
Exploring the Status of an Urban Coral and the Presence of Potential Probiotic Traits in Culturable Bacteria

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Permits
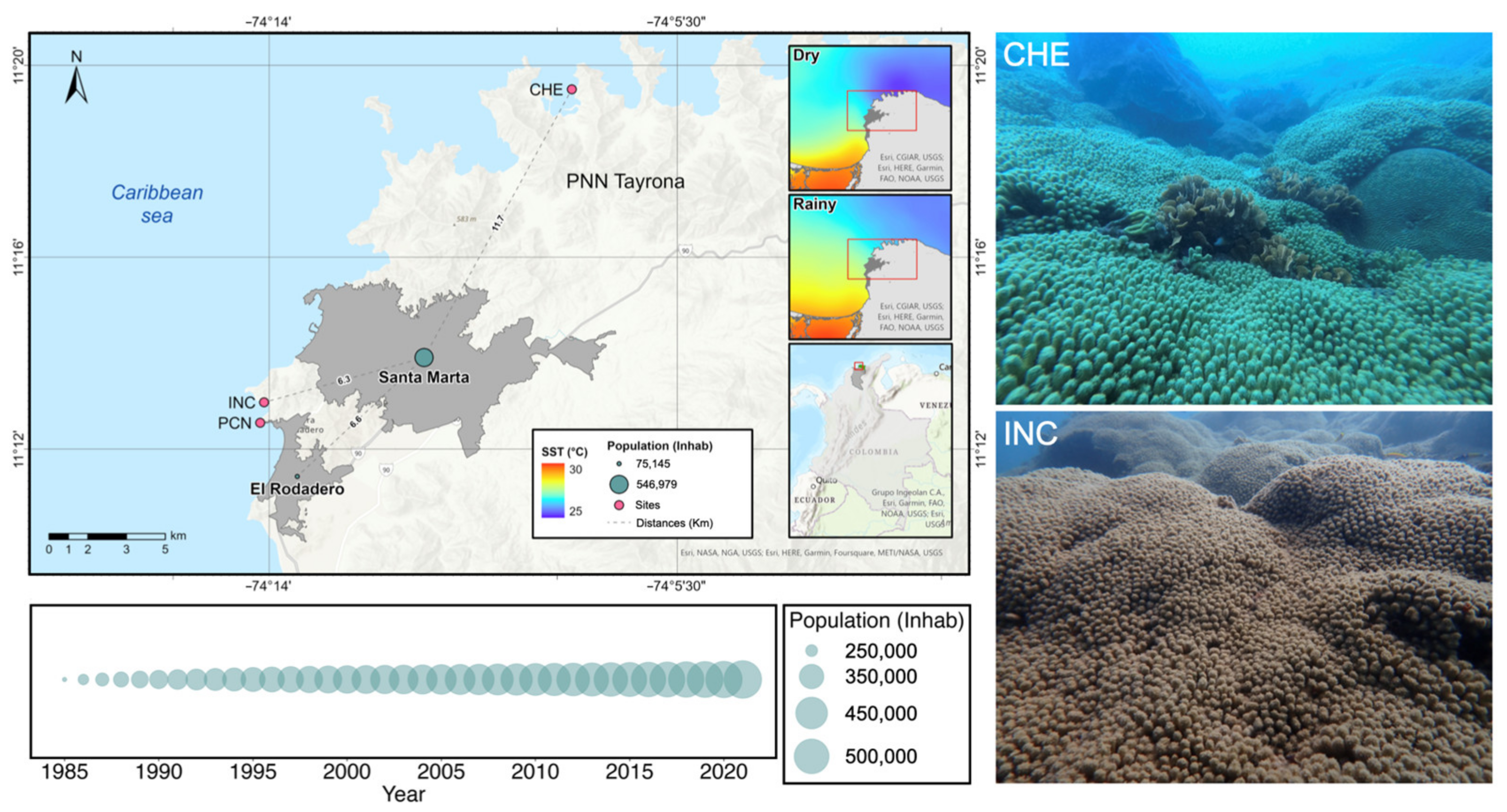
2.2. Study Area
2.3. Evaluation of Madracis auretenra Reef Patches Status
2.3.1. Sample Collection
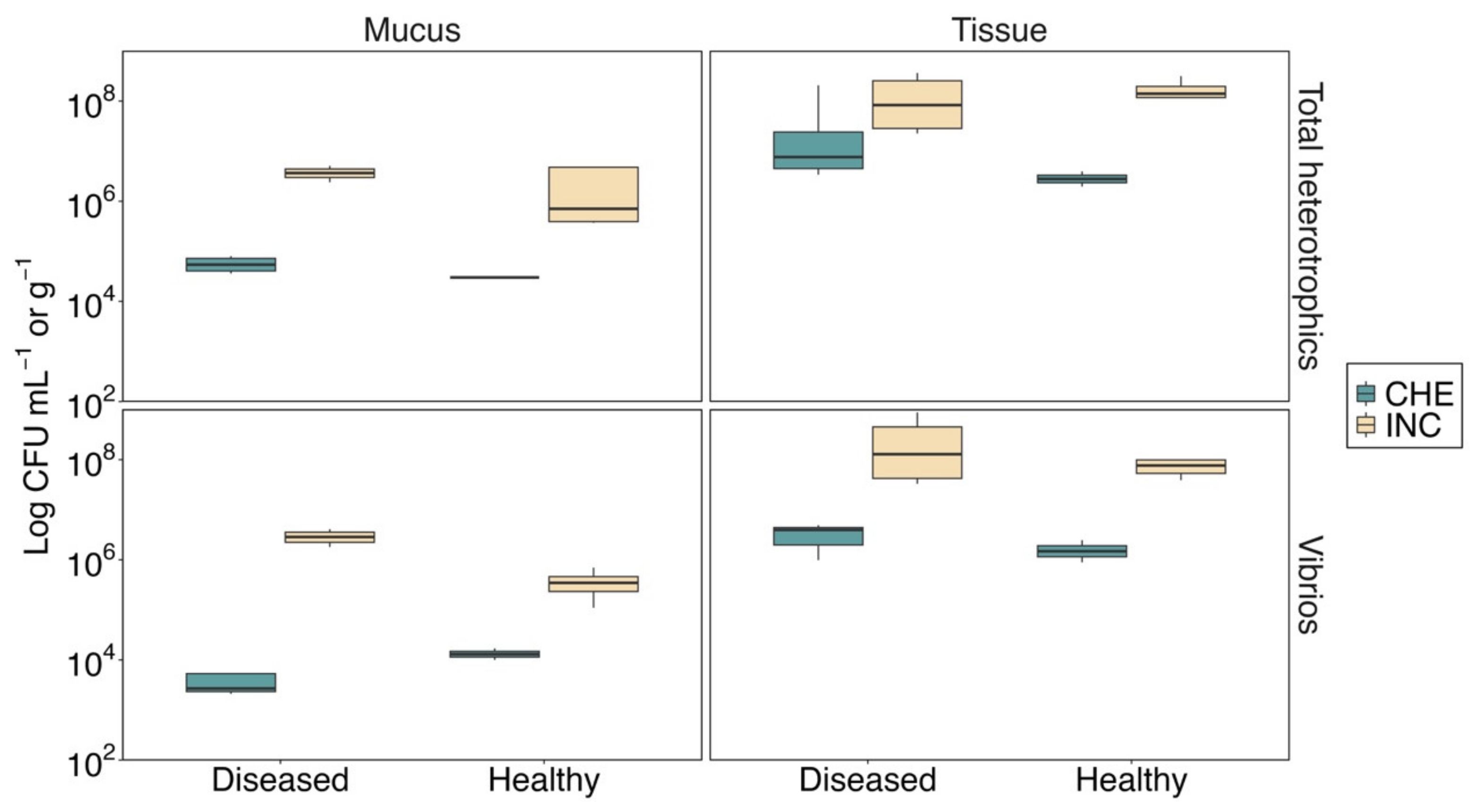
2.3.2. Counts of Total Heterotrophic Bacteria and Vibrios
2.3.3. Benthic Community and Health Status
2.3.4. Physicochemical Analysis
2.3.5. Statistical Analysis
2.4. Characterization of Culturable Bacteria and Screening of Probiotic Traits
2.4.1. Coral-Associated Bacterial Isolation
2.4.2. Qualitative Functional Screening of Coral Probiotic Traits
Antioxidant Activity and Pigment Production
Antagonistic and antiQS Activities
Siderophores Production
2.4.3. DNA Extraction and Identification via 16S rRNA Gene Sequence Analysis
2.4.4. Phylogenetic Analysis
2.4.5. Data Analysis
3. Results
3.1. Environmental and Microbiological Conditions
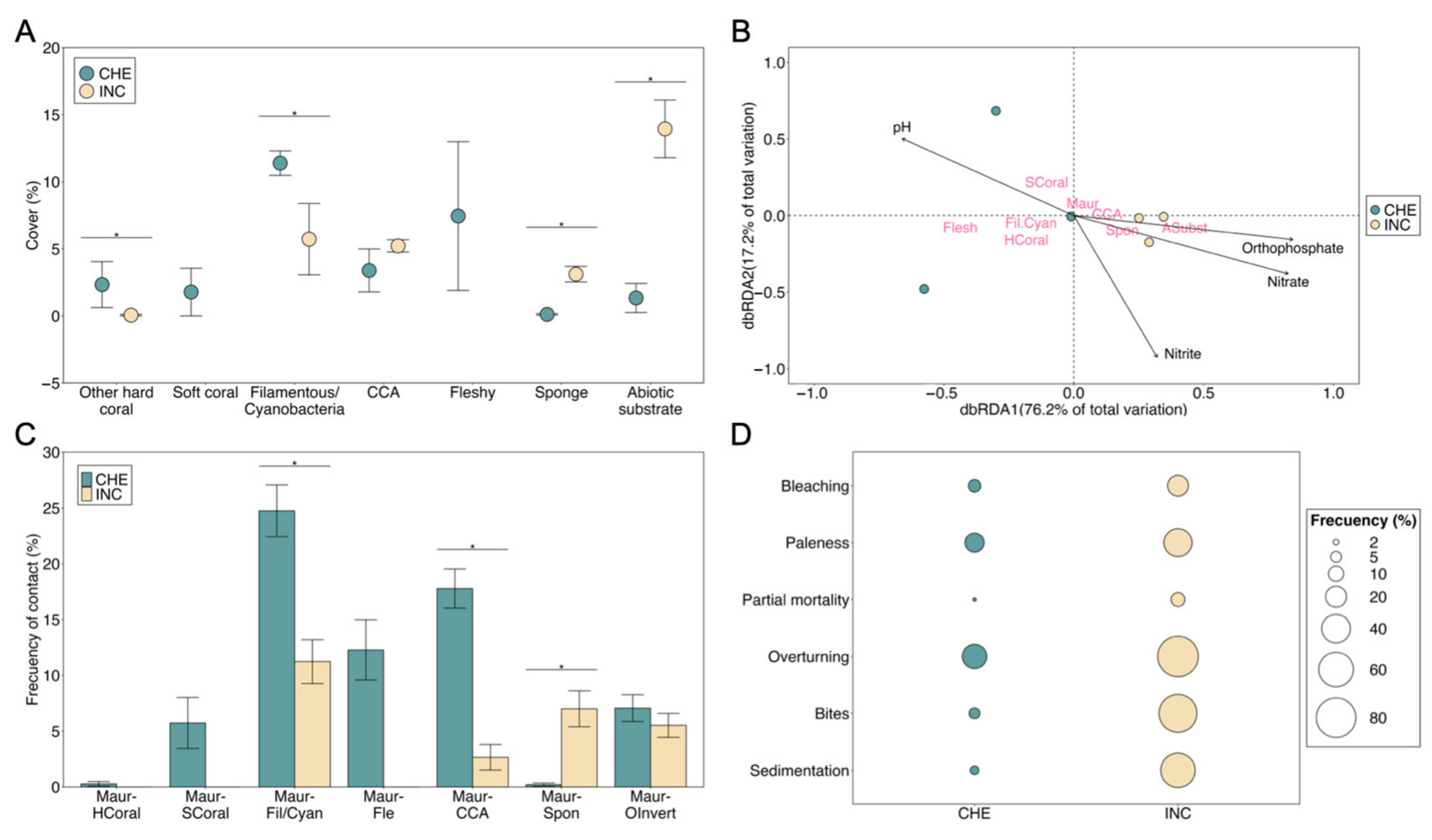
3.2. Benthic Community and Health Status
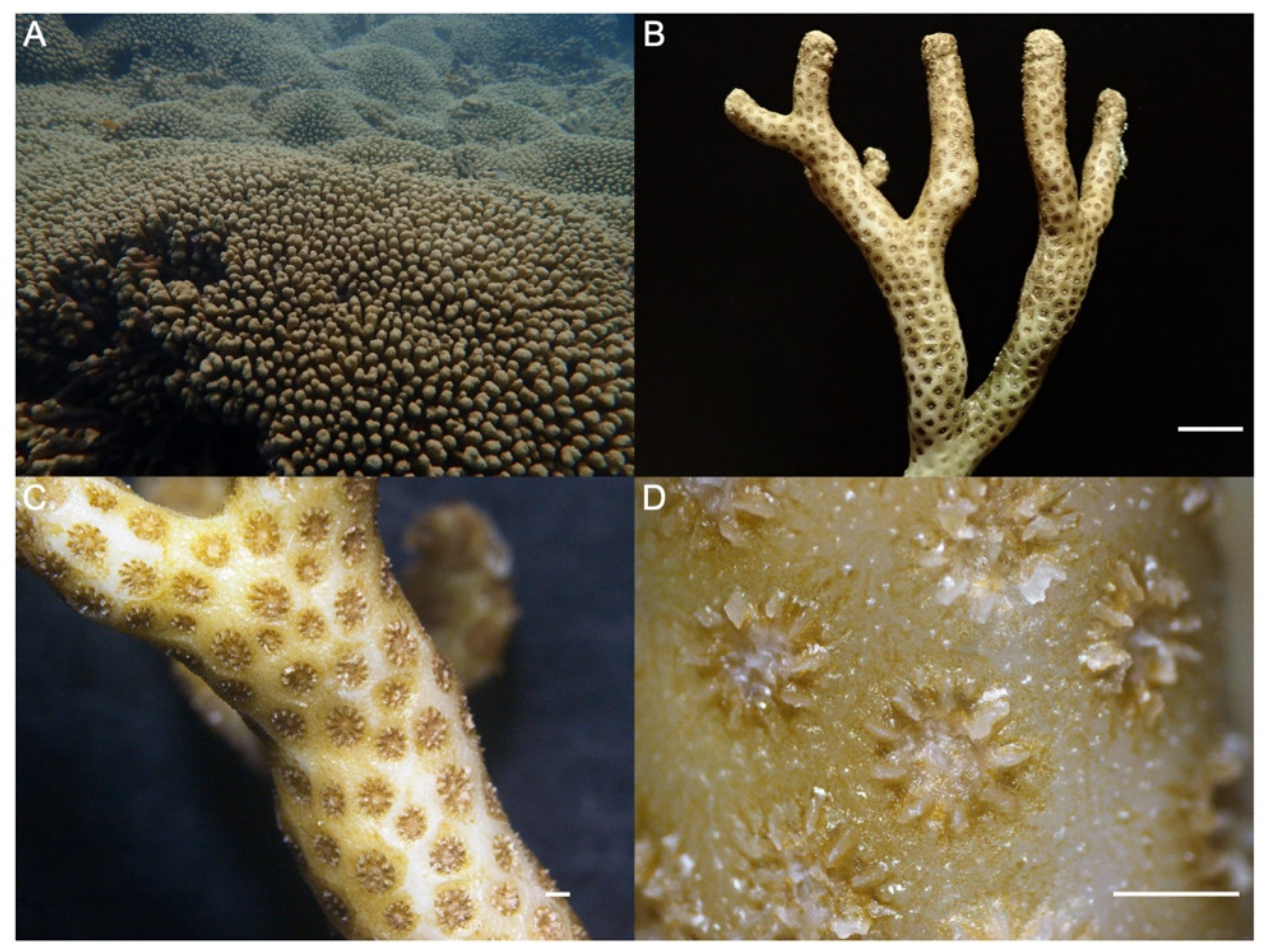
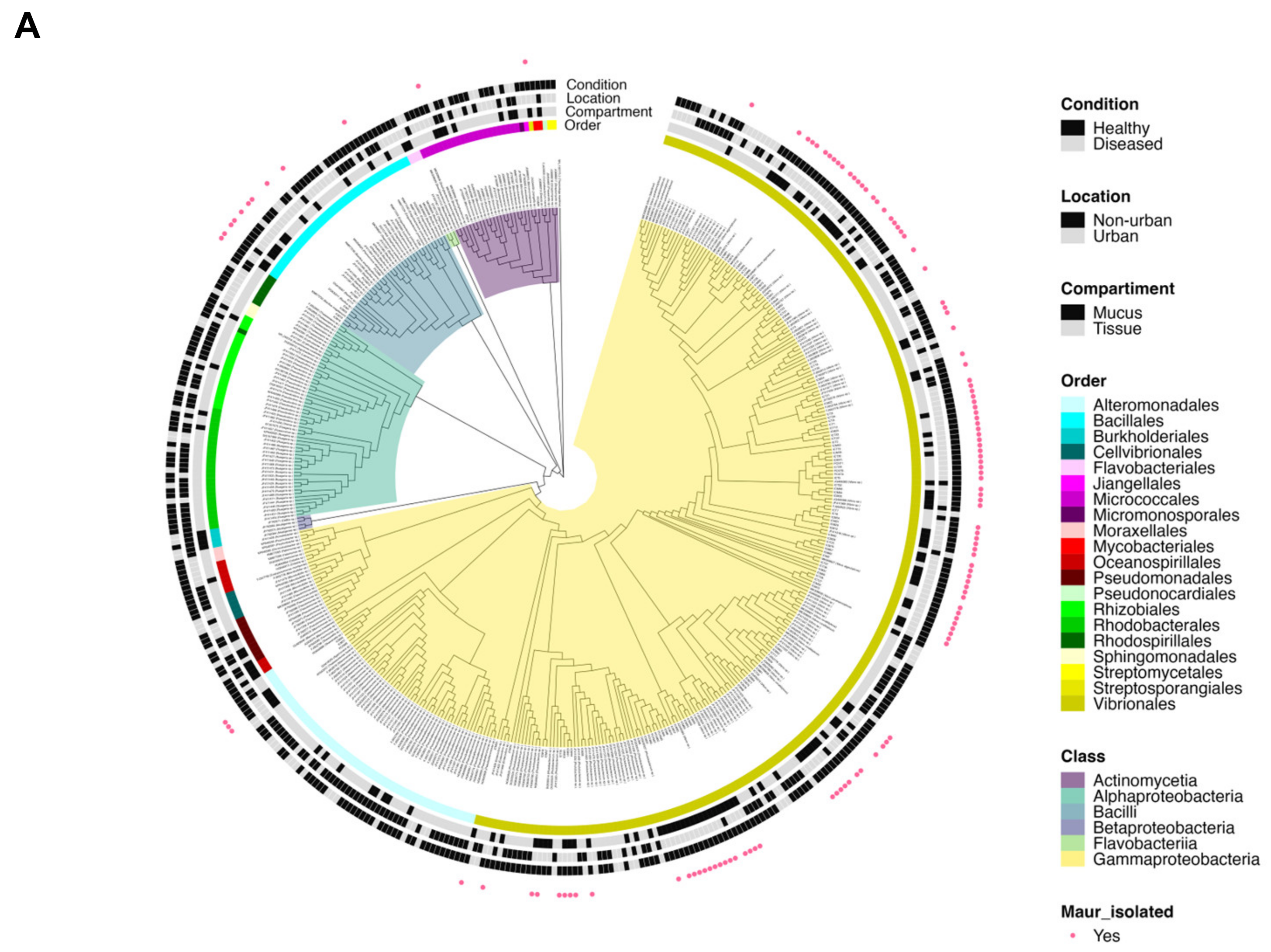
3.3. M. auretenra-Associated Culturable Bacteria and Phylogenetic Analysis
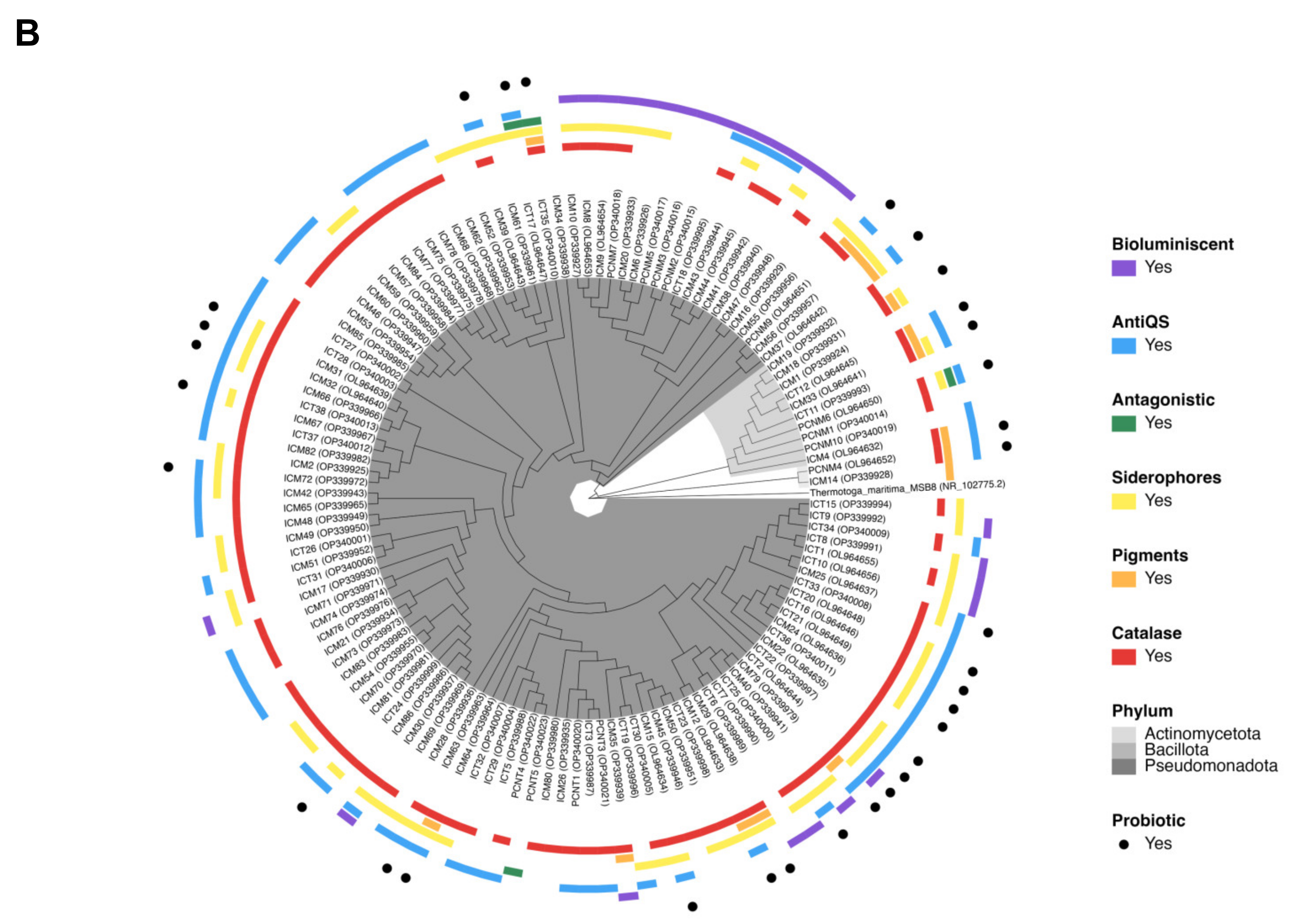
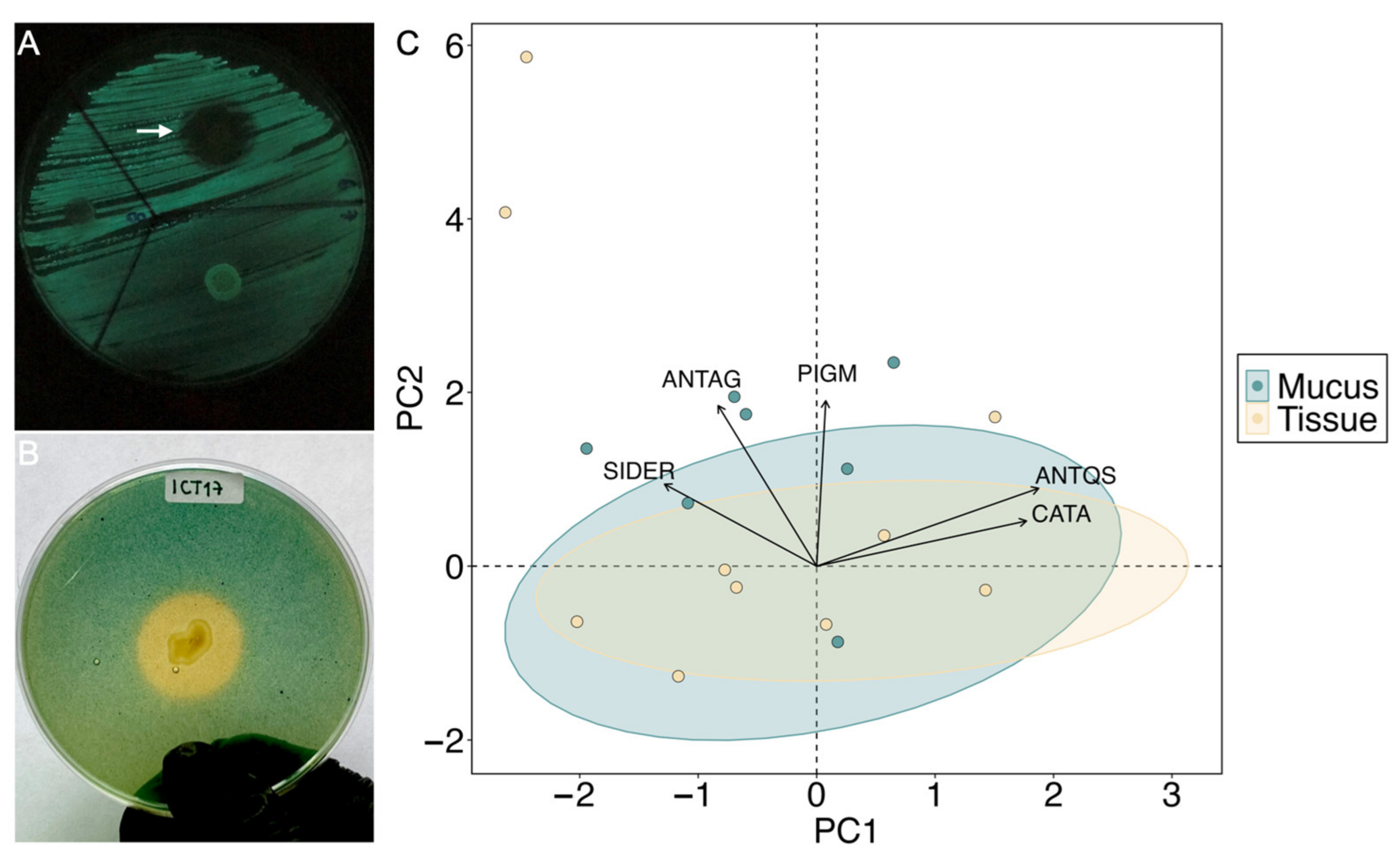
3.4. Probiotic Potential of M. auretenra-Associated Bacteria
4. Discussion
4.1. Status of Urban Madracis auretenra Coral and Culturable Bacteria
4.2. Presence of Potential Probiotic Traits in M. auretenra-Associated Bacteria

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probiotic Candidates (Genera) | Source of Isolation | Compartment and Condition | Location | Exhibited Probiotic Trait(s) | Tested Efficacy | References |
|---|---|---|---|---|---|---|
| Vibrio and Pseudoalteromonas | Various scleractinian corals | Healthy mucus | Gulf of Eilat, Israel | Anti-QS activity (Quorum sensing disruption of different biosensors) | No | [136] |
| Bacillus, Acinetobacter, Paracoccus, Pseudomonas, Vibrio, and Psychrobacter | Mussismilia harttii | Healthy tissue | Natural Park, Recife de Fora, Porto Seguro, Bahia, Brazil | Degradation of pollutants (oils) | Yes | [24] |
| Cobetia, Halomonas, and Pseudoalteromonas | Aquaria-maintained Pocillopora damicornis | Healthy tissue and surrounding water | Rio de Janeiro, Brazil | Catalase, antagonism against V. coralliilyticus and nifH and nirK | Yes | [25] |
| Bacillus, Pseudomonas, Photobacterium, Acinetobacter, Vibrio, and Enterobacter | Exposed low-tide Porites spp., and Turbinaria | Healthy and diseased mucus | Poshitra Natural Park, GoK, Jamnagar, Gujarat, India | Siderophore production | No | [22] |
| Shewanella, Cobetia, Halomonas, and Pseudoalteromonas | Oil-influenced Siderastrea stellata | Healthy tissue | Armação dos Búzios, Rio de Janeiro, Brazil | Degradation of pollutants (oils) | Yes | [7] |
| Bacillus, Planococcus, Salinivibrio, and Brachybacterium | Thermally resistant Mussismilia hispida | Healthy tissue | Maraú, Bahia, Brazil | Catalase, antagonism against V. coralliilyticus and V. alginolyticus, and nifH and nirK | Yes | [26] |
| Yangia, Roseobacter, Phytobacter, and Salinicola | Various scleractinian corals | Healthy tissue | Gaven Reef, Fiery Cross Reef, Johnson South Reef, South China Sea | Nitrogen-cycling genes nifH | Yes | [27] |
| Vibrio, Shewanella, Exiguobacterium, Bacillus, Fictibacillus, and Priestia | Urban Madracis auretenra | Healthy mucus and tissue | Inca Inca, Santa Marta, Colombia | Catalase, pigment, antagonism and anti-QS, and siderophore production | Not yet | Present study |
