The Genetic Variability of Floral and Agronomic Characteristics of Newly-Bred Cytoplasmic Male Sterile Rice


Abstract
:1. Introduction
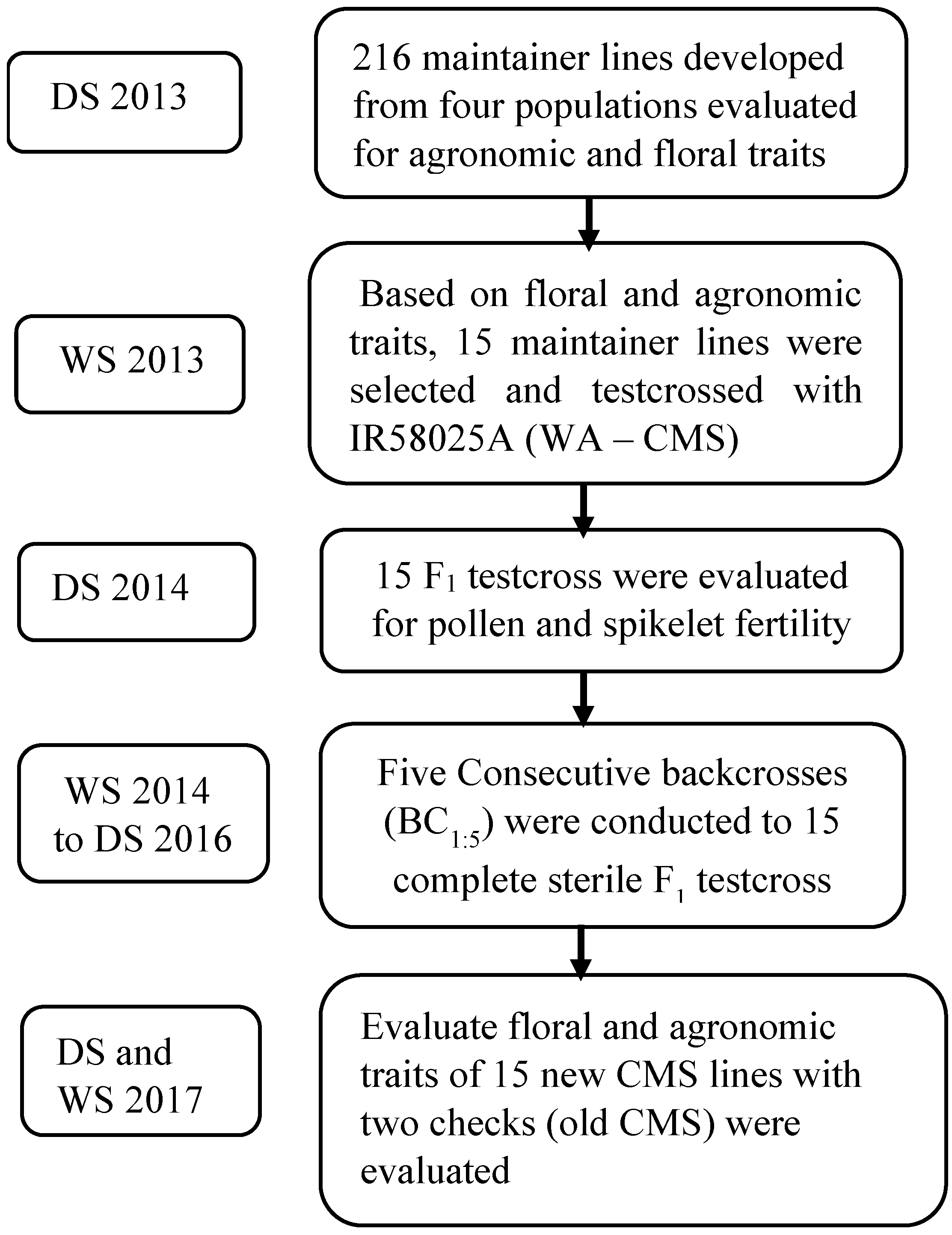
2. Materials and Methods
2.1. Backcrossing and Derived Offspring Assessment
2.2. Outcrossing Experiment
- EGA = the expected genetic advance under selection;
- σph = the phenotypic standard deviation;
- h2b = heritability in a broad sense;
- k = selection intensity.
