
Detection of Haplosporidium pinnae from Pinna nobilis Faeces

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Biological Materials
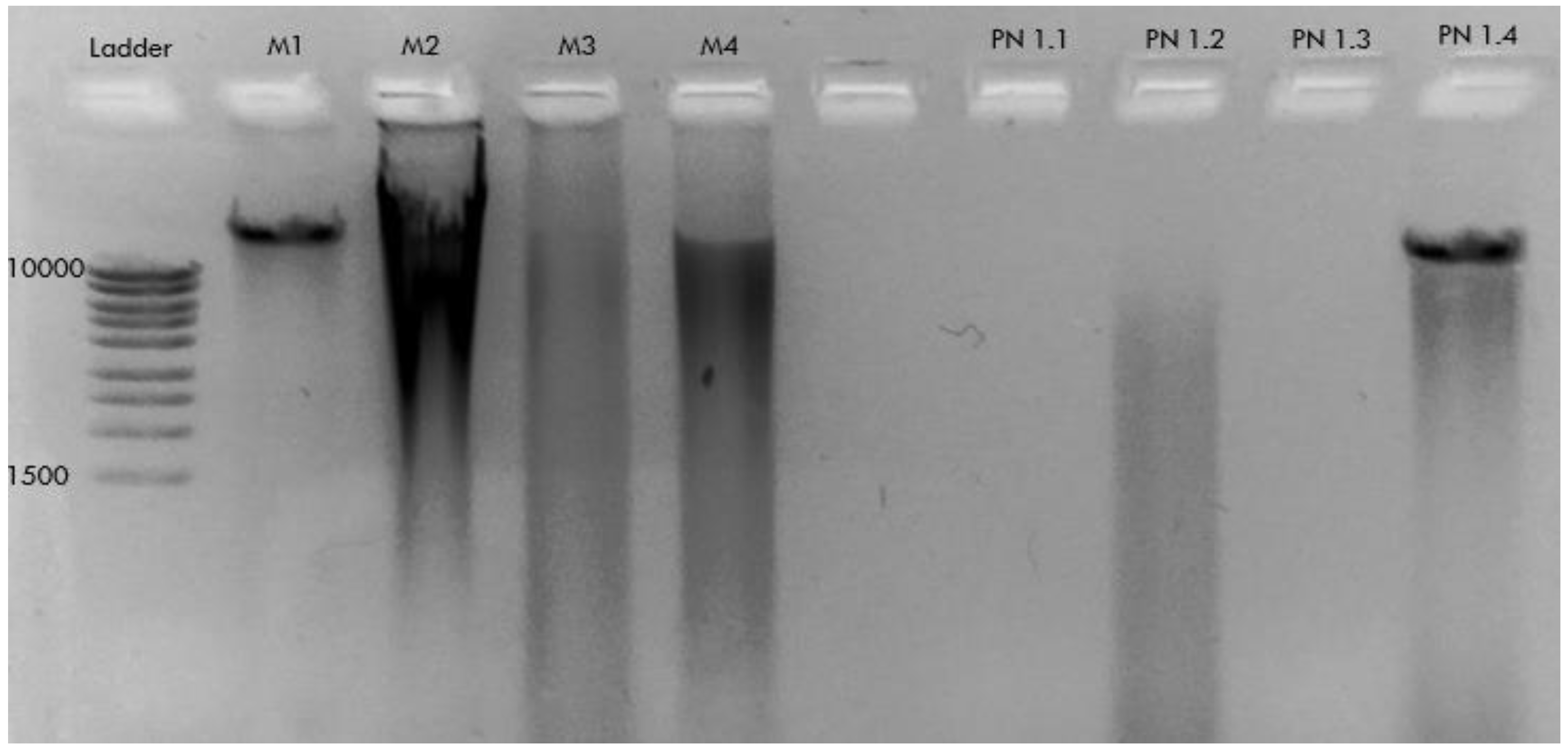
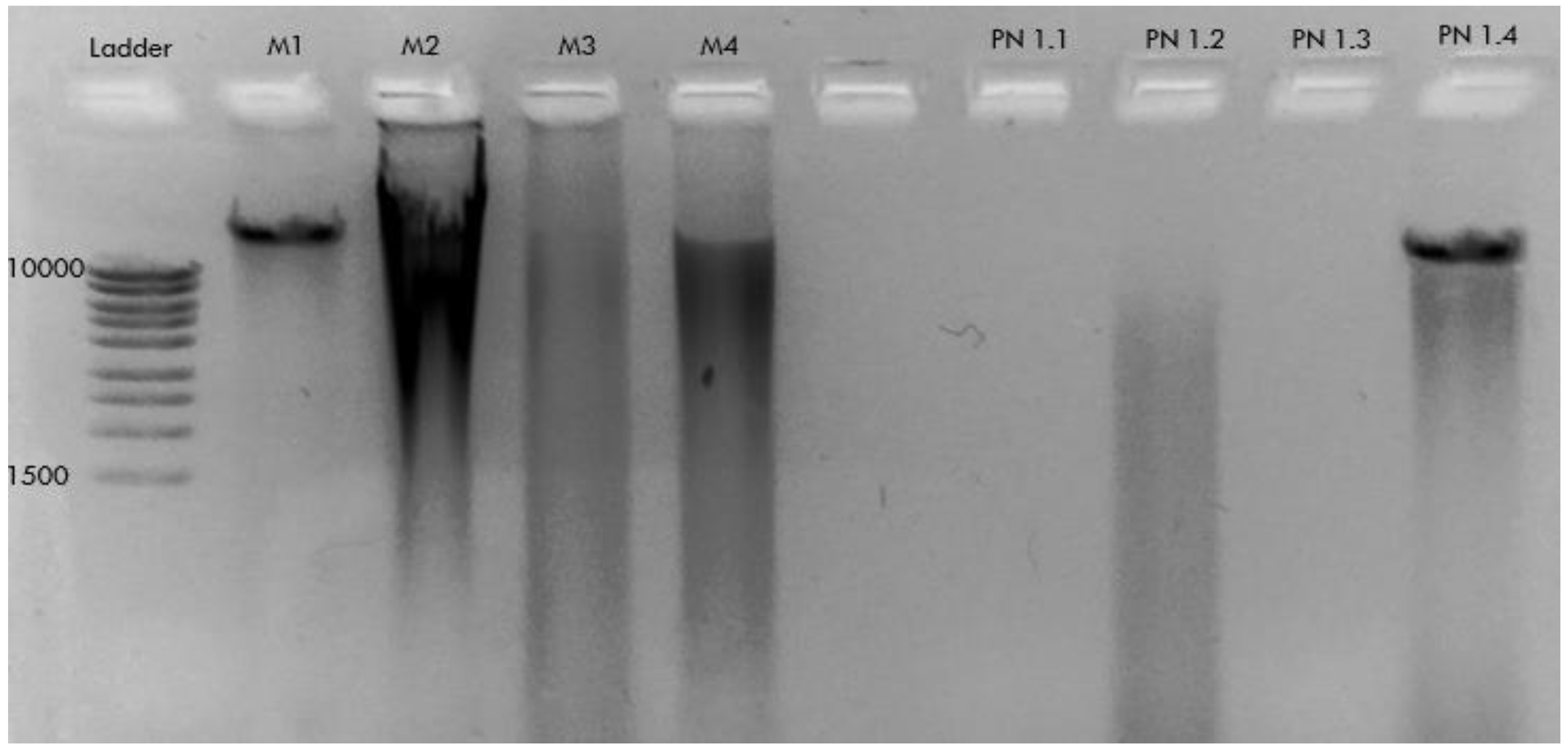
2.2. DNA Extraction
2.3. Gene Amplification and Sequencing