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Uthicke, S.; Ebert, T.; Liddy, M.; Johansson, C.; Fabricius, K.E.; Lamare, M. Echinometra Sea Urchins Acclimatized to Elevated PCO2 at Volcanic Vents Outperform Those under Present-Day PCO2 Conditions. Glob. Chang. Biol. 2016, 22, 2451–2461. [Google Scholar] [CrossRef]
- Wenger, A.S.; Williamson, D.H.; da Silva, E.T.; Ceccarelli, D.M.; Browne, N.K.; Petus, C.; Devlin, M.J. Effects of Reduced Water Quality on Coral Reefs in and out of No-Take Marine Reserves. Conserv. Biol. 2016, 30, 142–153. [Google Scholar] [CrossRef]
- Hughes, T.P.; Anderson, K.D.; Connolly, S.R.; Heron, S.F.; Kerry, J.T.; Lough, J.M.; Baird, A.H.; Baum, J.K.; Berumen, M.L.; Bridge, T.C.; et al. Spatial and Temporal Patterns of Mass Bleaching of Corals in the Anthropocene. Science 2018, 359, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.I.; Jamil, T.; Anlauf, H.; Coker, D.J.; Curdia, J.; Hewitt, J.; Jones, B.H.; Krokos, G.; Kürten, B.; Hariprasad, D.; et al. Multiple Stressor Effects on Coral Reef Ecosystems. Glob. Chang. Biol. 2019, 25, 4131–4146. [Google Scholar] [CrossRef]
- National Academies of Sciences, Engineering, and Medicine. A Research Review of Interventions to Increase the Persistence and Resilience of Coral Reefs; The National Academies Press: Washington, DC, USA, 2019; ISBN 978-0-309-48535-7. [Google Scholar]
- Villela, H.D.M.; Vilela, C.L.S.; Assis, J.M.; Varona, N.; Burke, C.; Coil, D.A.; Eisen, J.A.; Peixoto, R.S. Prospecting Microbial Strains for Bioremediation and Probiotics Development for Metaorganism Research and Preservation. J. Vis. Exp. 2019, 152, e60238. [Google Scholar] [CrossRef]
- Silva, D.P.; Villela, H.D.M.; Santos, H.F.; Duarte, G.A.S.; Ribeiro, J.R.; Ghizelini, A.M.; Vilela, C.L.S.; Rosado, P.M.; Fazolato, C.S.; Santoro, E.P.; et al. Multi-Domain Probiotic Consortium as an Alternative to Chemical Remediation of Oil Spills at Coral Reefs and Adjacent Sites. Microbiome 2021, 9, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, R.S.; Sweet, M.; Villela, H.D.M.; Cardoso, P.; Thomas, T.; Voolstra, C.R.; Høj, L.; Bourne, D.G. Coral Probiotics: Premise, Promise, Prospects. Annu. Rev. Anim. Biosci. 2021, 9, 265–288. [Google Scholar] [CrossRef] [PubMed]
- Reshef, L.; Koren, O.; Loya, Y.; Zilber-Rosenberg, L.; Rosenberg, E. The Coral Probiotic Hypothesis. Environ. Microbiol. 2006, 8, 2068–2073. [Google Scholar] [CrossRef]
- Ziegler, M.; Grupstra, C.G.B.; Barreto, M.M.; Eaton, M.; BaOmar, J.; Zubier, K.; Al-Sofyani, A.; Turki, A.J.; Ormond, R.; Voolstra, C.R. Coral Bacterial Community Structure Responds to Environmental Change in a Host-Specific Manner. Nat. Commun. 2019, 10, 4921. [Google Scholar] [CrossRef]
- Roitman, S.; López-Londoño, T.; Pollock, J.F.; Ritchie, K.B.; Galindo-Martínez, C.T.; Gómez-Campo, K.; González-Guerrero, L.A.; Pizarro, V.; López-Victoria, M.; Iglesias-Prieto, R.; et al. Surviving Marginalized Reefs: Assessing the Implications of the Microbiome on Coral Physiology and Survivorship. Coral Reefs 2020, 39, 795–807. [Google Scholar] [CrossRef]
- Deignan, L.K.; McDougald, D. Differential Response of the Microbiome of Pocillopora Acuta to Reciprocal Transplantation within Singapore. Microb. Ecol. 2022, 83, 608–618. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, M.; Seneca, F.O.; Yum, L.K.; Palumbi, S.R.; Voolstra, C.R. Bacterial Community Dynamics Are Linked to Patterns of Coral Heat Tolerance. Nat. Commun. 2017, 8, 14213. [Google Scholar] [CrossRef] [PubMed]
- Savary, R.; Barshis, D.J.; Voolstra, C.R.; Cárdenas, A.; Evensen, N.R.; Banc-Prandi, G.; Fine, M.; Meibom, A. Fast and Pervasive Transcriptomic Resilience and Acclimation of Extremely Heat-Tolerant Coral Holobionts from the Northern Red Sea. Proc. Natl. Acad. Sci. USA 2021, 118, e2023298118. [Google Scholar] [CrossRef]
