

Evaluation of Genotype × Environment Interactions in Quinoa Genotypes (Chenopodium quinoa Willd.)


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Conditions
2.2. Statistical Analysis
3. Results
3.1. Analysis of Variance
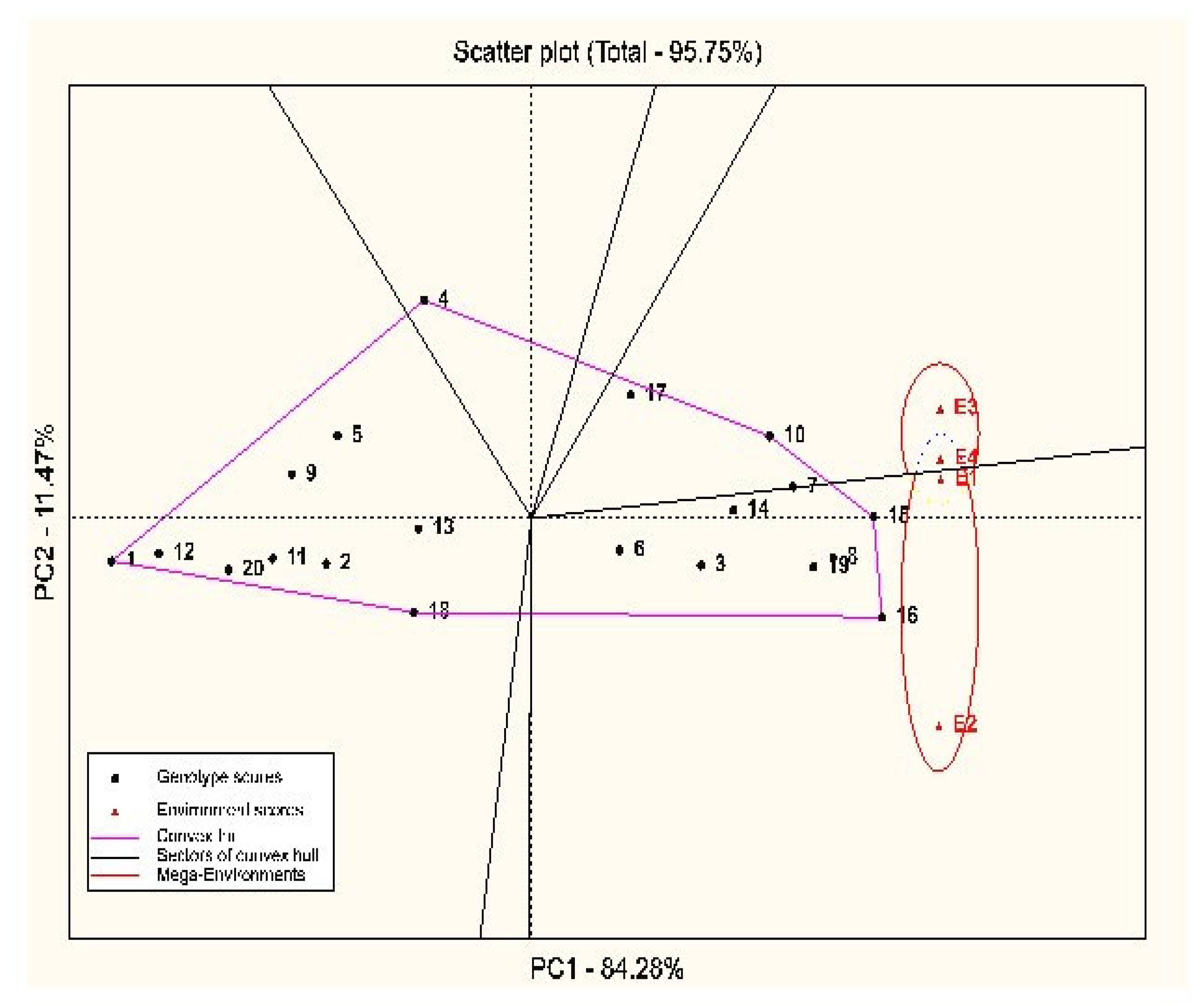
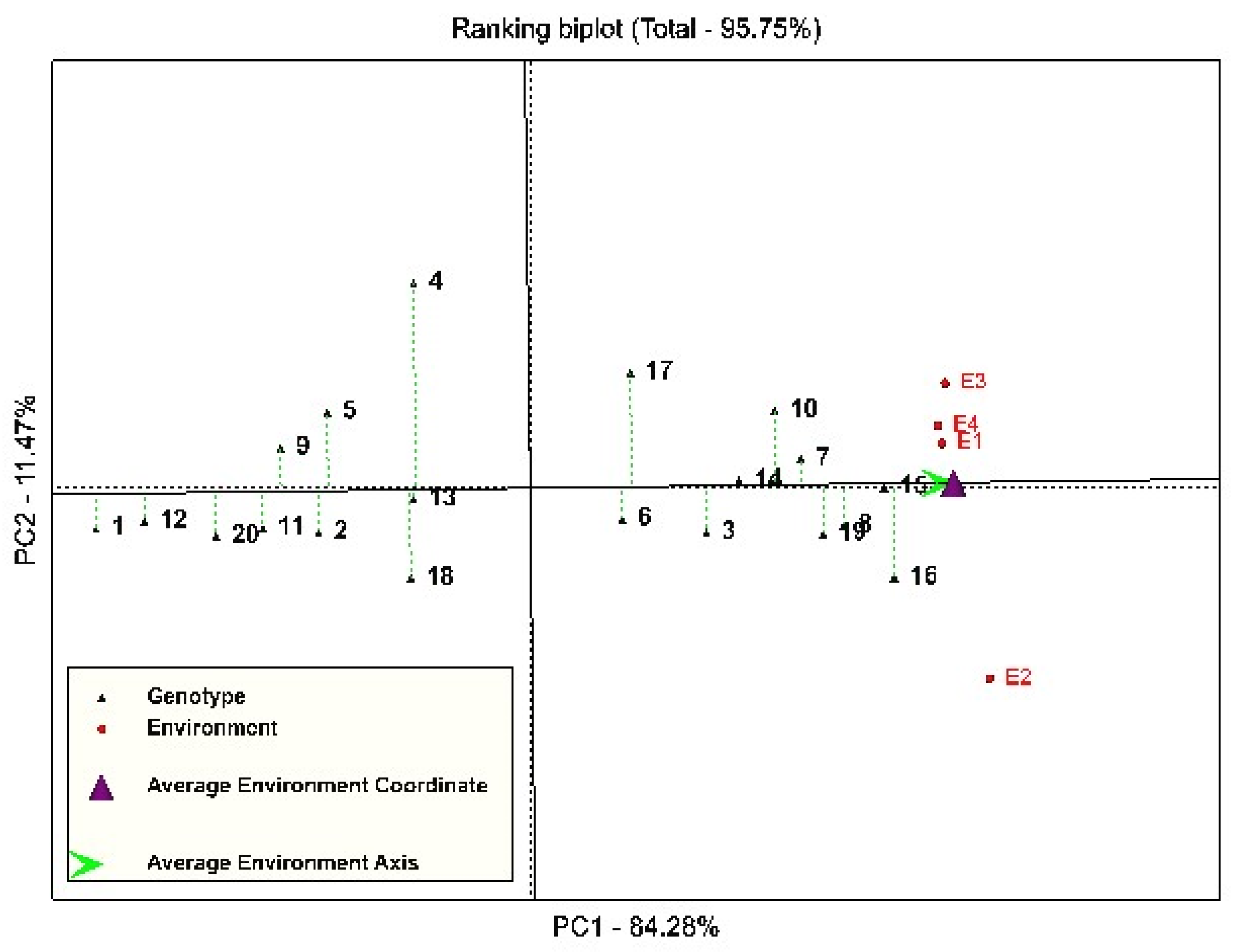
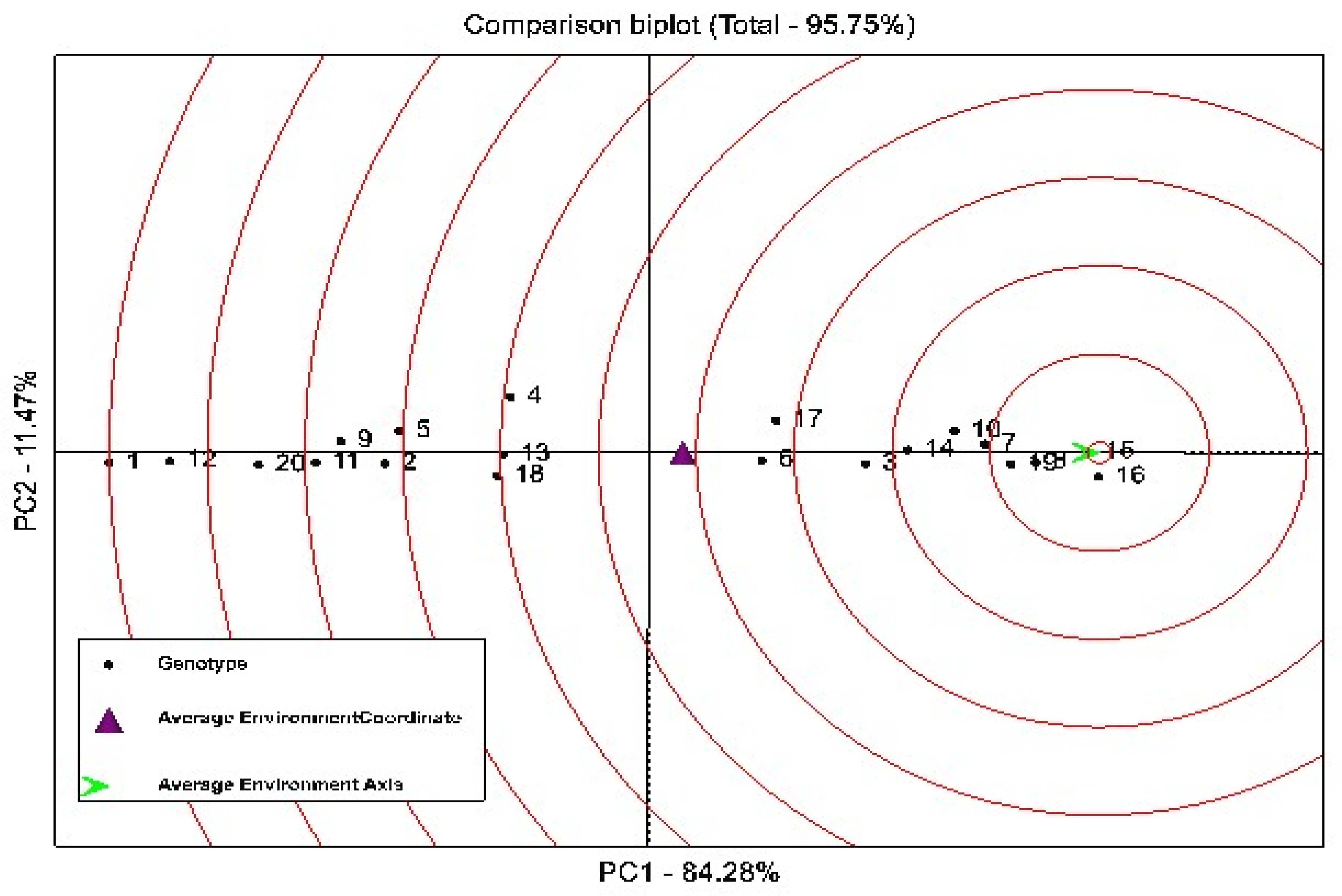
3.2. Stability Analysis
3.3. Morphological Traits
4. Discussion
4.1. Genetic Diversity
4.2. Identifying Stable Genotypes
4.3. Comparison of Morphological Traits
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Maughan, P.J.; Kolano, B.A.; Maluszynska, J.; Coles, N.D.; Bonifacio, A.; Rojas, J.; Coleman, C.E.; Stevens, M.R.; Fairbanks, D.J.; Parkinson, S.E.; et al. Molecular and cytological characterization of ribosomal RNA genes in Chenopodium quinoa and Chenopodium berlandieri. Genome 2006, 49, 825–839. [Google Scholar] [CrossRef]
- Anchico-Jojoa, W.; Peixoto, J.R.; de Oliveira Júnior, A.A. Agronomic characterization and interaction of genotype by environment of quinoa under conditions of Brazil and Colombia. Hortic. Bras. 2023, 41, e2629. [Google Scholar] [CrossRef]
