Effect of Ascophyllum nodosum, Sideritis scardica and Fucus vesiculosus Extracts on Germination, Initial Growth and Antioxidant Potential of Red Russian Kale Microgreens

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Extracts
2.2. Assay of Extracts (Screening)
2.3. Germination Test
2.4. Pot Experiment
2.5. Assays for Antioxidant Activity
3. Results
3.1. Chemical Content of the Extracts
3.2. Germination Indexes
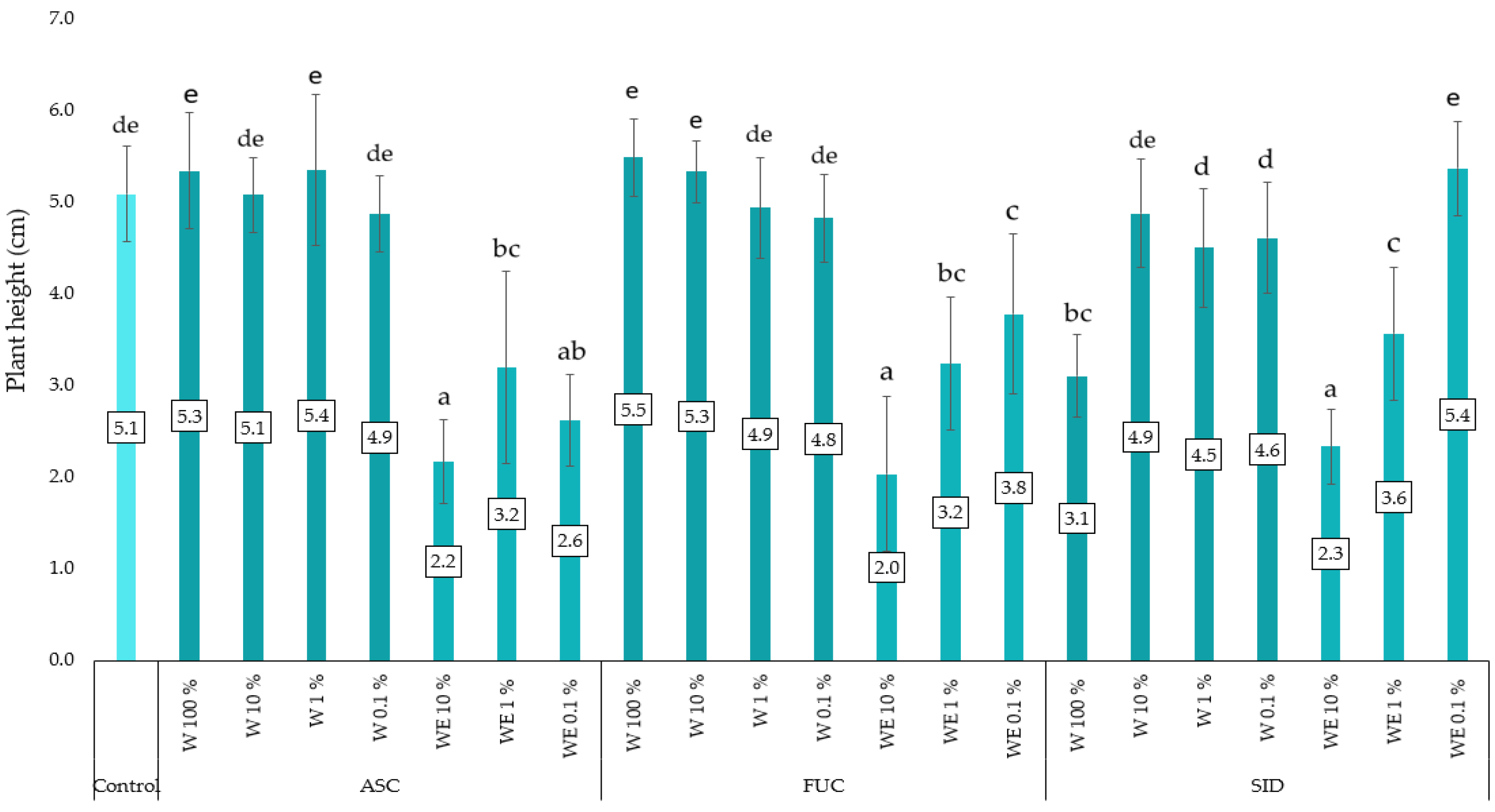
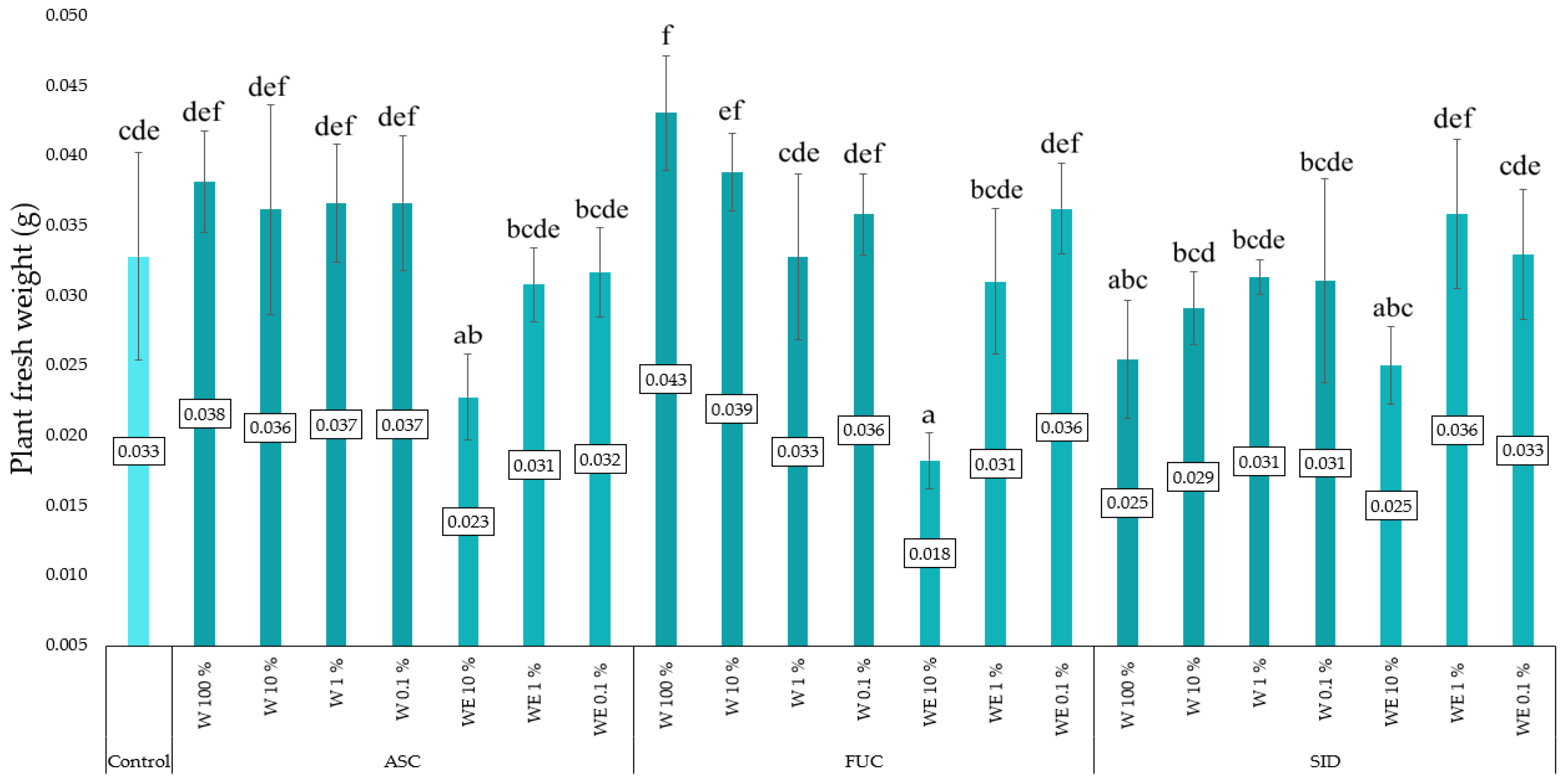
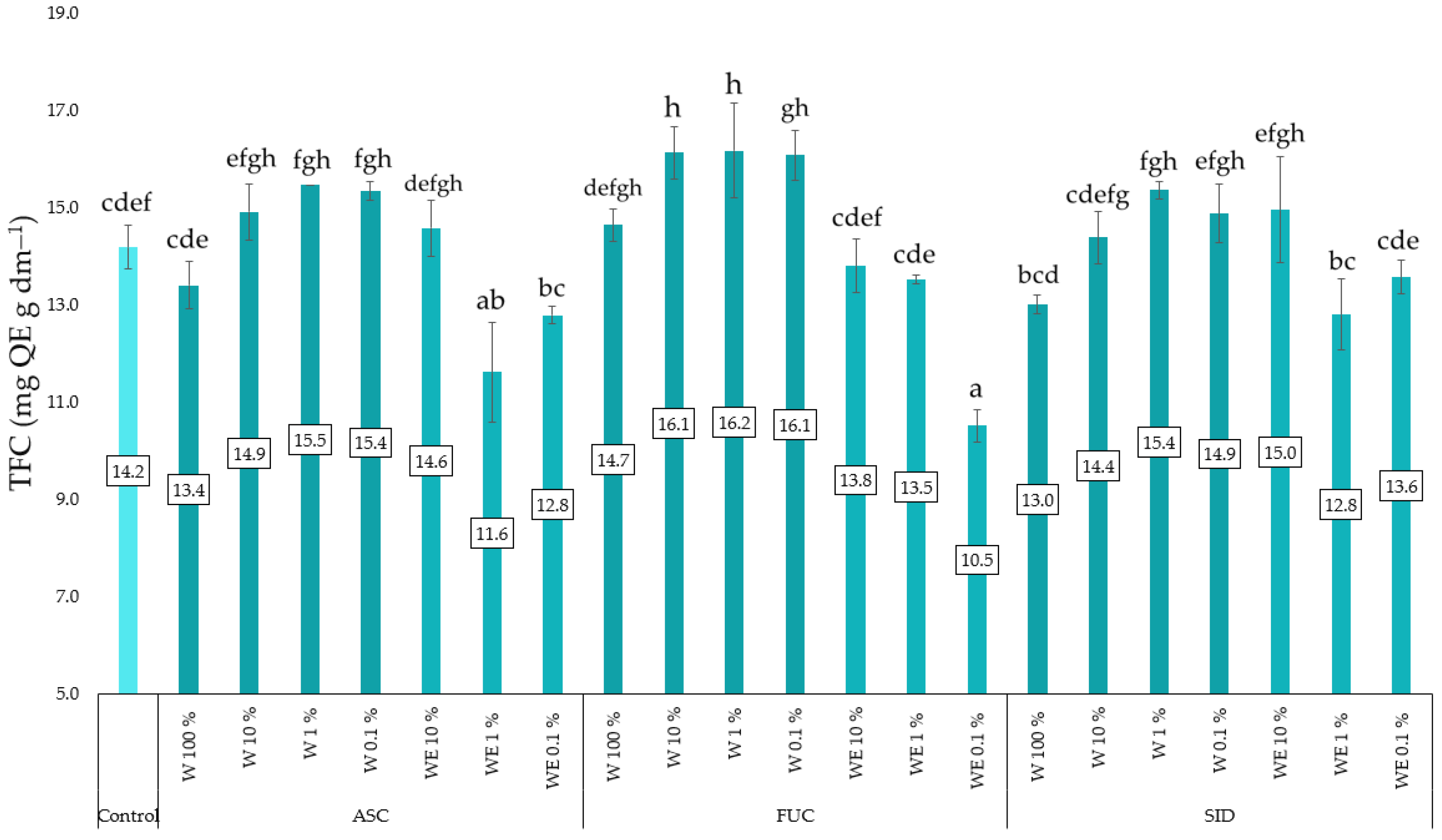
3.3. Linen Mat Experiment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| TPC | Total Phenolic Compound |
| TFC | Total Flavonoids |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl radical |
| ASC | Ascophyllum nodosum |
| FUC | Fucus vesiculosus |
| SID | Sideritis scardica |
| W | Water (aqueous) extract |
| WE | Water–ethanol extract |
| E | Ethanol extract |
| TE | Trolox equivalent |
| GAE | Gallic acid equivalent |
| QE | Gallic acid equivalent |
References
- Cowden, R.J.; Ghaley, B.B.; Henriksen, C.B. Analysis of light recipe, seeding density, and fertilization effects on secondary metabolite accumulation and growth-defense responses in Brassicaceae microgreens. Food Biosci. 2024, 59, 104071. [Google Scholar] [CrossRef]
- Goryńska-Goldmann, E.; Murawska, A.; Balcerowska-Czerniak, G. Consumer Profiles of Sustainable Fruit and Vegetable Consumption in the European Union. Sustainability 2023, 15, 15512. [Google Scholar] [CrossRef]
- Figueroa-Macías, J.P.; García, Y.C.; Núñez, M.; Díaz, K.; Olea, A.F.; Espinoza, L. Plant Growth-Defense Trade-Offs: Molecular Processes Leading to Physiological Changes. Int. J. Mol. Sci. 2021, 22, 693. [Google Scholar] [CrossRef]
- Li, T.; Lalk, G.T.; Bi, G. Fertilization and Pre-Sowing Seed Soaking Affect Yield and Mineral Nutrients of Ten Microgreen Species. Horticulturae 2021, 7, 14. [Google Scholar] [CrossRef]
- Mallor, C.; Bertolín, J.R.; Paracuellos, P.; Juan, T. Nutraceutical Potential of Leafy Vegetables Landraces at Microgreen, Baby, and Adult Stages of Development. Foods 2023, 12, 3173. [Google Scholar] [CrossRef]
- Gunjal, M.; Singh, J.; Kaur, J.; Kaur, S.; Nanda, V.; Sharma, A.; Rasane, P. Microgreens: Cultivation practices, bioactive potential, health benefits, and opportunities for its utilization as value-added food. Food Biosci. 2024, 62, 105133. [Google Scholar] [CrossRef]
- Bhaswant, M.; Shanmugam, D.K.; Miyazawa, T.; Abe, C.; Miyazawa, T. Microgreens—A Comprehensive Review of Bioactive Molecules and Health Benefits. Molecules 2023, 28, 867. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Colla, G.; De Pascale, S. Sprouts, Microgreens and Edible Flowers as Novel Functional Foods. Agronomy 2021, 11, 2568. [Google Scholar] [CrossRef]
