
Comparison of Viral Production and Decay Rates at the Surface and Bottom of the Euphotic Zone in the Summertime in the Southern East China Sea



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Samplings
2.2. Experiments on Viral Production and Decay
2.3. Statistical Analysis
3. Results
3.1. Environmental Dynamics
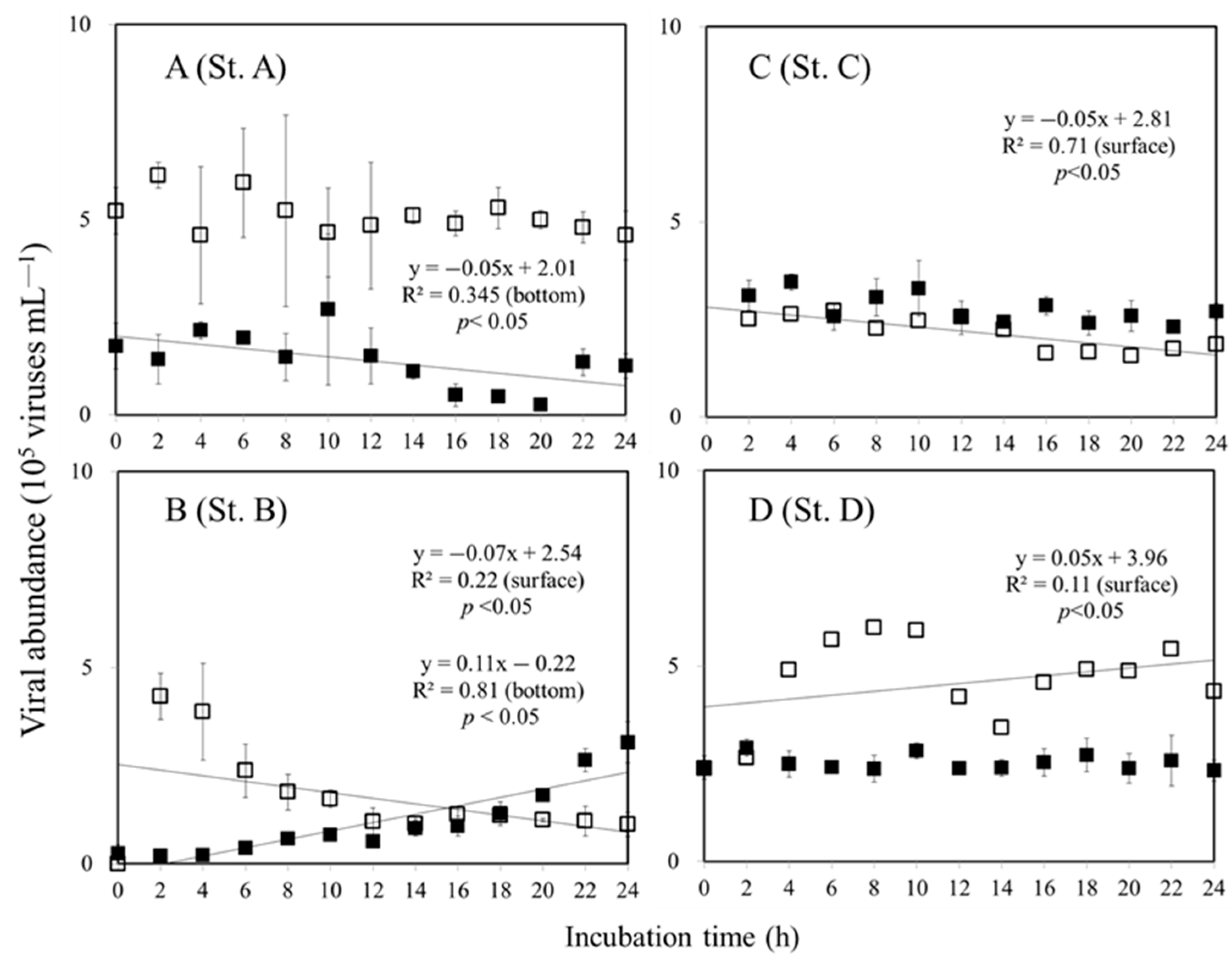
3.2. Viral Production and Decay Rate in Surface and Bottom of the Euphotic Zone
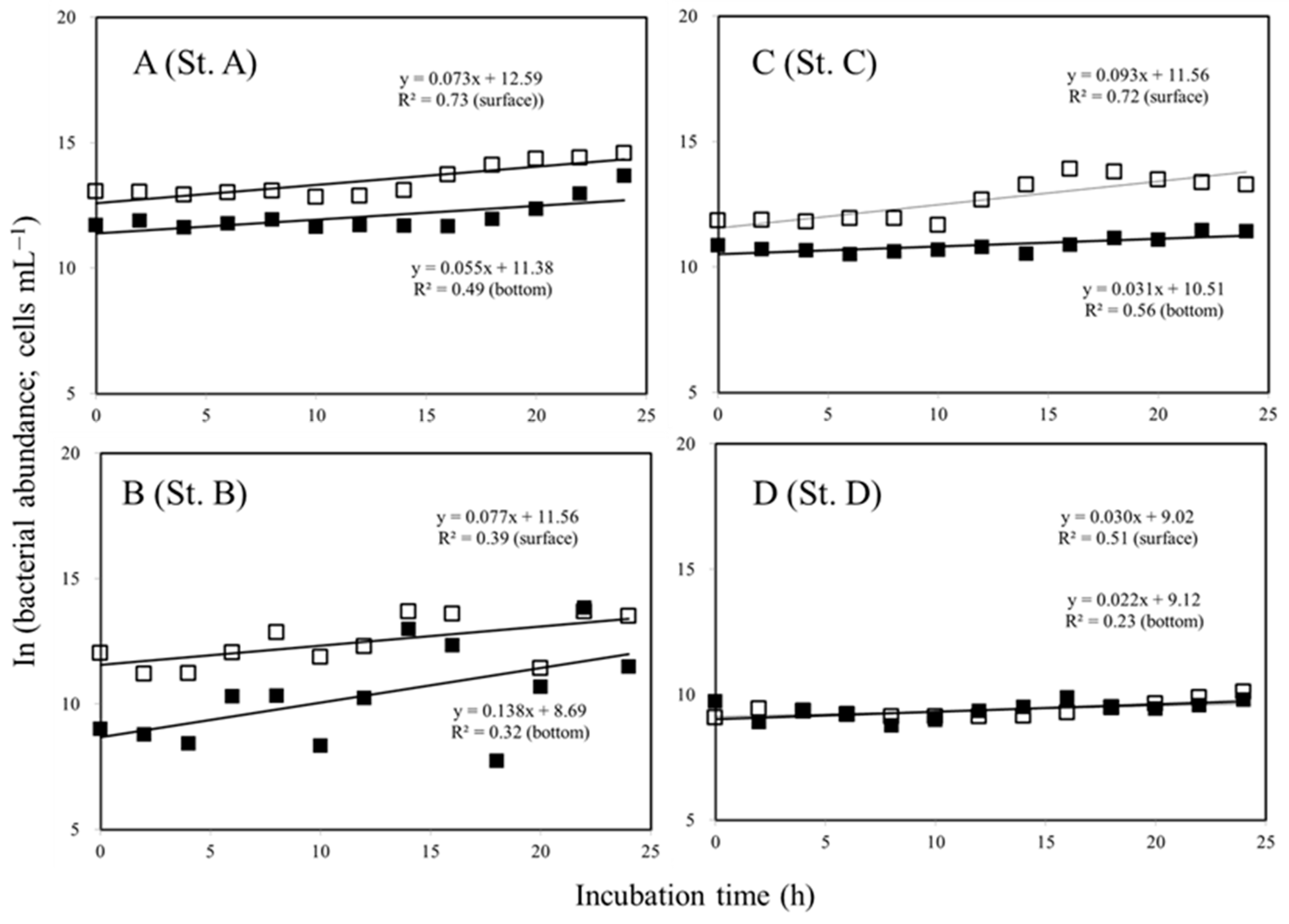
3.3. Variations in the Bacterial Growth in the Surface and Bottom Water
4. Discussion
4.1. Variations in Virus to Bacteria Ratio (VBR)
4.2. Variations in Viral Production
4.3. Variations in Viral Decay
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Azam, F.; Fenchel, T.; Field, J.G.; Gray, J.S.; Meyer-Reil, L.A.; Thingstad, F. The ecological role of water column microbes in the sea. Mar. Ecol. Prog. Ser. 1983, 10, 257–263. [Google Scholar] [CrossRef]
- Fuhrman, J.A.; Noble, R.T. Viruses and protists cause similar bacterial mortality in coastal seawater. Limnol. Oceanogr. 1995, 40, 1236–1242. [Google Scholar] [CrossRef]
- Tsai, A.Y.; Chiang, K.P.; Chang, J.; Gong, G.C. Seasonal diel variations of picoplankton and nanoplankton in a subtropical western Pacific coastal ecosystem. Limnol. Oceanogr. 2005, 50, 1221–1231. [Google Scholar] [CrossRef]
- Pasulka, A.L.; Samo, T.J.; Landry, M.R. Grazer and viral impacts on microbial growth and mortality in the southern California Current Ecosystem. J. Plankton Res. 2015, 37, 320–336. [Google Scholar] [CrossRef]
- Weinbauer, M.G.; Höfle, M.G. Significance of viral lysis and flagellate grazing as factors controlling bacterioplankton production in a eutrophic lake. Appl. Environ. Microbiol. 1998, 64, 431–438. [Google Scholar] [CrossRef]
- Lymer, D.; Lindstrom, E.S.; Vrede, K. Changing importance of viral induced bacterial mortality in lakes along gradients in trophic status and humic content. Freshw. Biol. 2008, 53, 1101–1113. [Google Scholar] [CrossRef]
- Taira, Y.; Uchimiya, M.; Kudo, I. Simultaneous estimation of viral lysis and protozoan grazing on bacterial mortality using a modified virus-dilution method. Mar. Ecol. Prog. Ser. 2009, 379, 23–32. [Google Scholar] [CrossRef]
- Tsai, A.Y.; Gong, G.C.; Chao, C.F. Contribution of viral lysis and nanoflagellate grazing to bacterial mortality at surface waters and deeper depths in the coastal ecosystem of subtropical Western Pacific. Estuar. Coasts 2016, 39, 1357–1366. [Google Scholar] [CrossRef]
