
Transcriptome Analysis of Multiple Tissues in the Shrimp Penaeus vannamei Reveals the Typical Physiological Response to Three Pathogens



Abstract
1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Transcriptome Assembly and Differential Expression Gene Analysis
3. Results
3.1. Significant Differentially Expressed Genes (DEGs)
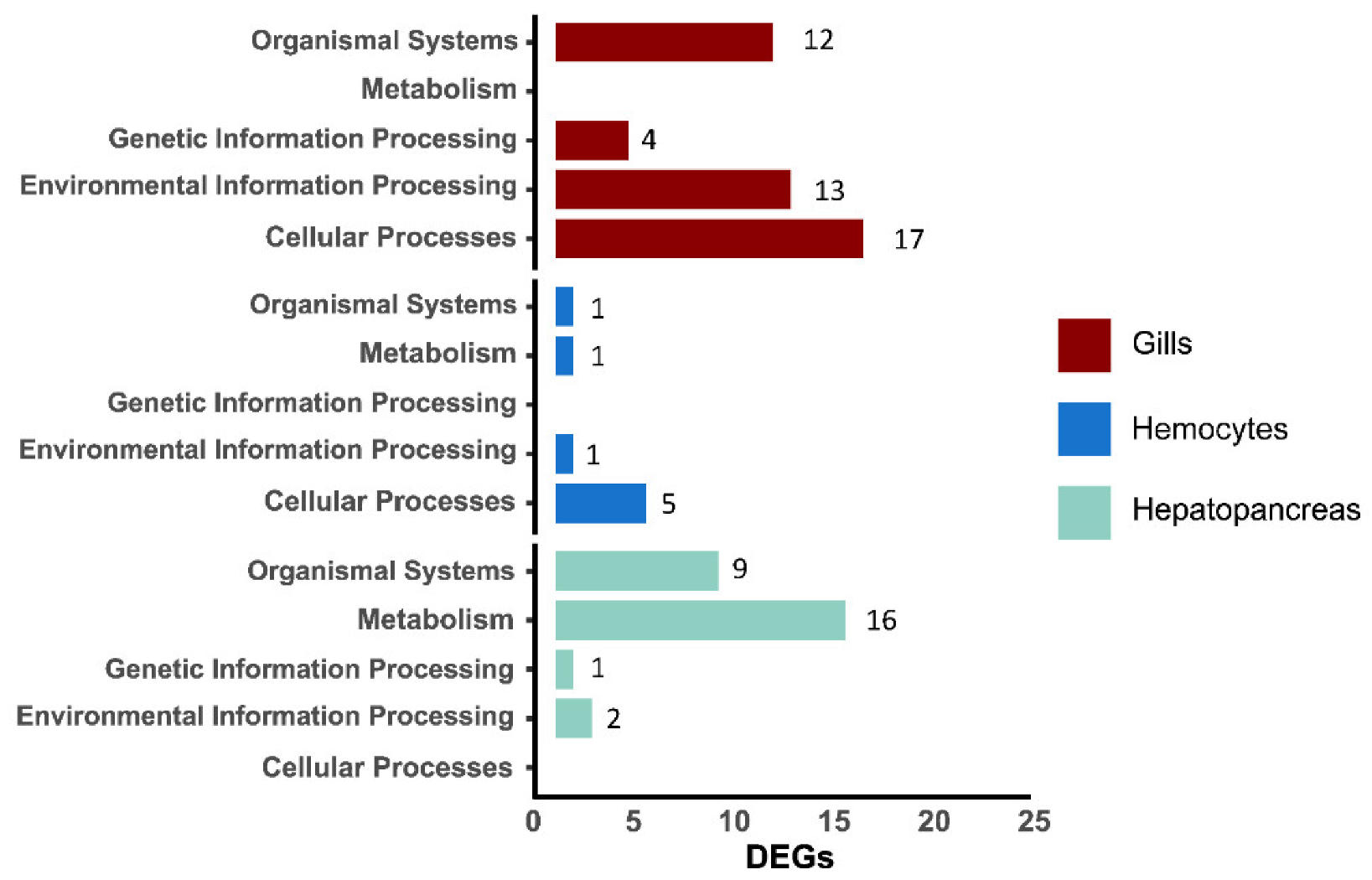
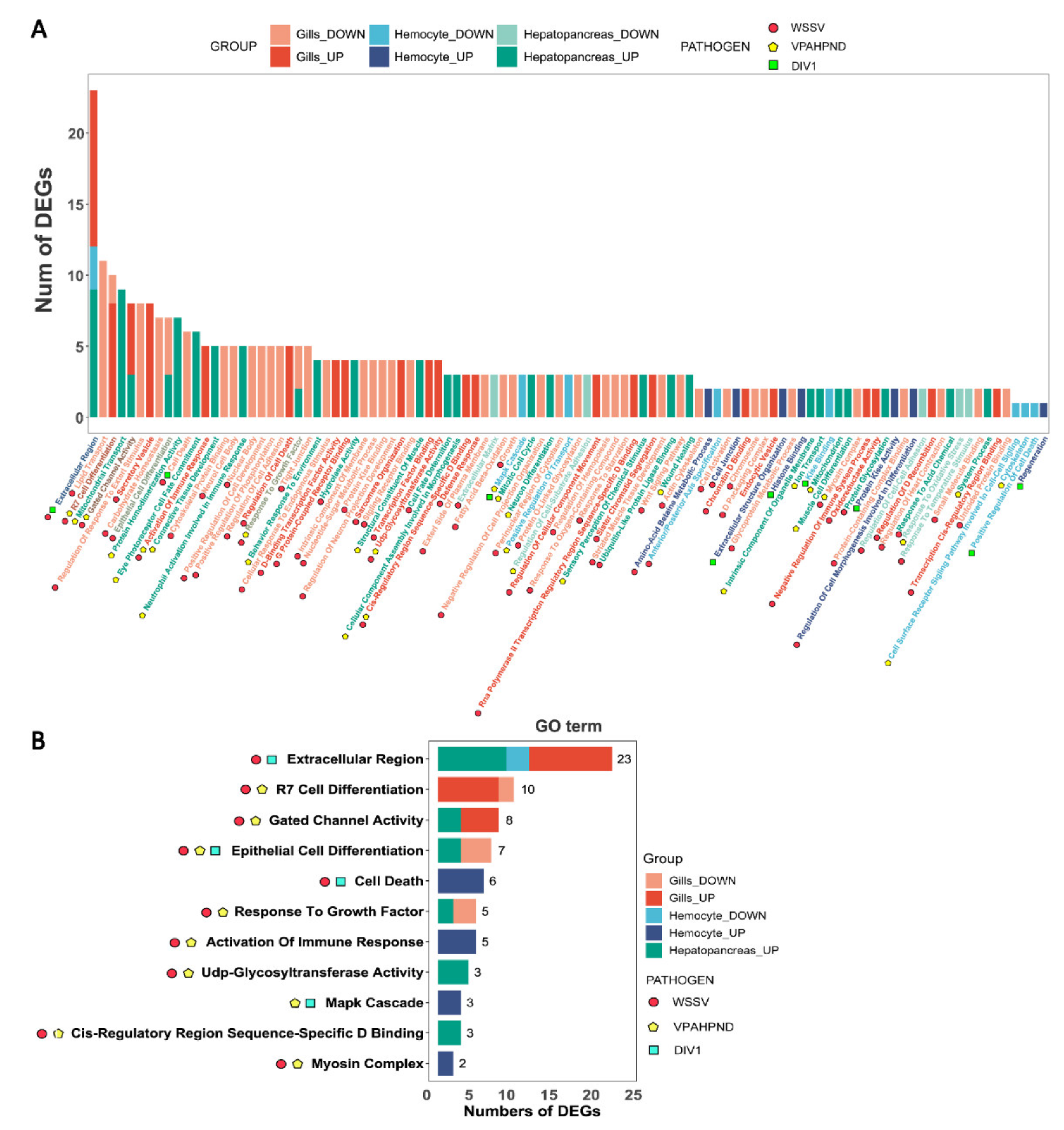
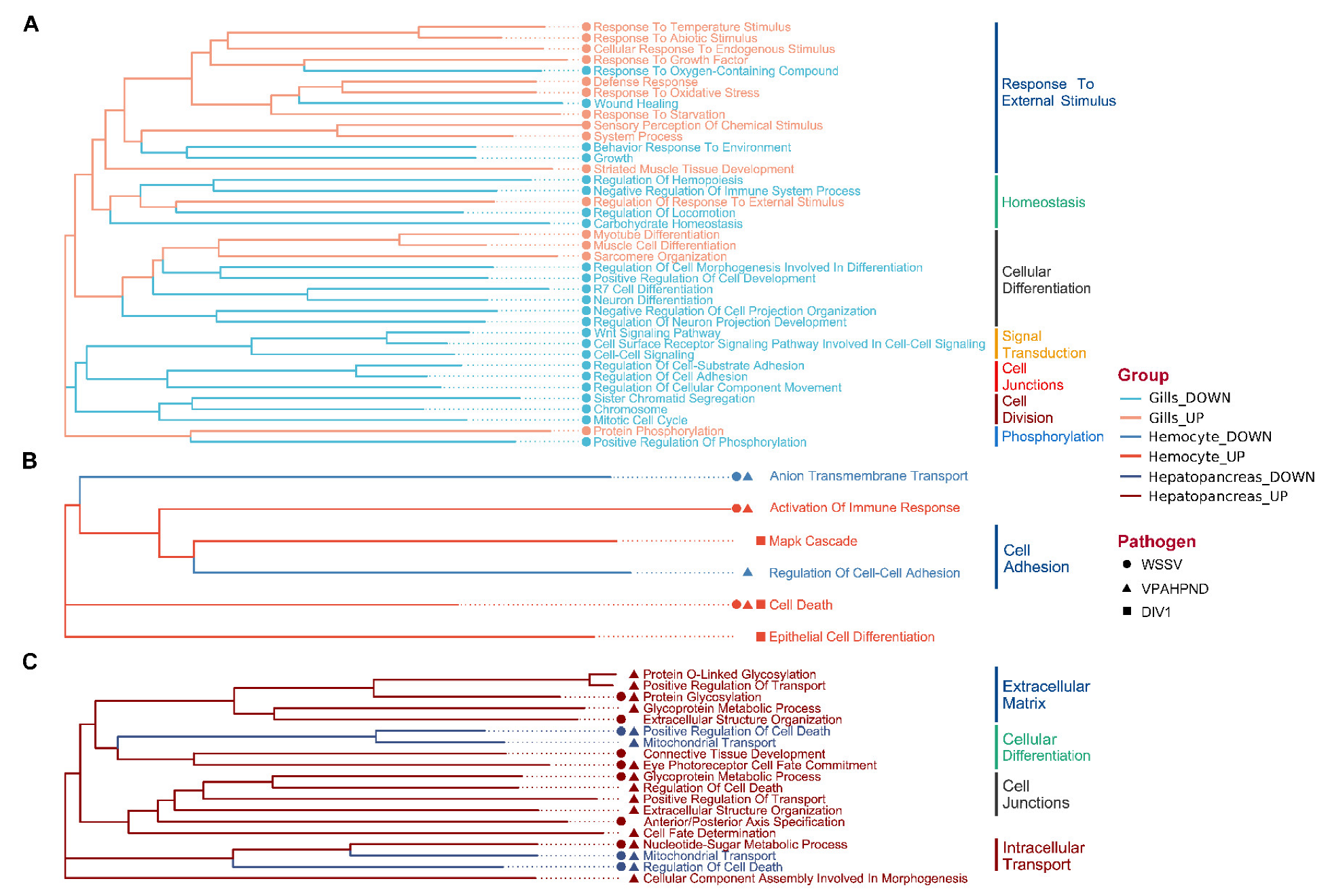
3.2. GO Analysis of DEGs
3.3. Pathway Enrichment Analysis of DEGs
4. Discussion
4.1. Tissue-Specific Responses
4.2. Immune Responses Shared among Tissues
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cai, J.; Galli, G. Top 10 Species Groups in Global Aquaculture 2019; Food and Agriculture Organization of the United Nations: Rome, Italy, 2021. [Google Scholar]
- Zhang, S.; Fang, S.; Song, S.; Zheng, Y.; Tan, B.; Shi, L. Effect of changes in the activity of Wnt/β-catenin signalling pathway on the growth performance, immunity and transcriptome response in Litopenaeus vannamei. Aquac. Rep. 2021, 20, 100774. [Google Scholar] [CrossRef]
- Flegel, T.W. Historic emergence, impact and current status of shrimp pathogens in Asia. J. Invertebr. Pathol. 2012, 110, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Millard, R.; Bickley, L.; Bateman, K.; Farbos, A.; Minardi, D.; Moore, K.; Ross, S.; Stentiford, G.; Tyler, C.; van Aerle, R.; et al. Global mRNA and miRNA Analysis reveal key processes in the initial response to infection with WSSV in the Pacific whiteleg shrimp. Viruses 2021, 13, 1140. [Google Scholar] [CrossRef]
- Yan, X.; Liao, X.; He, J.; Yin, B.; Li, C. Effects of WSSV and DIV1 infection on hemolymph coagulation in Litopenaeus vannamei. Shengwuxuezazhi 2022, 39, 18. [Google Scholar]
- Caro, L.F.A.; Mai, H.N.; Noble, B.; Dhar, A.K. Acute hepatopancreatic necrosis disease (VPAHPND), a chronic disease in shrimp (Penaeus vannamei) population raised in latin America. J. Invertebr. Pathol. 2020, 174, 107424. [Google Scholar] [CrossRef]
- Lightner, D.V. The penaeid shrimp viruses TSV, IHHNV, WSSV, and YHV: Current status in the Americas, available diagnostic methods, and management strategies. J. Appl. Aquac. 1999, 9, 27–52. [Google Scholar] [CrossRef]
- Chen, X.; Qiu, L.; Wang, H.; Zou, P.; Dong, X.; Li, F.; Huang, J. Susceptibility of Exopalaemon carinicauda to the infection with shrimp hemocyte iridescent virus (SHIV 20141215), a strain of Decapod Iridescent Virus 1 (DIV1). Viruses 2019, 11, 387. [Google Scholar] [CrossRef] [PubMed]