- Teng, J.; Liao, P.; Wang, M. The role of emerging micro-scale vegetables in human diet and health benefits—An updated review based on microgreens. Food Funct. 2021, 12, 1914–1932. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, Z.; Ager, E.; Kong, L.; Tan, L. Nutritional quality and health benefits of microgreens, a crop of modern agriculture. J. Future Foods 2021, 1, 58–66. [Google Scholar] [CrossRef]
- Partap, M.; Sharma, D.; Deekshith, H.N.; Thakur, M.; Verma, V.; Bhargava, U.B. Microgreen: A tiny plant with superfood potential. J. Funct. Foods 2023, 107, 105697. [Google Scholar] [CrossRef]
- Gupta, A.; Sharma, T.; Singh, S.P.; Bhardwaj, A.; Srivastava, D.; Kumar, R. Prospects of microgreens as budding living functional food: Breeding and biofortification through OMICS and other approaches for nutritional security. Front. Genet. 2023, 14, 1053810. [Google Scholar] [CrossRef]
- Toscano, S.; Romano, D.; Patanè, C. Effect of Application of Biostimulants on the Biomass, Nitrate, Pigments, and Antioxidants Content in Radish and Turnip Microgreens. Agronomy 2023, 13, 145. [Google Scholar] [CrossRef]
- Di Gioia, F.; Hong, J.C.; Pisani, C.; Petropoulos, S.A.; Bai, J.; Rosskopf, E.N. Yield performance, mineral profile, and nitrate content in a selection of seventeen microgreen species. Front. Plant Sci. 2023, 14, 1220691. [Google Scholar] [CrossRef]
- Carillo, P.; Ciarmiello, L.F.; Woodrow, P.; Corrado, G.; Chiaiese, P.; Rouphael, Y. Enhancing Sustainability by Improving Plant Salt Tolerance through Macro- and Micro-Algal Biostimulants. Biology 2020, 9, 253. [Google Scholar] [CrossRef]
- Jithesh, M.N.; Shukla, P.; Kant, P.; Joshi, J.; Critchley, A.T.; Prithiviraj, B. Physiological And Transcriptomics Analysis reveal that asco phyllum nodosum extracts induce salinity tolerance in Arabidopsis by regulating the expression of stress responsive genes. J. Plant Growth Regul. 2018, 38, 463–478. [Google Scholar] [CrossRef]
- Drygaś, B.; Piechowiak, T.; Balawejder, M.; Matłok, N.; Kreczko, J.; Puchalski, C. The Eliciting Effect of Aqueous Extracts from Ascophyllum nodosum Algae on the Cultivation of Arugula (Eruca sativa Mill.) Microgreens. Sustainability 2024, 16, 7436. [Google Scholar] [CrossRef]
- Marchica, A.; Cotrozzi, L.; Detti, R.; Lorenzini, G.; Pellegrini, E.; Petersen, M.; Nali, C. The Biosynthesis of Phenolic Compounds Is an Integrated Defence Mechanism to Prevent Ozone Injury in Salvia officinalis. Antioxidants 2020, 9, 1274. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef]
- Constabel, C.P.; Barbehenn, R. Defensive Roles of Polyphenol Oxidase in Plants. In Induced Plant Resistance to Herbivory; Schaller, A., Ed.; Springer: New York, NY, USA, 2008; pp. 253–270. [Google Scholar]