- Maire, J.; van Oppen, M.J.H. A Role for Bacterial Experimental Evolution in Coral Bleaching Mitigation? Trends Microbiol. 2022, 30, 217–228. [Google Scholar] [CrossRef]
- Bourne, D.G.; Morrow, K.M.; Webster, N.S. Insights into the Coral Microbiome: Underpinning the Health and Resilience of Reef Ecosystems. Annu. Rev. Microbiol. 2016, 70, 317–340. [Google Scholar] [CrossRef]
- Thatcher, C.; Høj, L.; Bourne, D.G. Probiotics for Coral Aquaculture: Challenges and Considerations. Curr. Opin. Biotechnol. 2022, 73, 380–386. [Google Scholar] [CrossRef]
- Ravindran, J.; Kannapiran, E.; Manikandan, B.; Francis, K.; Arora, S.; Karunya, E.; Kumar, A.; Singh, S.K.; Jose, J. UV-Absorbing Bacteria in Coral Mucus and Their Response to Simulated Temperature Elevations. Coral Reefs 2013, 32, 1043–1050. [Google Scholar] [CrossRef]
- Dungan, A.M.; Bulach, D.; Lin, H.; Van Oppen, M.J.H.; Blackall, L.L. Development of a Free Radical Scavenging Probiotic to Mitigate Coral Bleaching. bioRxiv 2020, 1, 1–42. [Google Scholar] [CrossRef]
- Raina, J.B.; Tapiolas, D.; Motti, C.A.; Foret, S.; Seemann, T.; Tebben, J.; Willis, B.L.; Bourne, D.G. Isolation of an Antimicrobial Compound Produced by Bacteria Associated with Reef-Building Corals. PeerJ 2016, 4, e2275. [Google Scholar] [CrossRef]
- Ma, Z.P.; Song, Y.; Cai, Z.H.; Lin, Z.J.; Lin, G.H.; Wang, Y.; Zhou, J. Anti-Quorum Sensing Activities of Selected Coral Symbiotic Bacterial Extracts from the South China Sea. Front. Cell Infect. Microbiol. 2018, 8, 144. [Google Scholar] [CrossRef]
- Patel, N.P.; Shimpi, G.G.; Haldar, S. Evaluation of Heterotrophic Bacteria Associated with Healthy and Bleached Corals of Gulf of Kutch, Gujarat, India for Siderophore Production and Their Response to Climate Change Factors. Ecol. Indic. 2020, 113, 106219. [Google Scholar] [CrossRef]
- Peixoto, R.S.; Rosado, P.M.; Leite, D.C.A.; Rosado, A.S.; Bourne, D.G. Beneficial Microorganisms for Corals (BMC): Proposed Mechanisms for Coral Health and Resilience. Front. Microbiol. 2017, 8, 341. [Google Scholar] [CrossRef]
- Fragoso Ados Santos, H.; Duarte, G.A.S.; Rachid, C.T.D.C.; Chaloub, R.M.; Calderon, E.N.; Marangoni, L.F.D.B.; Bianchini, A.; Nudi, A.H.; Do Carmo, F.L.; Van Elsas, J.D.; et al. Impact of Oil Spills on Coral Reefs Can Be Reduced by Bioremediation Using Probiotic Microbiota. Sci. Rep. 2015, 5, 18268. [Google Scholar] [CrossRef] [PubMed]
- Rosado, P.M.; Leite, D.C.A.; Duarte, G.A.S.; Chaloub, R.M.; Jospin, G.; Nunes da Rocha, U.; Saraiva, J.P.; Dini-Andreote, F.; Eisen, J.A.; Bourne, D.G.; et al. Marine Probiotics: Increasing Coral Resistance to Bleaching through Microbiome Manipulation. ISME J. 2019, 13, 921–936. [Google Scholar] [CrossRef]
- Santoro, E.P.; Borges, R.M.; Espinoza, J.L.; Freire, M.; Messias, C.S.M.A.; Villela, H.D.M.; Pereira, L.M.; Vilela, C.L.S.; Rosado, J.G.; Cardoso, P.M.; et al. Coral Microbiome Manipulation Elicits Metabolic and Genetic Restructuring to Mitigate Heat Stress and Evade Mortality. Sci. Adv. 2021, 7, 3088–3101. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, Q.; Ling, J.; Long, L.; Huang, H.; Yin, J.; Wu, M.; Tang, X.; Lin, X.; Zhang, Y.; et al. Shifting the Microbiome of a Coral Holobiont and Improving Host Physiology by Inoculation with a Potentially Beneficial Bacterial Consortium. BMC Microbiol. 2021, 21, 130. [Google Scholar] [CrossRef]
- Camp, E.F.; Schoepf, V.; Suggett, D.J. How Can “Super Corals” Facilitate Global Coral Reef Survival under Rapid Environmental and Climatic Change? Glob. Change Biol. 2018, 24, 2755–2757. [Google Scholar] [CrossRef] [PubMed]