- Sangpo, P.; Thitamadee, S.; Dong, H.T.; Senapin, S. Aeromonas schubertii, a novel bacterium recovered from AHPND affected farm is lethal to whiteleg shrimp, Penaeus vannamei. Microb. Pathog. 2020, 149, 104501. [Google Scholar] [CrossRef]
- Qiu, L.; Chen, M.-M.; Wan, X.-Y.; Li, C.; Zhang, Q.-L.; Wang, R.-Y.; Cheng, D.-Y.; Dong, X.; Yang, B.; Wang, X.-H.; et al. Characterization of a new member of Iridoviridae, Shrimp hemocyte iridescent virus (SHIV), found in white leg shrimp (Litopenaeus vannamei). Sci. Rep. 2017, 7, 11834. [Google Scholar] [CrossRef] [PubMed]
- Pradeep, B.; Rai, P.; Mohan, S.A.; Shekhar, M.S.; Karunasagar, I. Biology, host range, pathogenesis and diagnosis of white spot syndrome virus. Indian J. Virol. 2012, 23, 161–174. [Google Scholar] [CrossRef]
- Yan, D.C.; Huang, J.; Yang, B.; Sun, H.S.; Wang, Y.Y.; Liu, X. Competition of infectious hypodermal and haematopoietic necrosis virus (IHHNV) with white spot syndrome virus (WSSV) for binding to shrimp cellular membrane. J. Fish Dis. 2016, 39, 1225–1229. [Google Scholar] [CrossRef]
- Jiravanichpaisal, P.; Söderhäll, K.; Söderhäll, I. Characterization of white spot syndrome virus replication in in vitro-cultured haematopoietic stem cells of freshwater crayfish, Pacifastacus leniusculus. J. Gen. Virol. 2006, 87, 847–854. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Li, S.; Li, F. Different immune responses of the lymphoid organ in shrimp at early challenge stage of Vibrio parahaemolyticus and WSSV. Animals 2021, 11, 2160. [Google Scholar] [CrossRef]
- Escobedo-Bonilla, C.M.; Alday-Sanz, V.; Wille, M.; Sorgeloos, P.; Pensaert, M.B.; Nauwynck, H.J. A review on the morphology, molecular characterization, morphogenesis and pathogenesis of white spot syndrome virus. J. Fish Dis. 2007, 31, 1–18. [Google Scholar] [CrossRef]
- Gong, H.-Y.; Li, Q.-Y.; Zhang, H.; Ye, L.; Shi, L.; Feng, Y.-H. Development and comparison of qPCR and qLAMP for rapid detection of the decapod iridescent virus 1 (DIV1). J. Invertebr. Pathol. 2021, 182, 107567. [Google Scholar] [CrossRef]
- Qiu, L.; Chen, X.; Gao, W.; Li, C.; Guo, X.-M.; Zhang, Q.-L.; Yang, B.; Huang, J. Molecular epidemiology and histopathological study of a natural infection with decapod iridescent virus 1 in farmed white leg shrimp, Penaeus vannamei. Aquaculture 2020, 533, 736105. [Google Scholar] [CrossRef]
- Srisala, J.; Sanguanrut, P.; Thaiue, D.; Laiphrom, S.; Siriwattano, J.; Khudet, J.; Powtongsook, S.; Flegel, T.W.; Sritunyalucksana, K. Infectious myonecrosis virus (IMNV) and Decapod iridescent virus 1 (DIV1) detected in captured, wild Penaeus monodon. Aquaculture 2021, 545, 737262. [Google Scholar] [CrossRef]