- De Bock, P.; Van Bockstaele, F.; Muylle, H.; Quataert, P.; Vermeir, P.; Eeckhout, M.; Cnops, G. Yield and nutritional characterization of thirteen quinoa (Chenopodium quinoa Willd.) varieties grown in North-West Europe—Part I. Plants 2021, 10, 2689. [Google Scholar] [CrossRef]
- Manjarres-Hernández, E.H.; Arias-Moreno, D.M.; Morillo-Coronado, A.C.; Ojeda-Pérez, Z.Z.; Cárdenas-Chaparro, A. Phenotypic characterization of quinoa (Chenopodium quinoa Willd.) for the selection of promising materials for breeding programs. Plants 2021, 10, 1339. [Google Scholar] [CrossRef]
- Taaime, N.; El Mejahed, K.; Moussafir, M.; Bouabid, R.; Oukarroum, A.; Choukr-Allah, R.; El Gharous, M. Early sowing of quinoa cultivars, benefits from rainy season and enhances quinoa development, growth, and yield under arid condition in Morocco. Sustainability 2022, 14, 4010. [Google Scholar] [CrossRef]
- FAO FAOSTAT. Quinoa 2013 International Year. Food and Agriculture Organization of the United Nations (FAO). Available online: https://www.fao.org/quinoa-2013 (accessed on 4 October 2013).
