
Regulatory Effects of Mowing on Biomass Allocation and Compensation Growth Mechanisms in Elymus Species

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling and Measurement of Indicators
2.3. Statistical Analysis
3. Results
3.1. Effects of Mowing on the Individual Biomass and Component Allocation of Elymus Species
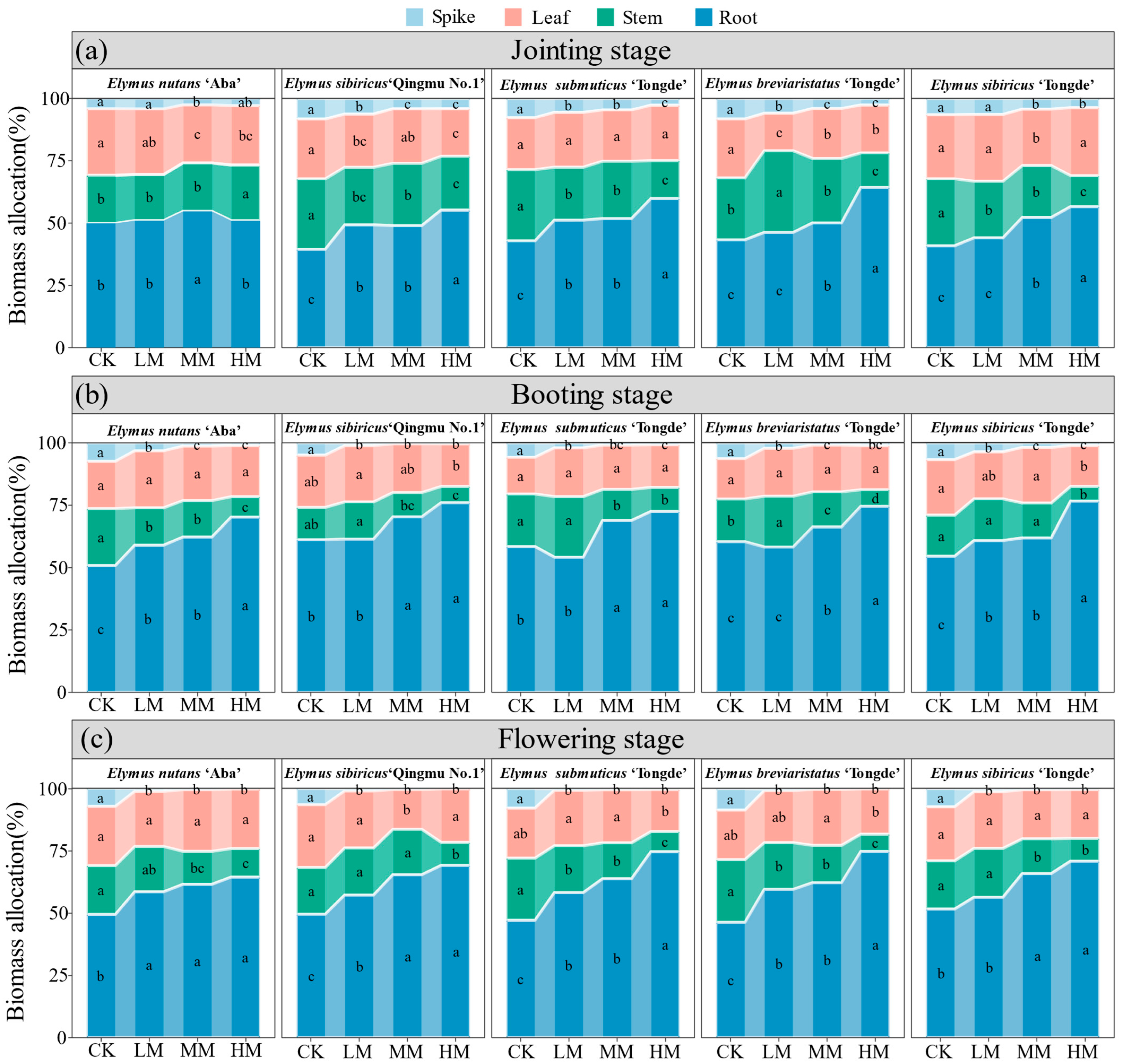
3.2. Effects of Mowing on the Component Biomass Allocation in Elymus Species
3.3. The Allometric Growth Relationships of Biomass Among Different Components of Elymus Species Under Mowing Treatments
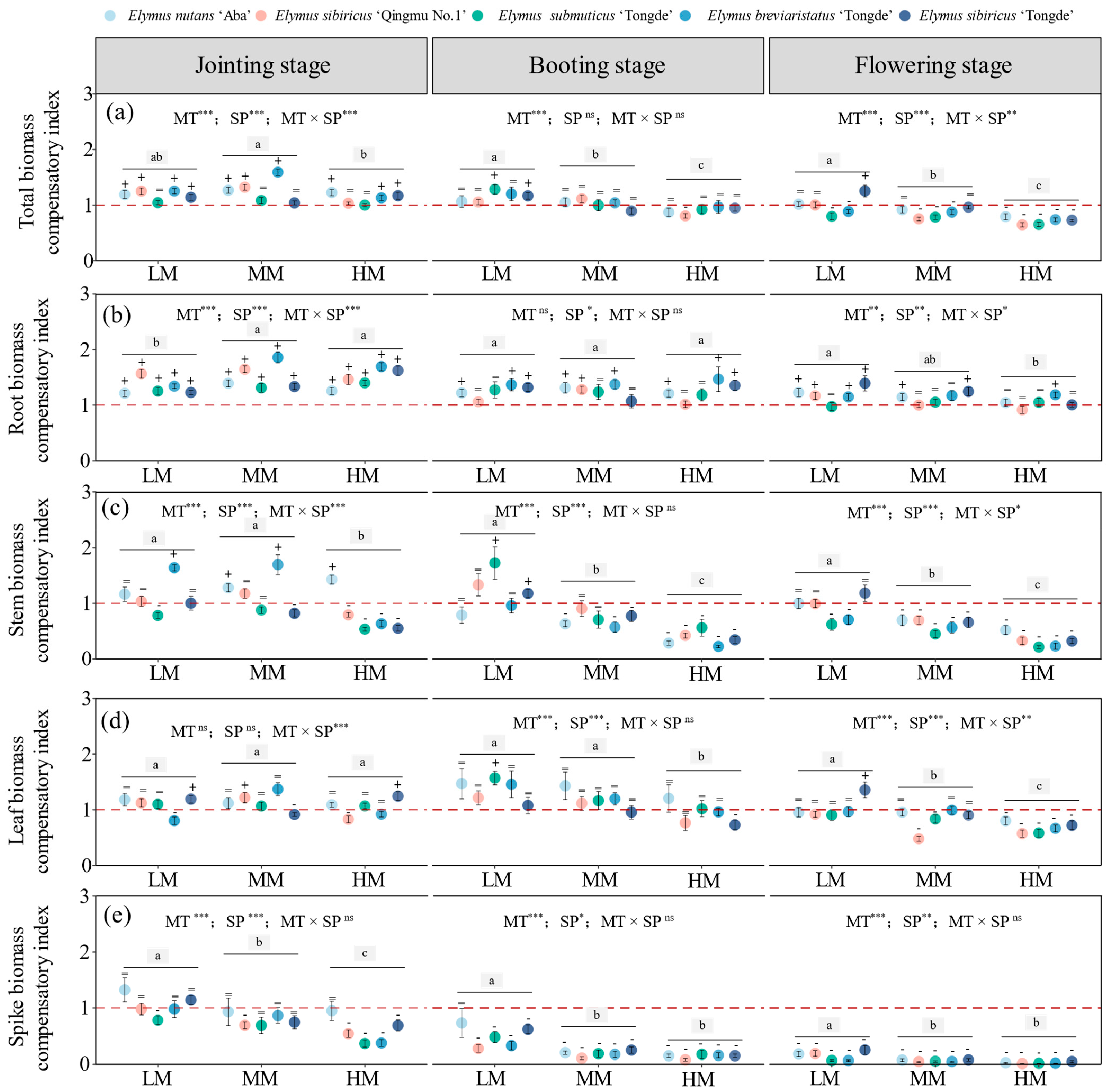
3.4. Effects of Mowing on the Compensatory Growth of Biomass Components in Elymus Species
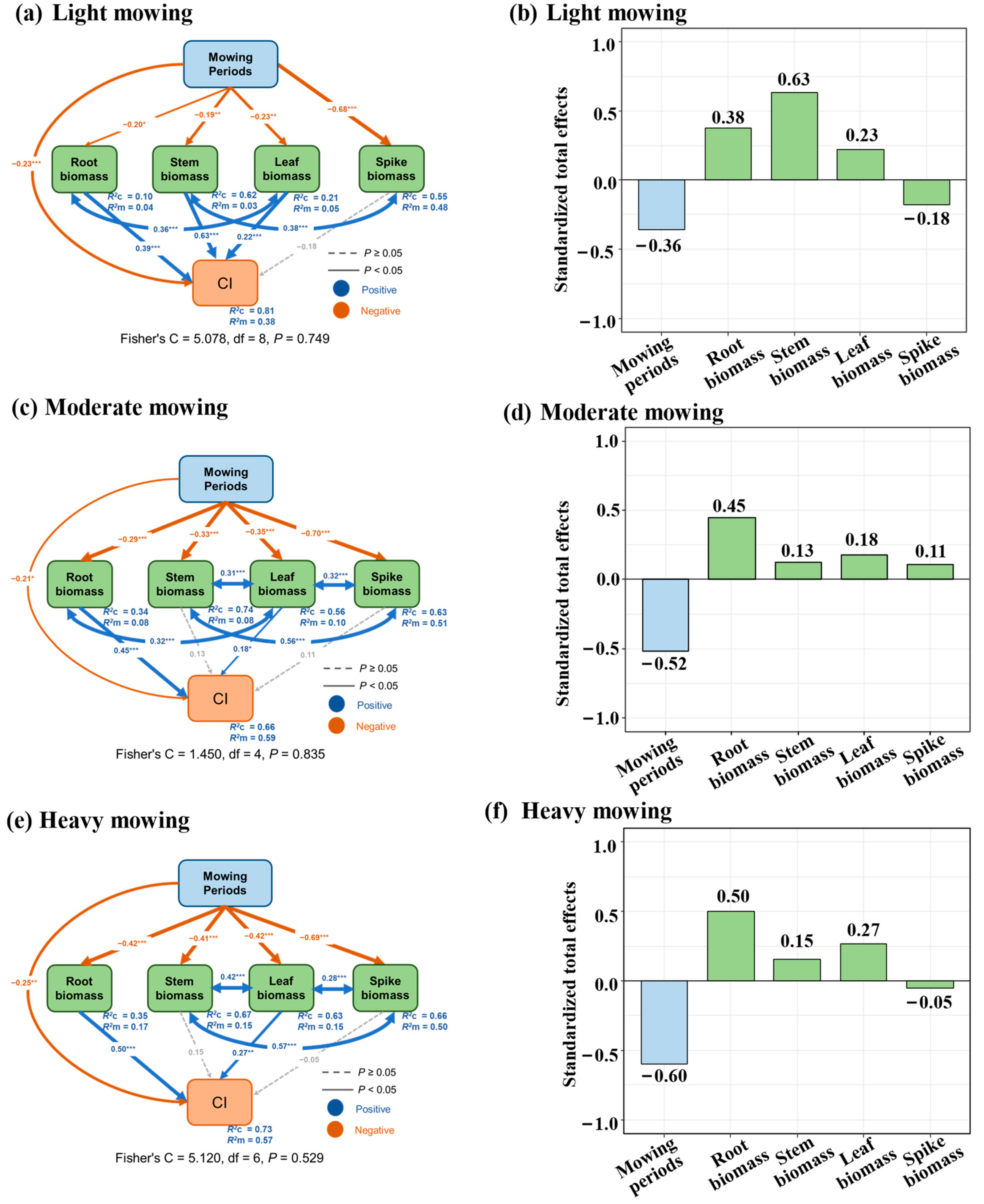
3.5. The Pathways Through Which Mowing Affects Compensatory Growth in Elymus Species
4. Discussion
4.1. Effects of Mowing on the Biomass Accumulation in Various Plant Organs
4.2. Effects of Mowing on the Biomass Allocation Among Plant Organs
4.3. Mowing Timing and Organ Biomass Jointly Drive the Total Biomass Compensation in Elymus Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, G.; Wang, Y.; Hautier, Y.; Song, X.; Zhou, Y.; Zhang, M.; Wang, L. Moderate grassland use counteracts the combined negative impact of nitrogen deposition and plant diversity decline on carbon exchange. Catena 2024, 247, 108558. [Google Scholar] [CrossRef]
- Wan, Z.; Yang, J.; Gu, R.; Liang, Y.; Yan, Y.; Gao, Q.; Yang, J. Influence of different mowing systems on community characteristics and the compensatory growth of important species of the Stipa grandis steppe in Inner Mongolia. Sustainability 2016, 8, 1121. [Google Scholar] [CrossRef]
- Peng, F.; Lai, C.; Li, C.; Ji, C.; Zhang, P.; Sun, J.; Chen, X.; You, Q.; Xue, X. Plasticity in over-Compensatory growth along an alpine meadow degradation gradient on the Qinghai-Tibetan Plateau. J. Environ. Manag. 2023, 325, 116499. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zheng, Q.; Zhang, Y.; Ganjurjav, H.; Yue, H.; Wang, X.; Wu, K.; Liang, K.; Zeng, H.; Wu, H. Short-term robust plant overcompensatory growth was observed in a degraded alpine meadow on the southeastern Qinghai-Tibetan Plateau. Sci. Total Environ. 2024, 918, 170607. [Google Scholar] [CrossRef]