2.4. qPCR Quantification of Haplosporidium Pinnae in Pinna Nobilis
3. Results
3.1. PCR Detection of Haplosporidium sp. in Pinna nobilis
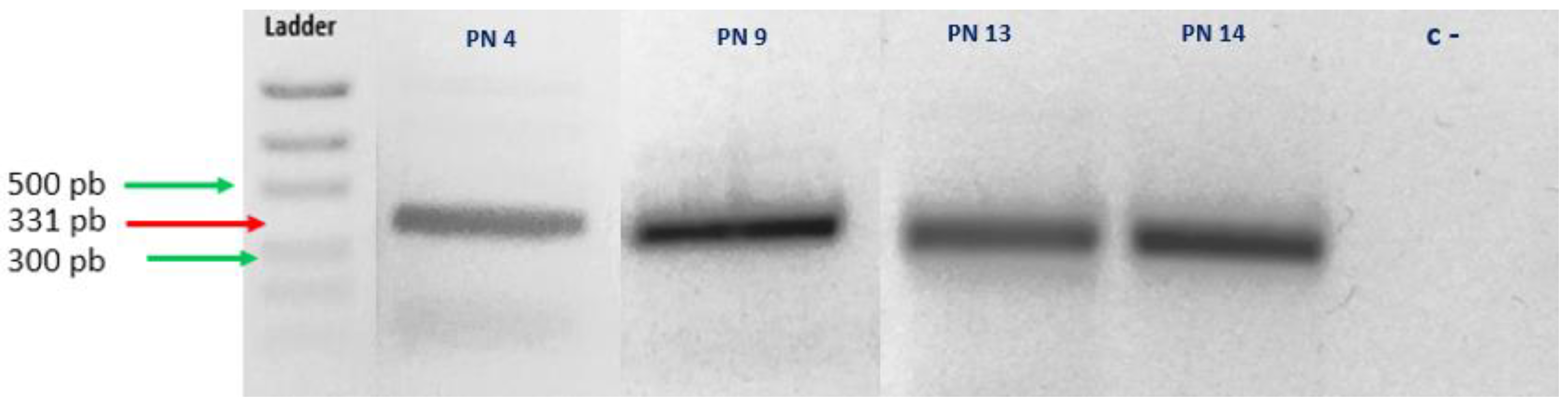
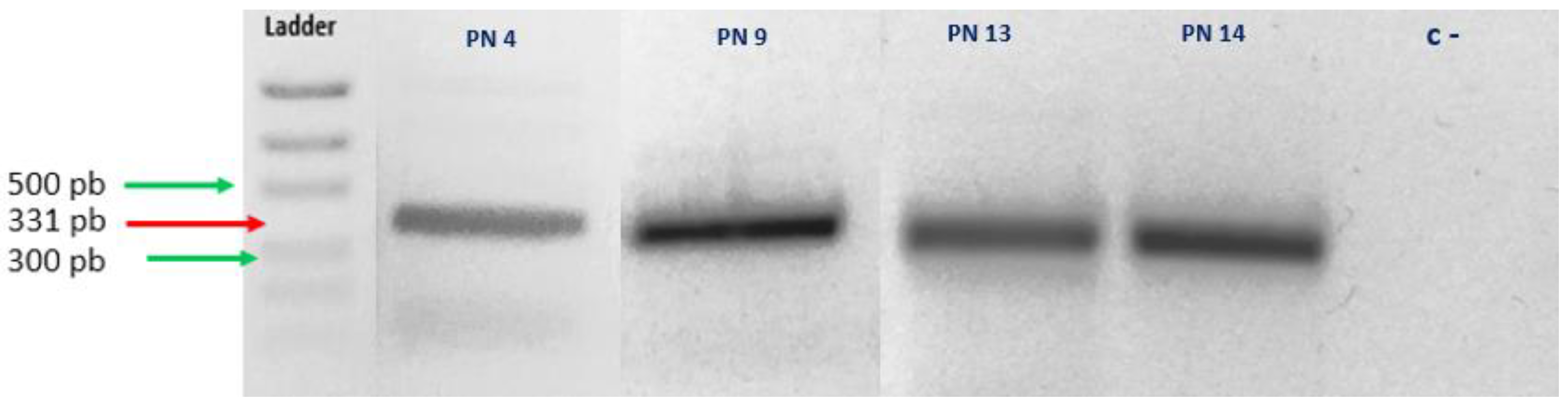
3.2. Confirmation of Haplosporidium pinnae Infection of P. nobilis by Sanger Sequencing
3.3. Quantification of Haplosporidium pinnae in Pinna nobilis
4. Discussion
5. Conclusions
- − A kit for blood and tissue DNA extraction renders high-quality DNA from P. nobilis mantle/digestive epithelium and faeces samples.