3. Results
3.1. Maintainer Lines
Variance and Heritability
3.2. Newly-Bred CMS Lines
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- El-Namaky, R.; Nwilene, F.; Coulibaly, M.B.; Alhassan, M.; Traore, K.; Dieng, I.; Ortiz, R.; Manneh, B. Putting plant genetic diversity and variability at work for breeding: Hybrid rice suitability in West Africa. Diversity 2017, 9, 27. [Google Scholar] [CrossRef]
- Duvick, D.N. Heterosis: Feeding people and protecting natural resources. In Genetics and Exploitation of Heterosis; Coors, J.G., Pandey, S., Eds.; American Society of Agronomy—Crop Science Society of America: Madison, WI, USA, 1999; pp. 19–24. ISBN 9780891185499. [Google Scholar]
- Li, S.; Yang, D.; Zhu, Y. Characterization and Use of Male Sterility in Hybrid Rice Breeding. J. Integr. Plant Biol. 2007, 49, 791–804. [Google Scholar] [CrossRef]
- Guo, L.; Zhuang, J.; Qiu, F.; Gandhi, H.; Kadaru, S.; Jon, E.; Asis, D.; Xie, F. Genome-wide association study of outcrossing in cytoplasmic male sterile lines of rice. Sci. Rep. 2017, 7, 3223. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Kaur, R.; Yadav, S.S. Phenotypic characterization of new cytoplasmic male sterile lines in rice (Oryza sativa L.). J. Pharmacogn. Phytochem. 2017, 6, 166–167. [Google Scholar]
- Ma, X.; Zheng, Z.; Lin, F.; Ge, T.; Sun, H. Genetic analysis and gene mapping of a low stigma exposed mutant gene by high throughput sequencing. PLoS ONE 2018, 13, e0186942. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Li, P.B.; Xie, W.B.; Hussain, S.; Li, Y.B.; Xia, D. Genome-wide association analyses reveal the genetic basis of stigma exertion in rice. Mol. Plant 2017, 10, 634–644. [Google Scholar] [CrossRef] [PubMed]
- Parmar, K.S.; Swaminathan, M.S.; Siddiq, E.A. Variation in reproductive organs of rice with reference to male incompatibility index. Indian J. Genet. Plant Breed. 1980, 40, 262–271. [Google Scholar]
- Virmani, S.S.; Khush, G.S.; Yang, R.C. Rice cultivars possessing some desirable floral traits influencing outcrossing. Int. Rice Res. Newsl. 1980, 5, 4–5. [Google Scholar]
- Sarkar, H.C.; Miah, N.M. Extent of natural outcrossing of V20a, a cytoplasmic male-sterile (CMS) line in Bangladesh. Int. Rice Res. Newsl. 1983, 8, 7–8. [Google Scholar]
- Taillebois, J. New prospects for the production of F1 hybrid seed: Transfer of allogamous characters of O. longistaminata A. Chev. to O. sativa L. Agron. Trop. Nogent 1983, 38, 303–307. [Google Scholar]
- Pandey, P.; Tiwari, D.K.; Dwivedi, J.L. Evaluation of CMS lines for different allogamic traits that influence out crossing in rice (Oryza sativa L.). AAB Bioflux 2010, 2, 247–250. [Google Scholar]
- Li, T.; Chen, Y. Genetics of stigma exertion in rice. Rice Genet. Newsl. 1985, 2, 84–85. [Google Scholar]
