Apatite Stimulates the Deposition of Glomalin-Related Soil Protein in a Lowbush Blueberry Commercial Field


Abstract
1. Introduction
2. Materials and Methods
2.1. Sand Bag Preparation
2.2. Field Soil Incubation
2.3. Glomalin-Related Soil Protein (GRSP) Extraction and Quantification
2.4. Statistical Analysis
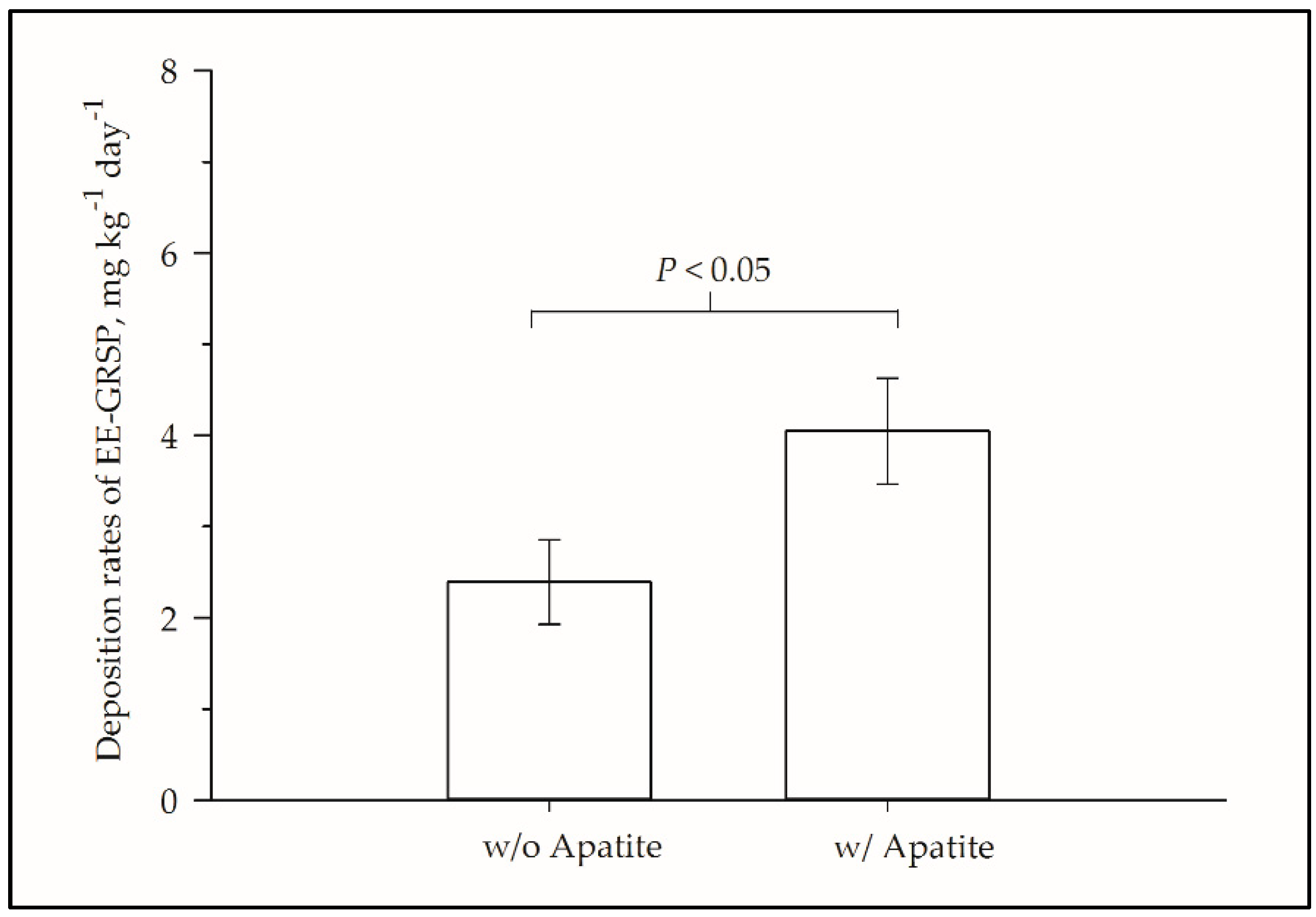
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- MAPAQ. Monographie de L’industrie du Bleuet Sauvage au Québec; Sous-ministériat aux Politiques Agroalimentaire. Direction du Développement des Secteurs Agroalimentaire; Ministère de L’Agriculture, des Pêcheries et de L’Alimentation du Québec (MAPAQ): Québec, QC, Canada, 2016; p. 32. [Google Scholar]
- Raymond, R.; Mailloux, A.; Dubé, A. Pédologie de la Région du Lac-Saint-Jean; Ministère de L’Agriculture et de la Colonisation du Québec, Division des Sols, Bulletin Technique No 11; La Pocatiére, QC, Canada, 1965; p. 159. [Google Scholar]
- Lafond, J. La restauration des zones dénudées dans les bleuetières, feuillet 3.4. In Guide de Production du Bleuet Sauvage Dans une Perspective de Développement Durable; Gagnon, S., Ed.; Centre de Référence en Agriculture et Agroalimentaire du Québec (CRAAQ): Québec, QC, Canada, 2014; p. 3. [Google Scholar]
- Hurisso, T.T.; Moebius-Clune, D.J.; Culman, S.W.; Moebius-Clune, B.N.; Thies, J.E.; Van Es, H.M. Soil protein as a rapid soil health indicator of potentially available organic nitrogen. Agric. Environ. Lett. 2018, 3, 1–5. [Google Scholar]
- Wright, S.F.; Upadhyaya, A. A survey of soils for aggregate stability and glomalin, a glycoprotein produced by hyphae of arbuscular mycorrhizal fungi. Plant Soil 1998, 198, 97–107. [Google Scholar] [CrossRef]
- Rillig, M.C. Arbuscular mycorrhizae, glomalin, and soil aggregation. Can. J. Soil Sci. 2004, 84, 355–363. [Google Scholar] [CrossRef]
- Wilson, G.W.T.; Rice, C.W.; Rillig, M.C.; Springer, A.; Hartnett, D.C. Soil aggregation and carbon sequestration are tightly correlated with the abundance of arbuscular mycorrhizal fungi: Results from long-term field experiments. Ecol. Lett. 2009, 12, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Bedini, S.; Pellegrino, E.; Avio, L.; Pellegrini, S.; Bazzoffi, P.; Argese, E.; Giovannetti, M. Changes in soil aggregation and glomalin-related soil protein content as affected by the arbuscular mycorrhizal fungal species Glomus mosseae and Glomus intraradices. Soil Biol. Biochem. 2009, 41, 1491–1496. [Google Scholar] [CrossRef]
- Yang, Y.; He, C.; Huang, L.; Ban, Y.; Tang, M. The effects of arbuscular mycorrhizal fungi on glomalin-related soil protein distribution, aggregate stability and their relationships with soil properties at different soil depths in lead-zinc contaminated area. PLoS ONE 2017, 12, e0182264. [Google Scholar] [CrossRef] [PubMed]
- Borie, F.; Rubio, R.; Rouanet, J.L.; Morales, A.; Borie, G.; Rojas, C. Effects of tillage systems on soil characteristics, glomalin and mycorrhizal propagules in a Chilean Ultisol. Soil Tillage Res. 2006, 88, 253–261. [Google Scholar] [CrossRef]