- Drygaś, B.; Piechowiak, T.; Kreczko, J.; Matłok, N.; Saletnik, B.; Balawejder, M. The Utilisation of Fucus vesiculosus Algae Extracts in the Production of Microgreens Hordeum vulgare L. with an Increased Content of Selected Bioactive Compounds. Plants 2024, 13, 2871. [Google Scholar] [CrossRef]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Biostimulant Properties of Seaweed Extracts in Plants: Implications towards Sustainable Crop Production. Plants 2021, 10, 531. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Patel, J.S. Seaweed extract: Biostimulator of plant defense and plant productivity. Int. J. Environ. Sci. Technol. 2020, 17, 553–558. [Google Scholar] [CrossRef]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed Extracts as Biostimulants of Plant Growth and Development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Ahmad, A.; Blasco, B.; Martos, V. Combating Salinity Through Natural Plant Extracts Based Biostimulants: A Review. Front. Plant Sci. 2022, 13, 862034. [Google Scholar] [CrossRef]
- Zulfigar, F.; Casadesús, A.; Brockman, H.; Munné-Bosch, S. An overview of plant-based natural biostimulants for sustainable horticulture with a particular focus on moringa leaf extracts. Plant Sci. 2020, 295, 110194. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Sehrawat, K.D.; Phogat, D.; Sehrawat, A.R.; Chaudhary, R.; Sushkova, S.N.; Voloshina, M.S.; Rajput, V.D.; Shmaraeva, A.N.; Marc, R.A.; et al. Ascophyllum nodosum (L.) Le Jolis, a Pivotal Biostimulant toward Sustainable Agriculture: A Comprehensive Review. Agriculture 2023, 13, 1179. [Google Scholar] [CrossRef]
- Subramaniyan, L.; Veerasamy, R.; Prabhakaran, J.; Selvaraj, A.; Algarswamy, S.; Karuppasami, K.M.; Thangavel, K.; Nalliappan, S. Biostimulation Effects of Seaweed Extract (Ascophyllum nodosum) on Phytomorpho-Physiological, Yield, and Quality Traits of Tomato (Solanum lycopersicum L.). Horticulturae 2023, 9, 348. [Google Scholar] [CrossRef]
- Shakya, R.; Capilla, E.; Torres-Pagán, N.; Muñoz, M.; Boscaiu, M.; Lupuţ, I.; Vicente, O.; Verdeguer, M. Effect of Two Biostimulants, Based on Ascophyllum nodosum Extracts, on Strawberry Performance under Mild Drought Stress. Agriculture 2023, 13, 2108. [Google Scholar] [CrossRef]
- Campobenedetto, C.; Agliassa, C.; Mannino, G.; Vigliante, I.; Contartese, V.; Secchi, F.; Bertea, C.M. A Biostimulant Based on Seaweed (Ascophyllum nodosum and Laminaria digitata) and Yeast Extracts Mitigates Water Stress Effects on Tomato (Solanum lycopersicum L.). Agriculture 2021, 11, 557. [Google Scholar] [CrossRef]
- Krautforst, K.; Szymczycha-Madeja, A.; Wełna, M.; Michalak, I. Brown seaweed: Fucus vesiculosus as a feedstock for agriculture and environment protection. Sci. Rep. 2023, 13, 10065. [Google Scholar] [CrossRef]
- Walasek-Janusz, M.; Papliński, R.; Mysiak, B.; Nurzyńska-Wierdak, R. Phenolic Profile and Antioxidant Activity of Extracts from Aerial Parts of Thymus vulgaris L. and Sideritis scardica Griseb. Appl. Sci. 2025, 15, 3842. [Google Scholar] [CrossRef]