- El-Namaky, R.A.; Demont, M. Hybrid rice in Africa: Challenges and prospects. In Realizing Africa’s Rice Promise; Wopereis, M.C.S., Ed.; CAB International: Wallingford, UK, 2013; pp. 173–178. ISBN 9781845938123. [Google Scholar]
- El-Namaky, R.; Van Oort, P. Phenology, sterility and inheritance of two environment genic male sterile (EGMS) lines for hybrid rice. Rice 2017, 10, 31. [Google Scholar] [CrossRef] [PubMed]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1983; p. 690. ISBN 0471-87092-7. [Google Scholar]
- Yang, B. Study on stigma exertion rate of sterile line and its seed setting rate. Hybrid Rice 1997, 12, 13–15. [Google Scholar]
- IRRI. Standard Evaluation System for Rice; International Rice Research Institute: Los Baños, Philippines, 2014; ISBN 9712203042. [Google Scholar]
- Burton, G.W.; Devane, E.M. Genetic variability, correlation and path coefficient analysis in rice. Agron. J. 1953, 45, 478–481. [Google Scholar] [CrossRef]
- Dilnesaw, Z.; Abadi, S.; Getahun, A. Genetic variability and heritability of soybean (Glycine max (L.) Merrill) genotypes in Pawe district, Metekel zone, Benishangule Gumuz regional state, Northwestern Ethiopia. Wudpecker J. Agric. Res. 2013, 2, 240–245. [Google Scholar]
- Hanson, C.H.; Robinson, H.F.; Comstock, R.E. Biometrical studies of yield in segregating populations of Korean Lespedeza. Agron. J. 1956, 48, 268–272. [Google Scholar] [CrossRef]
- Jonson, H.W.; Robinson, H.F.; Comstock, R.E. Estimates of genetic and environmental variability in soybean. Agron. J. 1955, 47, 314–318. [Google Scholar] [CrossRef]
- Robinson, H.F.; Akkaya, M.S.; Bhagwat, A.A.; Lavi, V.; Cregan, P.B. Estimation of genetic variability, heritability and genetic advance in “new plant type” tropical japonica rice under upland environment. Agron. J. 1951, 43, 262–267. [Google Scholar]
- Savitha, P.; Kumari, U.R.; Vanniarajan, C. Correlation between selected morphological character and yield components related traits in Oryza sativa L. Indian J. Exp. Biol. 2017, 55, 642–647. [Google Scholar]
- Majumder, R.R.; Roy, R.K.; Kader, M.A.; Aditya, T.L.; Ansari, A. Variability, correlation and path analysis in maintainer lines of aromatic rice for outcrossing traits. Bangladesh J. Crop Sci. 2013, 25, 139–146. [Google Scholar]
- Singh, R.K.; Chowdhury, B.D. Variability, heritability, and genetic advance in cultivars of rice (Oryza sativa L.). Crop Res. Hisar 1996, 12, 165–167. [Google Scholar]
- Iftekharuddaula, K.M.; Hasan, M.S.; Islam, M.J.; Badshah, M.A.; Islam, M.R.; Khaleda, A. Genetic evaluation and selection criteria of hybrid rice in irrigated ecosystem of Bangladesh. Pak. J. Biol. Sci. 2001, 4, 790–792. [Google Scholar]
- Khaleda, A.; Iftekharuddaula, K.M.; Basar, M.K.; Kabir, M.H.; Sarkar, M.Z.A. Genetic variability, correlation and path analysis in irrigated hybrid rice. J. Subtrop. Agric. Res. Dev. 2004, 2, 17–23. [Google Scholar]