- Ando, H.; Chen, Y.C.; Tang, H.; Shimizu, M.; Watanabe, K.; Mitsunaga, T. Food components in fractions of quinoa seed. Food Sci. Technol. Res. 2002, 8, 80–84. [Google Scholar] [CrossRef]
- Pathan, S.; Eivazi, F.; Valliyodan, B.; Paul, K.; Ndunguru, G.; Clark, K. Nutritional composition of the green leaves of quinoa (Chenopodium quinoa Willd.). J. Food Res. 2019, 8, 55–65. [Google Scholar] [CrossRef]
- Abdelshafy, A.M.; Rashwan, A.K.; Osman, A.I. Potential food applications and biological activities of fermented quinoa: A review. Trends Food Sci. Technol. 2024, 144, 104339. [Google Scholar] [CrossRef]
- Alandia, G.; Odone, A.; Rodriguez, J.P.; Bazile, D.; Condori, B. Quinoa—Evolution and future perspectives. In The Quinoa Genome; Springer Nature: Cham, Switzerland, 2021; pp. 179–195. [Google Scholar]
- Cui, H.; Yao, Q.; Xing, B.; Zhou, B.; Shah, S.S.; Qin, P. The performance of agronomic and quality traits of quinoa under different altitudes in Northwest of China. Agronomy 2024, 14, 1194. [Google Scholar] [CrossRef]
- FAO FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 1 September 2024).