- Tsai, A.Y. Effects of bacteria-virus interaction on Synechococcus spp. growth in the coastal waters. Terr. Atmo. Ocean. Sci. 2020, 31, 691–696. [Google Scholar] [CrossRef]
- Chen, X.; Liu, H.; Weinbauer, M.; Chen, B.; Jiao, N. Viral dynamics in the surface water of the western South China Sea in summer 2007. Aquat. Microb. Ecol. 2011, 63, 145–160. [Google Scholar] [CrossRef]
- Weinbauer, M.G.; Rassoulzadegan, F. Are viruses driving microbial diversification and diversity? Environ. Microbiol. 2004, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Danovaro, R.; Dell’Anno, A.; Corinaldesi, C.; Magagnini, M.; Noble, R.; Tamburini, C.; Weinbauer, M. Major viral impact on the functioning of benthic deep-sea ecosystems. Nature 2008, 454, 1084–1087. [Google Scholar] [CrossRef] [PubMed]
- Haaber, J.; Middelboe, M. Viral lysis of Phaeocystis pouchetii: Implications for algal population dynamics and heterotrophic C, N and P cycling. ISME J. 2009, 3, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Shelford, E.J.; Middelboe, M.; Moller, E.F.; Suttle, C.A. Virus-driven nitrogen cycling enhances phytoplankton growth. Aquat. Microb. Ecol. 2012, 66, 41–46. [Google Scholar] [CrossRef]
- Salter, I.; Böttjet, D.; Christaki, U. The effect of inorganic particle concentration on bacteria–virus–nanoflagellate dynamics. Environ. Microbiol. 2011, 13, 2768–2777. [Google Scholar] [CrossRef]
- Tsai, A.-Y.; Lin, Y.-T.; Gong, G.-C. Effect of the presence of Virus-like Particles on Bacterial Growth in Sunlit Surface and Dark Deep Ocean Environments in the Southern East China Sea. Water 2021, 13, 2934. [Google Scholar] [CrossRef]
- Gobler, C.J.; Davis, T.W.; Deonarine, S.N.; Saxton, M.A.; Lavrentyev, P.J.; Jochem, F.J.; Wilhelm, S.W. Grazing and virus induced mortality of microbial populations before and during the onset of annual hypoxia in Lake Erie. Aquat. Microb. Ecol. 2008, 51, 117–128. [Google Scholar] [CrossRef]
- Colombet, J.; Sime-Ngando, T.; Cauchie, H.M.; Fonty, G.; Hoffmann, L.; Demeure, G. Depth-related gradients of viral activity in Lake Pavin. Appl. Environ. Microbiol. 2006, 72, 4440–4445. [Google Scholar] [CrossRef]
- He, L.; Yin, K.; Yuan, X.; Li, D.; Zhang, D.; Harrison, P.J. Spatial distribution of viruses, bacteria and chlorophyll in the northern South China Sea. Aquat. Microb. Ecol. 2009, 54, 153–162. [Google Scholar] [CrossRef]