- Guo, M.; Guo, T.; Zhou, J.; Liang, J.; Yang, G.; Zhang, Y. Restored legume acts as a “nurse” to facilitate plant compensatory growth and biomass production in mown grasslands. Agron. Sustain. Dev. 2024, 44, 60. [Google Scholar] [CrossRef]
- Tozer, K.; Douglas, G.; Dodd, M.; Müller, K. Vegetation options for increasing resilience in pastoral hill country. Front. Sustain. Food Syst. 2021, 5, 550334. [Google Scholar] [CrossRef]
- Li, X.; Png, G.K.; Sun, S.; Shi, H.; Jin, K.; Li, Y. Positive microbial legacy and short-term clonal plasticity aid grazing tolerance of a widespread grass species. Plant Soil 2022, 473, 291–303. [Google Scholar] [CrossRef]
- Fill, J.M.; Crandall, R.M. High survival promotes bunchgrass persistence in old-growth savannas under different fire regimes. Ecosphere 2024, 15, e70092. [Google Scholar] [CrossRef]
- Eklöf, J.S.; McMahon, K.; Lavery, P.S. Effects of multiple disturbances in seagrass meadows: Shading decreases resilience to grazing. Mar. Freshw. Res. 2009, 60, 1317–1327. [Google Scholar] [CrossRef]
- Shahba, M.; Alshammary, S.; Abbas, M. Effects of salinity on seashore paspalum cultivars at different mowing heights. Crop Sci. 2012, 52, 1358–1370. [Google Scholar] [CrossRef]
- Brys, R.; Shefferson, R.P.; Jacquemyn, H. Impact of herbivory on flowering behaviour and life history trade-offs in a polycarpic herb: A 10-year experiment. Oecologia 2011, 166, 293–303. [Google Scholar] [CrossRef]
- Schroeder, J.; Piersma, T.; Groen, N.M.; Kentie, R.; Lourenco, P.M.; Schekkerman, H.; Both, C. Reproductive timing and investment in relation to spring warming and advancing agricultural schedules. J. Ornithol. 2012, 153, 327–336. [Google Scholar] [CrossRef]
- Bricca, A.; Catorci, A.; Tardella, F.M. Intra-specific multi-trait approach reveals scarce ability in the variation of resource exploitation strategies for a dominant tall-grass under intense disturbance. Flora 2020, 270, 151665. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, L.; Zhou, H.; Ren, L.; Li, W.; Bai, W.; Zhang, W. Carbon allocation patterns in forbs and grasses differ in responses to mowing and nitrogen fertilization in a temperate grassland. Ecol. Indic. 2022, 135, 108588. [Google Scholar] [CrossRef]
