
Transcriptome Analysis of Resistant and Susceptible Sorghum Lines to the Sorghum Aphid (Melanaphis sacchari (Zehntner))

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plants and Aphid Cultivation
2.2. Assessment of Sorghum Resistance to Aphids
2.3. Management of Sorghum Inbred Lines with Varying Resistance Levels
2.4. RNA Extraction and Transcriptome Sequencing
2.5. Gene Expression Profiling
2.6. Quantitative qRT-PCR Expression Analysis
3. Results
3.1. Evaluation of the Sorghum Lines for Aphid Resistance
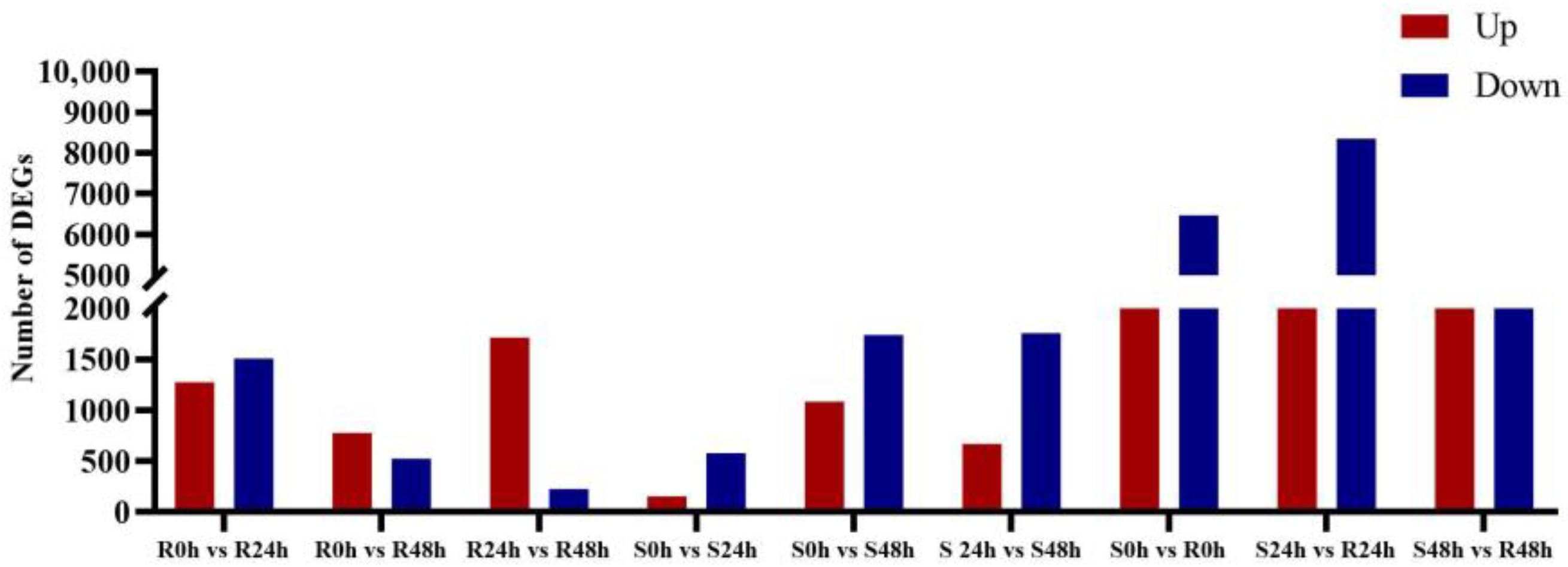
3.2. Differential Gene Expression in the Resistant and Susceptible Sorghum Lines
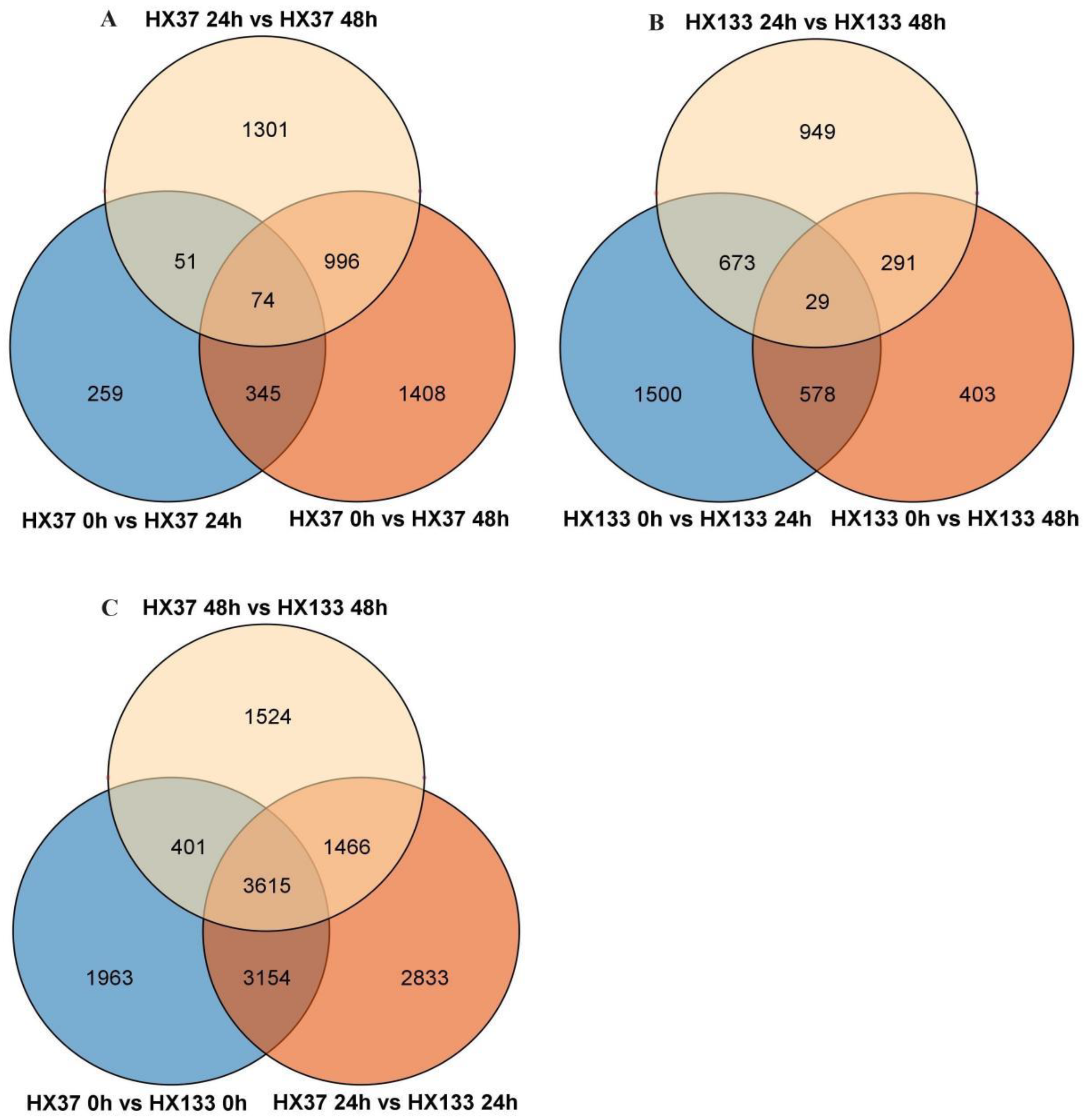
