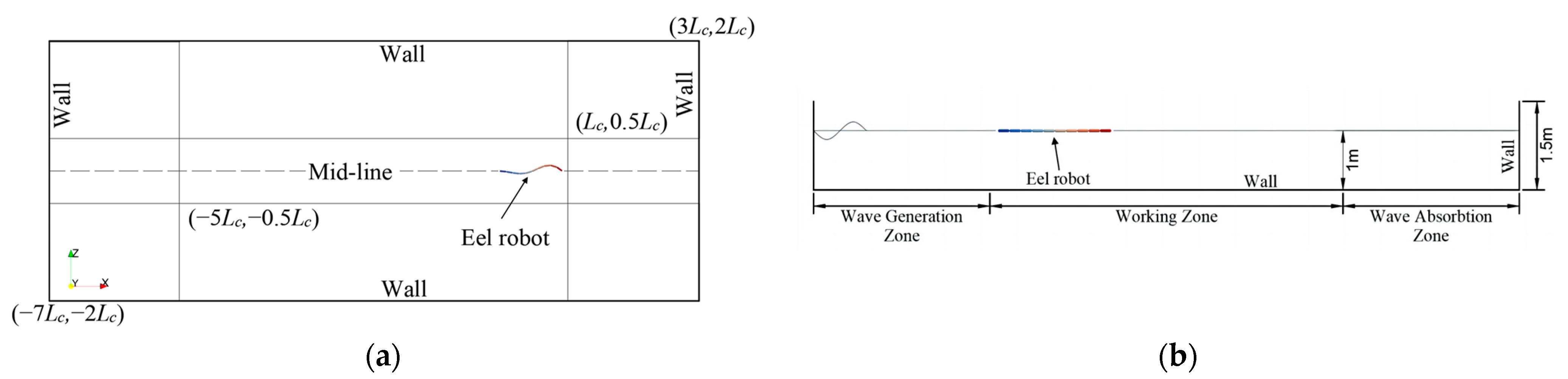
3. Simulation Domain and Resolution Verification
3.1. Simulation Setting
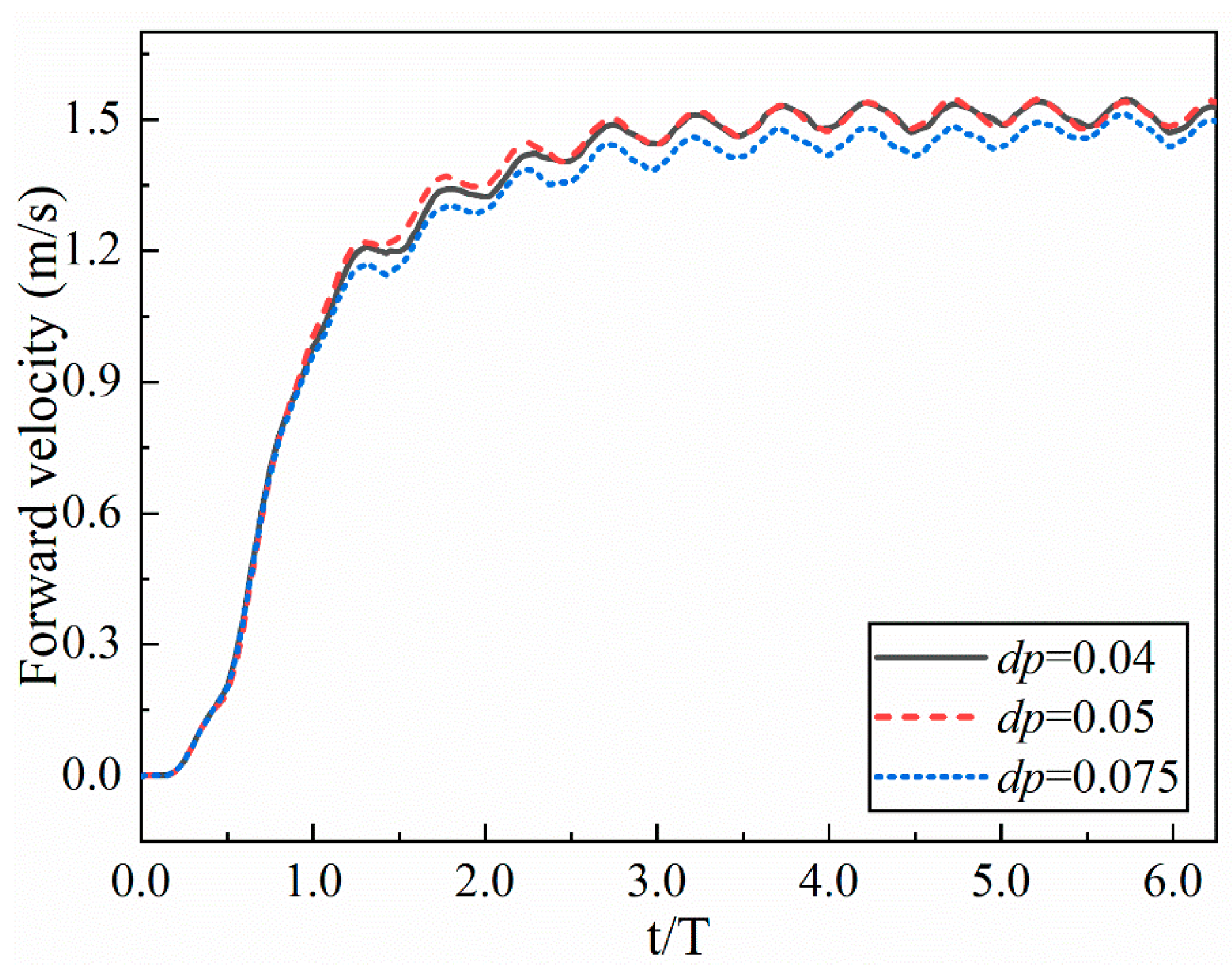
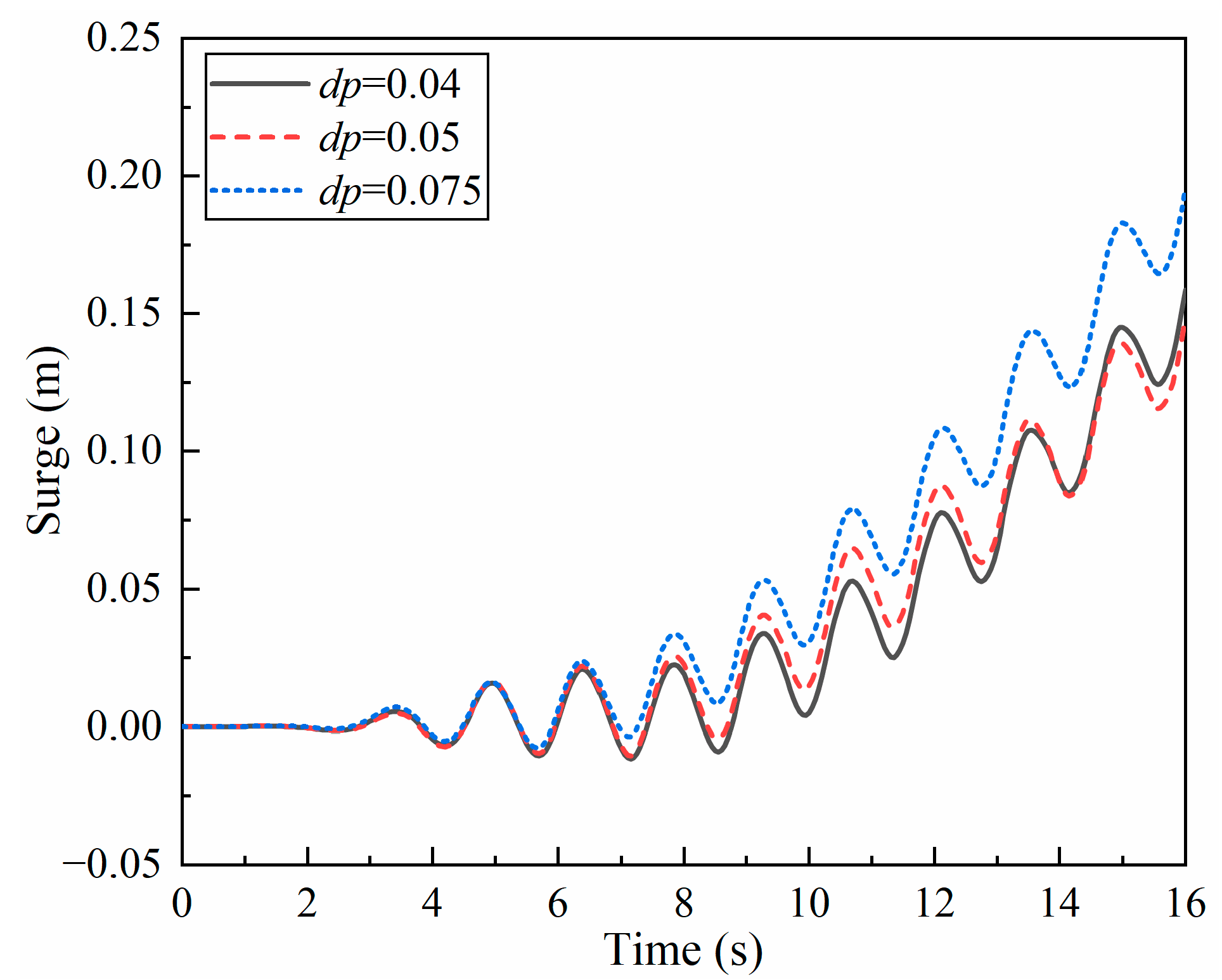
3.2. Resolution Verification
4. Results and Discussion
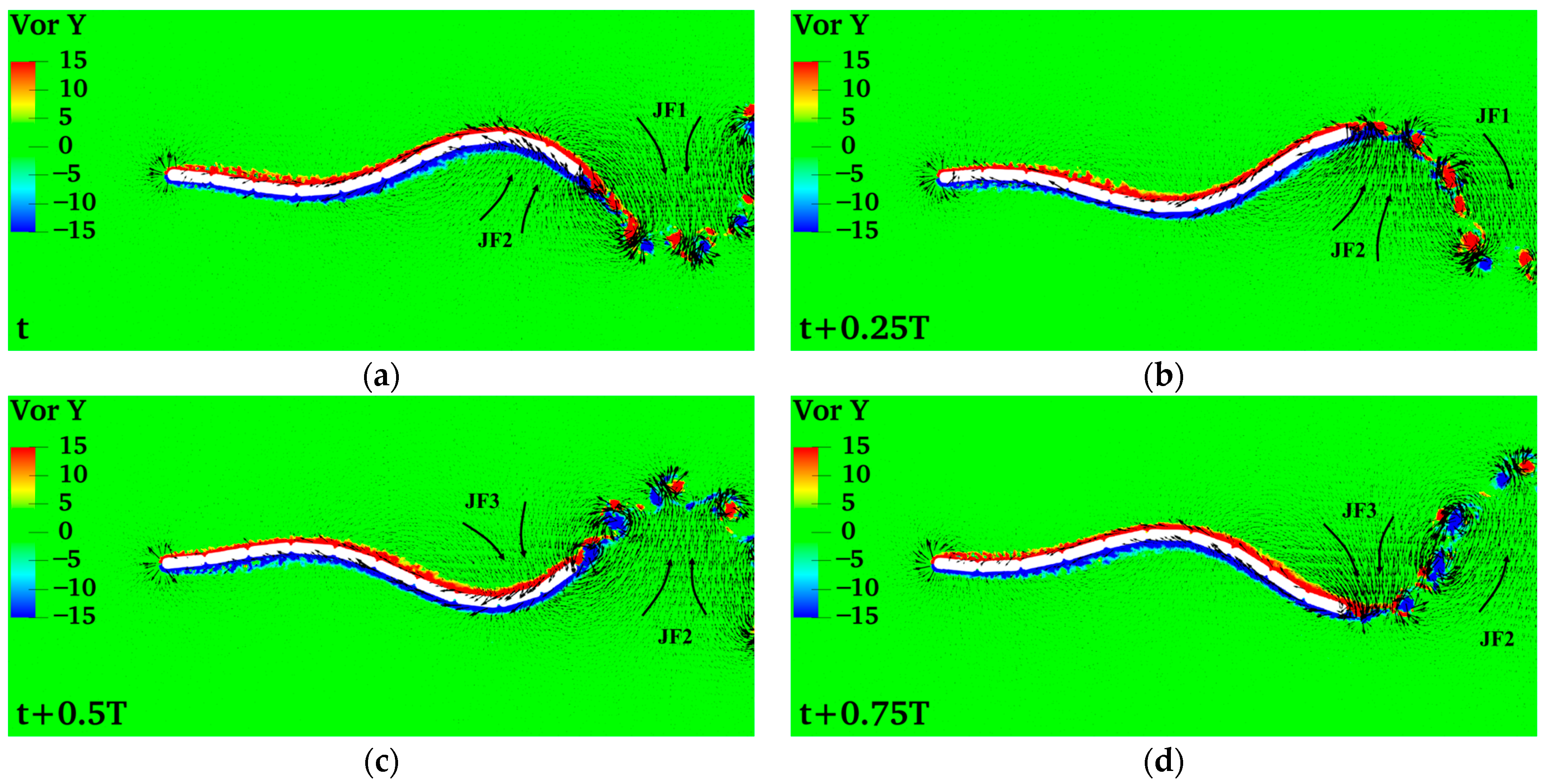