- − We detected Haplosporidium sp. in P. nobilis mantle and faeces using conventional PCR.
- − Sanger sequencing confirmed H. pinnae in mantle and faeces of infected P. nobilis.
- − About 7.1% of P. nobilis individuals (n = 28) from Mar Menor lagoon (SE Spain) are infected with H. pinnae.
- − P. nobilis-infected individuals contain 2.4 × 10−10–7.3 × 10−9 pg H. pinnae DNA µg−1 tissue.
- − P. nobilis faecal DNA analysis using PCR and qPCR can be used to monitor infection and recovery from H. pinnae.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Butler, A.; Vicente, N.; de Gaulejac, B. Ecology of the pterioid bivalves Pinna bicolor Gmelin and Pinna nobilis L. Mar. Life 1993, 3, 37–45. [Google Scholar]
- Zavodnik, D. Contribution to the Ecology of Pinna nobilis L. (Moll. bivalvia) in the Northern Adriatic. JAZU (Yugoslav Academy of Sciences and Arts (Jugoslavenska akademija znanosti i umjetnosti) 1967. Available online: https://scholar.google.com/scholar_lookup?title=Contribution+to+the+ecology+of+Pinna+nobilis+L.+(Moll.+Bivalvia)+in+the+Northern+Adriatic+sea&author=Zavodnik+D.&publication+year=1967&journal=Thalassia+Jugoslavica&volume=3&pages=93-102 (accessed on 21 December 2021).
- Zavodnik, D.; Hrs-Brenko, M.; Legac, M. Synopsis on the fan shell Pinna nobilis L. in the eastern Adriatic Sea. In Les Espèces Marines à Protéger en Méditerranée; Boudourescque, C.F., Avon, M., Gravez, V., Eds.; GIS Posidonie: Marseilles, France, 1991; pp. 169–178. [Google Scholar]
- Prado, P.; Grau, A.; Catanese, G.; Cabanes, P.; Carella, F.; Fernández-Tejedor, M.; Añón, K.T.; Hernandis, S.; Tena, J.; García-March, J.R. Pinna nobilis in suboptimal environments are more tolerant to disease but more vulnerable to severe weather phenomena. Mar. Environ. Res. 2021, 163, 105220. [Google Scholar] [CrossRef] [PubMed]
- Kersting, D.; Benabdi, M.; Čižmek, H.; Grau, A.; Jimenez, C.; Katsanevakis, S.; Öztürk, S.; Tuncer, L.; Tunesi, M.; Vázquez-Luis, N.; et al. Pinna nobilis. The IUCN Red List of Threatened Species. 2019, 2019-3. Available online: https://www.iucnredlist.org/species/160075998/160081499 (accessed on 21 December 2021).
- García-March, J.; Tena, J.; Henandis, S.; Vázquez-Luis, M.; López, D.; Téllez, C.; Prado, P.; Navas, J.I.; Bernal, J.; Catanese, G.; et al. Can we save a marine species affected by a highly infective, highly lethal, waterborne disease from extinction? Biol. Conserv. 2020, 243, 108498. [Google Scholar] [CrossRef]
- Orden TEC/1078/2018. Por la Que se Declara la Situación Crítica de Cistus Heterophyllus Subsp. Carthaginensis, Lanius Minor, Margaritifera Auricularia, Marmaronetta Angustirostris, Mustela Lutreola, Pinna nobilis y Tetrao Urogallus Cantabricus En España, y se Declaran de Interés General Las Obras y Proyectos Encaminados a la Recuperación de Dichos Taxones. Boletin Oficial de Estado (BOE). 28 September 2018. Available online: https://www.boe.es/eli/es/o/2018/09/28/tec1078 (accessed on 21 December 2021).