- Wommack, K.E.; Colwell, R.R. Virioplankton: Viruses in aquatic ecosystems. Microbiol. Mol. Biol. Rev. 2000, 64, 69–114. [Google Scholar] [CrossRef]
- Alonso, M.C.; Jimenez-Gomez, F.; Rodriguez, J.; Borrego, J.J. Distribution of virus-like particles in an oligotrophic marine environment (Alboran Sea, western Mediterranean). Microb. Ecol. 2001, 42, 407–415. [Google Scholar] [CrossRef]
- Culley, A.I.; Welschmeyer, N.A. The abundance, distribution, and correlation of viruses, phytoplankton, and prokaryotes along a Pacific Ocean transect. Limnol. Oceanogr. 2002, 47, 1508–1513. [Google Scholar] [CrossRef]
- Jiang, S.C.; Paul, J.H. Seasonal and diel abundance of viruses and the occurrence of lysogeny/bacteriocinogeny in the marine environment. Mar. Ecol. Prog. Ser. 1994, 104, 163–172. [Google Scholar] [CrossRef]
- Maranger, R.; Bird, D.F. Viral abundances in aquatic systems: A comparison between marine and fresh waters. Mar. Ecol. Prog. Ser. 1995, 121, 217–226. [Google Scholar] [CrossRef]
- Taylor, G.T.; Hein, C.; Iabichella, M. Temporal variations in viral distributions in the anoxic Cariaco Basin. Aquat. Microb. Ecol. 2003, 30, 103–116. [Google Scholar] [CrossRef]
- Magiopoulos, I.; Pitta, P. Viruses in a deep oligotrophic sea: Seasonal distribution of marine viruses in the epi-, meso- and bathypelagic waters of the Eastern Mediterranean Sea. Deep Sea Res. Part I 2012, 66, 1–10. [Google Scholar] [CrossRef]
- Yager, P.L.; Connelly, T.L.; Mortazavi, B.; Wommack, K.E.; Bano, N.; Bauer, J.E.; Opsahl, S.; Hollibaugh, J.T. Dynamic bacterial and viral response to an algal bloom at subzero temperatures. Limnol. Oceanogr. 2001, 46, 790–801. [Google Scholar] [CrossRef]
- Clasen, J.L.; Brigden, S.M.; Payet, J.P.; Suttle, C.A. Evidence that viral abundance across oceans and lakes is driven by different biological factors. Freshw. Biol. 2008, 53, 1090–1100. [Google Scholar] [CrossRef]
- Parada, V.; Sintes, E.; Van Aken, H.M.; Weinbauer, M.G.; Herndl, N.J. Viral abundance, decay, and diversity in the meso- and bathypelagic waters of the north atlantic. Appl. Environ. Microbiol. 2007, 73, 4429–4438. [Google Scholar] [CrossRef]
- Wei, W.; Chen, X.; Weinbauer, M.G.; Jiao, N.; Zhang, R. Reduced bacterial mortality and enhanced viral productivity during sinking in the ocean. ISME J. 2022, 16, 1668–1675. [Google Scholar] [CrossRef]