- Heery, E.C.; Hoeksema, B.W.; Browne, N.K.; Reimer, J.D.; Ang, P.O.; Huang, D.; Friess, D.A.; Chou, L.M.; Loke, L.H.L.; Saksena-Taylor, P.; et al. Urban Coral Reefs: Degradation and Resilience of Hard Coral Assemblages in Coastal Cities of East and Southeast Asia. Mar. Pollut. Bull. 2018, 135, 654–681. [Google Scholar] [CrossRef]
- Wainwright, B.J.; Afiq-Rosli, L.; Zahn, G.L.; Huang, D. Characterisation of Coral-Associated Bacterial Communities in an Urbanised Marine Environment Shows Strong Divergence over Small Geographic Scales. Coral Reefs 2019, 38, 1097–1106. [Google Scholar] [CrossRef]
- Röthig, T.; Ochsenkühn, M.A.; Roik, A.; Van Der Merwe, R.; Voolstra, C.R. Long-Term Salinity Tolerance Is Accompanied by Major Restructuring of the Coral Bacterial Microbiome. Mol. Ecol. 2016, 25, 1308–1323. [Google Scholar] [CrossRef]
- Brown, K.T.; Bender-Champ, D.; Hoegh-Guldberg, O.; Dove, S. Seasonal Shifts in the Competitive Ability of Macroalgae Influence the Outcomes of Coral-Algal Competition. R. Soc. Open Sci. 2020, 7, 201797. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Castillo, V.; Rodríguez-Troncoso, A.P.; Santiago-Valentín, J.D.; Cupul-Magaña, A.L. The Influence of Urban Pressures on Coral Physiology on Marginal Coral Reefs of the Mexican Pacific. Coral Reefs 2020, 39, 625–637. [Google Scholar] [CrossRef]
- Soares, M.O.; Rossi, S.; Gurgel, A.R.; Lucas, C.C.; Tavares, T.C.L.; Diniz, B.; Feitosa, C.V.; Rabelo, E.F.; Pereira, P.H.C.; de Kikuchi, R.K.P.; et al. Impacts of a Changing Environment on Marginal Coral Reefs in the Tropical Southwestern Atlantic. Ocean. Coast. Manag. 2021, 210, 105692. [Google Scholar] [CrossRef]
- Burt, J.A.; Camp, E.F.; Enochs, I.C.; Johansen, J.L.; Morgan, K.M.; Riegl, B.; Hoey, A.S. Insights from Extreme Coral Reefs in a Changing World. Coral Reefs 2020, 39, 495–507. [Google Scholar] [CrossRef]
- McDevitt-Irwin, J.M.; Baum, J.K.; Garren, M.; Vega Thurber, R.L. Responses of Coral-Associated Bacterial Communities to Local and Global Stressors. Front. Mar. Sci. 2017, 4, 262. [Google Scholar] [CrossRef]
- Ouédraogo, D.Y.; Delaunay, M.; Sordello, R.; Hédouin, L.; Castelin, M.; Perceval, O.; Domart-Coulon, I.; Burga, K.; Ferrier-Pagès, C.; Multon, R.; et al. Evidence on the Impacts of Chemicals Arising from Human Activity on Tropical Reef-Building Corals; a Systematic Map. Environ. Evid. 2021, 10, 22. [Google Scholar] [CrossRef]
- Hussein, E.I.; Juhmani, A.S.F.; Jacob, J.H.; Telfah, M.A.; Al-Razaq, M.A.A.; Al-Horani, F.A.; Al Zoubi, M.S.; Malkawi, H.I. Effect of Various Local Anthropogenic Impacts on the Diversity of Coral Mucus-Associated Bacterial Communities. J. Mar. Sci. Eng. 2022, 10, 863. [Google Scholar] [CrossRef]
- Winston, M.; Oliver, T.; Couch, C.; Donovan, M.K.; Asner, G.P.; Conklin, E.; Fuller, K.; Grady, B.W.; Huntington, B.; Kageyama, K.; et al. Coral Taxonomy and Local Stressors Drive Bleaching Prevalence across the Hawaiian Archipelago in 2019. PLoS ONE 2022, 17, e0269068. [Google Scholar] [CrossRef]
- Márquez, A. Estudio Sedimentológico y Morfológico de La Plataforma Continental Caribe Entre El Sector de Santa Marta y Punta Morro Hermoso. Boletín Cient. CIOH 1993, 1, 67–78. [Google Scholar] [CrossRef]
- Cámara de Comercio de Santa Marta. Informe Económico Anual de La Jurisdicción; Cámara de Comercio de Santa Marta: Magdalena, Colombia, 2022. [Google Scholar]
- Parques Nacionales Naturales de Colombia. Unidad Administrativa Especial del Sistema de Parques Naturales; Parques Nacionales Naturales de Colombia: Bogotá, Colombia, 2006. [Google Scholar]