- Cabanellas-Reboredo, M.; Vázquez-Luis, M.; Mourre, B.; Álvarez, E.; Deudero, S.; Amores, Á.; Addis, P.; Ballesteros, E.; Barrajón, A.; Coppa, S.; et al. Tracking a mass mortality outbreak of pen shell Pinna nobilis populations: A collaborative effort of scientists and citizens. Sci. Rep. 2019, 9, 1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carella, F.; Aceto, S.; Pollaro, F.; Miccio, A.; Iaria, C.; Carrasco, N.; Prado, P.; De Vico, G. A mycobacterial disease is associated with the silent mass mortality of the pen shell Pinna nobilis along the Tyrrhenian coastline of Italy. Sci. Rep. 2019, 9, 2725. [Google Scholar] [CrossRef] [PubMed]
- Lattos, A.; Bitchava, K.; Giantsis, I.; Theodorou, J.; Batargias, C.; Michaelidis, B. The Implication of Vibrio Bacteria in the Winter Mortalities of the Critically Endangered Pinna nobilis. Microorganisms 2021, 9, 922. [Google Scholar] [CrossRef] [PubMed]
- Giménez-Casalduero, F.; Gomariz-Castillo, F.; Alonso-Sarría, F.; Cortés, E.; Izquierdo-Muñoz, A.; Ramos-Esplá, A. Pinna nobilis in the Mar Menor coastal lagoon: A story of colonization and uncertainty. Mar. Ecol. Prog. Ser. 2020, 652, 77–94. [Google Scholar] [CrossRef]
- Nebot-Colomer, E.; Álvarez, E.; Belando, M.D.; Deudero, S.; Catanese, G.; Bernardeau-Esteller, J.; García-Muñoz, R.; Ramos-Segura, A.; Ruiz, J.M.; Vázquez-Luis, M. Living under threat: Will one of the last Pinna nobilis populations be able to survive? Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 32, 1–13. [Google Scholar] [CrossRef]
- Chiou, K.L.; Bergey, C.M. Methylation-based enrichment facilitates low-cost, noninvasive genomic scale sequencing of populations from feces. Sci. Rep. 2018, 8, 1975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, K.; Fujiwara, H.; Zajac, C.; Sandford, E.; Reddy, P.; Choi, S.W.; Tewari, M. A Pipeline for Faecal Host DNA Analysis by Absolute Quantification of LINE-1 and Mitochondrial Genomic Elements Using ddPCR. Sci. Rep. 2019, 9, 5599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Sanmartín, M.; Catanese, G.; Grau, A.; Valencia, J.M.; García-March, J.R.; Navas, J.I. Real-Time PCR based test for the early diagnosis of Haplosporidium pinnae affecting fan mussel Pinna nobilis. PLoS ONE 2019, 14, e0212028. [Google Scholar] [CrossRef] [PubMed]
- Sanna, D.; Cossu, P.; Dedola, G.L.; Scarpa, F.; Maltagliati, F.; Castelli, A.; Franzoi, P.; Lai, T.; Cristo, B.; Curini-Galletti, M.; et al. Mitochondrial DNA Reveals Genetic Structuring of Pinna nobilis across the Mediterranean Sea. PLoS ONE 2013, 8, e67372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jódar-Pérez, A.B.; Terradas-Fernández, M.; López-Moya, F.; Asensio-Berbegal, L.; López-Llorca, L.V. Multidisciplinary Analysis of Cystoseira sensu lato (SE Spain) Suggest a Complex Colonization of the Mediterranean. J. Mar. Sci. Eng. 2020, 8, 961. [Google Scholar] [CrossRef]