- Baker, R.J. Test for cross over genotype-environment interactions. Can. J. Plant Sci. 1988, 68, 405–441. [Google Scholar] [CrossRef]
- Maliro, M.F.; Guwela, V.F.; Nyaika, J.; Murphy, K.M. Preliminary studies of the performance of quinoa (Chenopodium quinoa Willd.) genotypes under irrigated and rainfed conditions of central Malawi. Front. Plant Sci. 2017, 8, 227. [Google Scholar] [CrossRef]
- Mustafa, T.A.N.; Temel, S. Studies on the adaptation of quinoa (Chenopodium quinoa Willd.) to Eastern Anatolia Region of Turkey. Agrofor 2017, 2, 33–39. [Google Scholar]
- Thiam, E.; Allaoui, A.; Benlhabib, O. Quinoa productivity and stability evaluation through varietal and environmental interaction. Plants 2021, 10, 714. [Google Scholar] [CrossRef] [PubMed]
- Wricke, G. Übereine methode zur erfassung der ökologischen streubreite in feldversuchen. Z. Pflanzenzücht. 1962, 47, 92–96. [Google Scholar]
- Finlay, K.W.; Wilkinson, G.N. Adaptation in a plant breeding programme. Aust. J. Agric. Res. 1963, 14, 742–754. [Google Scholar] [CrossRef]
- Eberhart, S.A.T.; Russell, W.A. Stability parameters for comparing varieties. Crop Sci. 1966, 6, 36–40. [Google Scholar] [CrossRef]
- Roemer, J. Sinde die ertagdreichen sorten ertagissicherer? Mitt DLG 1917, 32, 87–89. [Google Scholar]
- Shukla, G.K. Some statistical aspects of partitioning genotype-environmental components of variability. Heredity 1972, 28, 237–245. [Google Scholar] [CrossRef]
- Francis, T.R.; Kannenberg, L.W. Yield stability studies in short-season maize. I. A descriptive method for grouping genotypes. Can. J. Plant Sci. 1978, 58, 1029–1034. [Google Scholar] [CrossRef]
- Lin, C.S.; Binns, M.R. A superiority measure of cultivar performance for cultivar x location data. Can. J. Plant Sci. 1988, 68, 193–198. [Google Scholar] [CrossRef]
- Yan, W.; Kang, M.S.; Ma, B.; Woods, S.; Cornelius, P.L. GGE biplot vs. AMMI analysis of genotype-by-environment data. Crop Sci. 2007, 4, 643–653. [Google Scholar] [CrossRef]
- SAS Institute. SAS® 9.4 System Options: Reference; SAS Institute Inc.: Cary, NC, USA, 2019. [Google Scholar]
- Pour-Aboughadareh, A.; Yousefian, M.; Moradkhani, H.; Poczai, P.; Siddique, K.H. STABILITYSOFT: A new online program to calculate parametric and non-parametric stability statistics for crop traits. Appl. Plant Sci. 2019, 7, e01211. [Google Scholar] [CrossRef]
- Payne, R.W.; Baird, D.B.; Cherry, M.; Gilmour, A.R.; Harding, S.A.; Lane, P.W.; Morgan, G.W.; Murray, D.A.; Soutar, D.M.; Thompson, R.; et al. GenStat Release 6.1 Reference Manual; Directives; Hemel Hempstead VSN International: Hemel Hempstead, UK, 2002; 404p. [Google Scholar]
- Miranda, M.; Vega-Gálvez, A.; Quispe-Fuentes, I.; Rodríguez, M.J.; Maureira, H.; Martínez, E.A. Nutritional aspects of six quinoa (Chenopodium quinoa Willd.) ecotypes from three geographical areas of Chile. Chil. J. Agric. Res. 2012, 72, 175–181. [Google Scholar] [CrossRef]
- Al-Naggar, A.M.M.; Younis, A.E.S.M.; Atta, M.M.; El-Moneim, M.L.A.; Al-Metwally, M.S. Stability of Chenopodium quinoa genotypes under different nitrogen fertilizer source and level using AMMI and GGE-biplot models. Plant Cell Biotechnol. Mol. Biol. 2022, 23, 1–20. [Google Scholar] [CrossRef]
- Bertero, H.D.; de la Vega, A.J.; Correa, G.; Jacobsen, S.E.; Mujica, A. Genotype and genotype-by-environment interaction effects for grain yield and grain size of quinoa (Chenopodium quinoa Willd.) as revealed by pattern analysis of international multi-environment trials. Field Crop. Res. 2004, 89, 299–318. [Google Scholar] [CrossRef]