- Zheleva-Dimitrova, D.; Voynikov, Y.; Gevrenova, R.; Balabanova, V. A Comprehensive Phytochemical Analysis of Sideritis scardica Infusion Using Orbitrap UHPLC-HRMS. Molecules 2024, 29, 204. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Hernández, E.; Antunes-Ricardo, M.; Jacobo-Velázquez, D.A. Improving the Health-Benefits of Kales (Brassica oleracea L. var. acephala DC) through the Application of Controlled Abiotic Stresses: A Review. Plants 2021, 10, 2629. [Google Scholar] [CrossRef]
- Satheesh, N.; Workneh Fanta, S.; Yildiz, F. Kale: Review on nutritional composition, bio-active compounds, anti-nutritional factors, health beneficial properties and value-added products. Cogent Food Agric. 2020, 6, 1811048. [Google Scholar] [CrossRef]
- Khalid, W.; Iqra; Afzal, F.; Rahim, M.A.; Rehman, A.A.; Rasul, H.F.U.; Arshad, M.S.; Ambreen, S.; Zubair, M.; Safdar, S.; et al. Industrial applications of kale (Brassica oleracea var. sabellica) as a functional ingredient: A review. Int. J. Food Prop. 2023, 26, 489–501. [Google Scholar] [CrossRef]
- Šamec, D.; Urlić, B.; Salopek-Sondi, B. Kale (Brassica oleracea var. acephala) as a superfood: Review of the scientific evidence behind the statement. Crit. Rev. Food Sci. 2018, 59, 2411–2422. [Google Scholar] [CrossRef]
- Piechowiak, T.; Sowa-Borowiec, P. Changes in redox status in raspberry (Rubus idaeus L.) fruit during ripening. Biocatal. Agric. Biotechnol. 2024, 61, 103380. [Google Scholar] [CrossRef]
- Silva, L.D.; Bahcevandziev, K.; Pereira, L. Production of bio-fertilizer from Ascophyllum nodosum and Sargassum muticum (Phaeophyceae). J. Ocean. Limnol. 2019, 37, 918–927. [Google Scholar] [CrossRef]
- Karbarz, M.; Piziak, M.; Żuczek, J.; Duda, M. Influence of Microalgae Planktochlorella nurekis Clones on Seed Germination. Agronomy 2023, 13, 9. [Google Scholar] [CrossRef]
- Salehi, M.R.; Ashiri, F.; Salehi, H. Effect of Different Ethanol Concentrations on Seed Germination of Three Turfgrass Genera. Adv. Nat. Appl. Sci. 2008, 2, 6–9. [Google Scholar]
- Miyoshi, K.; Sato, T. The Effects of Ethanol on the Germination of Seeds of Japonica and Indica Rice (Oryza sativa L.) under Anaerobic and Aerobic Conditions. Ann. Bot. 1997, 79, 391–395. [Google Scholar] [CrossRef]
- Park, Y.J.; Park, J.-E.; Truong, T.Q.; Koo, S.Y.; Choi, J.-H.; Kim, S.M. Effect of Chlorella vulgaris on the Growth and Phytochemical Contents of “Red Russian” Kale (Brassica napus var. Pabularia). Agronomy 2022, 12, 2138. [Google Scholar] [CrossRef]
- Monteiro, M.; Santos, R.A.; Iglesias, P.; Couto, A.; Serra, C.R.; Gouvinhas, I.; Oliva-Teles, A.; Enes, P.; Díaz-Rosales, P. Effect of extraction method and solvent system on the phenolic content and antioxidant activity of selected macro- and microalgae extracts. J. Appl. Phycol. 2020, 32, 349–362. [Google Scholar] [CrossRef]
- Ingle, K.P.; Deshmukh, A.G.; Padole, D.A.; Dudhare, M.S.; Moharil, M.P.; Khelurkar, V.C. Phytochemicals: Extraction methods, identification and detection of bioactive compounds from plant extracts. Pharmacogn. J. 2017, 6, 32–36. Available online: https://www.phytojournal.com/archives/2017/vol6issue1/PartA/6-1-23-924.pdf (accessed on 15 April 2025).