- Gastal, F.; Lemaire, G. Defoliation, shoot plasticity, sward structure and herbage utilization in pasture: Review of the underlying ecophysiological Processes. Agriculture 2015, 5, 1146–1171. [Google Scholar] [CrossRef]
- Li, X.; Wu, Z.; Liu, Z.; Hou, X.; Badgery, W.; Guo, H.; Zhao, Q.; Hu, N.; Duan, J.; Ren, W. Contrasting effects of long-term grazing and clipping on plant morphological plasticity: Evidence from a rhizomatous grass. PLoS ONE 2015, 10, e0141055. [Google Scholar] [CrossRef] [PubMed]
- Dar, B.A.; Assaeed, A.M.; Al-Rowaily, S.L.; Al-Doss, A.A.; Habib, M.M.; Malik, J.A.; Abd-ElGawad, A.M. Effect of simulated grazing on morphological plasticity and resource allocation of aeluropus lagopoides. Agronomy 2024, 14, 144. [Google Scholar] [CrossRef]
- Turner, C.L.; Seastedt, T.R.; Dyer, M.I. Maximization of aboveground grassland production: The role of defoliation frequency, intensity, and history. Ecol. Appl. 1993, 3, 175–186. [Google Scholar] [CrossRef]
- Zhao, C.; Li, G.; Li, Q.; Zhou, D. Effects of mowing frequency on biomass allocation and yield of Leymus Chinensis. Rangel. Ecol. Manag. 2022, 83, 102–111. [Google Scholar] [CrossRef]
- Qu, L.; Liu, J.; Yang, J.; Bai, L.; Huang, Y.; Lu, N.; Yu, H.; Wang, Z.; Li, Z. Soil saline-alkali heterogeneity is an important factor driving the spatial expansion of clonal plant in grassland. Front. Environ. Sci. 2023, 10, 1106825. [Google Scholar] [CrossRef]
- Tian, T.; Guo, J.; Yang, Z.; Yao, Z.; Liu, X.; Wang, Z. Effects of different grazing treatments on the root system of Stipa krylovii Steppe. Sustainability 2024, 16, 3975. [Google Scholar] [CrossRef]
- Ferraro, D.O.; Oesterheld, M. Effect of defoliation on grass growth. A quantitative review. Oikos 2002, 98, 125–133. [Google Scholar] [CrossRef]
- Belsky, A.J. Does herbivory benefit plants? A review of the evidence. Am. Nat. 1986, 127, 870–892. [Google Scholar] [CrossRef]
- Gao, Y.Z.; Wang, S.P.; Han, X.G.; Chen, Q.S.; Zhou, Z.Y.; Patton, B.D. Defoliation, nitrogen, and competition: Effects on plant growth and resource allocation of Cleistogenes squarrosa and Artemisia frigida. J. Plant Nutr. Soil Sci. 2007, 170, 115–122. [Google Scholar] [CrossRef]