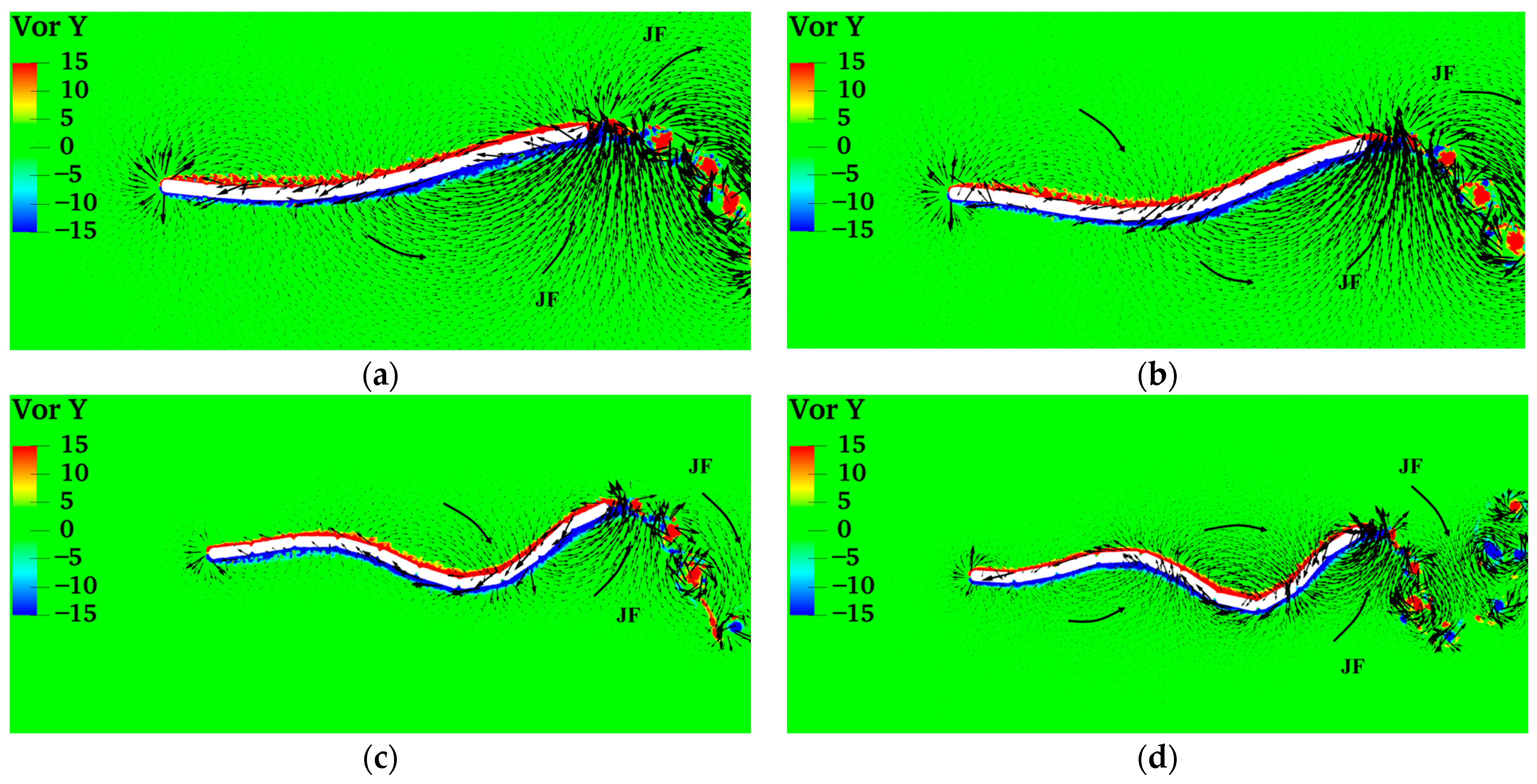
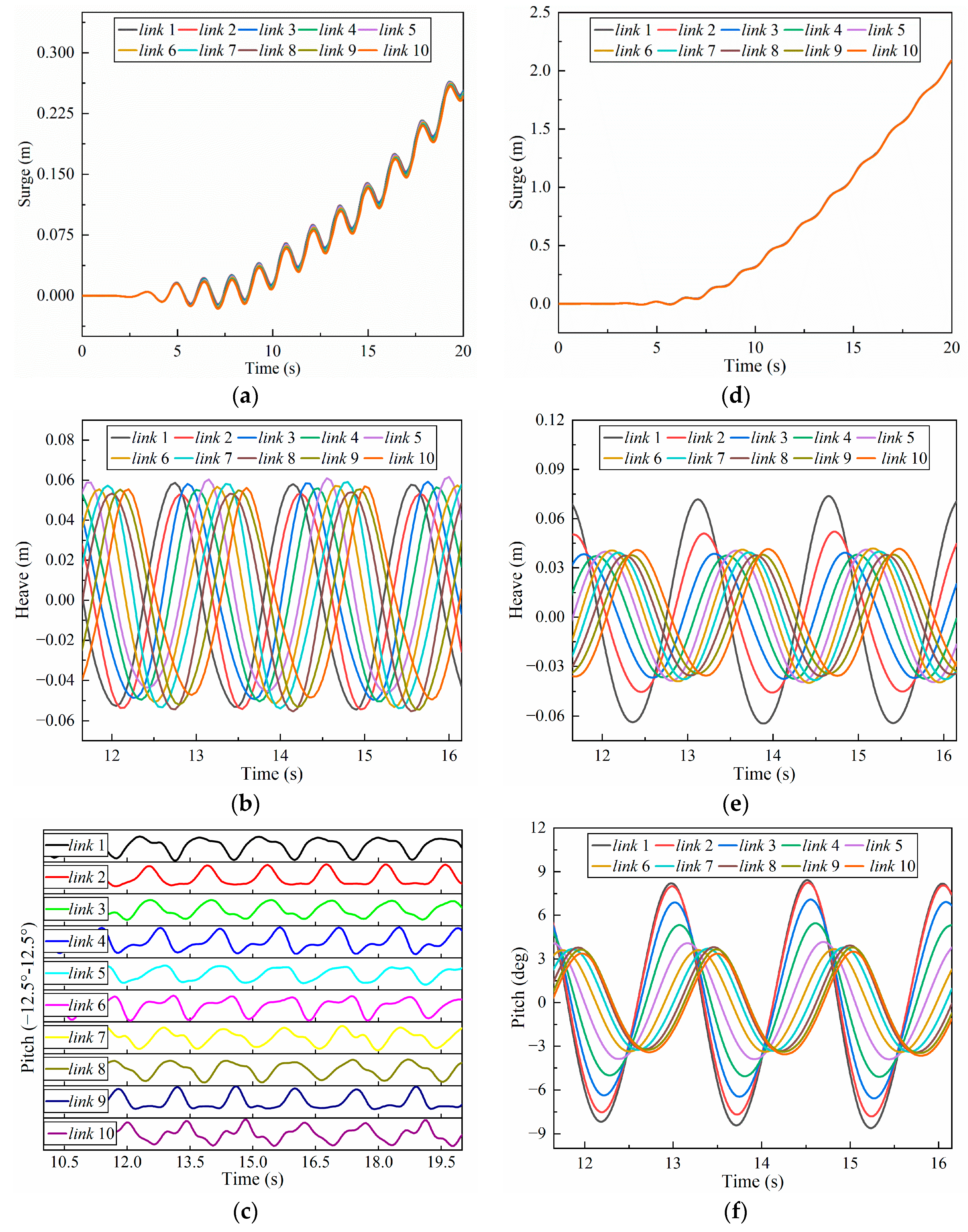
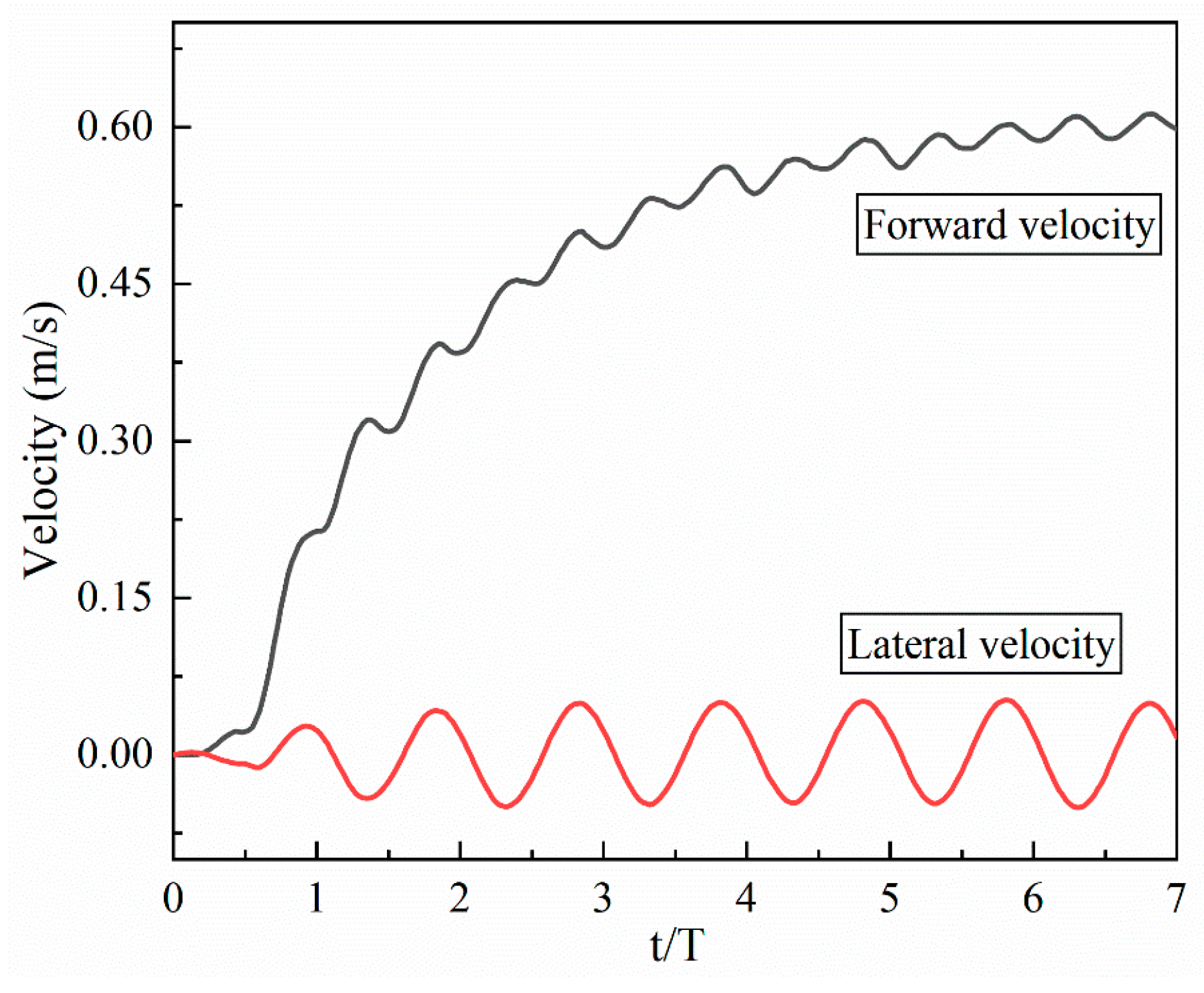
4.1. Swimming Model and Swimming Efficiency of Multi-Joint Robotic Fish
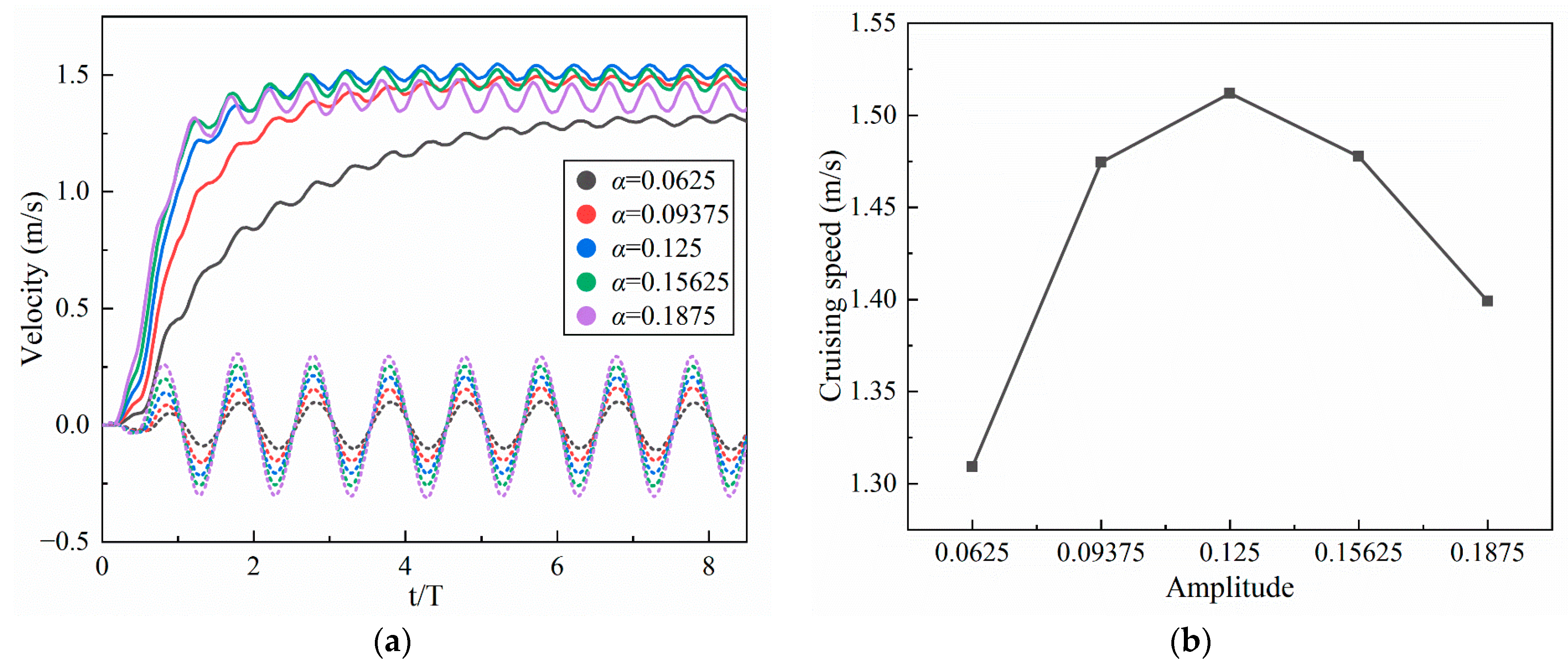
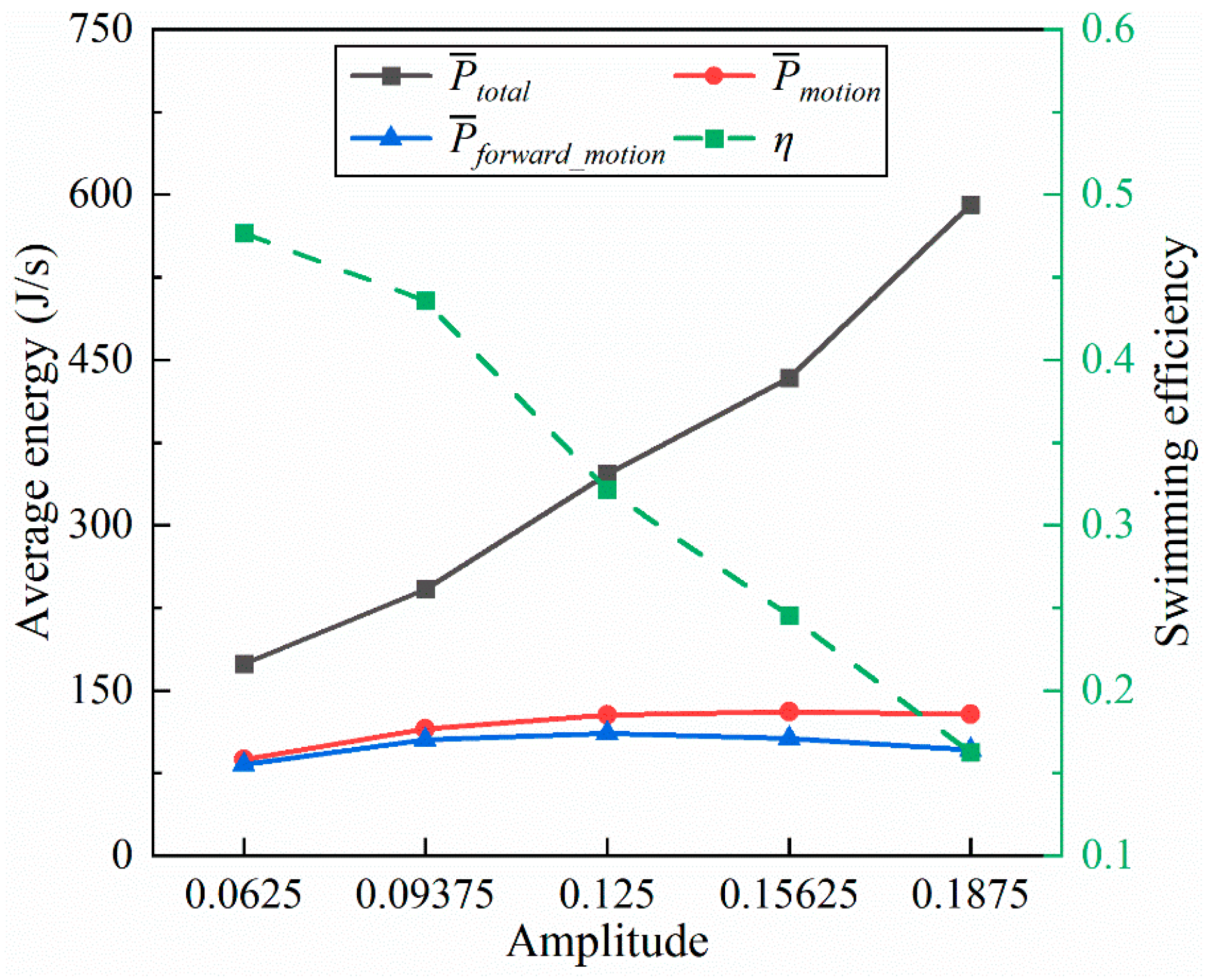