- Mei, M.L.; Danovaro, R. Virus production and life strategies in aquatic sediments. Limnol. Oceanogr. 2004, 49, 459–470. [Google Scholar] [CrossRef]
- Gong, G.C.; Shiah, F.K.; Liu, K.K.; Wen, Y.H.; Liang, M.H. Spatial and temporal variation of chlorophyll a, primary productivity and chemical hydrography in the southern East China Sea. Cont. Shelf. Res. 2000, 20, 411–436. [Google Scholar] [CrossRef]
- Brussaard, C.P.D. Optimization of procedures for counting viruses by flow cytometry. Appl. Environ. Microbiol. 2004, 70, 1506–1513. [Google Scholar] [CrossRef]
- Hammes, F.; Egli, T. Cytometric methods for measuring bacteria in water: Advantages, pitfalls and applications. Anal. Bioanal. Chem. 2010, 397, 1083–1095. [Google Scholar] [CrossRef]
- Wilhelm, S.W.; Bridgen, S.M.; Suttle, C.A. A dilution technique for the direct measurement of viral production: A comparison in stratified and tidally mixed coastal waters. Microb. Ecol. 2002, 43, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Noble, R.T.; Fuhrman, J.A. Viral decay and its causes in coastal waters. Appl. Environ. Microbiol. 1997, 63, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.L.; Tseng, Y.H.; Jan, S. The formation and dynamics of the cold-dome off northeastern Taiwan. J. Mar. Syst. 2011, 86, 10–27. [Google Scholar] [CrossRef]
- Suttle, C.A. Viruses in the sea. Nature 2005, 437, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Auguet, J.C.; Montanié, H.; Delmas, D.; Hartmann, H.J.; Huet, V. Dynamic of virioplankton abundance and its environmental control in the Charente Estuary (France). Microb. Ecol. 2005, 50, 337–349. [Google Scholar] [CrossRef]
- Boehme, J.; Frischer, M.E.; Jiang, S.C.; Kellogg, C.A.; Pichard, S.; Rose, J.B.; Steinway, C.; Paul, J.H. Viruses, bacterioplankton, and phytoplankton in the southeastern Gulf of Mexico: Distribution and contribution to oceanic DNA pools. Mar. Ecol. Prog. Ser. 1993, 97, 1–10. [Google Scholar] [CrossRef]
- Wilcox, R.M.; Fuhrman, J.A. Bacterial viruses in coastal seawater: Lytic rather than lysogenic production. Mar. Ecol. Prog. Ser. 1994, 114, 35–45. [Google Scholar] [CrossRef]
- Wigington, C.H.; Sonderegger, D.; Brussaard, C.P.; Buchan, A.; Finke, J.F.; Fuhrman, J.A.; Lennon, J.T.; Middelboe, M.; Suttle, C.A.; Stock, C. Re-examination of the relationship between marine virus and microbial cell abundances. Nat. Microbiol. 2016, 1, 15024. [Google Scholar] [CrossRef] [PubMed]