- Wright, S.F.; Green, V.S.; Cavigelli, M.A. Glomalin in aggregate size classes from three different farming systems. Soil Tillage Res. 2007, 94, 546–549. [Google Scholar] [CrossRef]
- Kohler, J.; Caravaca, F.; Roldan, A. An AM fungus and a PGPR intensify the adverse effects of salinity on the stability of rhizosphere soil aggregates of Lactuca sativa. Soil Biol. Biochem. 2010, 42, 429–434. [Google Scholar] [CrossRef]
- Jorge-Araujo, P.; Quiquampoix, H.; Matumoto-Pintro, P.T.; Staunton, S. Glomalin-related soil protein in French temperate forest soils: Interference in the Bradford assay caused by co-extracted humic substances. Eur. J. Soil Sci. 2015, 66, 311–319. [Google Scholar] [CrossRef]
- Wright, S.F.; Upadhyaya, A. Extraction of an abundant and unusual protein from soil and comparison with hyphal protein of arbuscular mycorrhizal fungi. Soil Sci. 1996, 161, 575–586. [Google Scholar] [CrossRef]
- Driver, J.D.; Holben, W.E.; Rillig, M.C. Characterization of glomalin as a hyphal wall component of arbuscular mycorrhizal fungi. Soil Biol. Biochem. 2005, 37, 101–106. [Google Scholar] [CrossRef]
- Halvorson, J.J.; Gonzalez, J.M. Bradford reactive soil protein in Appalachian soils: Distribution and response to incubation, extraction reagent and tannins. Plant Soil 2006, 286, 339–356. [Google Scholar] [CrossRef]
- Purin, S.; Rillig, M.C. The arbuscular mycorrhizal fungal protein glomalin: Limitations, progress, and a new hypothesis for its function. Pedobiologia 2007, 51, 123–130. [Google Scholar] [CrossRef]
- Gillespie, A.W.; Farrell, R.E.; Walley, F.L.; Ross, A.R.S.; Leinweber, P.; Eckhardt, K.U.; Regier, T.Z.; Blyth, R.I.R. Glomalin-related soil protein contains non-mycorrhizal-related heat-stable proteins, lipids and humic materials. Soil Biol. Biochem. 2011, 43, 766–777. [Google Scholar] [CrossRef]
- Walley, F.L.; Gillespie, A.W.; Adetona, A.B.; Germida, J.J.; Farrell, R.E. Manipulation of rhizosphere organisms to enhance glomalin production and C sequestration: Pitfalls and promises. Can. J. Plant Sci. 2014, 94, 1025–1032. [Google Scholar] [CrossRef]
- Rillig, M.C. A connection between fungal hydrophobins and soil water repellency? Pedobiologia 2005, 49, 395–399. [Google Scholar] [CrossRef]
- Wu, Q.S.; Srivastava, A.K.; Cao, M.Q. Systematicness of glomalin in roots and mycorrhizosphere of a split-root trifoliate orange. Plant Soil Environ. 2016, 62, 508–514. [Google Scholar]
- Kubota, M.; McGonigle, T.P.; Hyakumachi, M. Clethra barbinervis, a member of the order Ericales, forms arbuscular mycorrhizae. Can. J. Bot. 2001, 79, 300–306. [Google Scholar]
- Obase, K.; Matsuda, Y.; Ito, S. Enkianthus campanulatus (Ericaceae) is commonly associated with arbuscular mycorrhizal fungi. Mycorrhiza 2013, 23, 199–208. [Google Scholar] [CrossRef]