- Lezoul, N.E.H.; Belkadi, M.; Habibi, F.; Guillén, F. Extraction Processes with Several Solvents on Total Bioactive Compounds in Different Organs of Three Medicinal Plants. Molecules 2020, 25, 4672. [Google Scholar] [CrossRef] [PubMed]
- Kaczorová, D.; Karalija, E.; Dahija, S.; Bešta-Gajević, R.; Parić, A.; Ćavar Zeljković, S. Influence of Extraction Solvent on the Phenolic Profile and Bioactivity of Two Achillea Species. Molecules 2021, 26, 1601. [Google Scholar] [CrossRef]
- García-Caparrós, P.; De Filippis, L.; Gul, A.; Hasanuzzaman, M.; Ozturk, M.; Altay, V.; Lao, M.T. Oxidative Stress and Antioxidant Metabolism under Adverse Environmental Conditions: A Review. Bot. Rev. 2021, 87, 421–466. [Google Scholar] [CrossRef]
- Huang, H.; Ullah, F.; Zhou, D.-X.; Yi, M.; Zhao, Y. Mechanisms of ROS Regulation of Plant Development and Stress Responses. Front. Plant Sci. 2019, 10, 800. [Google Scholar] [CrossRef]
- Haoujar, I.; Cacciola, F.; Abrini, J.; Mangraviti, D.; Giuffrida, D.; Oulad El Majdoub, Y.; Kounnoun, A.; Miceli, N.; Fernanda Taviano, M.; Mondello, L.; et al. The Contribution of Carotenoids, Phenolic Compounds, and Flavonoids to the Antioxidative Properties of Marine Microalgae Isolated from Mediterranean Morocco. Molecules 2019, 24, 4037. [Google Scholar] [CrossRef]
- Liu, Z.; Shi, J.; Wan, J.; Pham, Q.; Zhang, Z.; Sun, J.; Yu, L.; Luo, Y.; Wang, T.T.Y.; Chen, P. Profiling of Polyphenols and Glucosinolates in Kale and Broccoli Microgreens Grown under Chamber and Windowsill Conditions by Ultrahigh-Performance Liquid Chromatography High-Resolution Mass Spectrometry. ACS Food Sci. Technol. 2022, 2, 101–113. Available online: https://pubs.acs.org/doi/pdf/10.1021/acsfoodscitech.1c00355 (accessed on 15 April 2025). [CrossRef]
- Shin, J.; Lee, Y.; Hahm, S.; Lee, K.; Park, J. Effects of Exogenous Ethanol Treatment in Nutrient Solution on Growth and Secondary Metabolite Contents of Three Herb Species in an Indoor Vertical Farming System. Plants 2023, 12, 3842. [Google Scholar] [CrossRef] [PubMed]
- Stoica, R.; Velea, S.; Ilie, L.; Calugareanu, M.; Ghimis, S.B.; Ion, R.-M. The Influence of Ethanol Concentration on the Total Phenolics and Antioxidant Activity of Scenedesmus Opoliensis Algal Biomass Extracts. Rev. Chim. 2013, 64, 304–306. [Google Scholar]
- Alara, O.R.; Abdurahman, N.H.; Ukaegbu, C.I. Extraction of phenolic compounds: A review. Curr. Res. Food Sci. 2021, 4, 200–2014. [Google Scholar] [CrossRef]
- Plaskova, A.; Mlcek, J. New insights of the application of water or ethanol-water plant extract rich in active compounds in food. Front. Nutr. 2023, 28, 1118761. [Google Scholar] [CrossRef]
- Fraga-Corral, M.; García-Oliveira, P.; Pereira, A.G.; Lourenço-Lopes, C.; Jimenez-Lopez, C.; Prieto, M.A.; Simal-Gandara, J. Technological Application of Tannin-Based Extracts. Molecules 2020, 25, 614. [Google Scholar] [CrossRef] [PubMed]
- Kusuma, S.B.; Wulandari, S.; Nurfitriani, R.A.; Awaludin, A. The potential solvent for tannin extraction as a feed additive made of coffee husk (Coffea canephora) using Soxhlet Method. IOP Conf. Ser. Earth Environ. Sci. 2022, 980, 012024. [Google Scholar] [CrossRef]
- Strugała, P.; Tronina, T.; Huszcza, E.; Gabrielska, J. Bioactivity In Vitro of Quercetin Glycoside Obtained in Beauveria bassiana Culture and Its Interaction with Liposome Membranes. Molecules 2017, 22, 1520. [Google Scholar] [CrossRef] [PubMed]
- Fereira, O.; Pinho, S.P. Solubility of Flavonoids in Pure Solvents. Ind. Eng. Chem. Res. 2012, 51, 6586–6590. [Google Scholar] [CrossRef]
- Xia, B.-H.; Yu, Z.-L.; Lu, Y.-A.; Liu, S.-J.; Li, Y.-M.; Xie, M.-X.; Lin, L.-M. Green and Efficient Extraction of Phenolic Components from Plants with Supramolecular Solvents: Experimental and Theoretical Studies. Molecules 2024, 29, 2067. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Name | Phytochemical Group Detected | Result Expected |
|---|---|---|
| FeCl3 test | tannins | gallic tannins—navy blue color catechol tannins—dark green |
| gelatin test | tannins/phenolic compounds | precipitation |
| alkaline test (NaOH) | anthocyanins | blue color after addition of NaOH, turning yellow or orange after addition of HCl |
| alkaline test (NaOH) | flavonoids | intense yellow color after addition of NaOH, color fades after addition of HCl |
| lead acetate test | tannins | white precipitate |
| lead acetate test | flavonoids | yellow precipitate |
| Shinod test | flavonoids | red color of the solution |
| Mayer test | alkaloids | cream-colored precipitation/green solution |
| Wagner test | alkaloids | brown-red precipitate |
| Dragendorff test | alkaloids | orange precipitate |
| Keller–Killiani test | cardiac glycosides | cardiac glycosides—red-brown ring (steroid aglycone—part of the glycoside) |
| Liebermann–Burchard test | cardiac glycosides | cardiac glycosides—green color/ring (steroid aglycone—part of the glycoside) |
| Liebermann–Burchard test | steroids | green color/ring (steroid aglycone—part of the glycoside) |
| Liebermann–Burchard test | terpenoids | red-purple color |
| Salkowski test | steroids | red-brown ring (steroid aglycone—part of the glycoside) |
| Salkowski test | terpenoids | red-brown ring on the interface |
| Fehling test | reducing sugars/carbohydrates | brick-red precipitation |
| biuret reaction | proteins and amino acids | violet coloration |