- Franco-Herrera, A. Oceanografía de La Ensenada de Gaira; Universidad Jorge Tadeo Lozano: Bogotá, Colombia, 2005; ISBN 958-9029-72-8. [Google Scholar]
- Vega-Sequeda, J.; Polo-Silva, C.J.; Franco-Herrera, A.; Paramo-Granados, J.; Sanjuan-Muñoz, A. Dynamics of Physicochemical Variables of the Northern Colombian Caribbean Coastal Waters. DYNA 2019, 86, 17–25. [Google Scholar] [CrossRef]
- Ramos-Ortega, L.M.; Vidal, L.A.; Vilardy, S.; Saavedra-Díaz, L. Análisis de La Contaminación Microbiológica (Coliformes Totales y Fecales) En La Bahía de Santa Marta, Caribe Colombiano. Acta Biol. Colomb. 2008, 13, 87–98. [Google Scholar]
- García, F.; Palacio, C.; Garcia, U. Calidad Del Agua En El Área Costera de Santa Marta (Colombia). DYNA 2012, 79, 85–94. [Google Scholar]
- Vega-Sequeda, J.; Rodríguez-Ramírez, A.; Reyes-Nivia, M.C.; Navas-Camacho, R. Formaciones Coralinas Del Área de Santa Marta: Estado y Patrones de Distribución Espacial de La Comunidad Bentónica. Boletín Investig. Mar. Costeras 2008, 37, 87–105. [Google Scholar] [CrossRef]
- Locke, J.M.; Weil, E.; Coates, K.A. A Newly Documented Species of Madracis (Scleractinia: Pocilloporidae) from the Caribbean. Proc. Biol. Soc. Wash. 2007, 120, 214–226. [Google Scholar] [CrossRef]
- Invemar (Ed.) Corales Escleractinios de Colombia; No. 14; Invemar Serie de Publicaciones Especiales: Santa Marta, Colombia, 2010; Volume 14, ISBN 978-958-8448-21-3. [Google Scholar]
- Cárdenas, A.; Rodriguez, R.L.M.; Pizarro, V.; Cadavid, L.F.; Arévalo-Ferro, C. Shifts in Bacterial Communities of Two Caribbean Reef-Building Coral Species Affected by White Plague Disease. ISME J. 2012, 6, 502–512. [Google Scholar] [CrossRef]
- Brown, K.T.; Bender-Champ, D.; Bryant, D.E.P.; Dove, S.; Hoegh-Guldberg, O. Human Activities Influence Benthic Community Structure and the Composition of the Coral-Algal Interactions in the Central Maldives. J. Exp. Mar. Biol. Ecol. 2017, 497, 33–40. [Google Scholar] [CrossRef]
- Trygonis, V.; Sini, M. PhotoQuad: A Dedicated Seabed Image Processing Software, and a Comparative Error Analysis of Four Photoquadrat Methods. J. Exp. Mar. Biol. Ecol. 2012, 424, 99–108. [Google Scholar] [CrossRef]
- Barott, K.; Smith, J.; Dinsdale, E.; Hatay, M.; Sandin, S.; Rohwer, F. Hyperspectral and Physiological Analyses of Coral-Algal Interactions. PLoS ONE 2009, 4, e8043. [Google Scholar] [CrossRef]
- Cuthbert, M.W.; Smith, D.W.; Arróliga, O.; Mendoza, J.; Torres, N.; Flores, O. Indice de Salud Arrecifal (ISA) En Los Arrecifes Coralinos de Cayos Miskitos. Rev. Univ. Del. Caribe 2019, 22, 16–23. [Google Scholar] [CrossRef]
- Rodríguez-Grimón, R.; Campos, N.H.; Castro, Í.B. Effect of Maritime Traffic on Water Quality Parameters in Santa Marta, Colombia. J. Mar. Sci. Eng. 2021, 9, 474. [Google Scholar] [CrossRef]
- Strickland, J.D.H.; Parsons, T.R. A Practical Handbook of Seawater Analysis; Fisheries Research Board of Canada: Ottawa, ON, Canada, 1972; Volume 167. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutation Tests for Univariate or Multivariate Analysis of Variance and Regression. Can. J. Fish. Aquat. Sci. 2001, 58, 626–639. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Getting Started with PRIMER v7. PRIMER-E; Plymouth Marine Laboratory: Plymouth, UK, 2015; pp. 1–20. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier Science: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Stevens, M.H.H.; Oksanen, M.J.; Suggests, M.A.S.S. The Vegan Package; 2008; Volume 10. Available online: https://www.researchgate.net/profile/Gavin-Simpson-2/publication/228339454_The_vegan_Package/links/0912f50be86bc29a7f000000/The-vegan-Package.pdf (accessed on 26 September 2023).