- Liu, Y.; Cordero, I.; Bardgett, R.D. Defoliation and fertilisation differentially moderate root trait effects on soil abiotic and biotic properties. J. Ecol. 2023, 111, 2733–2749. [Google Scholar] [CrossRef]
- Tong, Y.; Dong, Q.-M.; Yu, Y.; Cao, Q.; Yang, X.; Liu, W.; Yang, Z.; Zhang, X.; Yuzhen, L.; Zhang, C. Nitrogen Application Increases the Productivity of Perennial Alpine Cultivated Grassland by Improving Soil Physicochemical Properties and Microbial Community Characteristics. Plant Soil 2024, 505, 559–579. [Google Scholar] [CrossRef]
- Yuan, J.; Li, H.; Yang, Y. The compensatory tillering in the forage grass Hordeum brevisubulatum after simulated grazing of different severity. Front. Plant Sci. 2020, 11, 792. [Google Scholar] [CrossRef]
- Niklas, K.; Enquist, B. Invariant scaling relationships for interspecific plant biomass production rates and body size. Proc. Natl. Acad. Sci. USA 2001, 98, 2922–2927. [Google Scholar] [CrossRef]
- Warton, D.I.; Duursma, R.A.; Falster, D.S.; Taskinen, S. Smatr 3-an R package for estimation and inference about allometric lines. Methods Ecol. Evol. 2012, 3, 257–259. [Google Scholar] [CrossRef]
- Lefcheck, J.S. piecewiseSEM: Piecewise structural equation modelling in r for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Wilschut, R.A.; van Kleunen, M. Drought alters plant-soil feedback effects on biomass allocation but not on plant performance. Plant Soil 2021, 462, 285–296. [Google Scholar] [CrossRef]
- Zhao, W.; Chen, S.P.; Lin, G.H. Compensatory growth responses to clipping defoliation in Leymus Chinensis (Poaceae) under nutrient addition and water deficiency conditions. Plant Ecol. 2008, 196, 85–99. [Google Scholar] [CrossRef]
- Angassa, A.; Oba, G. Effects of grazing pressure, age of enclosures and seasonality on bush cover dynamics and vegetation composition in southern Ethiopia. J. Arid. Environ. 2010, 74, 111–120. [Google Scholar] [CrossRef]
- McNaughton, S.J. Compensatory plant growth as a response to herbivory. Oikos 1983, 40, 329–336. [Google Scholar] [CrossRef]
- Hamilton, E.W., III; Frank, D.A. Can plants stimulate soil microbes and their own nutrient supply? evidence from a grazing. Ecology 2001, 82, 2397–2402. [Google Scholar] [CrossRef]
- Morris, C.D. Buried but unsafe–defoliation depletes the underground storage organ (USO) of the mesic grassland geophyte, Hypoxis Hemerocallidea. S. Afr. J. Bot. 2021, 141, 265–272. [Google Scholar] [CrossRef]