- Curti, R.N.; de la Vega, A.J.; Andrade, A.J.; Bramardi, S.J.; Bertero, H.D. Multi-environmental evaluation for grain yield and its physiological determinants of quinoa genotypes across Northwest Argentina. Field Crop. Res. 2014, 166, 46–57. [Google Scholar] [CrossRef]
- Ali, M.; Elsadek, A.; Salem, E.M. Stability parameters and AMMI analysis of quinoa (Chenopodium quinoa Willd.). Egypt. J. Agron. 2018, 40, 59–74. [Google Scholar] [CrossRef]
- Vasconcelos, E.S.; Echer, M.M.; Kliemann, M.A.; Lang, M.J. Selection and recommend of quinoa (Chenopodium quinoa) genotypes based on the yield genotypic adaptability and stability. Rev. Ceres Viçosa 2019, 66, 117–123. [Google Scholar] [CrossRef]
- Khan, M.M.H.; Rafii, M.Y.; Ramlee, S.I.; Jusoh, M.; Mamun, M.A.; Halidu, J. DNA fingerprinting, fixation-index (Fst), and admixture mapping of selected bambara groundnut (Vigna subterranea [L.] verdc) accessions using ISSR markers system. Sci. Rep. 2021, 11, 14527. [Google Scholar]
- Yan, W.; Tinker, N.A. Biplot analysis of multi-environment trial data: Principles and applications. Can. J. Plant Sci. 2006, 86, 623–645. [Google Scholar] [CrossRef]
- Yan, W. GGE biplot: A windows application for graphical analysis of multi-environment trial data and other types of two-way data. Agron. J. 2001, 93, 1111–1118. [Google Scholar] [CrossRef]
- Hashim, N.; Rafii, M.Y.; Oladosu, Y.; Ismail, M.R.; Ramli, A.; Arolu, F.; Chukwu, S. Integrating multivariate and univariate statistical models to investigate genotype–environment interaction of advanced fragrant rice genotypes under rainfed condition. Sustainability 2021, 13, 4555. [Google Scholar] [CrossRef]
- Bhargava, A.; Shukla, S.; Ohri, D. Genetic variability and interrelationship among various morphological and quality traits in quinoa (Chenopodium quinoa Willd.). Field Crops Res. 2007, 101, 104–116. [Google Scholar] [CrossRef]
- Jorfi, A.; Alavifazel, M.; Gilani, A.; Ardakani, M.R.; Lak, S. Yield and morpho-physiological performance of quinoa (Chenopodium quinoa) genotypes as affected by phosphorus and zinc. J. Plant Nutr. 2022, 45, 2432–2446. [Google Scholar] [CrossRef]
- Shah, S.S.; Shi, L.; Li, Z.; Ren, G.; Zhou, B.; Qin, P. Yield, agronomic and forage quality traits of different quinoa (Chenopodium quinoa Willd.) genotypes in Northeast China. Agronomy 2020, 10, 1908. [Google Scholar] [CrossRef]
- Mastebroek, H.D.; Van Loo, E.N.; Dolstra, O. Combining ability for seed yield traits of Chenopodium quinoa breeding lines. Euphytica 2002, 125, 427–432. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| No. | Genotype | Accession Number a | Origin | Seed Color |
|---|---|---|---|---|
| 1 | CHEN67 | D2190 | Peru | Brown |
| 2 | CHEN68 | D2191 | Peru | Golden-brown |
| 3 | CHEN71 | D2196 | Bolivia | Light brown |
| 4 | CHEN83 | D2194 | Peru | Bright-white |
| 5 | CHEN84 | D2195 | Peru | White |
| 6 | CHEN89 | D5078 | Peru | Bright |
| 7 | CHEN90 | D5079 | Peru | White |
| 8 | CHEN112 | D11899 | Peru | Golden |
| 9 | CHEN119 | D9319 | Bolivia | White |
| 10 | CHEN126 | D9339 | Bolivia | Whitish-yellow |
| 11 | CHEN128 | D9320 | Bolivia | Yellow |
| 12 | CHEN133 | D9361 | Bolivia | White |
| 13 | CHEN151 | D9382 | Bolivia | Bright |
| 14 | CHEN159 | D9376 | Bolivia | Whitish-yellow |
| 15 | CHEN167 | D9346 | Peru | Yellow |
| 16 | CHEN171 | D9350 | Bolivia | Bright-white |
| 17 | CHEN182 | D9392 | Peru | White |
| 18 | CHEN189 | D9400 | Peru | White |
| 19 | CHEN212 | D9426 | Peru | Golden |
| 20 | CHEN223 | D9442 | Peru | Brown |