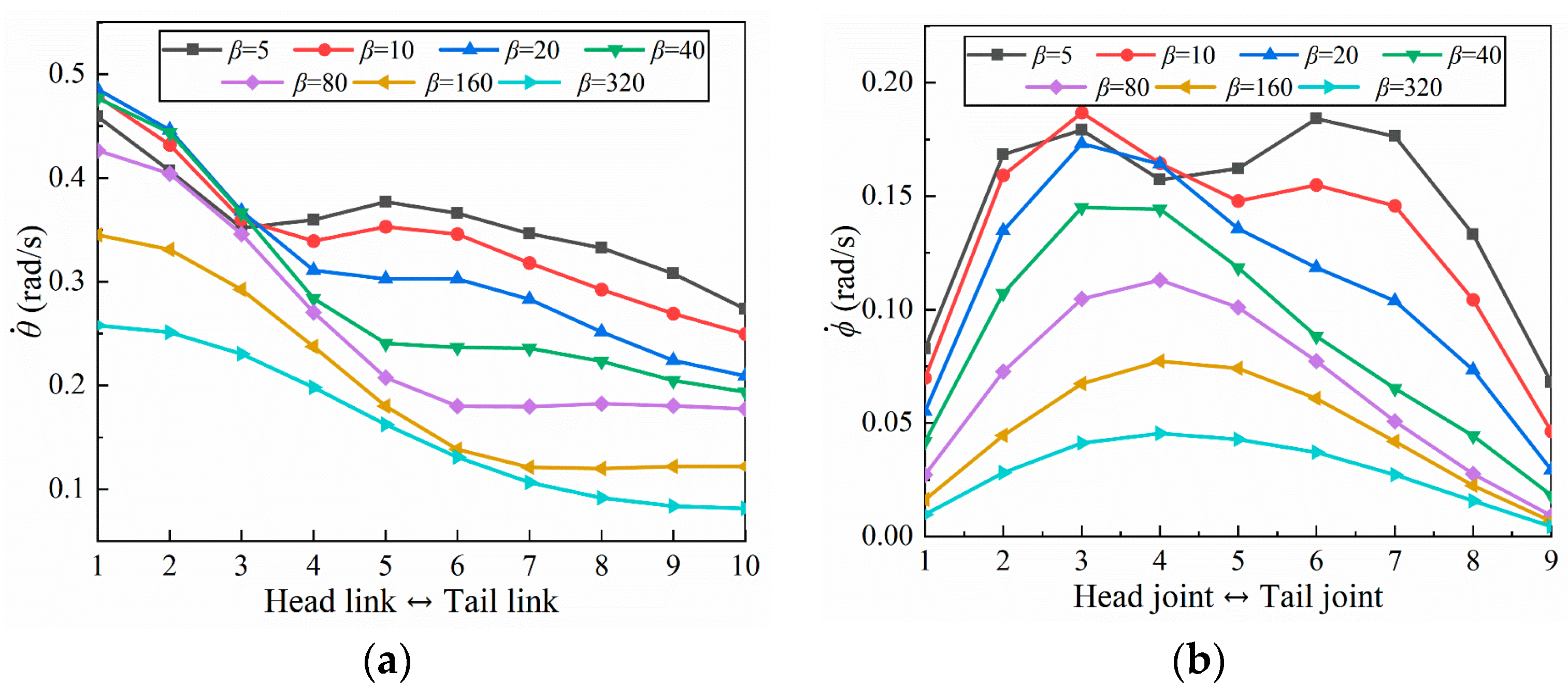
4.1.1. Amplitude Coefficient
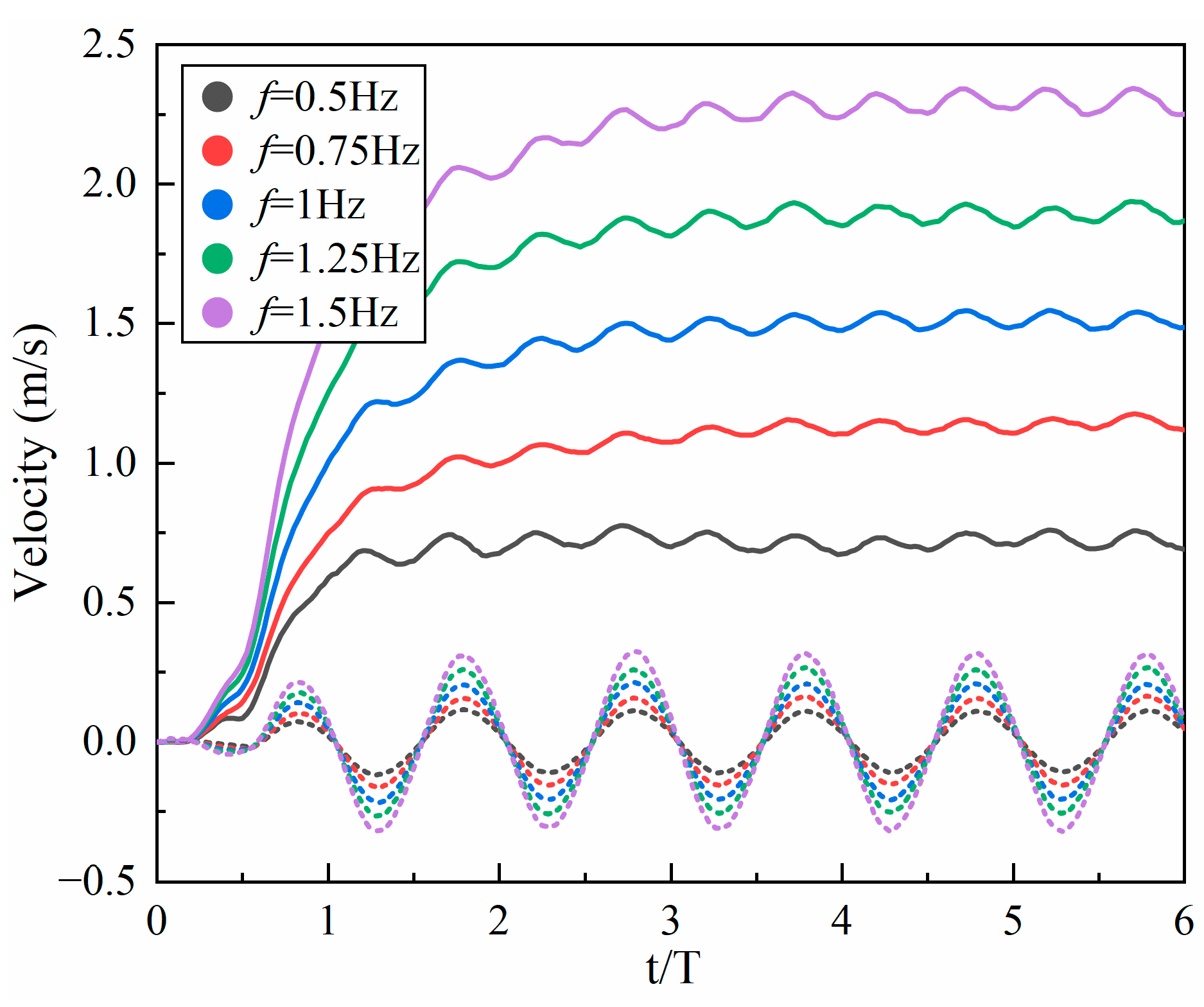
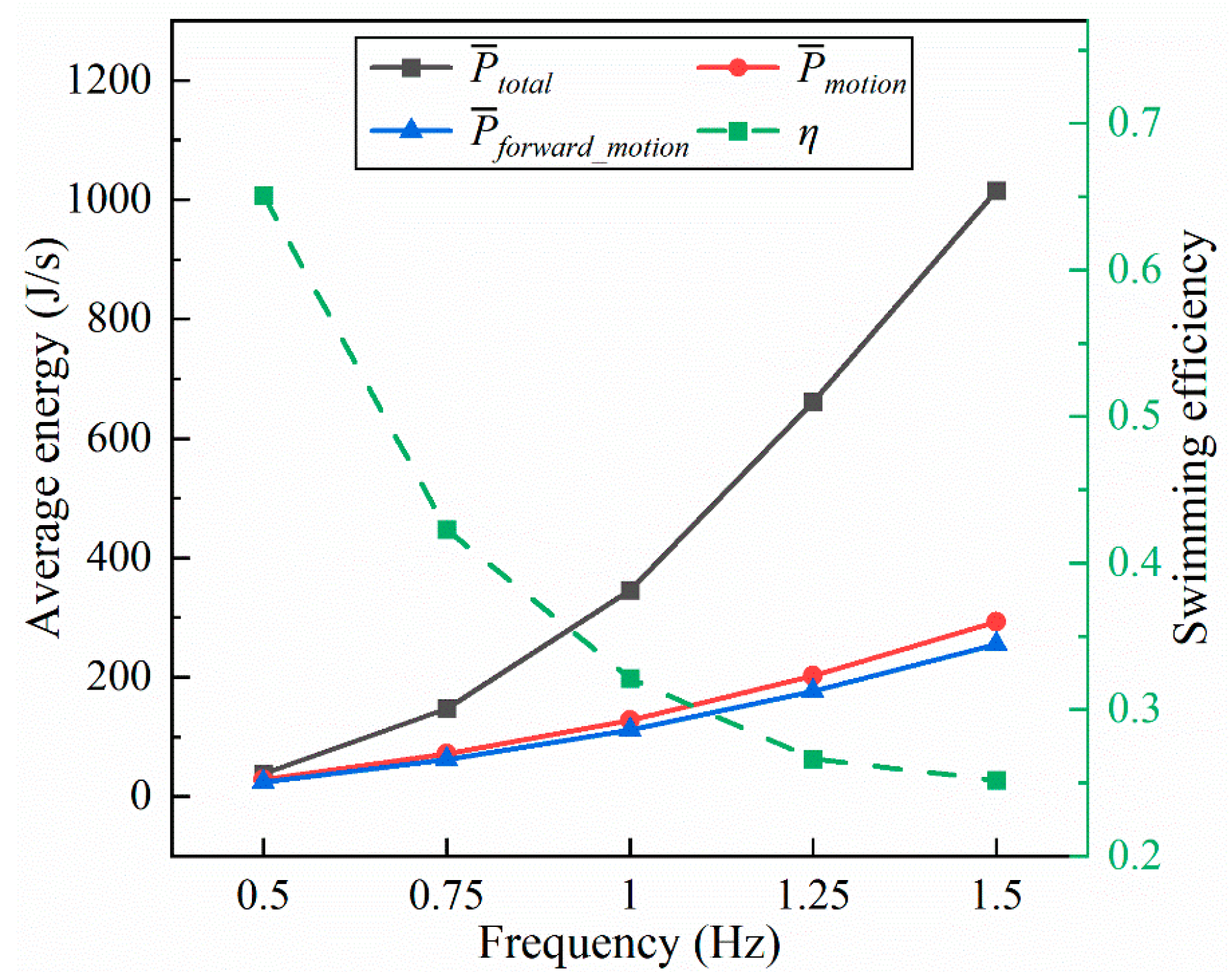
4.1.2. Frequency
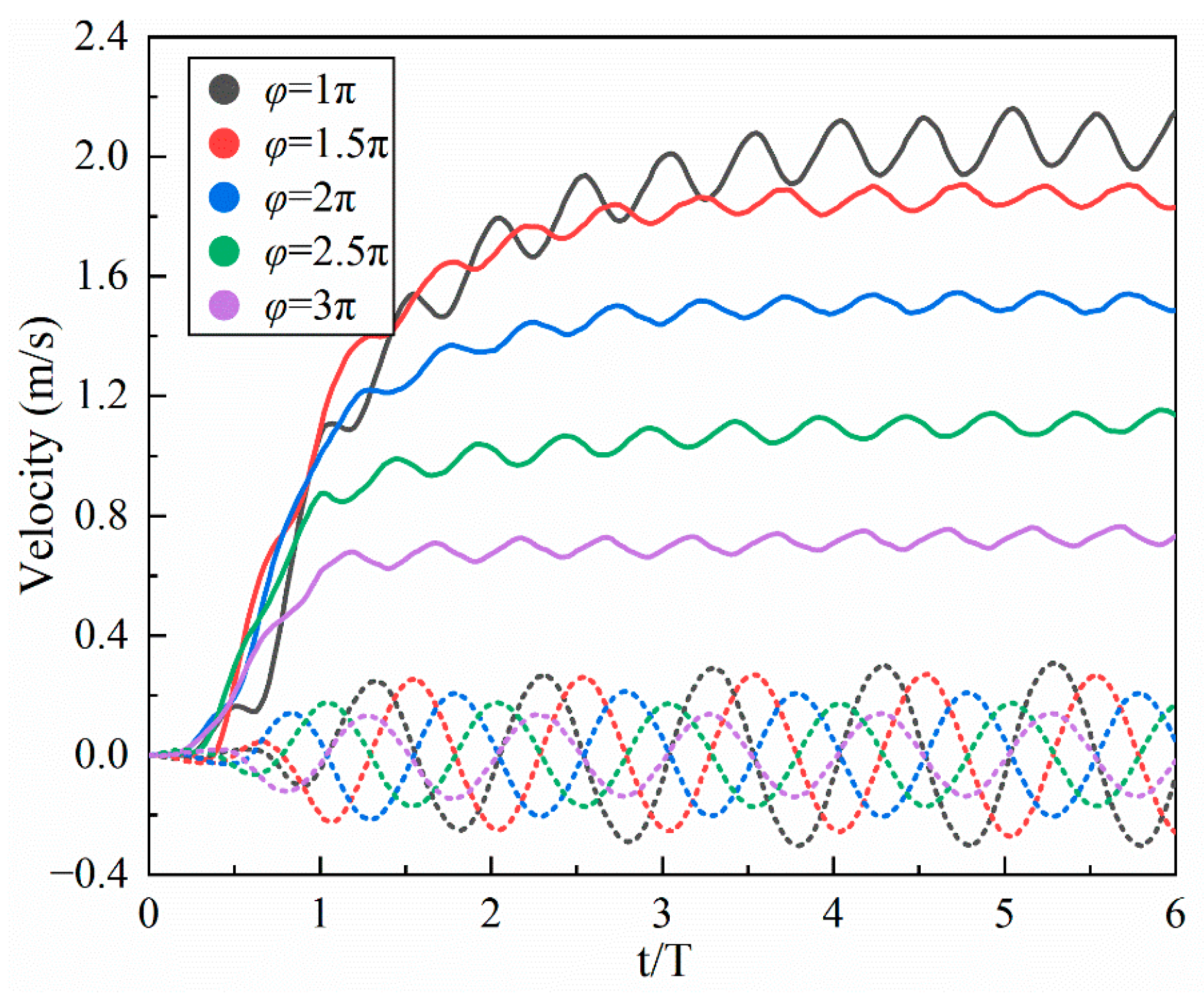
4.1.3. Phase
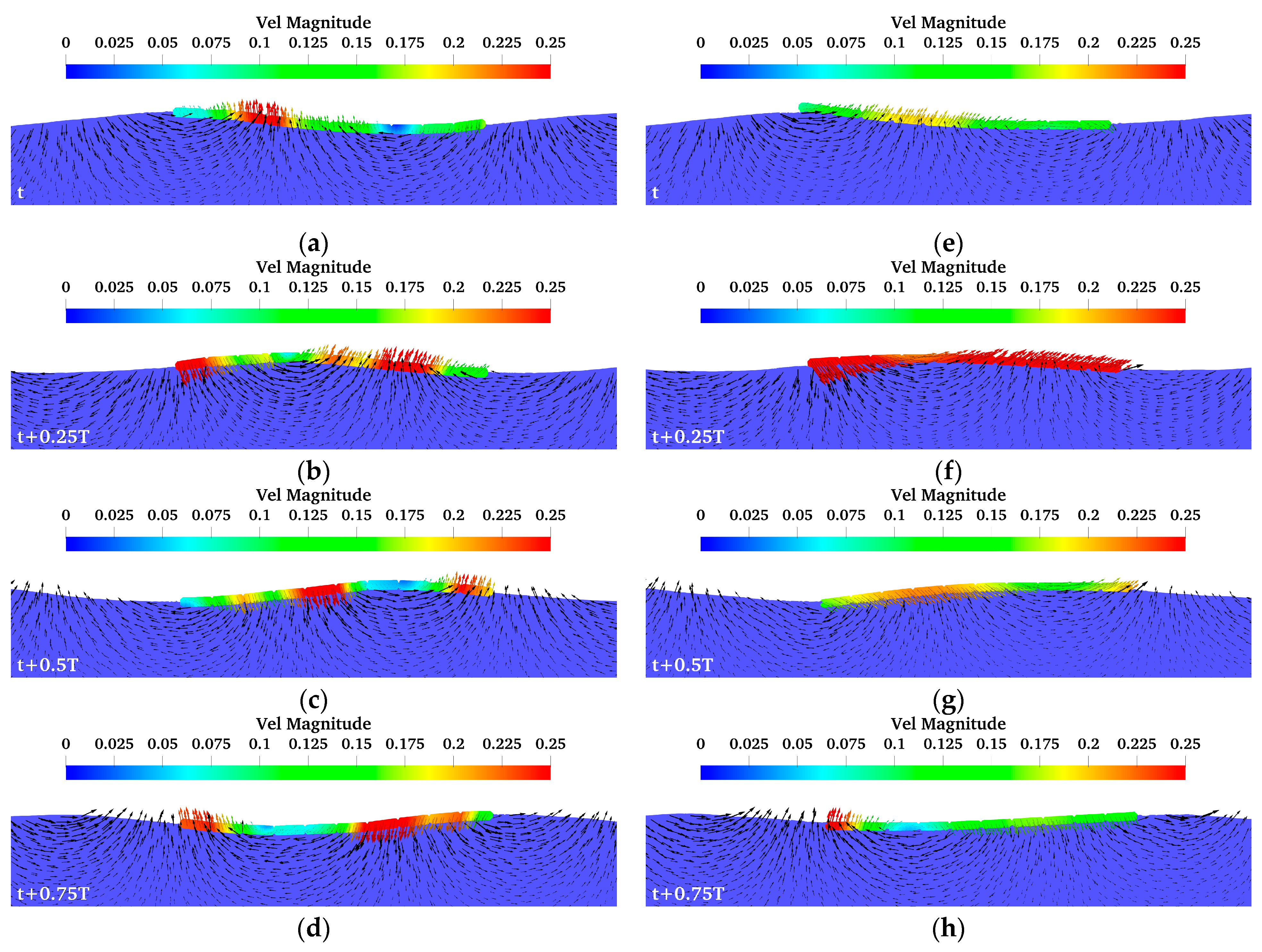
4.2. Energy Harvesting Model and Power in Waves
4.3. Feasibility Analysis of Energy Self-Sufficiency
5. Conclusions
- Inspired by the locomotion of eels, the thrust of multi-joint robotic fish primarily arises from the reactive force of the jet flow generated by tail oscillations. The undulation parameters, namely, the amplitude, frequency, and phase, of the robotic fish are crucial factors influencing the strength of the jet flow, and they strongly affect the velocity, energy costs, and swimming efficiency of the fish. It is observed that the frequency is the primary variable affecting both the energy costs and efficiency, followed by the amplitude and phase. Although low frequencies and amplitudes, along with phase adjustments to maintain a complete wavelength, could effectively reduce energy costs and promote body stability, there is a compromise in the forward speed.