- Zhang, Y.; Ni, J.; Tang, F.; Jiang, L.; Guo, T.; Pei, K.; Sun, L.; Liang, Y. Diversity of root-Associated fungi of Vaccinium mandarinorum along a human disturbance gradient in subtropical forests, China. J. Plant Ecol. 2017, 10, 56–66. [Google Scholar] [CrossRef]
- Urcelay, C. Co-occurrence of three fungal root symbionts in Gaultheria poeppiggi DC in Central Argentina. Mycorrhiza 2002, 12, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Bedini, S.; Avio, L.; Argese, E.; Giovannetti, M. Effects of long-term land use on arbuscular mycorrhizal fungi and glomalin-related soil protein. Agric. Ecosyst. Environ. 2007, 120, 463–466. [Google Scholar] [CrossRef]
- García-González, I.; Quemada, M.; Gabriel, J.L.; Hontoria, C. Arbuscular mycorrhizal fungal activity responses to winter cover crops in a sunflower and maize cropping system. Appl. Soil Ecol. 2016, 102, 10–18. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Wright, S.F.; Nichols, K.A. Using glomalin as an indicator for arbuscular mycorrhizal hyphal growth: An example from a tropical rain forest soil. Soil Biol. Biochem. 2004, 36, 1009–1012. [Google Scholar] [CrossRef]
- Terman, G.L.; Bouldin, D.R.; Webb, J.R. Phosphorus availability: Crop response to phosphorus in water-insoluble phosphates varying in citrate solubility and granule size. J. Agric. Food Chem. 1961, 9, 166–170. [Google Scholar] [CrossRef]
- Gilkes, R.J.; Lim-Nunez, R. Poorly soluble phosphates in australian superphosphate: Their nature and availability to plants. Aust. J. Soil Res. 1980, 18, 85–95. [Google Scholar] [CrossRef]
- Smith, S.E.; Jakobsen, I.; Grønlund, M.; Smith, F.A. Roles of arbuscular mycorrhizas in plant phosphorus nutrition: Interactions between pathways of phosphorus uptake in arbuscular mycorrhizal roots have important implications for understanding and manipulating plant phosphorus acquisition. Plant Physiol. 2011, 156, 1050–1057. [Google Scholar] [CrossRef] [PubMed]
- Taktek, S.; Trépanier, M.; Servin, P.M.; St-Arnaud, M.; Piché, Y.; Fortin, J.A.; Antoun, H. Trapping of phosphate solubilizing bacteria on hyphae of the arbuscular mycorrhizal fungus Rhizophagus irregularis DAOM 197198. Soil Biol. Biochem. 2015, 90, 1–9. [Google Scholar] [CrossRef]
- Rosling, A.; Suttle, K.B.; Johansson, E.; Van Hees, P.A.W.; Banfield, J.F. Phosphorous availability influences the dissolution of apatite by soil fungi. Geobiology 2007, 5, 265–280. [Google Scholar] [CrossRef]
- Warman, P.R. The effects of pruning, fertilizers, and organic amendments on lowbush blueberry production. Plant Soil 1987, 101, 67–72. [Google Scholar] [CrossRef]
- Lafond, J.; Ziadi, N. Nitrogen and phosphorus fertilization in wild lowbush blueberry in Quebec. Can. J. Plant Sci. 2011, 91, 535–544. [Google Scholar] [CrossRef]
- Preger, A.C.; Rillig, M.C.; Johns, A.R.; Du Preez, C.C.; Lobe, I.; Amelung, W. Losses of glomalin-related soil protein under prolonged arable cropping: A chronosequence study in sandy soils of the South African Highveld. Soil Biol. Biochem. 2007, 39, 445–453. [Google Scholar] [CrossRef]