| Location | Year | Spring Temperature (°C) | Summer Temperature (°C) | Spring Rainfall (mm) | Summer Rainfall (mm) | Total Rainfall (mm) | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Min | Max | Average | Min | Max | Average | |||||
| Kuhdasht | 2020 | 9.1 | 29 | 23.1 | 17.6 | 35.4 | 28.2 | 127.1 | 2.9 | 454.6 |
| 2021 | 9.3 | 29.2 | 22.2 | 18.1 | 36.5 | 28.9 | 142 | 1.7 | 507.2 | |
| Poldokhtar | 2020 | 19.2 | 33.8 | 27.5 | 24.2 | 38.2 | 33.3 | 103.5 | 1.4 | 368.8 |
| 2021 | 18.1 | 32.7 | 26.3 | 21.1 | 38.7 | 31.8 | 119.2 | 0.9 | 425.7 | |
| Location | Year | Soil type | EC (ds.m−1) | pH | N (%) | P (ppm) | K (mg.kg−1) | OC (%) | ||
| Kuhdasht | 2020 | Loam | 0.45 | 7.57 | 0.09 | 4.6 | 325 | 0.87 | ||
| 2021 | Loam | 0.51 | 7.76 | 0.1 | 3.4 | 260 | 1.07 | |||
| Poldokhter | 2020 | Loam | 0.46 | 7.5 | 0.1 | 13.2 | 295 | 0.91 | ||
| 2021 | Loam | 0.49 | 7.66 | 0.11 | 12.48 | 230 | 1.15 | |||
| Source of Variations | Df | Mean Squares | |||
|---|---|---|---|---|---|
| 2020 | 2021 | ||||
| Kuhdasht | Poldokhtar | Kuhdasht | Poldokhtar | ||
| Replication | 2 | 20,235.82 ** | 13,495.22 ** | 28,913.15 ns | 39,516.12 ** |
| Genotype | 19 | 2,327,079.39 ** | 2,359,481.80 ** | 2,161,112.64 ** | 2,015,355.29 ** |
| Error | 38 | 7033.19 | 45,278.57 | 3926.57 | 63,043.16 |
| CV (%) | - | 4.10 | 9.65 | 2.42 | 10.36 |
| Source of Variations | Df | Mean Squares |
|---|---|---|
| Year (Y) | 1 | 9,177,552.65 ** |
| Location (L) | 1 | 575,456.33 ** |
| Y × L | 1 | 4,129,126.75 ** |
| Replication/GY | 8 | 35,871.33 |
| Genotype (G) | 19 | 8,374,364.65 ** |
| G × Y | 19 | 204,816.03 ** |
| G × L | 19 | 137,420.84 ** |
| G × Y × L | 19 | 147,440.05 ** |
| Experimental error | 152 | 31,232.23 |
| Coefficient of variation | 7.21 | |
| Contribution of genotype to total variation (%) | 84.94 | |
| Contribution of environment to total variation (%) | 10.09 | |
| Contribution of GEI to total variation (%) | 4.97 |
| Genotype a | 2020 | 2021 | Total Mean b | ||
|---|---|---|---|---|---|
| Kuhdasht | Poldokhtar | Kuhdasht | Poldokhtar | ||
| 1 | 660.65 | 716.63 | 992.74 | 964.36 | 833.60 k |
| 2 | 1311.66 | 1511.66 | 1829.56 | 1687.53 | 1585.10 j |
| 3 | 2541.66 | 2731.43 | 3243.37 | 3058.85 | 2893.83 e |
| 4 | 1724.65 | 1929.48 | 1808.82 | 2201.42 | 1916.09 g |
| 5 | 1401.58 | 1600.52 | 1683.34 | 1789.23 | 1618.67 i |
| 6 | 2312.33 | 2510.73 | 2921.18 | 2698.12 | 2610.59 f |
| 7 | 2900.42 | 3160.61 | 3494.49 | 3306.70 | 3215.56 c |
| 8 | 3028.66 | 3213.38 | 3736.65 | 3446.64 | 3356.33 b |
| 9 | 1277.34 | 1415.33 | 1560.53 | 1590.09 | 1460.82 h |
| 10 | 2840.62 | 3040.53 | 3317.71 | 3321.29 | 3130.04 c |
| 11 | 1143.12 | 1343.34 | 1620.18 | 1480.52 | 1396.79 h |
| 12 | 773.33 | 853.33 | 1158.34 | 1206.23 | 997.81 k |
| 13 | 1662.21 | 1862.66 | 2129.95 | 1968.21 | 1905.76 j |
| 14 | 2725.44 | 2925.46 | 3293.36 | 3080.35 | 3006.15 d |
| 15 | 3217.25 | 3415.51 | 3836.63 | 3517.76 | 3496.79 a |
| 16 | 3309.39 | 3216.33 | 3968.86 | 3619.16 | 3528.44 a |
| 17 | 2460.76 | 2656.39 | 2738.28 | 2716.58 | 2643.00 f |
| 18 | 1611.65 | 1811.76 | 2236.61 | 1915.46 | 1893.87 h |
| 19 | 3012.43 | 3136.52 | 3670.76 | 3335.38 | 3288.77 b |
| 20 | 942.42 | 1035.28 | 1440.34 | 1543.31 | 1240.34 i |
| Mean | 2042.88 | 2204.34 | 2534.09 | 2422.36 | 2300.91 |
| Genotype a | ECV b | Finlay–Wilkinson | ||||||
|---|---|---|---|---|---|---|---|---|
| bi | t-Value (H0: b = 1) | |||||||
| 1 | 20.31 | 28,672.47 | 984.80 | 0.751 | 1.799 ns | 2777.67 ns | 4407.79 | 13,223.36 |
| 2 | 14.13 | 50,138.49 | 74,787.67 | 1.014 | 0.215 ns | 616.06 ns | 420.49 | 1261.46 |