- PTO systems with different damping coefficients affect the wave-induced motion of robotic fish by applying a resistance moment at the joints, thereby impacting the power and efficiency of energy harvesting. It is observed that determining the appropriate damping coefficient for multi-joint robotic fish is important for balancing body motion and joint resistance moment during energy harvesting, thereby maximizing the harvested energy from waves. In the wave environment in this study, the maximum power output can reach 4 W under optimal damping coefficients.
- The energy harvesting of multi-joint robotic fish is determined by the damping coefficients and environmental factors. Therefore, the key to achieving energy self-sufficiency for multi-joint robotic fish lies in adjusting the undulation parameters to reduce energy costs and enhance swimming efficiency. Under the examined conditions of wave energy harvesting and undulation parameters, the ratio of the average energy harvested by the multi-joint robotic fish to the average energy cost is 1:5.8, demonstrating that energy self-sufficiency is feasible.
- This research demonstrates that it is feasible for multi-joint robotic fish to achieve energy self-sufficiency in wave environments, and there is great potential for development. Additionally, it provides an effective and sustainable solution for the long-term operation of underwater robots.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Breder, C.M. The Locomotion of Fishes. Zoologica 1926, 4, 159–297. [Google Scholar] [CrossRef]
- Van Ginneken, V.; Antonissen, E.; Müller, U.K.; Booms, R.; Eding, E.; Verreth, J.; Van Den Thillart, G. Eel Migration to the Sargasso: Remarkably High Swimming Efficiency and Low Energy Costs. J. Exp. Biol. 2005, 208, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Gillis, G.B. Undulatory Locomotion in Elongate Aquatic Vertebrates: Anguilliform Swimming since Sir James Gray. Am. Zool. 1996, 36, 656–665. [Google Scholar] [CrossRef]
- Gray, J. Studies in Animal Locomotion: I. The Movement of Fish with Special Reference to the Eel. J. Exp. Biol. 1933, 10, 88–104. [Google Scholar] [CrossRef]
- Fish, F.E.; Lauder, G.V. Passive and Active Flow Control by Swimming Fishes and Mammals. Annu. Rev. Fluid Mech. 2006, 38, 193–224. [Google Scholar] [CrossRef]
- Müller, U.K.; Van Leeuwen, J.L. Undulatory Fish Swimming: From Muscles to Flow. Fish Fish. 2006, 7, 84–103. [Google Scholar] [CrossRef]
- Triantafyllou, M.S.; Triantafyllou, G.S.; Yue, D.K.P. Hydrodynamics of Fishlike Swimming. Annu. Rev. Fluid Mech. 2000, 32, 33–53. [Google Scholar] [CrossRef]
- Minami, M.; Jingyu, G.; Mae, Y. Chaos-Driving Robotic Intelligence for Catching Fish. In Proceedings of the 2007 IEEE International Conference on Robotics and Automation, Rome, Italy, 10–14 April 2007; pp. 85–91. [Google Scholar]
- Carling, J.; Williams, T.L.; Bowtell, G. Self-Propelled Anguilliform Swimming: Simultaneous Solution of the Two-Dimensional Navier–Stokes Equations and Newton’s Laws of Motion. J. Exp. Biol. 1998, 201, 3143–3166. [Google Scholar] [CrossRef] [PubMed]
- Taylo, G.I. Analysis of the Swimming of Long and Narrow Animals. Proc. R. Soc. Lond. Ser. A Math. Phys. Sci. 1952, 214, 158–183. [Google Scholar] [CrossRef]
- Lighthill, M.J. Large-Amplitude Elongated-Body Theory of Fish Locomotion. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1971, 179, 125–138. [Google Scholar] [CrossRef]
- McIsaac, K.A.; Ostrowski, J.P. Motion Planning for Anguilliform Locomotion. IEEE Trans. Robot. Autom. 2003, 19, 637–652. [Google Scholar] [CrossRef]
- Boyer, F.; Porez, M.; Khalil, W. Macro-Continuous Computed Torque Algorithm for a Three-Dimensional Eel-like Robot. IEEE Trans. Robot. 2006, 22, 763–775. [Google Scholar] [CrossRef]
- Ekeberg, Ö. A Combined Neuronal and Mechanical Model of Fish Swimming. Biol. Cybern. 1993, 69, 363–374. [Google Scholar] [CrossRef]
- Wiens, A.J.; Nahon, M. Optimally Efficient Swimming in Hyper-Redundant Mechanisms: Control, Design, and Energy Recovery. Bioinspiration Biomim. 2012, 7, 046016. [Google Scholar] [CrossRef] [PubMed]
- Khalil, W.; Gallot, G.; Boyer, F. Dynamic Modeling and Simulation of a 3-D Serial Eel-Like Robot. IEEE Trans. Syst. Man Cybern. C 2007, 37, 1259–1268. [Google Scholar] [CrossRef]
- Candelier, F.; Porez, M.; Boyer, F. Note on the Swimming of an Elongated Body in a Non-Uniform Flow. J. Fluid Mech. 2013, 716, 616–637. [Google Scholar] [CrossRef]
- Kelasidi, E.; Pettersen, K.Y.; Gravdahl, J.T. Modeling of Underwater Snake Robots. In Proceedings of the 2014 IEEE International Conference on Robotics and Automation (ICRA), Hong Kong, China, 31 May–7 June 2014; pp. 4540–4547. [Google Scholar] [CrossRef]
- Mikołajczyk, T.; Mikołajewski, D.; Kłodowski, A.; Łukaszewicz, A.; Mikołajewska, E.; Paczkowski, T.; Macko, M.; Skornia, M. Energy Sources of Mobile Robot Power Systems: A Systematic Review and Comparison of Efficiency. Appl. Sci. 2023, 13, 7547. [Google Scholar] [CrossRef]
- Prudell, J.; Stoddard, M.; Amon, E.; Brekken, T.K.A.; Von Jouanne, A. A Permanent-Magnet Tubular Linear Generator for Ocean Wave Energy Conversion. IEEE Trans. Ind. Appl. 2010, 46, 2392–2400. [Google Scholar] [CrossRef]
- Sodano, H.A.; Park, G.; Inman, D.J. Estimation of Electric Charge Output for Piezoelectric Energy Harvesting. Strain 2004, 40, 49–58. [Google Scholar] [CrossRef]
- Wang, J.; Tang, L.; Zhao, L.; Zhang, Z. Efficiency Investigation on Energy Harvesting from Airflows in HVAC System Based on Galloping of Isosceles Triangle Sectioned Bluff Bodies. Energy 2019, 172, 1066–1078. [Google Scholar] [CrossRef]
- Michelin, S.; Doare, O. Energy Harvesting Efficiency of Piezoelectric Flags in Axial Flows. J. Fluid Mech. 2013, 714, 489–504. [Google Scholar] [CrossRef]
- Hirt, C.W.; Nichols, B.D. Volume of Fluid (VOF) Method for the Dynamics of Free Boundaries. J. Comput. Phys. 1981, 39, 201–225. [Google Scholar] [CrossRef]
- Lakzian, E.; Ramezani, M.; Shabani, S.; Salmani, F.; Majkut, M.; Kim, H.D. The Search for an Appropriate Condensation Model to Simulate Wet Steam Transonic Flows. Int. J. Numer. Methods Heat Fluid Flow 2023, 33, 2853–2876. [Google Scholar] [CrossRef]
- Dalrymple, R.A.; Rogers, B.D. Numerical Modeling of Water Waves with the SPH Method. Coast. Eng. 2006, 53, 141–147. [Google Scholar] [CrossRef]