- Xu, T.; Johnson, D.; Bardgett, R.D. Defoliation modifies the impact of drought on the transfer of recent plant-assimilated carbon to soil and arbuscular mycorrhizal fungi. Plant Soil 2024, 507, 693–711. [Google Scholar] [CrossRef]
- Merino, V.M.; Aguilar, R.I.; Rivero, M.J.; Ordóñez, I.P.; Piña, L.F.; López-Belchí, M.D.; Schoebitz, M.I.; Noriega, F.A.; Pérez, C.I.; Cooke, A.S.; et al. Distribution of non-structural carbohydrates and root structure of Plantago lanceolata L. under different defoliation frequencies and intensities. Plants 2024, 13, 2773. [Google Scholar] [CrossRef]
- Strauss, S.Y.; Agrawal, A.A. The ecology and evolution of plant tolerance to herbivory. Trends Ecol. Evol. 1999, 14, 179–185. [Google Scholar] [CrossRef]
- Meuriot, F.; Morvan-Bertrand, A.; Noiraud-Romy, N.; Decau, M.L.; Escobar Gutiérrez, A.J.; Gastal, F.; Prud’homme, M.P. Short-term effects of defoliation intensity on sugar remobilization and N fluxes in ryegrass. J. Exp. Bot. 2018, 69, 3975–3986. [Google Scholar] [CrossRef]
- Medeiros, C.; Schmitt, D.; Duchini, P.; Miqueloto, T.; Sbrissia, A. Defoliation intensity and leaf area index recovery in defoliated swards: Implications for forage accumulation. Sci. Agric. 2020, 78, e20190095. [Google Scholar]
- Albacete, A.A.; Martínez-Andújar, C.; Pérez-Alfocea, F. Hormonal and metabolic regulation of source–sink relations under salinity and drought: From plant survival to crop yield stability. Biotechnol. Adv. 2014, 32, 12–30. [Google Scholar] [CrossRef] [PubMed]
- Lambers, H.; Oliveira, R.S. Plant Physiological Ecology, 3rd ed.; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Iqbal, N.; Masood, A.; Khan, N.A. Analyzing the significance of defoliation in growth, photosynthetic compensation and source-Sink Relations. Photosynthetica 2012, 50, 161–170. [Google Scholar] [CrossRef]
- Deslauriers, A.; Caron, L.; Rossi, S. Carbon allocation during defoliation: Testing a defense-growth trade-off in balsam fir. Front. Plant Sci. 2015, 6, 338. [Google Scholar] [CrossRef]
- Cavagnaro, R.A. Species-specific trade-offs between regrowth and mycorrhizas in the face of defoliation and phosphorus addition. Fungal Ecol. 2021, 51, 101058. [Google Scholar] [CrossRef]