- Renault, T.; Stokes, N.; Chollet, B.; Cochennec, N.; Berthe, F.; Gerard, A.; Burreson, E. Haplosporidiosis in the Pacific oyster Crassostrea gigas from the French Atlantic coast. Dis. Aquat. Org. 2000, 42, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Catanese, G.; Grau, A.; Valencia, J.; March, J.R.G.; Vázquez-Luis, M.; Alvarez, E.; Deudero, S.; Darriba, S.; Carballal, M.J.; Villalba, A. Haplosporidium pinnae sp. nov., a haplosporidan parasite associated with mass mortalities of the fan mussel, Pinna nobilis, in the Western Mediterranean Sea. J. Invertebr. Pathol. 2018, 157, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Fernández, V.; Dietrich, D.E.; Haney, R.L.; Tintoré, J. Mesoscale, seasonal and interannual variability in the Mediterranean Sea using a numerical ocean model. Prog. Oceanogr. 2005, 66, 321–340. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
| Sample Code | Date | Location | Sample | DNA Extraction Method | PCR (HapF1/HapR2) | PCR (LCOI1490/HC02198) |
|---|---|---|---|---|---|---|
| M1 | FEB-2021 | Local Supermarket | Mytilus edulis (Mantle necropsy) | CTAB | nt | nt |
| M2 | FEB-2021 | Local Supermarket | DNeasy PowerSoil Pro Kit | nt | nt | |
| M3 | FEB-2021 | Local Supermarket | DNeasy Mini Plant Kit | nt | nt | |
| M4 | FEB-2021 | Local Supermarket | DNeasy Blood & Tissue Kit | nt | nt | |
| PN1.1 | JUL-2021 | Pueblo Cálido (Mar Menor) | Pinna nobilis (Mantle biopsy) | CTAB | nt | nt |
| PN1.2 | JUL-2021 | Pueblo Cálido (Mar Menor) | DNeasy PowerSoil Pro Kit | nt | nt | |
| PN1.3 | JUL-2021 | Pueblo Cálido (Mar Menor) | DNeasy Mini Plant Kit | nt | nt | |
| PN1.4 | JUL-2021 | Pueblo Cálido (Mar Menor) | DNeasy Blood & Tissue Kit | nt | nt | |
| PNE 1 | JUL-2021 | Perdiguera Island NW (Mar Menor) | DNeasy Blood & Tissue Kits | nt | nt | |
| CP1 | FEB-2020 | Perdiguera Island NW (Mar Menor) | nt | nt | ||
| CP2 | FEB-2020 | Perdiguera Island SW (Mar Menor) | nt | nt | ||
| CP4 | FEB-2020 | Perdiguera Island SW (Mar Menor) | nt | nt | ||
| PN1 | FEB-2020 | Perdiguera Island SW (Mar Menor) | - | + | ||
| PN2 | FEB-2020 | Baron Island (Mar Menor) | - | + | ||
| PN3 | FEB-2020 | Baron Island (Mar Menor) | - | + | ||
| PN5 | MAY-2020 | Santiago de la Ribera (Mar Menor) | - | + | ||
| PN6 | MAY-2020 | Baron Island (Mar Menor) | - | + | ||
| PN7 | MAY-2020 | Baron Island (Mar Menor) | - | + | ||
| PN8 | MAY-2020 | Baron Island (Mar Menor) | - | + | ||
| PN9 | JUN_2020 | Pueblo Cálido (Mar Menor) | Pinna nobilis (Mantle necropsy and digestive gland) | + | + | |
| PN10 | JUN_2020 | Pueblo Cálido (Mar Menor) | - | + | ||
| PN11 | JUN_2020 | Baron Island (Mar Menor) | - | + | ||
| PN12 | JUN_2020 | Baron Island (Mar Menor) | - | + | ||
| PN13 * | JUL_2020 | IRTA (Delta del Ebro) | + | + | ||
| PN14 * | JUL_2020 | IRTA (Delta del Ebro) | + | + | ||
| PN16 | APR-2021 | Baron Island (Mar Menor) | - | + | ||
| PN17 | APR-2021 | Baron Island (Mar Menor) | - | + | ||