| Xanthoproteic reaction | proteins and amino acids | after adding HNO3, a white precipitate forms. After dissolving it in ammonia, it turns yellow |
| Borntrager test | anthraquinones | pink, purple or violet coloring |
| Molish test | reducing sugars/carbohydrates | reddish-purple ring at the phase boundary |
| foam/emulsion test | saponins | persistent foam/formation of an emulsion with oil |
| Symbol | Solvent, Concentration | Description |
|---|---|---|
| Control | Control | distilled water only |
| ASC (Ascophyllum nodosum) | W 100% | undiluted aqueous extract |
| W 10% | aqueous extract diluted 10 times (10% extract) | |
| W 1% | aqueous extract diluted 100 times (1% extract) | |
| W 0.1% | aqueous extract diluted 1000 times (0.1% extract) | |
| W 0.01% | aqueous extract diluted 10,000 times (0.01% extract) | |
| WE 10% | water–ethanol extract diluted 10 times (10% extract) | |
| WE 1% | water–ethanol extract diluted 100 times (1% extract) | |
| WE 0.1% | water–ethanol extract diluted 1000 times (0.1% extract) | |
| WE 0.01% | water–ethanol diluted 10,000 times (0.01% extract) | |
| E 1% | ethanol extract diluted 100 times (1% extract) | |
| E 0.1% | ethanol extract diluted 1000 times (0.1% extract) | |
| E 0.01% | ethanol extract diluted 10,000 times (0.01% extract) | |
| FUC (Fucus vesiculosus) | W 100% | undiluted aqueous extract |
| W 10% | aqueous extract diluted 10 times (10% extract) | |
| W 1% | aqueous extract diluted 100 times (1% extract) | |
| W 0.1% | aqueous extract diluted 1000 times (0.1% extract) | |
| W 0.01% | aqueous extract diluted 10,000 times (0.01% extract) | |
| WE 10% | water–ethanol extract diluted 10 times (10% extract) | |
| WE 1% | water–ethanol extract diluted 100 times (1% extract) | |
| WE 0.1% | water–ethanol extract diluted 1000 times (0.1% extract) | |
| WE 0.01% | water–ethanol diluted 10,000 times (0.01% extract) | |
| E 1% | ethanol extract diluted 100 times (1% extract) | |
| E 0.1% | ethanol extract diluted 1000 times (0.1% extract) | |
| E 0.01% | ethanol extract diluted 10,000 times (0.01% extract) | |
| SID (Sideritis scardica) | W 100% | undiluted aqueous extract |
| W 10% | aqueous extract diluted 10 times (10% extract) | |
| W 1% | aqueous extract diluted 100 times (1% extract) | |
| W 0.1% | aqueous extract diluted 1000 times (0.1% extract) | |
| W 0.01% | aqueous extract diluted 10,000 times (0.01% extract) | |
| WE 10% | water–ethanol extract diluted 10 times (10% extract) | |
| WE 1% | water–ethanol extract diluted 100 times (1% extract) | |
| WE 0.1% | water–ethanol extract diluted 1000 times (0.1% extract) | |
| WE 0.01% | water–ethanol diluted 10,000 times (0.01% extract) | |
| E 1% | ethanol extract diluted 100 times (1% extract) | |
| E 0.1% | ethanol extract diluted 1000 times (0.1% extract) | |
| E 0.01% | ethanol extract diluted 10,000 times (0.01% extract) |
| Test Name | Phytochemical Group Detected | Result Obtained (+ Occurs; − Does Not Occur, +/− Inconclusive) | |||
|---|---|---|---|---|---|
| SID W(A) | SID W-E | ASC W-E | FUC W-E | ||
| FeCl3 test | tannins | 13B +/− dark brown solution with greenish tint | 11B +/− dark brown solution with greenish tint | 5B − brown solution with precipitate | 8B − light brown solution with gelatinous precipitate |
| gelatin test | tannins/phenolic compounds | 13C − no precipitate | 11C − no precipitate | 5C + light orange precipitate | 8C − no precipitate |
| alkaline test (NaOH) | anthocyanins | 13D − solution darkened to brownish-orange after addition of NaOH faded after addition of HCl | 11D − solution became brick red after addition of NaOH faded after addition of HCl | 5D − solution darkened to brownish-yellow after addition of NaOH faded after addition of HCl | 8D − solution darkened to light brownish-yellow after addition of NaOH faded after addition of HCl |
| alkaline test (NaOH) | flavonoids | 13D + solution darkened to brownish-orange after addition of NaOH faded after addition of HCl | 11D + solution became brick red after addition of NaOH faded after addition of HCl | 5D +/− Solution darkened to brownish-yellow after addition of NaOH faded after addition of HCl | 8D +/− solution darkened to light brownish-yellow after addition of NaOH faded after addition of HCl |
| lead acetate test | tannins | 13E − yellow precipitate | 11E − yellowish precipitate | 5E +/− cream precipitate | 8E + white precipitate |
| lead acetate test | flavonoids | 13E + yellow precipitate | 11E + yellowish precipitate | 5E +/− cream precipitate | 8E − white precipitate |
| Shinod test | flavonoids | 13F + light reddish coloration of the solution | 11F + intense reddish coloration of the solution | 5F − hazy brownish solution, foam present | 8F − no visible color change observed |
| Mayer test | alkaloids | 13G − after addition of HCl, precipitate appeared, after addition of Mayer reagent no visible changes were observed | 11G +/− after addition of HCl, turbidity appeared, after addition of Mayer reagent slight precipitation was observed | 5G − light turbidity appeared | 8G − no precipitate, color turned slightly yellowish |
| Wagner test | alkaloids | 13H +/− after addition of HCl, white precipitate was observed after addition of Wagner’s reagent, light reddish precipitate was observed | 11H − no precipitation observed, solution turned orange | 5H − no precipitation observed, solution turned orange | 8H − no precipitation observed, solution turned orange |