- Thitamadee, S.; Prachumwat, A.; Srisala, J.; Jaroenlak, P.; Salachan, P.V.; Sritunyalucksana, K.; Flegel, T.W.; Itsathitphaisarn, O. Review of current disease threats for cultivated penaeid shrimp in Asia. Aquaculture 2016, 452, 69–87. [Google Scholar] [CrossRef]
- Tassanakajon, A.; Somboonwiwat, K.; Supungul, P.; Tang, S. Discovery of immune molecules and their crucial functions in shrimp immunity. Fish Shellfish Immunol. 2013, 34, 954–967. [Google Scholar] [CrossRef]
- Chen, Y.-H.; He, J.-G. Effects of environmental stress on shrimp innate immunity and white spot syndrome virus infection. Fish Shellfish Immunol. 2018, 84, 744–755. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.; Dawood, M.A.; Alagawany, M.; Faggio, C.; Nowosad, J.; Kucharczyk, D. Health benefits and potential applications of fucoidan (FCD) extracted from brown seaweeds in aquaculture: An updated review. Fish Shellfish Immunol. 2022, 122, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Latif, H.M.; Abdel-Daim, M.M.; Shukry, M.; Nowosad, J.; Kucharczyk, D. Benefits and applications of Moringa oleifera as a plant protein source in Aquafeed: A review. Aquaculture 2021, 547, 737369. [Google Scholar] [CrossRef]
- Liu, X.-L.; Xi, Q.-Y.; Yang, L.; Li, H.-Y.; Jiang, Q.-Y.; Shu, G.; Wang, S.-B.; Gao, P.; Zhu, X.-T.; Zhang, Y.-L. The effect of dietary Panax ginseng polysaccharide extract on the immune responses in white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2011, 30, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.-L.; Hsieh, Y.-T. Immunostimulation of tiger shrimp (Penaeus monodon) hemocytes for generation of microbicidal substances: Analysis of reactive oxygen species. Dev. Comp. Immunol. 1994, 18, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Ji, P.-F.; Yao, C.-L.; Wang, Z.-Y. Immune response and gene expression in shrimp (Litopenaeus vannamei) hemocytes and hepatopancreas against some pathogen-associated molecular patterns. Fish Shellfish Immunol. 2009, 27, 563–570. [Google Scholar] [CrossRef]
- Söderhäll, I. Crustacean hematopoiesis. Dev. Comp. Immunol. 2016, 58, 129–141. [Google Scholar] [CrossRef]
- Liu, S.; Zheng, S.-C.; Li, Y.-L.; Li, J.; Liu, H.-P. Hemocyte-Mediated Phagocytosis in Crustaceans. Front. Immunol. 2020, 11, 268. [Google Scholar] [CrossRef]