- Hossain, M.A.; Mianm, A.K.; Rasul, M.G.; Hasan, M.J.; Kulsum, M.U.; Karim, M.A. Genetic variability in floral Traits of CMS lines and their relationship with outcrossing in rice. Trop. Agric. Dev. 2016, 60, 236–241. [Google Scholar]
- Virmani, S.S.; Athwal, D.S. Genetic variability for floral characters influencing outcrossing in Oryza sativa L. Crop Sci. 1973, 13, 66–67. [Google Scholar] [CrossRef]
- Tian, D.; Zhang, S.; Qin, C. Increase of stigma exertion rate is the key measure for high yielding in hybrid seed production of rice. Sichuan Agric. Sci. Technol. 1990, 2, 16–18. [Google Scholar]
- Dai, G.; Hua, Z.; Cai, W. Study on application of hormone in seed production of japonica hybrid rice. Hybrid Rice 1999, 14, 15–20. [Google Scholar]
- Kato, H.; Namai, H. Intervarietal variations of floral characteristics with special reference to F1 seed production in japonica rice (Oryza sativa L.). Jpn. J. Breed. 1987, 37, 75–87. [Google Scholar] [CrossRef]
- Virmani, S.S. Outcrossing mechanisms and hybrid seed production practices in rice. In Heterosis and Hybrid Rice Breeding; Springer Verlag—International Rice Research Institute: Manila, Philippines, 1994; pp. 79–96. [Google Scholar]
- Li, Q.L.; Lt, G.T.; Guo, G.Z.; Jiang, G.; Zhu, Y. Study on high yield breeding and genetic analysis of yield components of main rice cultivars in Jilin. Inst. Agric. Sci. 1991, 13, 3–6. [Google Scholar]


{kind=link}
{kind=link}
| Agronomic Characteristics | ||||||||
| Source of Variation | Degrees of Freedom | DAF | DAM | NPT | PH | GW | GYP | |
| Block | 11 | 63.05 | 1898.9 | 8.05 | 255.8 | 0.316 | 277 | |
| Lines | 217 | 34.35 ** | 46.5 ** | 2.33 NS | 40.73 ** | 0.11 ** | 59.16 ** | |
| Error | 11 | 0.678 | 1.041 | 1.5 | 6.06 | 0.01 | 12.56 | |
| Panicle and Floral Characteristics | ||||||||
| Source of Variation | Degrees of Freedom | SE | SOA | PnE | PnL | PnW | NS/Pn | SF% |
| Block | 11 | 465.1 | 15.62 | 20.5 | 8.58 | 0.758 | 1045.7 | 6.72 |
| Lines | 217 | 278.6 ** | 12.18 NS | 29.19 ** | 2.47 ** | 0.323 * | 269.4 ** | 25.21 * |
| Error | 11 | 10.1 | 6.74 | 0.5 | 0.4 | 0.12 | 26.79 | 10.21 |
| Characteristics | Range | Average | IR68897B | IR58025B | LSD0.05 | CV † |
|---|---|---|---|---|---|---|
| Days to 50% flowering | 69.46–108.46 | 96.95 | 85.67 | 104.25 | 1.89 | 0.01 |
| Days to maturity | 83.04–140.54 | 126.30 | 117.58 | 133.50 | 2.34 | 0.01 |
| Tillers per plant (#) | 10.87–20.57 | 15.44 | 17.00 | 18.33 | 2.80 | 0.08 |
| Plant height (cm) | 56.92–104.37 | 86.44 | 87.25 | 91.09 | 5.65 | 0.03 |
| 1000-grain weight (g) | 18.8–33.7 | 25.3 | 23.70 | 23.00 | 2.20 | 0.40 |
| Grain yield per plant (g) | 10.30–63.11 | 31.49 | 25.56 | 30.60 | 8.10 | 0.11 |
| Stigma exertion (%) | 1.13–76.16 | 30.14 | 24.86 | 29.84 | 7.27 | 0.11 |