| Dragendorff test | alkaloids | 13I − white precipitate appeared after addition of HCl, after addition of Dragendorff’s reagent solution turned orange and no other visible changes were observed | 11I +/− turbidity appeared after addition of HCl, after addition of Dragendorff’s reagent solution turned bright orange and light ring of precipitate was visible | 5I − no precipitate solution turned bright orange | 8I − no precipitate solution turned bright orange |
| Keller–Killiani test | cardiac glycosides | 13J + brown ring/zone was observed on the junction of layers | 11J + distinct brown ring/zone was observed on the junction of layers | 5J + light brownish ring/zone observed on the junction of layers | 8J +/− very subtle zone of discoloration observed on the junction of layers |
| Liebermann–Burchard test | cardiac glycosides | 13K + greenish ring was observed | 11K − Solution turned slightly darker orange | 5K − solution turned orange foam was produced | 8K − solution turned slightly darker precipitate was produced (blue at first than hastily turning white) |
| Liebermann–Burchard test | steroids | 13K + greenish ring was observed | 11K − solution turned slightly darker orange | 5K − solution turned orange foam was produced | 8K − solution turned slightly darker precipitate was produced (blue at first than hastily turning white) |
| Liebermann–Burchard test | terpenoids | 13K − greenish ring was observed | 11K − solution turned slightly darker orange | 5K − solution turned orange foam was produced | 8K − solution turned slightly darker precipitate was produced (blue at first than hastily turning white) |
| Salkowski test | steroids | 13L + clearly visible brown ring/zone was observed | 11L + clearly visible brown ring/zone was observed | 5L + clearly visible brown ring was observed | 8L + clearly visible brown ring was observed |
| Salkowski test | terpenoids | 13L + clearly visible brown ring/zone was observed | 11L + clearly visible brown ring/zone was observed | 5L + clearly visible brown ring was observed | 8L + clearly visible brown ring was observed |
| Fehling test | reducing sugars/carbohydrates | 13M + brick-red precipitate was produced, solution turned olive-green | 11M + brick-red precipitate was produced, solution turned deep brick-red | 5M − no precipitate solution turned green | 8M − no precipitate, blue solution |
| biuret reaction | proteins and amino acids | 13N − solution turned light green | 11N − solution turned yellow-brownish | 5N − solution turned light green | 8N − solution turned light green |
| xanthoproteic reaction | proteins and amino acids | 13O +/− after adding HNO3, no precipitate was formed, but solution turned deep yellow after adding ammonia | 11O +/− after adding HNO3, no precipitate was formed, but solution turned deep yellow after adding ammonia | 5O +/− after adding HNO3, no precipitate was formed, but solution turned yellow after adding ammonia | 8O +/− after adding HNO3, no precipitate was formed, but solution turned yellow after adding ammonia |
| Borntrager test | anthraquinones | 13P − clear, orange and brown-orange layers were observed | 11P − orange and brown-orange layers were observed | 5P − brown solution of two layers and ring of dark brown precipitate were observed | 8P − brown gelatinous precipitate was observed |
| Molish test | reducing sugars/carbohydrates | 13R + pink reddish ring was observed under greyish layer at the junction of phases | 11R + thick reddish ring, turning into layer overtime | 5R +/− light brown-reddish ring/zone at the junction of phases | 8R +/− light reddish ring/zone at the junction of phases |
| foam/emulsion test | saponins | 13A +/− light foamy ring, persistent after 1 min, visible emulsion forming | 11A +/− light foamy ring, persistent after 1 min, visible emulsion forming | 5A − stable foam but emulsion not forming | 8A +/− light foamy ring, persistent after 1 min, visible emulsion forming |
| Object | Germination Energy (%) | Germination Capacity (%) | Maguire Index | Pieper’s Index | Healthy Non-Germinating Seeds (%) | Infected, Rotten Seeds(%) | |||
|---|---|---|---|---|---|---|---|---|---|
| 10 d | 25 d | 10 d | 25 d | ||||||
| Control | 83.3 ± 4.2 h | 84.7 ± 4.2 f | 22.0 ± 1.6 gh | 22.4 ± 1.4 hi | 2.0 ± 0.1 ab | 3.7 ± 0.7 ab | 2.7 ± 2.3 a | 2.0 ± 2.0 a | |
| ASC | W 100% | 61.3 ± 13.6 cd | 68.7 ± 11.0 cde | 13.7 ± 3.1 bc | 14.6 ± 2.7 cd | 3.4 ± 0.7 abcde | 6.4 ± 1.6 abcdef | 0.0 ± 0.0 a | 4.7 ± 3.1 a |
| W 10% | 74.0 ± 6.9 defgh | 79.3 ± 4.2 ef | 18.2 ± 2.2 cdefg | 18.7 ± 2.1 defghi | 3.1 ± 0.9 abcd | 4.8 ± 0.8 abcd | 1.3 ± 1.2 a | 3.3 ± 1.2 a | |
| W 1% | 80.7 ± 9.9 fgh | 86.0 ± 7.2 f | 20.2 ± 2.6 efg | 20.7 ± 2.4 fghi | 3.2 ± 1.0 abcde | 4.2 ± 1.1 abc | 0.0 ± 0.0 a | 1.3 ± 2.3 a | |
| W 0.1% | 82.0 ± 5.3 gh | 84.7 ± 2.3 f | 22.4 ± 1.5 h | 22.7 ± 1.6 i | 2.4 ± 0.7 ab | 3.7 ± 0.8 ab | 0.0 ± 0.0 a | 5.3 ± 5.0 a | |
| W 0.01% | 78.7 ± 2.3 efgh | 81.3 ± 1.2 ef | 21.1 ± 0.9 fhg | 21.5 ± 0.9 ghi | 2.5 ± 0.7 ab | 4.2 ± 0.6 abc | 0.7 ± 1.2 a | 5.3 ± 4.2 a | |
| WE 10% | 1.3 ± 2.3 a | 2.7 ± 1.2 a | 0.2 ± 0.2 a | 0.6 ± 0.2 a | 7.0 ± 1.7 efg | 16.9 ± 1.7 jk | 67.3 ± 13.0 defgh | 16.7 ± 15.5 abc | |
| WE 1% | 2.7 ± 1.1 a | 2.7 ± 1.2 a | 0.3 ± 0.1 a | 0.6 ± 0.3 a | 5.0 ± 0.0 abcdef | 15.8 ± 2.0 ijk | 70.7 ± 13.8 ef ghi | 14.0 ± 11.1 abc | |
| WE 0.1% | 0.7 ± 1.1 a | 2.7 ± 1.2 a | 0.2 ± 0.1 a | 0.9 ± 0.3 a | 7.8 ± 1.3 fg | 16.9 ± 0.5 jk | 64.0 ± 10.6 defg | 8.7 ± 3.1 a | |