- Cooksey, R.W. Illustrating Statistical Procedures: Finding Meaning in Quantitative Data; Springer: Berlin/Heidelberg, Germany, 2020; ISBN 978-981-15-2537-7. [Google Scholar]
- Li, J.; Zou, Y.; Yang, J.; Li, Q.; Bourne, D.G.; Sweet, M.; Liu, C.; Guo, A.; Zhang, S. Cultured Bacteria Provide Insight into the Functional Potential of the Coral-Associated Microbiome. mSystems 2022, 7, e00327-22. [Google Scholar] [CrossRef] [PubMed]
- Morrow, K.M.; Pankey, M.S.; Lesser, M.P. Community Structure of Coral Microbiomes Is Dependent on Host Morphology. Microbiome 2022, 10, 113. [Google Scholar] [CrossRef] [PubMed]
- Reiner, K. Catalase Test Protocol; American Society for Microbiology: Washington, DC, USA, 2010; pp. 1–6. [Google Scholar]
- Ratnakaran, P.; Bhoir, M.; Durve-Gupta, A. Isolation and Characterization of Pigment Producing Bacteria Isolated from Waste. Int. J. Appl. Res. 2020, 6, 252–260. [Google Scholar]
- Velho-Pereira, S.; Kamat, N.M. Antimicrobial Screening of Actinobacteria Using a Modified Cross-Streak Method. Indian. J. Pharm. Sci. 2011, 73, 223–228. [Google Scholar] [CrossRef]
- Cesa-Luna, C.; Baez, A.; Aguayo-Acosta, A.; Llano-Villarreal, R.C.; Juárez-González, V.R.; Gaytán, P.; del Rocío Bustillos-Cristales, M.; Rivera-Urbalejo, A.; Muñoz-Rojas, J.; Quintero-Hernández, V. Growth Inhibition of Pathogenic Microorganisms by Pseudomonas Protegens EMM-1 and Partial Characterization of Inhibitory Substances. PLoS ONE 2020, 15, e0240545. [Google Scholar] [CrossRef]
- Brodl, E.; Winkler, A.; Macheroux, P. Molecular Mechanisms of Bacterial Bioluminescence. Comput. Struct. Biotechnol. J. 2018, 16, 551–564. [Google Scholar] [CrossRef]
- Dunlap, P. Biochemistry and Genetics of Bacterial Bioluminescence. Adv. Biochem. Eng. Biotechnol. 2014, 144, 37–64. [Google Scholar] [CrossRef]
- Rajivgandhi, G.; Vijayan, R.; Maruthupandy, M.; Vaseeharan, B.; Manoharan, N. Antibiofilm Effect of Nocardiopsis sp. GRG 1 (KT235640) Compound against Biofilm Forming Gram Negative Bacteria on UTIs. Microb. Pathog. 2018, 118, 190–198. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal Chemical Assay for the Detection and Determination of Siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Pérez-Miranda, S.; Cabirol, N.; George-Téllez, R.; Zamudio-Rivera, L.S.; Fernández, F.J. O-CAS, a Fast and Universal Method for Siderophore Detection. J. Microbiol. Methods 2007, 70, 127–131. [Google Scholar] [CrossRef]
- Frank, J.A.; Reich, C.I.; Sharma, S.; Weisbaum, J.S.; Wilson, B.A.; Olsen, G.J. Critical Evaluation of Two Primers Commonly Used for Amplification of Bacterial 16S RRNA Genes. Appl. Environ. Microbiol. 2008, 74, 2461–2470. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Yarza, P.; Yilmaz, P.; Pruesse, E.; Glöckner, F.O.; Ludwig, W.; Schleifer, K.; Whitman, W.; Euzéby, J.; Amann, R.; Rosselló-Móra, R. Uniting the Classification of Cultured and Uncultured Bacteria and Archaea Using 16S RRNA Gene Sequences. Nat. Rev. Microbiol. 2014, 12, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A Taxonomically United Database of 16S RRNA Gene Sequences and Whole-Genome Assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Sweet, M.; Villela, H.; Keller-Costa, T.; Costa, R.; Romano, S.; Bourne, D.G.; Cárdenas, A.; Huggett, M.J.; Kerwin, A.H.; Kuek, F.; et al. Insights into the Cultured Bacterial Fraction of Corals. mSystems 2021, 6, e01249-20. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v3: An Online Tool for the Display and Annotation of Phylogenetic and Other Trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef]
- Comba González, N.; Ramírez Hoyos, M.L.; López Kleine, L.; Montoya Castaño, D. Production of Enzymes and Siderophores by Epiphytic Bacteria Isolated from the Marine Macroalga Ulva Lactuca. Aquat. Biol. 2018, 27, 107–118. [Google Scholar] [CrossRef]
- Munn, C.B. The Role of Vibrios in Diseases of Corals. Microbiol. Spectr. 2015, 3, 3–4. [Google Scholar] [CrossRef]
- Schaffelke, B.; Klumpp, D.W. Nutrient-Limited Growth of the Coral Reef Macroalga Sargassum Baccularia and Experimental Growth Enhancement by Nutrient Addition in Continuous Flow Culture. Mar. Ecol. Prog. Ser. 1998, 164, 199–211. [Google Scholar] [CrossRef]
- D’Angelo, C.; Wiedenmann, J. Impacts of Nutrient Enrichment on Coral Reefs: New Perspectives and Implications for Coastal Management and Reef Survival. Curr. Opin. Environ. Sustain. 2014, 7, 82–93. [Google Scholar] [CrossRef]
- Hayashida, H.; Matear, R.J.; Strutton, P. Background Nutrient Concentration Determines Phytoplankton Bloom Response to Marine Heatwaves. Glob. Change Biol. 2020, 26, 4800–4811. [Google Scholar] [CrossRef]
- Nalley, E.M.; Tuttle, L.J.; Conklin, E.E.; Barkman, A.L.; Wulstein, D.M.; Schmidbauer, M.C.; Donahue, M.J. A Systematic Review and Meta-Analysis of the Direct Effects of Nutrients on Corals. Sci. Total Environ. 2023, 856, 159093. [Google Scholar] [CrossRef] [PubMed]
- Lipp, E.; Griffin, D.W. Analysis of Coral Mucus as an Improved Medium for Detection of Enteric Microbes and for Determining Patterns of Sewage Contamination in Reef Environments. Ecohealth 2004, 1, 317–323. [Google Scholar] [CrossRef]
- Beurmann, S.; Ushijima, B.; Videau, P.; Svoboda, C.M.; Chatterjee, A.; Aeby, G.S.; Callahan, S.M. Dynamics of Acute Montipora White Syndrome: Bacterial Communities of Healthy and Diseased M. Capitata Colonies during and after a Disease Outbreak. Microbiology 2018, 164, 1240–1253. [Google Scholar] [CrossRef]
- Buccheri, M.A.; Salvo, E.; Coci, M.; Quero, G.M.; Zoccarato, L.; Privitera, V.; Rappazzo, G. Investigating Microbial Indicators of Anthropogenic Marine Pollution by 16S and 18S High-Throughput Sequencing (HTS) Library Analysis. FEMS Microbiol. Lett. 2019, 366, fnz179. [Google Scholar] [CrossRef]
- Numberger, D.; Ganzert, L.; Zoccarato, L.; Mühldorfer, K.; Sauer, S.; Grossart, H.P.; Greenwood, A.D. Characterization of Bacterial Communities in Wastewater with Enhanced Taxonomic Resolution by Full-Length 16S RRNA Sequencing. Sci. Rep. 2019, 9, 9673. [Google Scholar] [CrossRef]
- Orel, N.; Fadeev, E.; Klun, K.; Ličer, M.; Tinta, T.; Turk, V. Bacterial Indicators Are Ubiquitous Members of Pelagic Microbiome in Anthropogenically Impacted Coastal Ecosystem. Front. Microbiol. 2022, 12, 765091. [Google Scholar] [CrossRef]
- Ritchie, K. Regulation of Microbial Populations by Coral Surface Mucus and Mucus-Associated Bacteria. Mar. Ecol. Prog. Ser. 2006, 332, 1–14. [Google Scholar] [CrossRef]
- Narváez, S.; Gómez, M.; Acosta, J. Coliformes Termotolerantes En Aguas de Las Poblaciones Costeras y Palafíticas de La Ciénaga Grande de Santa Marta, Colombia. Acta Biol. Colomb. 2008, 13, 113–122. [Google Scholar]
- INVEMAR Análisis Histórico 2001–2021 de La Calidad de Las Aguas Marinas y Costeras En El Caribe y Pacífico Colombianos. Red de Vigilancia para la Conservación y Protección de las Aguas Marinas y Costeras de Colombia; Serie de Publicaciones Periódicas No. 4 del INVEMAR. Invemar: Santa Marta, Colombia, 2022; p. 208. Available online: https://www.researchgate.net/publication/362157282_Analisis_historico_2001-2021_de_la_calidad_de_las_aguas_marinas_y_costeras_en_el_Caribe_y_Pacifico_Colombianos_Informe_REDCAM_2021 (accessed on 26 September 2023).
- INVEMAR Diagnóstico y Evaluación de La Calidad de Las Aguas Marinas y Costeras En El Caribe y Pacífico Colombianos. In Red. de Vigilancia para la Conservación y Protección de las Aguas Marinas y Costeras de Colombia; Serie de Publicaciones Periódicas No. 4 del INVEMAR; Invemar: Santa Marta, Colombia, 2020; p. 171. Available online: https://www.invemar.org.co/documents/10182/14479/Informe+REDCAM+2016.pdf/b21d50f5-cd2d-4926-a016-b321cc9659e7 (accessed on 26 September 2023).