- Guo, Y.J.; Han, L.; Li, G.-D.; Han, J.G.; Wang, G.L.; Li, Z.Y.; Wilson, B. The effects of defoliation on plant community, root biomass and nutrient allocation and soil chemical properties on semi-arid steppes in northern china. J. Arid. Environ. 2012, 78, 128–134. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, X.; Tian, D.; Cong, R.; Zhang, X.; Pan, Q.; Shi, Z. Resource reallocation of two grass species during regrowth after defoliation. Front. Plant Sci. 2018, 9, 1767. [Google Scholar] [CrossRef]
- Zhang, Z.; Gong, J.; Li, X.; Ding, Y.; Wang, B.; Shi, J.; Liu, M.; Yang, B. Underlying mechanism on source-sink carbon balance of grazed perennial grass during regrowth: Insights into optimal grazing regimes of restoration of degraded grasslands in a temperate steppe. J. Environ. Manag. 2021, 277, 111439. [Google Scholar] [CrossRef]
- Zhu, Y.; Delgado-Baquerizo, M.; Shan, D.; Yang, X.; Eldridge, D.J. Grazing impacts on ecosystem functions exceed those from mowing. Plant Soil 2021, 464, 579–591. [Google Scholar] [CrossRef]
- Qiao, J. Grazing intensity changes root traits and resource utilization strategies of Stipa breviflora in a desert steppe. Plant Soil 2024, 503, 475–488. [Google Scholar] [CrossRef]
- Clarke, P.J.; Lawes, M.J.; Midgley, J.J.; Lamont, B.B.; Ojeda, F.; Burrows, G.E.; Enright, N.J.; Knox, K.J.E. Resprouting as a key functional trait: How buds, protection and resources drive persistence after fire. New Phytol. 2013, 197, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Fang, J.; Ma, W.; Guo, D.; Mohammat, A. Large-scale pattern of biomass partitioning across china’s grasslands. Glob. Ecol. Biogeogr. 2010, 19, 268–277. [Google Scholar] [CrossRef]
- Yuan, J.; Wang, P.; Yang, Y. Effects of simulated herbivory on the vegetative reproduction and compensatory growth of Hordeum Brevisubulatum at different ontogenic stages. Int. J. Environ. Res. Public Health 2019, 16, 1663. [Google Scholar] [CrossRef]
- Li, H.; Cai, J.; Jiang, D.; Liu, F.; Dai, T.; Cao, W. Carbohydrates accumulation and remobilization in wheat plants as influenced by combined waterlogging and shading stress during grain filling. J. Agron. Crop Sci. 2013, 199, 38–48. [Google Scholar] [CrossRef]