- Brussaard, C.P.D.; Wilhelm, S.W.; Thingstad, F.; Weinbauer, M.G.; Bratbak, G.; Heldal, M.; Kimmance, S.A.; Middelboe, M.; Nagasaki, K.; Schroeder, D.C.; et al. Global-scale processes with a nanoscale drive: The role of marine viruses. ISME J. 2008, 2, 575–578. [Google Scholar] [CrossRef]
- Wilhelm, S.W.; Matteson, A.R. Freshwater and marine virioplankton: A brief overview of commonalities and differences. Freshw. Biol. 2008, 53, 1076–1089. [Google Scholar] [CrossRef]
- Thomas, R.; Berdjeb, L.; Sime-Ngando, T.; Jacquet, S. Viral abundance, production, decay rates and life strategies (lysogeny versus lysis) in Lake Bourget (France). Environ. Microbiol. 2011, 13, 616–630. [Google Scholar] [CrossRef]
- Bongiorni, L.; Magagnini, M.; Armeni, M.; Noble, R.; Danovaro, R. Viral production, decay rates, and life strategies along a trophic gradient in the north Adriatic sea. Appl. Environ. Microbiol. 2005, 71, 6644–6650. [Google Scholar] [CrossRef]
- Steward, G.F.; Wikner, J.; Cochlan, W.P.; Smith, D.C.; Azam, F. Estimation of virus production in the sea: II. Field results. Mar. Microb. Food Webs 1992, 6, 79–90. [Google Scholar]
- Weinbauer, M.G.; Fuks, D.; Peduzzi, P. Distribution of viruses and dissolved DNA along a coastal trophic gradient in the northern Adriatic Sea. Appl. Environ. Microbiol. 1993, 59, 4074–4082. [Google Scholar] [CrossRef]
- Holmfeldt, K.; Titelman, J.; Riemann, L. Virus productive and lysate recycling in different sub-basins of the Northern Baltic Sea. Microb. Ecol. 2010, 60, 572–580. [Google Scholar] [CrossRef]
- Suttle, C.A.; Chen, F. Mechanisms and rates of decay of manne viruses in seawater. Appl. Environ. Mlcrobiol. 1992, 58, 3721–3729. [Google Scholar] [CrossRef]
- Wilhelm, S.W.; Weinbauer, M.G.; Suttle, C.A.; Jeffrey, W.H. The role of sunlight in the removal and repair of viruses in the sea. Limnol. Oceanogr. 1998, 43, 586–592. [Google Scholar] [CrossRef]
- Lawrence, J.E.; Chan, A.M.; Suttle, C.A. Viruses causing lysis of the toxic bloom-forming alga, Heterosigma akashiwo (Raphidophyceae), are widespread in coastal sediments of British Columbia, Canada. Limnol. Oceanogr. 2002, 47, 545–550. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Depth (m) | Temperature (°C) | Salinity | PAR (μEm−2 s−1) | Chl a (µg L−1) | Virus (105 Viruses mL−1) | Bacteria (105 Cells mL−1) | VBR |