- Clavero-Salas, A.; Sotelo-Mundo, R.R.; Gollas-Galván, T.; Hernández-López, J.; Peregrino-Uriarte, A.B.; Muhlia-Almazán, A.; Yepiz-Plascencia, G. Transcriptome analysis of gills from the white shrimp Litopenaeus vannamei infected with White Spot Syndrome Virus. Fish Shellfish Immunol. 2007, 23, 459–472. [Google Scholar] [CrossRef]
- Frischer, M.E.; Landers, S.C.; Walker, A.N.; Powell, S.A.; Lee, R.F. Black gill in marine decapod crustaceans: A review. Rev. Fish. Sci. Aquac. 2022, 30, 498–519. [Google Scholar] [CrossRef]
- Rao, R.; Zhu, Y.B.; Alinejad, T.; Tiruvayipati, S.; Thong, K.L.; Wang, J.; Bhassu, S. RNA-seq analysis of Macrobrachium rosenbergii hepatopancreas in response to Vibrio parahaemolyticus infection. Gut Pathog. 2015, 7, 6. [Google Scholar] [CrossRef]
- Maralit, B.A.; Jaree, P.; Boonchuen, P.; Tassanakajon, A.; Somboonwiwat, K. Differentially expressed genes in hemocytes of Litopenaeus vannamei challenged with Vibrio parahaemolyticus AHPND (VPAHPND) and VPAHPND toxin. Fish Shellfish Immunol. 2018, 81, 284–296. [Google Scholar] [CrossRef]
- Peruzza, L.; Shekhar, M.S.; Kumar, K.V.; Swathi, A.; Karthic, K.; Hauton, C.; Vijayan, K.K. Temporal changes in transcriptome profile provide insights of White Spot Syndrome Virus infection in Litopenaeus vannamei. Sci. Rep. 2019, 9, 13509. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Sun, X.; Zhou, Z.; Zhang, T.; Yi, Q.; Liu, R.; Wang, M.; Song, L. The promotion of cytoskeleton integration and redox in the haemocyte of shrimp Litopenaeus vannamei after the successive stimulation of recombinant VP28. Dev. Comp. Immunol. 2014, 45, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Peruzza, L.; Thamizhvanan, S.; Vimal, S.; Kumar, K.V.; Shekhar, M.; Smith, V.; Hauton, C.; Vijayan, K.; Hameed, A.S. A comparative synthesis of transcriptomic analyses reveals major differences between WSSV-susceptible Litopenaeus vannamei and WSSV-refractory Macrobrachium rosenbergii. Dev. Comp. Immunol. 2019, 104, 103564. [Google Scholar] [CrossRef] [PubMed]
- Tinwongger, S.; Thawonsuwan, J.; Kondo, H.; Hirono, I. Identification of an anti-lipopolysaccharide factor AV-R isoform (LvALF AV-R) related to Vp_PirAB-like toxin resistance in Litopenaeus vannamei. Fish Shellfish Immunol. 2019, 84, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.; Andrade, S.; Fernandes, J.; Freitas, P. Shedding the light on Litopenaeus vannamei differential muscle and hepatopancreas immune responses in White Spot Syndrome Virus (WSSV) Exposure. Genes 2020, 11, 805. [Google Scholar] [CrossRef]