- Zhang, T.; Li, F.Y.; Wang, H.; Wu, L.; Shi, C.; Li, Y.; Hu, J. Effects of defoliation timing on plant nutrient resorption and hay production in a semi-arid steppe. J. Plant Ecol. 2021, 14, 44–57. [Google Scholar] [CrossRef]
- Obeso, J.R. The costs of reproduction in plants. New Phytol. 2002, 155, 321–348. [Google Scholar] [CrossRef]
- Le Roncé, I.; Toïgo, M.; Dardevet, E.; Venner, S.; Limousin, J.M.; Chuine, I. Resource manipulation through experimental defoliation has legacy effects on allocation to reproductive and vegetative organs in quercus Ilex. Ann. Bot. 2020, 126, 1165–1179. [Google Scholar] [CrossRef]
- Li, S.; Hussain, S.B.; Vincent, C. Response of carbon fixation, allocation, and growth to source-sink manipulation by defoliation in vegetative citrus trees. Physiol. Plant. 2024, 176, e14304. [Google Scholar] [CrossRef]
- Ma, Z.; Chang, S.X.; Bork, E.W.; Steinaker, D.F.; Wilson, S.D.; White, S.R.; Cahill, J.F. Climate change and defoliation interact to affect root length across northern temperate grasslands. Funct. Ecol. 2020, 34, 2611–2621. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Zhang, C.; Cao, Q.; Yu, Y.; Liu, Y.; Tong, Y.; Zhang, X.; Li, C.; Dong, Q. Regulatory Effects of Mowing on Biomass Allocation and Compensation Growth Mechanisms in Elymus Species. Agriculture 2025, 15, 820. https://doi.org/10.3390/agriculture15080820
Yang Z, Zhang C, Cao Q, Yu Y, Liu Y, Tong Y, Zhang X, Li C, Dong Q. Regulatory Effects of Mowing on Biomass Allocation and Compensation Growth Mechanisms in Elymus Species. Agriculture. 2025; 15(8):820. https://doi.org/10.3390/agriculture15080820
Chicago/Turabian StyleYang, Zengzeng, Chunping Zhang, Quan Cao, Yang Yu, Yuzhen Liu, Yongshang Tong, Xiaofang Zhang, Caidi Li, and Quanmin Dong. 2025. "Regulatory Effects of Mowing on Biomass Allocation and Compensation Growth Mechanisms in Elymus Species" Agriculture 15, no. 8: 820. https://doi.org/10.3390/agriculture15080820
APA StyleYang, Z., Zhang, C., Cao, Q., Yu, Y., Liu, Y., Tong, Y., Zhang, X., Li, C., & Dong, Q. (2025). Regulatory Effects of Mowing on Biomass Allocation and Compensation Growth Mechanisms in Elymus Species. Agriculture, 15(8), 820. https://doi.org/10.3390/agriculture15080820