|---|---|---|---|---|---|---|---|---|
| A | 3 | 27.5 | 34.0 | 400.5 | 0.31 | 27.6 ± 1.9 | 19.1 ± 0.9 | 1.4 ± 1.2 |
| 30 | 23.8 | 34.3 | 12.5 | 0.12 | 12.6 ± 1.4 | 6.1 ± 0.9 | 2.1 ± 1.2 | |
| B | 5 | 28.5 | 33.6 | 155.6 | 0.13 | 21.8 ± 1.8 | 8.4 ± 0.3 | 2.6 ± 1.3 |
| 100 | 17.7 | 34.6 | 1.7 | 0.04 | 2.9 ± 0.7 | 0.3 ± 0.06 | 9.7 ± 0.6 | |
| C | 5 | 25.8 | 34.0 | 568.3 | 0.40 | 14.2 ± 2.6 | 6.3 ± 1.2 | 2.3 ± 1.6 |
| 200 | 16.9 | 34.6 | 0.2 | 0.03 | 4.2 ± 1.5 | 0.3 ± 0.1 | 14.0 ± 1.2 | |
| D | 5 | 29.1 | 34.1 | 508.6 | 0.04 | 18.4 ± 1.1 | 5.8 ± 1.6 | 3.2 ± 1.1 |
| 200 | 19.7 | 34.8 | 0.3 | 0.03 | 8.1 ± 0.3 | 0.5 ± 0.2 | 16.2 ± 0.2 |
| Station | Depth (m) | NVP (105 Viruses mL−1 h−1) | VD (105 Viruses mL−1 h−1) | GVP (105 Viruses mL−1 h−1) |
|---|---|---|---|---|
| A | 3 | ND | 0.09 ± 0.02 | 0.09 ± 0.01 |
| 30 | −0.05 ± 0.02 | 0.08 ± 0.02 | 0.03 ± 0.01 | |
| B | 5 | −0.07 ± 0.03 | 0.23 ± 0.05 | 0.16 ± 0.02 |
| 100 | 0.11 ± 0.02 | ND | 0.11 ± 0.02 | |
| C | 5 | −0.05 ± 0.01 | 0.18 ± 0.04 | 0.13 ± 0.03 |
| 200 | ND | ND | ND | |
| D | 5 | 0.05 ± 0.01 | 0.13 ± 0.02 | 0.18 ± 0.03 |
| 200 | ND | ND | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, P.W.-Y.; Olivia, M.; Mukhanov, V.; Tsai, A.-Y. Comparison of Viral Production and Decay Rates at the Surface and Bottom of the Euphotic Zone in the Summertime in the Southern East China Sea. J. Mar. Sci. Eng. 2023, 11, 364. https://doi.org/10.3390/jmse11020364
Chen PW-Y, Olivia M, Mukhanov V, Tsai A-Y. Comparison of Viral Production and Decay Rates at the Surface and Bottom of the Euphotic Zone in the Summertime in the Southern East China Sea. Journal of Marine Science and Engineering. 2023; 11(2):364. https://doi.org/10.3390/jmse11020364
Chicago/Turabian StyleChen, Patrichka Wei-Yi, Madeline Olivia, Vladimir Mukhanov, and An-Yi Tsai. 2023. "Comparison of Viral Production and Decay Rates at the Surface and Bottom of the Euphotic Zone in the Summertime in the Southern East China Sea" Journal of Marine Science and Engineering 11, no. 2: 364. https://doi.org/10.3390/jmse11020364
APA StyleChen, P. W.-Y., Olivia, M., Mukhanov, V., & Tsai, A.-Y. (2023). Comparison of Viral Production and Decay Rates at the Surface and Bottom of the Euphotic Zone in the Summertime in the Southern East China Sea. Journal of Marine Science and Engineering, 11(2), 364. https://doi.org/10.3390/jmse11020364