- Stewart, J.R.; Gast, R.J.; Fujioka, R.S.; Solo-Gabriele, H.M.; Meschke, J.S.; Amaral-Zettler, L.A.; Del Castillo, E.; Polz, M.F.; Collier, T.K.; Strom, M.S.; et al. The Coastal Environment and Human Health: Microbial Indicators, Pathogens, Sentinels and Reservoirs. Environ. Health 2008, 7, S3. [Google Scholar] [CrossRef] [PubMed]
- Kraus, R.; Baljak, V.; Vukić Lušić, D.; Kranjčević, L.; Cenov, A.; Glad, M.; Kauzlarić, V.; Lušić, D.; Grbčić, L.; Alvir, M.; et al. Impacts of Atmospheric and Anthropogenic Factors on Microbiological Pollution of the Recreational Coastal Beaches Neighboring Shipping Ports. Int. J. Environ. Res. Public Health 2022, 19, 8552. [Google Scholar] [CrossRef] [PubMed]
- Fudjaja, L.; Viantika, N.M.; Rani, C.; Nurdin, N.; Priosambodo, D.; Tenriawaru, A.N. Anthropogenic Activity and the Destruction of Coral Reefs in the Waters of Small Islands. IOP Conf. Ser. Earth Environ. Sci. 2020, 575, 012057. [Google Scholar] [CrossRef]
- Tout, J.; Siboni, N.; Messer, L.F.; Garren, M.; Stocker, R.; Webster, N.S.; Ralph, P.J.; Seymour, J.R. Increased Seawater Temperature Increases the Abundance and Alters the Structure of Natural Vibrio Populations Associated with the Coral Pocillopora Damicornis. Front. Microbiol. 2015, 6, 432. [Google Scholar] [CrossRef]
- Leite, D.C.A.; Salles, J.F.; Calderon, E.N.; Castro, C.B.; Bianchini, A.; Marques, J.A.; van Elsas, J.D.; Peixoto, R.S. Coral Bacterial-Core Abundance and Network Complexity as Proxies for Anthropogenic Pollution. Front. Microbiol. 2018, 9, 833. [Google Scholar] [CrossRef]
- Dunphy, C.M.; Gouhier, T.C.; Chu, N.D.; Vollmer, S.V. Structure and Stability of the Coral Microbiome in Space and Time. Sci. Rep. 2019, 9, 6785. [Google Scholar] [CrossRef]
- Rohwer, F.; Seguritan, V.; Azam, F.; Knowlton, N. Diversity and Distribution of Coral-Associated Bacteria. Mar. Ecol. Prog. Ser. 2002, 243, 1–10. [Google Scholar] [CrossRef]
- Littman, R.A.; Willis, B.L.; Pfeffer, C.; Bourne, D.G. Diversities of Coral-Associated Bacteria Differ with Location, but Not Species, for Three Acroporid Corals on the Great Barrier Reef. FEMS Microbiol. Ecol. 2009, 68, 152–163. [Google Scholar] [CrossRef]
- Hong, M.J.; Yu, Y.T.; Chen, C.A.; Chiang, P.W.; Tang, S.L. Influence of Species Specificity and Other Factors on Bacteria Associated with the Coral Stylophora Pistillata in Taiwan. Appl. Environ. Microbiol. 2009, 75, 7797–7806. [Google Scholar] [CrossRef]
- Sunagawa, S.; Woodley, C.M.; Medina, M. Threatened Corals Provide Underexplored Microbial Habitats. PLoS ONE 2010, 5, e9554. [Google Scholar] [CrossRef] [PubMed]
- Blackall, L.L.; Wilson, B.; Van Oppen, M.J.H. Coral-the World’s Most Diverse Symbiotic Ecosystem. Mol. Ecol. 2015, 24, 5330–5347. [Google Scholar] [CrossRef] [PubMed]
- Bourne, D.G.; Munn, C.B. Diversity of Bacteria Associated with the Coral Pocillopora Damicornis from the Great Barrier Reef. Environ. Microbiol. 2005, 7, 1162–1174. [Google Scholar] [CrossRef] [PubMed]
- Pollock, F.J.; McMinds, R.; Smith, S.; Bourne, D.G.; Willis, B.L.; Medina, M.; Thurber, R.V.; Zaneveld, J.R. Coral-Associated Bacteria Demonstrate Phylosymbiosis and Cophylogeny. Nat. Commun. 2018, 9, 4921. [Google Scholar] [CrossRef]
- Hernandez-Agreda, A.; Gates, R.D.; Ainsworth, T.D. Defining the Core Microbiome in Corals’ Microbial Soup. Trends Microbiol. 2017, 25, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Agreda, A.; Leggat, W.; Bongaerts, P.; Herrera, C.; Ainsworth, T.D. Rethinking the Coral Microbiome: Simplicity Exists within a Diverse Microbial Biosphere. mBio 2018, 9, e00812-18. [Google Scholar] [CrossRef] [PubMed]
- Lampert, Y.; Kelman, D.; Dubinsky, Z.; Nitzan, Y.; Hill, R.T. Diversity of Culturable Bacteria in the Mucus of the Red Sea Coral Fungia Scutaria. FEMS Microbiol. Ecol. 2006, 58, 99–108. [Google Scholar] [CrossRef]
- van Oppen, M.J.H.; Blackall, L.L. Coral Microbiome Dynamics, Functions and Design in a Changing World. Nat. Rev. Microbiol. 2019, 17, 557–567. [Google Scholar] [CrossRef]
- Arboleda, M.; Reichardt, W. Epizoic Communities of Prokaryotes on Healthy and Diseased Scleractinian Corals in Lingayen Gulf, Philippines. Microb. Ecol. 2009, 57, 117–128. [Google Scholar] [CrossRef]
- Koenig, J.E.; Bourne, D.G.; Curtis, B.; Dlutek, M.; Stokes, H.W.; Doolittle, W.F.; Boucher, Y. Coral-Mucus-Associated Vibrio Integrons in the Great Barrier Reef: Genomic Hotspots for Environmental Adaptation. ISME J. 2011, 5, 962–972. [Google Scholar] [CrossRef]
- Kvennefors, E.C.E.; Sampayo, E.; Ridgway, T.; Barnes, A.C.; Hoegh-Guldberg, O. Bacterial Communities of Two Ubiquitous Great Barrier Reef Corals Reveals Both Site-and Species-Specificity of Common Bacterial Associates. PLoS ONE 2010, 5, e10401. [Google Scholar] [CrossRef] [PubMed]
- Solano, F. Melanin and Melanin-Related Polymers as Materials with Biomedical and Biotechnological Applications—Cuttlefish Ink and Mussel Foot Proteins as Inspired Biomolecules. Int. J. Mol. Sci. 2017, 18, 1561. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Lin, Z.J.; Cai, Z.H.; Zeng, Y.H.; Zhu, J.M.; Du, X.P. Opportunistic Bacteria Use Quorum Sensing to Disturb Coral Symbiotic Communities and Mediate the Occurrence of Coral Bleaching. Environ. Microbiol. 2020, 22, 1944–1962. [Google Scholar] [CrossRef] [PubMed]
- Anithajothi, R.; Duraikannu, K.; Umagowsalya, G.; Ramakritinan, C.M. The Presence of Biomarker Enzymes of Selected Scleractinian Corals of Palk Bay, Southeast Coast of India. Biomed. Res. Int. 2014, 2014, 684874. [Google Scholar] [CrossRef]
- Zimmer, B.L.; May, A.L.; Bhedi, C.D.; Dearth, S.P.; Prevatte, C.W.; Pratte, Z.; Campagna, S.R.; Richardson, L.L. Quorum Sensing Signal Production and Microbial Interactions in a Polymicrobial Disease of Corals and the Coral Surface Mucopolysaccharide Layer. PLoS ONE 2014, 9, e108541. [Google Scholar] [CrossRef]
- Zhang, T.; Diaz, J.M.; Brighi, C.; Parsons, R.J.; McNally, S.; Apprill, A.; Hansel, C.M. Dark Production of Extracellular Superoxide by the Coral Porites Astreoides and Representative Symbionts. Front. Mar. Sci. 2016, 3, 232. [Google Scholar] [CrossRef]
- Hansel, C.M.; Diaz, J.M. Production of Extracellular Reactive Oxygen Species by Marine Biota. Ann. Rev. Mar. Sci. 2021, 13, 177–200. [Google Scholar] [CrossRef]
- Nielsen, D.A.; Petrou, K.; Gates, R.D. Coral Bleaching from a Single Cell Perspective. ISME J. 2018, 12, 1558–1567. [Google Scholar] [CrossRef]
- Li, X.; Imlay, J.A. Improved Measurements of Scant Hydrogen Peroxide Enable Experiments That Define Its Threshold of Toxicity for Escherichia Coli. Free Radic. Biol. Med. 2018, 120, 217–227. [Google Scholar] [CrossRef]
- Wang, L.; Li, Y. Metal Ions Driven Production, Characterization and Bioactivity of Extracellular Melanin from Streptomyces Sp. ZL-24. Int. J. Biol. Macromol. 2019, 123, 521–530. [Google Scholar] [CrossRef]
- Scott, K.J. Detection and Measurement of Carotenoids by UV/VIS Spectrophotometry. Curr. Protoc. Food Anal. Chem. 2001, 1, F2.2.1–F2.2.10. [Google Scholar] [CrossRef]
- Sen Gupta, S.; Ghosh, M. In Vitro Antioxidative Evaluation of α- and β -Carotene, Isolated from Crude Palm Oil. J. Anal. Methods Chem. 2013, 2013, 351671. [Google Scholar] [CrossRef] [PubMed]
- Misawa, N.; Shimada, H. Metabolic Engineering for the Production of Carotenoids in Non-Carotenogenic Bacteria and Yeasts. J. Biotechnol. 1998, 59, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Dieser, M.; Greenwood, M.; Foreman, C.M. Carotenoid Pigmentation in Antarctic Heterotrophic Bacteria as a Strategy to Withstand Environmental Stresses. Arct. Antarct. Alp. Res. 2010, 42, 396–405. [Google Scholar] [CrossRef]