- Berner, C.; Johansson, T.; Wallander, H. Long-term effect of apatite on ectomycorrhizal growth and community structure. Mycorrhiza 2012, 22, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Hoogsteen, M.J.J.; Lantinga, E.A.; Bakker, E.J.; Groot, J.C.J.; Tittonell, P.A. Estimating soil organic carbon through loss on ignition: Effects of ignition conditions and structural water loss. Eur. J. Soil Sci. 2015, 66, 320–328. [Google Scholar] [CrossRef]
- Wallander, H.; Nilsson, L.O.; Hagerberg, D.; Bååth, E. Estimation of the biomass and seasonal growth of external mycelium of ectomycorrhizal fungi in the field. New Phytol. 2001, 151, 753–760. [Google Scholar] [CrossRef]
- Moreau, M.-È. Proline (prothioconazole), feuillet 4.5.2.5. In Guide de Production du Bleuet Sauvage Dans une Perspective de Développement Durable; Gagnon, S., Ed.; Centre de Référence en Agriculture et Agroalimentaire du Québec (CRAAQ): Québec, QC, Canada, 2016; p. 2. [Google Scholar]
- Reyna, D.L.; Wall, L.G. Revision of two colorimetric methods to quantify glomalin-related compounds in soils subjected to different managements. Biol. Fertil. Soils 2014, 50, 395–400. [Google Scholar] [CrossRef]
- Stoscheck, C.M. Quantitation of protein. Methods Enzymol. 1990, 182, 50–68. [Google Scholar]
- IBM Corp. IBM Corp. Released 2016. IBM SPSS Statistics for Windows, Version 24.0; IBM Corp: Armonk, NY, USA, 2016. [Google Scholar]
- Helgason, T.; Merryweather, J.W.; Young, J.P.W.; Fitter, A.H. Specificity and resilience in the arbuscular mycorrhizal fungi of a natural woodland community. J. Ecol. 2007, 95, 623–630. [Google Scholar] [CrossRef]
- Johansson, J.F.; Paul, L.R.; Finlay, R.D. Microbial interactions in the mycorrhizosphere and their significance for sustainable agriculture. FEMS Microbiol. Ecol. 2004, 48, 1–13. [Google Scholar] [CrossRef]
- Rosenstock, N.P.; Berner, C.; Smits, M.M.; Krám, P.; Wallander, H. The role of phosphorus, magnesium and potassium availability in soil fungal exploration of mineral nutrient sources in Norway spruce forests. New Phytol. 2016, 211, 542–553. [Google Scholar] [CrossRef] [PubMed]
- Bidartondo, M.I.; Ek, H.; Wallander, H.; Söderström, B. Do nutrient additions alter carbon sink strength of ectomycorrhizal fungi? New Phytol. 2001, 151, 543–550. [Google Scholar] [CrossRef]
- Singh, P.K.; Singh, M.; Tripathi, B.N. Glomalin: An arbuscular mycorrhizal fungal soil protein. Protoplasma 2013, 250, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, L.O.; Wallander, H. Production of external mycelium by ectomycorrhizal fungi in a norway spruce forest was reduced in response to nitrogen fertilization. New Phytol. 2003, 158, 409–416. [Google Scholar] [CrossRef]