| WE 0.01% | 4.0 ± 2.0 ab | 4.0 ± 2.0 a | 0.9 ± 0.7 a | 1.6 ± 0.5 a | 2.8 ± 0.4 abc | 16.6 ± 1.1 jk | 61.3 ± 5.8 def | 8.0 ± 2.0 a | |
| E 1% | 0.0 ± 0.0 a | 2.0 ± 0.0 a | 0.0 ± 0.1 a | 0.1 ± 0.1 a | 9.7 ± 0.6 g | 11.2 ± 3.0 fghi | 88.0 ± 6.9 hi | 8.7 ± 6.1 a | |
| E 0.1% | 2.7 ± 1.1 a | 2.7 ± 1.2 a | 0.7 ± 0.3 a | 0.7 ± 0.3 a | 2.5 ± 1.3 ab | 3.8 ± 3.0 ab | 86.7 ± 6.4 ghi | 10.0 ± 6.9 ab | |
| E 0.01% | 3.3 ± 1.2 ab | 3.3 ± 1.2 a | 0.4 ± 0.1 a | 0.4 ± 0.2 a | 4.0 ± 0.0 abcdef | 4.8 ± 1.3 abcd | 87.3 ± 4.6 ghi | 8.7 ± 3.1 a | |
| FUC | W 100% | 48.7 ± 8.3 c | 56.7 ± 6.4 c | 10.5 ± 2.1 b | 11.8 ± 1.8 c | 5.4 ± 1.3 bcdef | 8.5 ± 1.7 bcdefg | 3.3 ± 4.2 a | 2.7 ± 3.1 a |
| W 10% | 66.7 ± 6.4 defg | 68.7 ± 4.6 cde | 15.9 ± 1.1 cde | 16.5 ± 1.1def | 2.2 ± 0.1 ab | 5.5 ± 0.4 abcd | 10.7 ± 5.0 ab | 3.3 ± 4.2 a | |
| W 1% | 64.7 ± 9.9 cdef | 68.7 ± 8.1 cde | 17.4 ± 2.6 cdefg | 18.0 ± 2.6 defgh | 2.5 ± 0.6 ab | 5.6 ± 0.8 abcde | 8.0 ± 3.5 ab | 4.0 ± 5.3 a | |
| W 0.1% | 76.0 ± 5.3 defgh | 77.3 ± 4.2 def | 19.4 ± 2.5 defgh | 19.8 ± 2.5 efghi | 2.2 ± 0.2 ab | 5.0 ± 0.5 abcd | 4.7 ± 3.1 ab | 2.0 ± 2.0 a | |
| W 0.01% | 66.7 ± 11.0 defg | 68.7 ± 9.2 cde | 17.0 ± 3.1 cdef | 17.4 ± 3.2 defg | 2.6 ± 0.8 ab | 5.5 ± 0.8 abcde | 10.7 ± 10.1 ab | 4.0 ± 5.3 a | |
| WE 10% | 0.0 ± 0.0 a | 2.0 ± 0.0 a | 0.0 ± 0.1 a | 0.3 ± 0.0 a | 9.7 ± 0.6 g | 17.6 ± 1.2 k | 57.3 ± 23.9 de | 34.0 ± 23.6 c | |
| WE 1% | 2.0 ± 0.0 a | 2.0 ± 0.0 a | 0.8 ± 0.3 a | 1.3 ± 0.3 a | 1.3 ± 0.6 a | 16.6 ± 3.0 jk | 64.7 ± 5.0 defgh | 15.3 ± 9.0 abc | |
| WE 0.1% | 2.0 ± 0.0 a | 2.0 ± 0.0 a | 0.8 ± 0.3 a | 1.1 ± 0.4 a | 1.3 ± 0.6 a | 15.7 ± 0.9 ijk | 65.3 ± 4.6 defgh | 22.0 ± 3.5 abc | |
| WE 0.01% | 2.7 ± 1.2 a | 4.7 ± 3.1 a | 1.1 ± 0.5 a | 1.6 ± 0.3 a | 1.3 ± 0.8 a | 15.9 ± 1.7 efghi | 45.3 ± 8.1 cd | 31.3 ± 2.3 bc | |
| E 1% | 0.0 ± 0.0 a | 2.0 ± 0.0 a | 0.0 ± 0.1 a | 0.2 ± 0.1 a | 9.7 ± 0.6 g | 10.8 ± 1.3 defgh | 85.3 ± 1.2 ghi | 11.3 ± 3.1 ab | |
| E 0.1% | 0.0 ± 0.0 a | 2.0 ± 0.0 a | 0.0 ± 0.1 a | 0.1 ± 0.1 a | 9.7 ± 0.6 g | 10.0 ± 0.0 bcdefg | 91.3 ± 5.0 i | 6.0 ± 4.0 abc | |
| E 0.01% | 2.0 ± 0.0 a | 4.0 ± 0.0 a | 0.3 ± 0.2 a | 0.6 ± 0.4 a | 6.8 ± 0.8 defg | 8.4 ± 0.9 bcdefg | 78.7 ± 5.8 efghi | 14.0 ± 8.0 abc | |
| SID | W 100% | 19.3 ± 4.2 b | 26.7 ± 3.1 b | 4.5 ± 0.4 a | 6.2 ± 0.2 b | 3.0 ± 0.0 abc | 11.7 ± 1.0 ghij | 18.7 ± 4.2 ab | 4.7 ± 3.1 a |
| W 10% | 65.3 ± 7.0 def | 69.3 ± 7.0 cde | 16.2 ± 2.0 cde | 16.6 ± 1.9 def | 2.6 ± 0.7 ab | 5.3 ± 0.6 abcd | 7.3 ± 4.2 ab | 7.3 ± 4.2 a | |
| W 1% | 64.7 ± 6.4 cdef | 68.7 ± 6.4 cde | 16.1 ± 2.7 cde | 16.7 ± 2.6 def | 2.3 ± 0.1 ab | 5.8 ± 0.3 abcde | 10.0 ± 6.9 ab | 2.7 ± 1.2 a | |
| W 0.1% | 62.0 ± 6.9 cd | 64.7 ± 5.0 cd | 14.9 ± 2.2 bc | 15.4 ± 2.0 cde | 2.3 ± 0.3 ab | 5.7 ± 0.9 abcde | 10.7 ± 3.1 ab | 8.0 ± 0.0 a | |
| W 0.01% | 62.7 ± 4.2 cde | 64.7 ± 5.8 cd | 15.9 ± 1.5 cde | 16.0 ± 1.3 cde | 2.1 ± 0.1 ab | 3.1 ± 0.6 a | 28.0 ± 2.0 bc | 3.3 ± 2.3 a | |
| WE 10% | 0.0 ± 0.0 a | 2.0 ± 0.0 a | 0.0 ± 0.1 a | 0.2 ± 0.1 a | 9.7 ± 0.6 g | 15.9 ± 1.6 ijk | 74.0 ± 8.7 efghi | 19.3 ± 5.8 abc | |
| WE 1% | 4.7 ± 2.3 ab | 4.7 ± 2.3 a | 1.9 ± 0.8 a | 2.3 ± 0.9 ab | 1.3 ± 0.3 a | 15.2 ± 2.4 hijk | 68.7 ± 6.4 defghi | 14.0 ± 4.0 abc | |
| WE 0.1% | 4.0 ± 0.0 ab | 5.3 ± 2.3 a | 1.3 ± 0.1 a | 1.9 ± 0.2 ab | 3.6 ± 1.9 abcde | 15.4 ± 1.0 ijk | 58.0 ± 5.3 de | 14.7 ± 7.6 abc | |
| WE 0.01% | 2.0 ± 0.0 a | 3.3 ± 1.2 a | 0.9 ± 0.4 a | 1.3 ± 0.4 a | 3.7 ± 1.4 abcde | 14.9 ± 1.4 hijk | 61.3 ± 4.2 def | 22.0 ± 6.0 abc | |
| E 1% | 0.0 ± 0.0 a | 2.0 ± 0.0 a | 0.1 ± 0.0 a | 0.2 ± 0.1 a | 5.3 ± 4.6 bcdef | 9.3 ± 4.2 cdefg | 87.3 ± 6.4 ghi | 8.0 ± 2.0 a | |
| E 0.1% | 2.7 ± 3.1 a | 4.0 ± 3.5 a | 0.4 ± 0.3 a | 0.5 ± 0.4 a | 3.9 ± 0.1 abcde | 6.7 ± 2.6 abcdefg | 83.3 ± 5.8 fghi | 11.3 ± 5.8 ab | |
| E 0.01% | 0.7 ± 1.1 a | 4.0 ± 2.0 a | 0.2 ± 0.1 a | 0.4 ± 0.1 a | 6.6 ± 2.8 cdefg | 9.5 ± 1.2 defg | 86.0 ± 0.0 ghi | 8.0 ± 2.0 a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drygaś, B.; Szpunar-Krok, E.; Kreczko, J.; Piechowiak, T.; Puchalski, C.; Jańczak-Pieniążek, M. Effect of Ascophyllum nodosum, Sideritis scardica and Fucus vesiculosus Extracts on Germination, Initial Growth and Antioxidant Potential of Red Russian Kale Microgreens. Agriculture 2025, 15, 961. https://doi.org/10.3390/agriculture15090961
Drygaś B, Szpunar-Krok E, Kreczko J, Piechowiak T, Puchalski C, Jańczak-Pieniążek M. Effect of Ascophyllum nodosum, Sideritis scardica and Fucus vesiculosus Extracts on Germination, Initial Growth and Antioxidant Potential of Red Russian Kale Microgreens. Agriculture. 2025; 15(9):961. https://doi.org/10.3390/agriculture15090961
Chicago/Turabian StyleDrygaś, Barbara, Ewa Szpunar-Krok, Joanna Kreczko, Tomasz Piechowiak, Czesław Puchalski, and Marta Jańczak-Pieniążek. 2025. "Effect of Ascophyllum nodosum, Sideritis scardica and Fucus vesiculosus Extracts on Germination, Initial Growth and Antioxidant Potential of Red Russian Kale Microgreens" Agriculture 15, no. 9: 961. https://doi.org/10.3390/agriculture15090961
APA StyleDrygaś, B., Szpunar-Krok, E., Kreczko, J., Piechowiak, T., Puchalski, C., & Jańczak-Pieniążek, M. (2025). Effect of Ascophyllum nodosum, Sideritis scardica and Fucus vesiculosus Extracts on Germination, Initial Growth and Antioxidant Potential of Red Russian Kale Microgreens. Agriculture, 15(9), 961. https://doi.org/10.3390/agriculture15090961