| 3 | 10.92 | 99,935.76 | 149,900.20 | 1.435 | 6.752 * | 601.89 ns | 9544.64 | 28,633.92 |
| 4 | 10.85 | 43,249.54 | 20,258.99 | 0.398 | 0.992 ns | 53,378.61 ns | 53,096.10 | 159,288.30 |
| 5 | 10.13 | 26,910.14 | 28,750.04 | 0.645 | 1.340 ns | 10,184.93 ns | 12,877.73 | 38,633.19 |
| 6 | 9.97 | 67,686.23 | 101,453.33 | 1.170 | 1.350 ns | 2294.78 ns | 2924.05 | 8772.15 |
| 7 | 7.79 | 62,813.72 | 93,565.36 | 1.122 | 0.860 ns | 2922.22 ns | 2668.51 | 8005.53 |
| 8 | 9.11 | 93,533.84 | 138,915.02 | 1.365 | 1.941 ns | 5134.70 ns | 9344.24 | 28,032.72 |
| 9 | 9.87 | 20,795.69 | 27,684.41 | 0.625 | 2.677 ns | 2840.28 ns | 8680.75 | 26,042.26 |
| 10 | 7.46 | 54,524.19 | 76,610.26 | 1.036 | 0.222 ns | 3890.97 ns | 2659.46 | 7978.37 |
| 11 | 14.56 | 41,373.06 | 61,601.15 | 0.916 | 0.925 ns | 1189.29 ns | 1132.33 | 3396.99 |
| 12 | 21.69 | 46,824.84 | 68,192.78 | 0.928 | 0.310 ns | 7757.90 ns | 5420.92 | 16,262.77 |
| 13 | 10.29 | 38,445.12 | 57,480.38 | 0.875 | 1.022 ns | 2178.08 ns | 2210.18 | 6630.53 |
| 14 | 8.00 | 57,768.07 | 86,631.01 | 1.077 | 0.584 ns | 2528.35 ns | 1972.79 | 5918.36 |
| 15 | 7.40 | 66,893.34 | 98,521.66 | 1.115 | 0.437 ns | 10,118.25 ns | 7389.68 | 22,169.05 |
| 16 | 9.65 | 115,865.15 | 149,293.17 | 1.382 | 0.773 ns | 35,373.16 ns | 30,623.23 | 91,869.68 |
| 17 | 4.78 | 15,961.00 | 20,160.05 | 0.430 | 2.998 ns | 3562.20 ns | 16,993.57 | 50,980.71 |
| 18 | 13.78 | 68,109.01 | 100,332.17 | 1.126 | 0.474 ns | 10,234.73 ns | 7589.887 | 22,769.66 |
| 19 | 8.74 | 82,544.24 | 118,235.92 | 1.246 | 0.886 ns | 11,193.65 ns | 10,393.65 | 31,180.96 |
| 20 | 23.85 | 87,532.25 | 126,504.70 | 1.242 | 0.660 ns | 19,463.60 ns | 15,803.9 | 47,411.71 |
| Mean | 11.66 | 58478.8075 | 79,993.1535 | 0.9949 | 1.3607 | 9412.066 | 10,307.69 | 30,923.084 |
| Genotype a | Environment 1 | Environment 2 | Environment 3 | Environment 4 | AYR b | FYR c |
|---|---|---|---|---|---|---|
| 1 | 20 | 20 | 20 | 20 | 20 | 20 |
| 2 | 15 | 15 | 13 | 15 | 14.5 | 15 |
| 3 | 8 | 8 | 8 | 8 | 8 | 8 |
| 4 | 11 | 11 | 14 | 11 | 11.75 | 11 |
| 5 | 14 | 14 | 15 | 14 | 14.25 | 14 |
| 6 | 10 | 10 | 9 | 10 | 9.75 | 10 |
| 7 | 5 | 4 | 5 | 6 | 5 | 5 |
| 8 | 3 | 3 | 3 | 3 | 3 | 3 |
| 9 | 16 | 16 | 17 | 16 | 16.25 | 16 |
| 10 | 6 | 6 | 6 | 5 | 5.75 | 6 |
| 11 | 17 | 17 | 16 | 18 | 17 | 17 |
| 12 | 19 | 19 | 19 | 19 | 19 | 19 |
| 13 | 12 | 12 | 12 | 12 | 12 | 12 |
| 14 | 7 | 7 | 7 | 7 | 7 | 7 |
| 15 | 2 | 1 | 2 | 2 | 1.75 | 2 |
| 16 | 1 | 2 | 1 | 1 | 1.25 | 1 |
| 17 | 9 | 9 | 10 | 9 | 9.25 | 9 |
| 18 | 13 | 13 | 11 | 13 | 12.5 | 13 |
| 19 | 4 | 5 | 4 | 4 | 4.25 | 4 |
| 20 | 18 | 18 | 18 | 17 | 17.75 | 18 |
| Genotype a | CV | bi | ASR | FSR | FYR + FSR | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 4 | 18 | 5 | 13 | 7 | 8 | 8 | 9.00 | 8 | 28 |
| 2 | 9 | 16 | 9 | 1 | 2 | 1 | 1 | 5.57 | 1 | 16 |
| 3 | 19 | 14 | 20 | 18 | 1 | 14 | 14 | 14.29 | 17 | 25 |
| 4 | 7 | 13 | 2 | 20 | 20 | 20 | 20 | 14.57 | 18 | 29 |
| 5 | 3 | 11 | 4 | 14 | 15 | 16 | 16 | 11.29 | 13 | 27 |
| 6 | 14 | 10 | 15 | 10 | 5 | 7 | 7 | 9.71 | 9 | 19 |
| 7 | 12 | 4 | 12 | 7 | 9 | 6 | 6 | 8.00 | 6 | 11 |
| 8 | 18 | 7 | 18 | 15 | 12 | 13 | 13 | 13.71 | 15 | 18 |
| 9 | 2 | 9 | 3 | 16 | 8 | 12 | 12 | 8.86 | 7 | 23 |
| 10 | 10 | 3 | 10 | 2 | 11 | 5 | 5 | 6.57 | 5 | 11 |
| 11 | 6 | 17 | 7 | 5 | 3 | 2 | 2 | 6.00 | 2 | 19 |
| 12 | 8 | 19 | 8 | 3 | 13 | 9 | 9 | 9.86 | 12 | 31 |
| 13 | 5 | 12 | 6 | 8 | 4 | 4 | 4 | 6.14 | 3 | 15 |
| 14 | 11 | 5 | 11 | 4 | 6 | 3 | 3 | 6.14 | 3 | 10 |
| 15 | 13 | 2 | 13 | 6 | 14 | 10 | 10 | 9.71 | 9 | 11 |
| 16 | 20 | 8 | 19 | 17 | 19 | 19 | 19 | 17.29 | 20 | 21 |