- Liao, X.; Wang, C.; Wang, B.; Qin, H.; Hu, S.; Wang, P.; Sun, C.; Zhang, S. Comparative transcriptome analysis of Litopenaeus vannamei reveals that triosephosphate isomerase-like genes play an important role during Decapod Iridescent Virus 1 Infection. Front. Immunol. 2020, 11, 1904. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Huerta-Cepas, J.; Szklarczyk, D.; Heller, D.; Hernández-Plaza, A.; Forslund, S.K.; Cook, H.V.; Mende, D.R.; Letunic, I.; Rattei, T.; Jensen, L.J.; et al. eggNOG 5.0: A hierarchical, functionally and phylogenetically annotated orthology resource based on 5090 organisms and 2502 viruses. Nucleic Acids Res. 2018, 47, D309–D314. [Google Scholar] [CrossRef] [PubMed]
- Cantalapiedra, C.P.; Hernández-Plaza, A.; Letunic, I.; Bork, P.; Huerta-Cepas, J. eggNOG-mapper v2: Functional annotation, orthology assignments, and domain prediction at the metagenomic scale. Mol. Biol. Evol. 2021, 38, 5825–5829. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Yu, G. Gene ontology semantic similarity analysis using GOSemSim. In Stem Cell Transcriptional Networks: Methods and Protocols; Kidder, B.L., Ed.; Springer US: New York, NY, USA, 2020; pp. 207–215. [Google Scholar]
- Kidane, D.G.; Brækkan, E.H. Global Seafood demand growth differences across regions, income levels, and time. Mar. Resour. Econ. 2021, 36, 289–305. [Google Scholar] [CrossRef]
- Sarathi, M.; Ahmed, V.I.; Venkatesan, C.; Balasubramanian, G.; Prabavathy, J.; Hameed, A.S. Comparative study on immune response of Fenneropenaeus indicus to Vibrio alginolyticus and white spot syndrome virus. Aquaculture 2007, 271, 8–20. [Google Scholar] [CrossRef]
- Habib, Y.J.; Wan, H.; Sun, Y.; Shi, J.; Yao, C.; Lin, J.; Ge, H.; Wang, Y.; Zhang, Z. Genome-wide identification of toll-like receptors in Pacific white shrimp (Litopenaeus vannamei) and expression analysis in response to Vibrio parahaemolyticus invasion. Aquaculture 2021, 532, 735996. [Google Scholar] [CrossRef]
- Burnett, K.G.; Burnett, L.E. Respiratory and metabolic impacts of crustacean immunity: Are there implications for the insects? Integr. Comp. Biol. 2015, 55, 856–868. [Google Scholar] [CrossRef]
- Habib, Y.J.; Zhang, Z. The involvement of crustaceans toll-like receptors in pathogen recognition. Fish Shellfish Immunol. 2020, 102, 169–176. [Google Scholar] [CrossRef]
- Iwanaga, S.; Lee, B.-L. Recent advances in the innate immunity of invertebrate animals. BMB Rep. 2005, 38, 128–150. [Google Scholar] [CrossRef] [PubMed]