| PN18 | APR-2021 | Perdiguera Island NW (Mar Menor) | - | + | ||
| PN19 | APR-2021 | Perdiguera Island NW (Mar Menor) | - | + | ||
| PN20 | APR-2021 | Perdiguera Island SW (Mar Menor) | - | + | ||
| PN21 | APR-2021 | Baron Island (Mar Menor) | - | + | ||
| PN22 | APR-2021 | Baron Island (Mar Menor) | - | + | ||
| PN15 | FEB-2020 | Perdiguera Island NW (Mar Menor) | Pinna nobilis (Faeces) | - | + | |
| PN4 | MAY-2020 | Baron Island (Mar Menor) | + | + | ||
| PN 23 ** | APR-2021 | Galan beach (Mar Menor) | nt | + | ||
| PN 24 ** | APR-2021 | Galan beach (Mar Menor) | nt | + | ||
| PN25 | OCT-2021 | El Pedrucho (Mar Menor) | - | - | ||
| PN26 | OCT-2021 | El Pedrucho (Mar Menor) | - | - | ||
| PN27 | OCT-2021 | El Pedrucho (Mar Menor) | - | - | ||
| PN28 | OCT-2021 | El Pedrucho (Mar Menor) | - | - | ||
| PN29 | OCT-2021 | El Pedrucho (Mar Menor) | - | - | ||
| PN30 | OCT-2021 | El Pedrucho (Mar Menor) | - | + |
| Haplosporidium Primers | Sequence (5′-3′) | Amplicon Size (bp) | Tm | Reference |
|---|---|---|---|---|
| HapF1 | GTTCTTTCWTGATTCTATGMA | 331 | 49 | [18] |
| HapR2 | GATGAAYAATTGCAATCAYCT | 331 | 49 | |
| HPNF3 | CATTAGCATGGAATAATAAAACACGAC | 600 | 55 | |
| HPNR3 | GCGACGGCTATTTAGATGGCTGA | 600 | 55 | |
| Pinna nobilis Primers | Sequence (5′-3′) | Amplicon Size (bp) | Tm | Reference |
| LCOI1490 | GGTCAACAAATCATAAAGATATTGG | 710 | 40 | [19] |
| HC02198 | TAAACTTCAGGGTGACCAAAAAATCA | 710 | 40 |
| Sample | Most Similar Species (BLAST) | Query Cover | Identity |
|---|---|---|---|
| PN4 | Haplosporidium pinnae (MN689716.1) | 98% | 96.08% |
| PN9 | Haplosporidium pinnae (MN689716.1) | 99% | 90.83% |
| PN13 | Haplosporidium pinnae (MN689716.1) | 100% | 100% |
| PN14 | Haplosporidium pinnae (MN689716.1) | 100% | 100% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopez-Nuñez, R.; Cortés Melendreras, E.; Giménez Casalduero, F.; Prado, P.; Lopez-Moya, F.; Lopez-Llorca, L.V. Detection of Haplosporidium pinnae from Pinna nobilis Faeces. J. Mar. Sci. Eng. 2022, 10, 276. https://doi.org/10.3390/jmse10020276
Lopez-Nuñez R, Cortés Melendreras E, Giménez Casalduero F, Prado P, Lopez-Moya F, Lopez-Llorca LV. Detection of Haplosporidium pinnae from Pinna nobilis Faeces. Journal of Marine Science and Engineering. 2022; 10(2):276. https://doi.org/10.3390/jmse10020276
Chicago/Turabian StyleLopez-Nuñez, Raquel, Emilio Cortés Melendreras, Francisca Giménez Casalduero, Patricia Prado, Federico Lopez-Moya, and Luis Vicente Lopez-Llorca. 2022. "Detection of Haplosporidium pinnae from Pinna nobilis Faeces" Journal of Marine Science and Engineering 10, no. 2: 276. https://doi.org/10.3390/jmse10020276
APA StyleLopez-Nuñez, R., Cortés Melendreras, E., Giménez Casalduero, F., Prado, P., Lopez-Moya, F., & Lopez-Llorca, L. V. (2022). Detection of Haplosporidium pinnae from Pinna nobilis Faeces. Journal of Marine Science and Engineering, 10(2), 276. https://doi.org/10.3390/jmse10020276