- Hillyer, K.E.; Dias, D.A.; Lutz, A.; Wilkinson, S.P.; Roessner, U.; Davy, S.K. Metabolite Profiling of Symbiont and Host during Thermal Stress and Bleaching in the Coral Acropora Aspera. Coral Reefs 2017, 36, 105–118. [Google Scholar] [CrossRef]
- Bana, H.Z. Isolation of Pigment-Producing Bacteria from Surface Water and Study Sun Protection Factor (SPF) of the Purified Pigments. Int. J. Adv. Sci. Eng. Technol. 2017, 5, 2321–9009. [Google Scholar]
- Reis-Mansur, M.C.P.P.; Cardoso-Rurr, J.S.; Silva, J.V.M.A.; de Souza, G.R.; Cardoso, V.S.; Mansoldo, F.R.P.; Pinheiro, Y.; Schultz, J.; Lopez Balottin, L.B.; da Silva, A.J.R.; et al. Carotenoids from UV-Resistant Antarctic Microbacterium Sp. LEMMJ01. Sci. Rep. 2019, 9, 9554. [Google Scholar] [CrossRef]
- Bythell, J.C.; Wild, C. Biology and Ecology of Coral Mucus Release. J. Exp. Mar. Biol. Ecol. 2011, 408, 88–93. [Google Scholar] [CrossRef]
- Sweet, M.; Ramsey, A.; Bulling, M. Designer Reefs and Coral Probiotics: Great Concepts but Are They Good Practice? Biodiversity 2017, 18, 19–22. [Google Scholar] [CrossRef][Green Version]
- Patel, N.P.; Kumar, S.B.; Haldar, S. Role of Bacteria in Coral Ecosystem. In Systems Biology of Marine Ecosystems; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 317–341. ISBN 9783319620947. [Google Scholar]
- Matthews, J.L.; Raina, J.B.; Kahlke, T.; Seymour, J.R.; van Oppen, M.J.H.; Suggett, D.J. Symbiodiniaceae-Bacteria Interactions: Rethinking Metabolite Exchange in Reef-Building Corals as Multi-Partner Metabolic Networks. Environ. Microbiol. 2020, 22, 1675–1687. [Google Scholar] [CrossRef]
- Höfte, M. Classes of Microbial Siderophores. In Iron Chelation in Plants and Soil Microorganisms; Academic Press: Cambridge, MA, USA, 1993; pp. 3–26. [Google Scholar]
- Golberg, K.; Pavlov, V.; Marks, R.S.; Kushmaro, A. Coral-Associated Bacteria, Quorum Sensing Disrupters, and the Regulation of Biofouling. Biofouling 2013, 29, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Golberg, K.; Eltzov, E.; Shnit-Orland, M.; Marks, R.S.; Kushmaro, A. Characterization of Quorum Sensing Signals in Coral-Associated Bacteria. Microb. Ecol. 2011, 61, 783–792. [Google Scholar] [CrossRef]
- Vega Thurber, R.; Mydlarz, L.D.; Brandt, M.; Harvell, D.; Weil, E.; Raymundo, L.; Willis, B.L.; Langevin, S.; Tracy, A.M.; Littman, R.; et al. Deciphering Coral Disease Dynamics: Integrating Host, Microbiome, and the Changing Environment. Front. Ecol. Evol. 2020, 8, 575927. [Google Scholar] [CrossRef]
- Liu, J.; Fu, K.; Wu, C.; Qin, K.; Li, F.; Zhou, L. “In-Group” Communication in Marine Vibrio: A Review of N-Acyl Homoserine Lactones-Driven Quorum Sensing. Front. Cell Infect. Microbiol. 2018, 8, 139. [Google Scholar] [CrossRef]
- Cervino, J.M.; Thompson, F.L.; Gomez-Gil, B.; Lorence, E.A.; Goreau, T.J.; Hayes, R.L.; Winiarski-Cervino, K.B.; Smith, G.W.; Hughen, K.; Bartels, E. The Vibrio Core Group Induces Yellow Band Disease in Caribbean and Indo-Pacific Reef-Building Corals. J. Appl. Microbiol. 2008, 105, 1658–1671. [Google Scholar] [CrossRef]
- Certner, R.H.; Vollmer, S.V. Inhibiting Bacterial Quorum Sensing Arrests Coral Disease Development and Disease-Associated Microbes. Environ. Microbiol. 2018, 20, 645–657. [Google Scholar] [CrossRef]
- Certner, R.H.; Vollmer, S.V. Evidence for Autoinduction and Quorum Sensing in White Band Disease-Causing Microbes on Acropora Cervicornis. Sci. Rep. 2015, 5, 11134. [Google Scholar] [CrossRef] [PubMed]
- Ringø, E.; Van Doan, H.; Lee, S.H.; Soltani, M.; Hoseinifar, S.H.; Harikrishnan, R.; Song, S.K. Probiotics, Lactic Acid Bacteria and Bacilli: Interesting Supplementation for Aquaculture. J. Appl. Microbiol. 2020, 129, 116–136. [Google Scholar] [CrossRef]
- Glasl, B.; Herndl, G.J.; Frade, P.R. The Microbiome of Coral Surface Mucus Has a Key Role in Mediating Holobiont Health and Survival upon Disturbance. ISME J. 2016, 10, 2280–2292. [Google Scholar] [CrossRef]
- Ruiz-Toquica, J.S.; Becerra-Real, L.M.; Díaz, L.M.V. Effect of Bacillus Firmus C101 on the Growth of Litopenaeus Vannamei Boone (White Shrimp) Post-Larvae, and Brachionus Plicatilis s.s. Müller (Rotifer). Bol. Investig. Mar. Costeras 2020, 49, 63–80. [Google Scholar] [CrossRef]
- Tao, X.; He, J.; Lu, J.; Chen, Z.; Jin, M.; Jiao, L.; Masagounder, K.; Liu, W.; Zhou, Q. Effects of Bacillus Subtilis DSM 32315 (Gutcare®) on the Growth Performance, Antioxidant Status, Immune Ability and Intestinal Function for Juvenile Litopenaeus Vannamei Fed with High/Low-Fishmeal Diets. Aquac. Rep. 2022, 26, 101282. [Google Scholar] [CrossRef]
- Chimetto, L.A.; Brocchi, M.; Thompson, C.C.; Martins, R.C.R.; Ramos, H.R.; Thompson, F.L. Vibrios Dominate as Culturable Nitrogen-Fixing Bacteria of the Brazilian Coral Mussismilia Hispida. Syst. Appl. Microbiol. 2008, 31, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Olson, N.D.; Ainsworth, T.D.; Gates, R.D.; Takabayashi, M. Diazotrophic Bacteria Associated with Hawaiian Montipora Corals: Diversity and Abundance in Correlation with Symbiotic Dinoflagellates. J. Exp. Mar. Biol. Ecol. 2009, 371, 140–146. [Google Scholar] [CrossRef]







| CHE Chengue (PNN Tayrona) | INC Inca Inca | |
|---|---|---|
| Geographic Location | 11°19′30″ N; 74°07′41.8″ W | 11°12′58.4″ N; 74°14′6.3″ W |
| Depth (m) | 9.67 ± 0.87 | 4.45 ± 0.49 |
| Temperature (°C) | 26.37 ± 0.09 | 26.30 ± 0.21 |
| Salinity | 36.20 ± 0.06 | 36.27 ± 0.09 |
| pH | 8.57 ± 0.21 | 8.37 ± 0.03 |
| Oxygen saturation (%) | 73.03 ± 3.47 | 69.90 ± 5.93 |
| Ammonia NH4+ (µg L−1) | 10.53 ± 0.53 | 12.27 ± 0.57 |
| Orthophosphate PO43− (µg L−1) | <2.00 | 3.07 ± 0.13 |
| Nitrate NO3− (µg L−1) | 4.98 ± 1.52 | 21.12 ± 0.53 |
| Nitrite NO2− (µg L−1) | 1.32 ± 0.21 | 1.54 ± 0.06 |
| TSS (mg L−1) | 3.62 ± 0.04 | 1.60 ± 0.11 |
| Isolate ID | Sequence Size (bp) | Genus | Similarity (%) | Access Code GenBank |
|---|---|---|---|---|
| ICM4 | 742 | Exiguobacterium | 99.33 | OL964632 |
| ICM12-15-22-24-31-32-63-64-66-67-72-79, ICT20-21-22-25-35 | 703–805 | Vibrio | 98.57–100 | OL964633-36, 39, 40, 48, 49, OP339963, 64, 66, 67, 72, 79, 97, OP340000, 10 |
| ICM18 | 796 | Priestia | 99.23 | OP339931 |
| ICM33, PCNM6 | 794–799 | Bacillus | 97.72–99.12 | OL964641, 50 |
| ICM37 | 788 | Shewanella | 99.49 | OL964642 |
| ICT12 | 799 | Fictibacillus | 98.74 | OL964645 |
| PCNM9 | 799 | Shewanella | 99.12 | OL964651 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-Toquica, J.S.; Yañez-Dukon, L.A.; Herrera Khenayzir, C.; Romero Borja, I.; Sanjuan-Muñoz, A.; Medina, M.; Franco-Herrera, A. Exploring the Status of an Urban Coral and the Presence of Potential Probiotic Traits in Culturable Bacteria. J. Mar. Sci. Eng. 2023, 11, 2006. https://doi.org/10.3390/jmse11102006
Ruiz-Toquica JS, Yañez-Dukon LA, Herrera Khenayzir C, Romero Borja I, Sanjuan-Muñoz A, Medina M, Franco-Herrera A. Exploring the Status of an Urban Coral and the Presence of Potential Probiotic Traits in Culturable Bacteria. Journal of Marine Science and Engineering. 2023; 11(10):2006. https://doi.org/10.3390/jmse11102006
Chicago/Turabian StyleRuiz-Toquica, Jordan Steven, Luis Alejandro Yañez-Dukon, Carolina Herrera Khenayzir, Isaac Romero Borja, Adolfo Sanjuan-Muñoz, Monica Medina, and Andres Franco-Herrera. 2023. "Exploring the Status of an Urban Coral and the Presence of Potential Probiotic Traits in Culturable Bacteria" Journal of Marine Science and Engineering 11, no. 10: 2006. https://doi.org/10.3390/jmse11102006
APA StyleRuiz-Toquica, J. S., Yañez-Dukon, L. A., Herrera Khenayzir, C., Romero Borja, I., Sanjuan-Muñoz, A., Medina, M., & Franco-Herrera, A. (2023). Exploring the Status of an Urban Coral and the Presence of Potential Probiotic Traits in Culturable Bacteria. Journal of Marine Science and Engineering, 11(10), 2006. https://doi.org/10.3390/jmse11102006