- Ji, P.; Yao, C.; Wang, Z. Reactive oxygen system plays an important role in shrimp Litopenaeus vannamei defense against Vibrio parahaemolyticus and WSSV infection. Dis. Aquat. Org. 2011, 96, 9–20. [Google Scholar] [CrossRef]
- Karthick, S.; Sunil Kumar, K.; Meiyalagan, V.; Arumugam, M. Comparative study on immunological responses in Scylla serrata and Litopenaeus vannamei upon WSSV infection. Aquac. Res. 2022, 53, 2249–2262. [Google Scholar] [CrossRef]
- Liu, C.-H.; Chang, C.-C.; Chiu, Y.-C.; Cheng, W.; Yeh, M.-S. Identification and cloning of a transglutaminase from giant freshwater prawn, Macrobrachium rosenbergii, and its transcription during pathogen infection and moulting. Fish Shellfish Immunol. 2011, 31, 871–880. [Google Scholar] [CrossRef]
- Yoganandhan, K.; Thirupathi, S.; Hameed, A.S. Biochemical, physiological and hematological changes in white spot syndrome virus-infected shrimp, Penaeus indicus. Aquaculture 2003, 221, 1–11. [Google Scholar] [CrossRef]
- Cheesman, H.K.; Feinbaum, R.L.; Thekkiniath, J.; Dowen, R.H.; Conery, A.L.; Pukkila-Worley, R. Aberrant activation of p38 MAP kinase-dependent innate immune responses is toxic to Caenorhabditis elegans. G3 Genes Genomes Genet. 2016, 6, 541–549. [Google Scholar] [CrossRef]
- Sun, B.; Wang, Z.; Wang, Z.; Ma, X.; Zhu, F. A Proteomic Study of hemocyte proteins from mud crab (Scylla paramamosain) infected with white spot syndrome virus or Vibrio alginolyticus. Front. Immunol. 2017, 8, 468. [Google Scholar] [CrossRef]
- Kumar, R.; Khandelwal, N.; Thachamvally, R.; Tripathi, B.N.; Barua, S.; Kashyap, S.K.; Maherchandani, S.; Kumar, N. Role of MAPK/MNK1 signaling in virus replication. Virus Res. 2018, 253, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Zhang, Y.; Liu, P.; Li, J. Comparative proteomic investigation of Marsupenaeus japonicus hepatopancreas challenged with Vibrio parahaemolyticus and white spot syndrome virus. Fish Shellfish Immunol. 2019, 93, 851–862. [Google Scholar] [CrossRef]
- Lin, C.; Zhang, Y.; Zhang, K.; Zheng, Y.; Lu, L.; Chang, H.; Yang, H.; Yang, Y.; Wan, Y.; Wang, S.; et al. Fever promotes t lymphocyte trafficking via a thermal sensory pathway involving heat shock protein 90 and α4 integrins. Immunity 2019, 50, 137–151.e6. [Google Scholar] [CrossRef] [PubMed]