| Spikelet opening angle (°) | 15.30–36.17 | 25.64 | 24.84 | 26.67 | 5.95 | 0.10 |
| Panicle exertion (%) | 78.0–100.0 | 96.84 | 100.00 | 100.00 | 0.00 | 0.00 |
| Panicle length (cm) | 20.46–29.76 | 26.06 | 24.45 | 25.64 | 1.47 | 0.02 |
| Panicle weight (g) | 1.79–5.88 | 3.25 | 3.23 | 3.24 | 0.79 | 0.11 |
| Spikelets per panicle | 91.82–197.67 | 126.10 | 115.92 | 125.30 | 12.47 | 0.04 |
| Spikelet fertility (%) | 73.76–97.74 | 84.78 | 87.54 | 88.09 | 7.28 | 0.04 |
| Characteristic | σ2G | σ2E | σ2P | GCV | PCV | H2 | GA |
|---|---|---|---|---|---|---|---|
| Days to 50% flowering | 27.53 | 0.63 | 28.15 | 5.41 | 5.42 | 75.0 | 11.14 |
| Days to maturity | 6.61 | 35.65 | 42.26 | 2.04 | 2.45 | 69.0 | 3.48 |
| Tillers per plant (#) | 0.99 | 0.91 | 1.90 | 6.43 | 6.68 | 93.0 | 12.77 |
| Plant height (cm) | 30.32 | 11.45 | 41.77 | 6.37 | 6.47 | 97.0 | 12.92 |
| 1000 grain weight (g) | 1.10 | 0.10 | 1.20 | 12.90 | 12.93 | 99.0 | 26.50 |
| Grain yield per plant (g) | 49.58 | 11.60 | 61.18 | 22.36 | 22.57 | 98.0 | 45.62 |
| Stigma exertion (%) | 295.52 | 5.99 | 301.51 | 57.03 | 57.08 | 97.0 | 117.39 |
| Spikelet opening angle (°) | 7.24 | 5.37 | 12.61 | 10.50 | 10.82 | 94.0 | 20.98 |
| Panicle exertion (%) | 26.72 | 0.12 | 26.84 | 5.24 | 5.34 | 98.0 | 11.00 |
| Panicle length (cm) | 2.07 | 0.35 | 2.41 | 5.52 | 5.56 | 99.0 | 11.29 |
| Panicle weight (g) | 0.23 | 0.11 | 0.34 | 14.75 | 15.04 | 96.0 | 29.81 |
| Spikelets per panicle (#) | 265.18 | 28.90 | 294.08 | 12.91 | 12.97 | 99.0 | 26.48 |
| Spikelet fertility (%) | 12.57 | 11.37 | 23.95 | 4.18 | 4.34 | 93.0 | 8.31 |
| Source of Variation | DF † | DAF | NPT | PH | PS | SpS |
| Replication | 2 | 1.24 | 1.92 | 3.42 | 0.0022 | 0.00005 |
| Line | 16 | 278.12 ** | 12.01 ** | 269.90 ** | 0.0004 | 0.00024 |
| Season | 1 | 10,360.60 ** | 0.09 | 23.83 * | 0.0004 | 1.6 × 10−5 |
| Season × Line | 16 | 17.79 ** | 1.30 | 4.33 | 0.0014 ** | 0.00182 ** |
| Residual | 66 | 4.12 | 1.82 | 4.62 | 0.0055 | 0.00050 |
| Source of Variation | DF | SE | SOA | SOT | PnE | OCR |
| Replication | 2 | 11.92 | 3.21 | 0.004 | 18.30 | 3.31 |
| Line | 16 | 1631.90 ** | 106.60 ** | 1.800 ** | 382.90 ** | 520.90 ** |
| Season | 1 | 4.96 | 12.50 | 0.004 | 5.99 | 5.52 |
| Season × Line | 16 | 16.91 | 5.83 | 0.009 | 17.12 | 5.19 |
| Residual | 66 | 25.93 | 8.80 | 0.240 | 15.81 | 6.84 |
| CMS Line | DAF | NPT | PH | PS% | SpS | SE | SOA | SOT | PnE | OCR |
|---|---|---|---|---|---|---|---|---|---|---|
| CMS 1 | 93.83 | 16.67 | 68.60 | 99.96 | 99.95 | 31.16 | 26.89 | 10.69 | 62.69 | 36.09 |
| CMS 2 | 81.83 | 17.00 | 78.17 | 99.96 | 99.95 | 23.17 | 29.67 | 9.32 | 55.00 | 31.67 |
| CMS 3 | 84.50 | 14.50 | 84.50 | 99.96 | 99.95 | 64.67 | 39.83 | 10.16 | 72.67 | 56.17 |
| CMS 4 | 82.00 | 15.67 | 79.67 | 99.95 | 99.96 | 24.83 | 32.67 | 10.18 | 58.00 | 36.17 |