- Monaghan, J.J.; Kos, A.; Issa, N. Fluid Motion Generated by Impact. J. Waterw. Port Coast. Ocean. Eng. 2003, 129, 250–259. [Google Scholar] [CrossRef]
- Falcão, A.F. Wave Energy Utilization: A Review of the Technologies. Renew. Sustain. Energy Rev. 2010, 14, 899–918. [Google Scholar] [CrossRef]
- McCormick, M.E. Ocean Wave Energy Conversion; Courier Corporation: Chelmsford, MA, USA, 2013. [Google Scholar]
- Salter, S.H. World Progress in Wave Energy—1988. Int. J. Ambient. Energy 1989, 10, 3–24. [Google Scholar] [CrossRef]
- Dalton, G.J.; Alcorn, R.; Lewis, T. Case Study Feasibility Analysis of the Pelamis Wave Energy Convertor in Ireland, Portugal and North America. Renew. Energy 2010, 35, 443–455. [Google Scholar] [CrossRef]
- Dong, E.; Yan, Q.; Zhang, S.; Yang, J. Self-Powered System of Articulated Bionic Robotic Fish Using Wave Energy Harvesting. Robot 2009, 31, 501–505. [Google Scholar]
- Zhu, W.; Wang, X.; Xu, M.; Yang, J.; Si, T.; Zhang, S. A Wave Energy Conversion Mechanism Applied in Robotic Fish. In Proceedings of the 2013 IEEE/ASME International Conference on Advanced Intelligent Mechatronics, Wollongong, NSW, Australia, 9–12 July 2013; pp. 319–324. [Google Scholar] [CrossRef]
- Ruffini, G.; Briganti, R.; De Girolamo, P.; Stolle, J.; Ghiassi, B.; Castellino, M. Numerical Modelling of Flow-Debris Interaction during Extreme Hydrodynamic Events with DualSPHysics-CHRONO. Appl. Sci. 2021, 11, 3618. [Google Scholar] [CrossRef]
- Domínguez, J.M.; Fourtakas, G.; Altomare, C.; Canelas, R.B.; Tafuni, A.; García-Feal, O.; Martínez-Estévez, I.; Mokos, A.; Vacondio, R.; Crespo, A.J.C.; et al. DualSPHysics: From Fluid Dynamics to Multiphysics Problems. Comput. Part. Mech. 2022, 9, 867–895. [Google Scholar] [CrossRef]
- Lo, E.Y.; Shao, S. Simulation of Near-Shore Solitary Wave Mechanics by an Incompressible SPH Method. Appl. Ocean Res. 2002, 24, 275–286. [Google Scholar] [CrossRef]
- Fourtakas, G.; Dominguez, J.M.; Vacondio, R.; Rogers, B.D. Local Uniform Stencil (LUST) Boundary Condition for Arbitrary 3-D Boundaries in Parallel Smoothed Particle Hydrodynamics (SPH) Models. Comput. Fluids 2019, 190, 346–361. [Google Scholar] [CrossRef]
- Tasora, A.; Anitescu, M. A Matrix-Free Cone Complementarity Approach for Solving Large-Scale, Nonsmooth, Rigid Body Dynamics. Comput. Methods Appl. Mech. Eng. 2011, 200, 439–453. [Google Scholar] [CrossRef]
- Canelas, R.B.; Domínguez, J.M.; Crespo, A.J.C.; Gómez-Gesteira, M.; Ferreira, R.M.L. A Smooth Particle Hydrodynamics Discretization for the Modelling of Free Surface Flows and Rigid Body Dynamics. Numer. Methods Fluids 2015, 78, 581–593. [Google Scholar] [CrossRef]
- Brito, M.; Canelas, R.B.; García-Feal, O.; Domínguez, J.M.; Crespo, A.J.C.; Ferreira, R.M.L.; Neves, M.G.; Teixeira, L. A Numerical Tool for Modelling Oscillating Wave Surge Converter with Nonlinear Mechanical Constraints. Renew. Energy 2020, 146, 2024–2043. [Google Scholar] [CrossRef]
- Tasora, A.; Serban, R.; Mazhar, H.; Pazouki, A.; Melanz, D.; Fleischmann, J.; Taylor, M.; Sugiyama, H.; Negrut, D. Chrono: An Open Source Multi-Physics Dynamics Engine. In High Performance Computing in Science and Engineering; Lecture Notes in Computer Science; Springer International Publishing: Cham, Switzerland, 2016; Volume 9611, pp. 19–49. ISBN 978-3-319-40360-1. [Google Scholar]
- Canelas, R.B.; Brito, M.; Feal, O.G.; Domínguez, J.M.; Crespo, A.J.C. Extending DualSPHysics with a Differential Variational Inequality: Modeling Fluid-Mechanism Interaction. Appl. Ocean Res. 2018, 76, 88–97. [Google Scholar] [CrossRef]
- Kern, S.; Koumoutsakos, P. Simulations of Optimized Anguilliform Swimming. J. Exp. Biol. 2006, 209, 4841–4857. [Google Scholar] [CrossRef] [PubMed]
- Bua, L.; Magis, B.; Ronsse, R.; Chatelain, P.; Bernier, C.; Jeanmart, H. Swimming Eel-like Robot: Design Improvement and Control. Master’s Thesis, Ecole polytechnique de Louvain, Louvain-la-Neuve, Belgium, 2018. [Google Scholar]
- Kelasidi, E.; Pettersen, K.Y.; Gravdahl, J.T. Energy Efficiency of Underwater Snake Robot Locomotion. In Proceedings of the 2015 23rd Mediterranean Conference on Control and Automation (MED), Torremolinos, Spain, 16–19 June 2015; pp. 1124–1131. [Google Scholar] [CrossRef]
- Tagliafierro, B.; Martínez-Estévez, I.; Crego-Loureiro, C.; Domínguez, J.M.; Crespo, A.J.C.; Coe, R.G.; Bacelli, G.; Gómez-Gesteira, M.; Viccione, G. Numerical Modeling of Moored Floating Platforms for Wave Energy Converters Using DualSPHysics. Ocean. Eng. 2022, 5, V05AT06A016. [Google Scholar] [CrossRef]
- Brooke, J. Wave Energy Conversion; Elsevier: Amsterdam, The Netherlands, 2003. [Google Scholar]
- Lu, K.; Xie, Y.H.; Zhang, D. Numerical Study of Large Amplitude, Nonsinusoidal Motion and Camber Effects on Pitching Airfoil Propulsion. J. Fluids Struct. 2013, 36, 184–194. [Google Scholar] [CrossRef]
- Ren, B.; He, M.; Dong, P.; Wen, H. Nonlinear Simulations of Wave-Induced Motions of a Freely Floating Body Using WCSPH Method. Appl. Ocean Res. 2015, 50, 1–12. [Google Scholar] [CrossRef]
- Feng, H.; Wang, Z.; Todd, P.A.; Lee, H.P. Simulations of Self-Propelled Anguilliform Swimming Using the Immersed Boundary Method in OpenFOAM. Eng. Appl. Comput. Fluid Mech. 2019, 13, 438–452. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (J/s) | (J/s) | (J/s) | |
|---|---|---|---|
| 23.3 | 18.7 | 17.53 | 0.75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, G.; Xin, Z.; Ding, Q.; Liu, S.; Ren, L. Numerical Study on the Swimming and Energy Self-Sufficiency of Multi-Joint Robotic Fish. J. Mar. Sci. Eng. 2024, 12, 701. https://doi.org/10.3390/jmse12050701
Liang G, Xin Z, Ding Q, Liu S, Ren L. Numerical Study on the Swimming and Energy Self-Sufficiency of Multi-Joint Robotic Fish. Journal of Marine Science and Engineering. 2024; 12(5):701. https://doi.org/10.3390/jmse12050701
Chicago/Turabian StyleLiang, Guodu, Zhiqiang Xin, Quanlin Ding, Songyang Liu, and Liying Ren. 2024. "Numerical Study on the Swimming and Energy Self-Sufficiency of Multi-Joint Robotic Fish" Journal of Marine Science and Engineering 12, no. 5: 701. https://doi.org/10.3390/jmse12050701
APA StyleLiang, G., Xin, Z., Ding, Q., Liu, S., & Ren, L. (2024). Numerical Study on the Swimming and Energy Self-Sufficiency of Multi-Joint Robotic Fish. Journal of Marine Science and Engineering, 12(5), 701. https://doi.org/10.3390/jmse12050701