- Yuan, K.; Yuan, F.-H.; He, H.-H.; Bi, H.-T.; Weng, S.-P.; He, J.-G.; Chen, Y.-H. Heat shock 70 kDa protein cognate 5 involved in WSSV toleration of Litopenaeus vannamei. Dev. Comp. Immunol. 2017, 72, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, F.; Sun, Z.; Zhang, X.; Xiang, Z. Differentially proteomic analysis of the Chinese shrimp at WSSV latent and acute infection stages by iTRAQ ap-proach. Fish Shellfish Immunol. 2016, 54, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Wang, Z.; Wang, X. Characterization of myosin light chain in shrimp hemocytic phagocytosis. Fish Shellfish Immunol. 2010, 29, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhu, F. Different roles of a novel shrimp microRNA in white spot syndrome virus (WSSV) and Vibrio alginolyticus infection. Dev. Comp. Immunol. 2018, 79, 21–30. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathogen | Type of Infection | Major Target Tissues | Diagnosis |
|---|---|---|---|
| WSSV | Vira | Gills, hematopoietic tissues, lymph | White spots on the shell, with longer blood coagulation time, pale, swollen and erosive hepatopancreas [11,12,13,14,15]. |
| DIV1 | Viral | Hematopoietic tissues hemocytes, hepatopancreas, gills and muscles | Softening of the shell, the yellowish-whitening of hepatopancreas, and empty stomach and intestine [16,17,18]. |
| VPAHPND | Bacterial | Gut-associated tissues and organs, including the hepatopancreas, stomach, and intestine | Softening of the shrimp body, atrophy and whitening of hepatopancreas, reduced feeding with empty stomach and intestine [6,19]. |
| BioProject | Tissue | Pathogen | N | Remarks: Homogenization of Data (H) or Limitation (L) | DOI of Publications |
|---|---|---|---|---|---|
| PRJNA524934 | Gills | WSSV | 18 | H: Data from 3 times points of WSSV challenge treatment were grouped as the “WSSV-challenged group”, and data from the 3 time points of control treatment were grouped as “control group”. | 10.1038/s41598-019-49836-0 [33] |
| PRJNA716175 | Gills | WSSV | 48 | H: Data from 6 times points of WSSV challenge treatment were grouped as the “WSSV-challenged group”, and data from the 6 time points of control treatment were grouped as “control group”. | 10.3390/v13061140 [4] |
| PRJNA233549 | Hemocyte | WSSV | 8 | H: No homogenization needed. L: WSSV isolate was not used in this study; instead, VP28, the major envelope proteins of WSSV was injected to shrimp to reveal immune response at the initial stage of infection. | 10.1016/j.dci.2014.02.013 [34] |
| PRJNA448614 | Hemocyte | VPAHPND | 3 | H: Data from 2 times points of VPAHPND challenge treatment were grouped as the “VPAHPND-challenged group”, and data from the 2 time points of control treatment were grouped as “control group”. L: There were only 3 RNA-seq libraries in this study as it was difficult to yield adequate amount of RNA for library construction from hemocyte samples, but each library includes 10 individuals of VPAHPND-challenged shrimp. | 10.1016/j.fsi.2018.06.054 [32] |