| CMS 5 | 82.83 | 14.83 | 80.50 | 99.95 | 99.95 | 30.83 | 31.33 | 9.32 | 65.67 | 38.33 |
| CMS 6 | 84.00 | 15.50 | 85.50 | 99.94 | 99.95 | 33.67 | 34.50 | 9.48 | 65.83 | 40.83 |
| CMS 7 | 85.00 | 15.17 | 89.17 | 99.94 | 99.94 | 40.67 | 35.00 | 9.68 | 70.33 | 44.00 |
| CMS 8 | 87.33 | 14.17 | 89.00 | 99.95 | 99.95 | 42.33 | 34.50 | 10.25 | 67.00 | 43.33 |
| CMS 9 | 86.83 | 14.33 | 85.67 | 99.96 | 99.96 | 51.28 | 34.17 | 10.20 | 70.50 | 45.67 |
| CMS 10 | 94.00 | 16.67 | 80.27 | 99.96 | 99.95 | 19.15 | 26.30 | 10.79 | 58.25 | 32.28 |
| CMS 11 | 96.50 | 18.00 | 88.47 | 99.95 | 99.94 | 19.01 | 26.93 | 10.58 | 55.27 | 30.31 |
| CMS12 | 88.67 | 18.00 | 74.13 | 99.95 | 99.95 | 58.98 | 31.50 | 10.43 | 69.10 | 51.42 |
| CMS 13 | 78.50 | 13.83 | 78.37 | 99.94 | 99.95 | 19.69 | 25.47 | 11.03 | 50.87 | 26.87 |
| CMS14 | 97.33 | 17.50 | 90.55 | 99.94 | 99.94 | 53.61 | 29.83 | 10.43 | 69.54 | 48.44 |
| CMS 15 | 86.33 | 17.67 | 69.50 | 99.95 | 99.95 | 9.13 | 23.33 | 10.87 | 47.96 | 22.87 |
| IR58025A | 97.67 | 17.17 | 75.60 | 99.96 | 99.96 | 19.79 | 28.67 | 9.92 | 51.72 | 27.55 |
| IR68897A | 79.50 | 15.50 | 78.17 | 99.97 | 99.96 | 16.00 | 29.67 | 9.38 | 54.33 | 31.83 |
| Standard Error | 1.15 | 1.11 | 1,71 | 0.02 | 0.14 | 4.13 | 2.41 | 0.17 | 3.17 | 2.06 |
| LSD0.05 | 1.82 | 1.86 | 2.45 | 0.32 | 0.26 | 7.18 | 4.32 | 0.24 | 6.43 | 3.99 |
| NPT | PH | PS | SpS | SE | SOA | SOT | PnE | OCR | |
|---|---|---|---|---|---|---|---|---|---|
| DAF | 0.70 ** | −0.026 | −0.089 | −0.058 | 0.22 | −0.23 | 0.39 | 0.14 | 0.15 |
| NPT | −0.33 | −0.067 | −0.033 | −0.21 | −0.52 | 0.2 | −0.31 | −0.24 | |
| PH | −0.29 | −0.52 | 0.42 ** | 0.56 | −0.22 | 0.54 * | 0.48 | ||
| PS | 0.62 ** | −0.16 | 0.031 | −0.19 | −0.16 | −0.14 | |||
| SpS | −0.21 | −0.1 | −0.16 | −0.31 | −0.26 | ||||
| SE | 0.75 ** | −0.051 | 0.92 ** | 0.97 ** | |||||
| SOA | −0.49 | 0.79 ** | 0.82 ** | ||||||
| SOT | −0.21 | −0.16 | |||||||
| PnE | 0.95 ** |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Namaky, R. The Genetic Variability of Floral and Agronomic Characteristics of Newly-Bred Cytoplasmic Male Sterile Rice. Agriculture 2018, 8, 68. https://doi.org/10.3390/agriculture8050068
El-Namaky R. The Genetic Variability of Floral and Agronomic Characteristics of Newly-Bred Cytoplasmic Male Sterile Rice. Agriculture. 2018; 8(5):68. https://doi.org/10.3390/agriculture8050068
Chicago/Turabian StyleEl-Namaky, Raafat. 2018. "The Genetic Variability of Floral and Agronomic Characteristics of Newly-Bred Cytoplasmic Male Sterile Rice" Agriculture 8, no. 5: 68. https://doi.org/10.3390/agriculture8050068
APA StyleEl-Namaky, R. (2018). The Genetic Variability of Floral and Agronomic Characteristics of Newly-Bred Cytoplasmic Male Sterile Rice. Agriculture, 8(5), 68. https://doi.org/10.3390/agriculture8050068