- Smits, M.M.; Bonneville, S.; Benning, L.G.; Banwart, S.A.; Leake, J.R. Plant-driven weathering of apatite—The role of an ectomycorrhizal fungus. Geobiology 2012, 10, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Rillig, M.C.; Wright, S.F.; Eviner, V.T. The role of arbuscular mycorrhizal fungi and glomalin in soil aggregation: Comparing effects of five plant species. Plant Soil 2002, 238, 325–333. [Google Scholar] [CrossRef]
- Rillig, M.C.; Wright, S.F.; Nichols, K.A.; Schmidt, W.F.; Torn, M.S. Large contribution of arbuscular mycorrhizal fungi to soil carbon pools in tropical forest soils. Plant Soil 2001, 233, 167–177. [Google Scholar] [CrossRef]
- Steinberg, P.D.; Rillig, M.C. Differential decomposition of arbuscular mycorrhizal fungal hyphae and glomalin. Soil Biol. Biochem. 2003, 35, 191–194. [Google Scholar] [CrossRef]
- Feeney, D.S.; Daniell, T.; Hallett, P.D.; Illian, J.; Ritz, K.; Young, I.M. Does the presence of glomalin relate to reduced water infiltration through hydrophobicity? Can. J. Soil Sci. 2004, 84, 365–372. [Google Scholar] [CrossRef]
- Wallander, H.; Johansson, U.; Sterkenburg, E.; Brandström Durling, M.; Lindahl, B.D. Production of ectomycorrhizal mycelium peaks during canopy closure in Norway spruce forests. New Phytol. 2010, 187, 1124–1134. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Soil Properties | Method | Value |
|---|---|---|
| Soil pH | In water (1:1) | 4.4 |
| Soil organic matter (%) | Combustion | 5.1 |
| P (mg·kg−1) | Mehlich 3-Extractable | 37.1 |
| Al (mg·kg−1) | Mehlich 3-Extractable | 1864 |
| P saturation index (%) | (P/Al)×100 | 2.0 |
| K (mg·kg−1) | Mehlich 3-Extractable | 21.4 |
| Mg (mg·kg−1) | Mehlich 3-Extractable | 9.8 |
| Ca (mg·kg−1) | Mehlich 3-Extractable | 121.0 |
| Source of Variation | df | SS | F-Value | Probability |
|---|---|---|---|---|
| Apatite (w/ vs. w/o apatite) | 1 | 0.338 | 5.10 | 0.038 |
| Fungicide (w/ vs. w/o fungicide) | 1 | 0.000 | 0.01 | 0.941 |
| Apatite × Fungicide | 1 | 0.096 | 1.45 | 0.246 |
| Error | 16 | 1.061 | ||
| Total | 19 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paré, M.C.; Chagnon, P.-L.; Plourde, J.; Legendre-Guillemin, V. Apatite Stimulates the Deposition of Glomalin-Related Soil Protein in a Lowbush Blueberry Commercial Field. Agriculture 2019, 9, 52. https://doi.org/10.3390/agriculture9030052
Paré MC, Chagnon P-L, Plourde J, Legendre-Guillemin V. Apatite Stimulates the Deposition of Glomalin-Related Soil Protein in a Lowbush Blueberry Commercial Field. Agriculture. 2019; 9(3):52. https://doi.org/10.3390/agriculture9030052
Chicago/Turabian StyleParé, Maxime C., Pierre-Luc Chagnon, Joanne Plourde, and Valérie Legendre-Guillemin. 2019. "Apatite Stimulates the Deposition of Glomalin-Related Soil Protein in a Lowbush Blueberry Commercial Field" Agriculture 9, no. 3: 52. https://doi.org/10.3390/agriculture9030052
APA StyleParé, M. C., Chagnon, P.-L., Plourde, J., & Legendre-Guillemin, V. (2019). Apatite Stimulates the Deposition of Glomalin-Related Soil Protein in a Lowbush Blueberry Commercial Field. Agriculture, 9(3), 52. https://doi.org/10.3390/agriculture9030052