| PRJNA612147 | Hemocyte | DIV1 | 6 | H: No homogenization needed. | 10.3389/fimmu.2020.01904 [38] |
| PRJNA428228 | Hepatopancreas | WSSV | 8 | H: Only hepatopancreas RNA-seq expression data was used. | 10.3390/genes11070805 [37] |
| PRJNA554075 | Hepatopancreas | WSSV | 6 | H: Only RNA-seq expression data in P. vannamei was used. | 10.1016/j.dci.2019.103564 [35] |
| PRJNA413606 | Hepatopancreas | VPAHPND | 6 | H: Only hepatopancreas RNA-seq expression data was used. | 10.1016/j.fsi.2018.10.005 [36] |
| PRJNA421143 | Hepatopancreas | VPAHPND | 6 | H: No homogenization needed. | Not available |
| BioProject | EdgeR | Limma | Deseq2 | DEGs | Unigenes (DEGs) in Tissue | Tissue | Pathogen |
|---|---|---|---|---|---|---|---|
| PRJNA524934 | 1842 | 78 | 630 | 473 | 400 | Gills | WSSV |
| PRJNA716175 | 353 | 2 | 179 | 156 | Gills | WSSV | |
| PRJNA233549 | 173 | 239 | - | 27 | 84 | Hemocyte | WSSV |
| PRJNA448614 | 29 | 83 | - | 7 | Hemocyte | VPAHPND | |
| PRJNA612147 | 186 | 55 | 13 | 50 | Hemocyte | DIV1 | |
| PRJNA413606 | 53 | 54 | 1 | 19 | 434 | Hepatopancreas | VPAHPND |
| PRJNA421143 | 612 | 1289 | 240 | 609 | Hepatopancreas | VPAHPND | |
| PRJNA428228 | 19 | 231 | - | 17 | Hepatopancreas | WSSV | |
| PRJNA554075 | 83 | 280 | 44 | 78 | Hepatopancreas | WSSV |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Z.; Chu, K.H.; Ma, K.Y. Transcriptome Analysis of Multiple Tissues in the Shrimp Penaeus vannamei Reveals the Typical Physiological Response to Three Pathogens. J. Mar. Sci. Eng. 2023, 11, 389. https://doi.org/10.3390/jmse11020389
Wu Z, Chu KH, Ma KY. Transcriptome Analysis of Multiple Tissues in the Shrimp Penaeus vannamei Reveals the Typical Physiological Response to Three Pathogens. Journal of Marine Science and Engineering. 2023; 11(2):389. https://doi.org/10.3390/jmse11020389
Chicago/Turabian StyleWu, Ziwei, Ka Hou Chu, and Ka Yan Ma. 2023. "Transcriptome Analysis of Multiple Tissues in the Shrimp Penaeus vannamei Reveals the Typical Physiological Response to Three Pathogens" Journal of Marine Science and Engineering 11, no. 2: 389. https://doi.org/10.3390/jmse11020389
APA StyleWu, Z., Chu, K. H., & Ma, K. Y. (2023). Transcriptome Analysis of Multiple Tissues in the Shrimp Penaeus vannamei Reveals the Typical Physiological Response to Three Pathogens. Journal of Marine Science and Engineering, 11(2), 389. https://doi.org/10.3390/jmse11020389