| 17 | 1 | 1 | 1 | 19 | 10 | 18 | 18 | 9.71 | 9 | 18 |
| 18 | 15 | 15 | 14 | 9 | 16 | 11 | 11 | 13.00 | 14 | 27 |
| 19 | 16 | 6 | 16 | 12 | 17 | 15 | 15 | 13.86 | 16 | 20 |
| 20 | 17 | 20 | 17 | 11 | 18 | 17 | 17 | 16.71 | 19 | 37 |
| Genotype a | SD (mm) | PH (cm) | PL (cm) | NPP | NGP | TGW (g) | DM (Day) | HI (%) | SP (%) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 5.86 j | 109.75 i | 12.24 n | 11.23 h | 14.82 i | 2.2 m | 112.29 i | 31.21 kij | 1.41 e |
| 2 | 5.96 g | 115.25 h | 17.4 k | 13.91 ef | 20.44 h | 2.44 l | 116.34 h | 35.20 hi | 1.59 c |
| 3 | 6.56 e | 105.91 j | 29.86 f | 15.25 dc | 27.74 d | 2.88 f | 103.77 k | 48.98 dc | 0.21 1 m |
| 4 | 5.47 k | 119.41 e | 21.24 ij | 12.66 g | 19.86 h | 2.48 jlk | 120.18 f | 39.95 fg | 1.64 b |
| 5 | 5.34 l | 118.91 ef | 18.44 k | 12.75 g | 24.62 efg | 2.55 jhki | 119.04 fg | 34.21 hij | 1.02 h |
| 6 | 6.18 f | 116.91 hg | 27.93 g | 13.91 ef | 22.86 g | 2.58 hi | 117.79 gh | 45.91 de | 1.21 f |
| 7 | 7.12 b | 99.08 m | 32.16 cd | 16.08 bc | 34.19 c | 3.41 d | 99.28 m | 54.36 b | 0.27 l |
| 8 | 7.31 a | 99.58 m | 34.11 b | 17.33 a | 38.71 b | 3.67 c | 101.72 l | 58.21 ab | 0.07 n |
| 9 | 5.73 ij | 125.5 c | 16.17 l | 13.16 gf | 26.65 de | 2.73 g | 126.42 c | 37.79 hg | 1.36 e |
| 10 | 7.16 b | 117.33 fg | 31.15 de | 16.5 ab | 38.64 b | 3.67 c | 118.46 fg | 58.12 ab | 0.18 m |
| 11 | 5.82 hi | 124.41 c | 15.22 lm | 13.58 efg | 23.55 fg | 2.56 ihj | 124.59 d | 31.07 jk | 1.78 a |
| 12 | 5.38 kl | 129 b | 14.48 mn | 11.58 h | 23.00 fg | 2.48 kl | 129.75 b | 29.37 k | 0.59 j |
| 13 | 5.91 hg | 116.25 hg | 20.62 j | 13.33 gef | 25.15 ef | 2.51 iklj | 117.56 gh | 39.62 fg | 1.14 g |
| 14 | 6.82 d | 105.16 j | 30.27 ef | 15.91 bc | 34.19 c | 3.30 e | 105.85 j | 55.56 ab | 1.52 d |
| 15 | 7.20 b | 100.16 ml | 36.29 a | 17.16 a | 40.68 b | 3.85 b | 99.55 m | 57.93 ab | 0.07 n |
| 16 | 7.32 a | 103.25 k | 38.16 a | 17.41 a | 43.84 a | 3.96 a | 96.33 o | 58.70 a | 0.07 n |
| 17 | 6.13 f | 122.50 b | 25.11 h | 14.33 de | 23.75 fg | 2.54 jki | 122.84 e | 50.07 c | 1.68 b |
| 18 | 5.76 ji | 132.16 a | 20.4 j | 13.08 gf | 24.47 efg | 2.63 h | 132.58 a | 45.88 ef | 0.80 i |
| 19 | 6.99 c | 101.83 kl | 33.07 bc | 16.66 ab | 40.85 b | 3.70 c | 98.33 n | 56.02 ab | 0.08 n |
| 20 | 6.19 f | 124.41 c | 22.14 i | 13.91 ef | 23.86 fg | 2.54 jki | 124.55 d | 43.06 ef | 0.50 k |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souri Laki, E.; Rabiei, B.; Jokarfard, V.; Shahbazi Miyangaskari, M.; Marashi, H.; Börner, A. Evaluation of Genotype × Environment Interactions in Quinoa Genotypes (Chenopodium quinoa Willd.). Agriculture 2025, 15, 515. https://doi.org/10.3390/agriculture15050515
Souri Laki E, Rabiei B, Jokarfard V, Shahbazi Miyangaskari M, Marashi H, Börner A. Evaluation of Genotype × Environment Interactions in Quinoa Genotypes (Chenopodium quinoa Willd.). Agriculture. 2025; 15(5):515. https://doi.org/10.3390/agriculture15050515
Chicago/Turabian StyleSouri Laki, Ebrahim, Babak Rabiei, Vahid Jokarfard, Mahboubeh Shahbazi Miyangaskari, Hassan Marashi, and Andreas Börner. 2025. "Evaluation of Genotype × Environment Interactions in Quinoa Genotypes (Chenopodium quinoa Willd.)" Agriculture 15, no. 5: 515. https://doi.org/10.3390/agriculture15050515
APA StyleSouri Laki, E., Rabiei, B., Jokarfard, V., Shahbazi Miyangaskari, M., Marashi, H., & Börner, A. (2025). Evaluation of Genotype × Environment Interactions in Quinoa Genotypes (Chenopodium quinoa Willd.). Agriculture, 15(5), 515. https://doi.org/10.3390/agriculture15050515