3.3. Venn Diagram Analysis
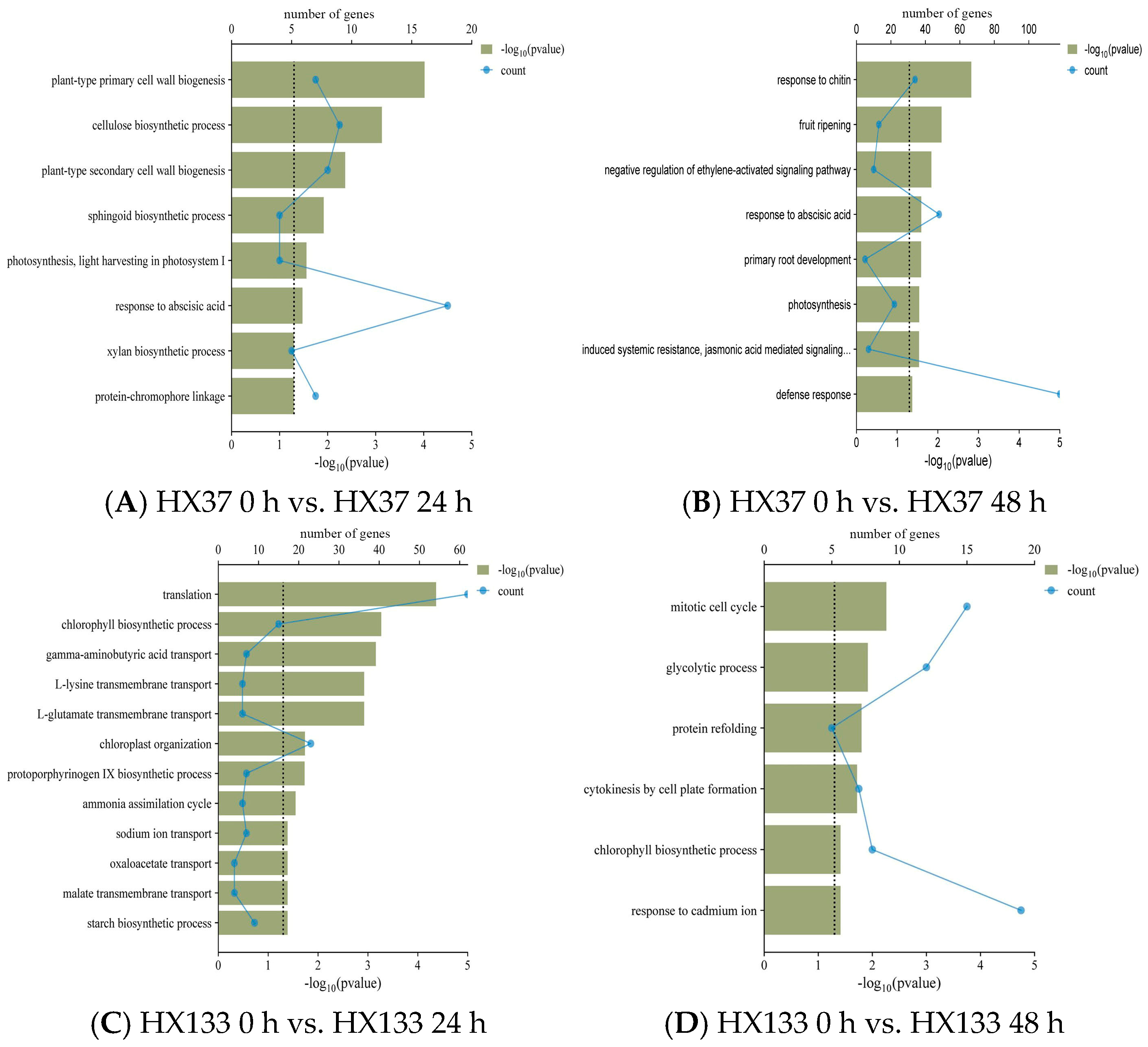
3.4. Gene Ontology Analysis
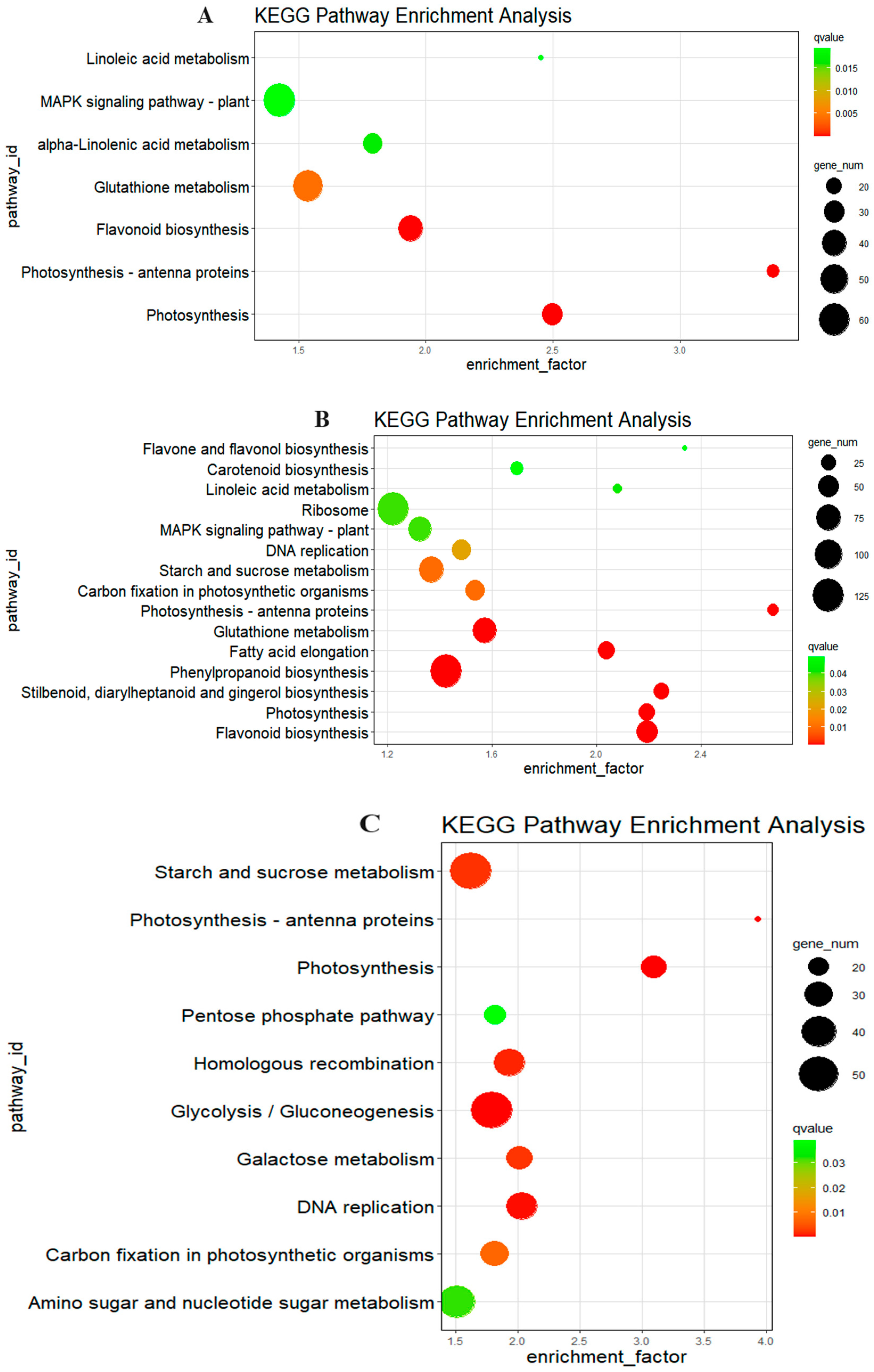
3.5. KEGG Pathway Analysis
3.6. Differentially Expressed Genes of Hormone Metabolism Post-Aphid Infestation
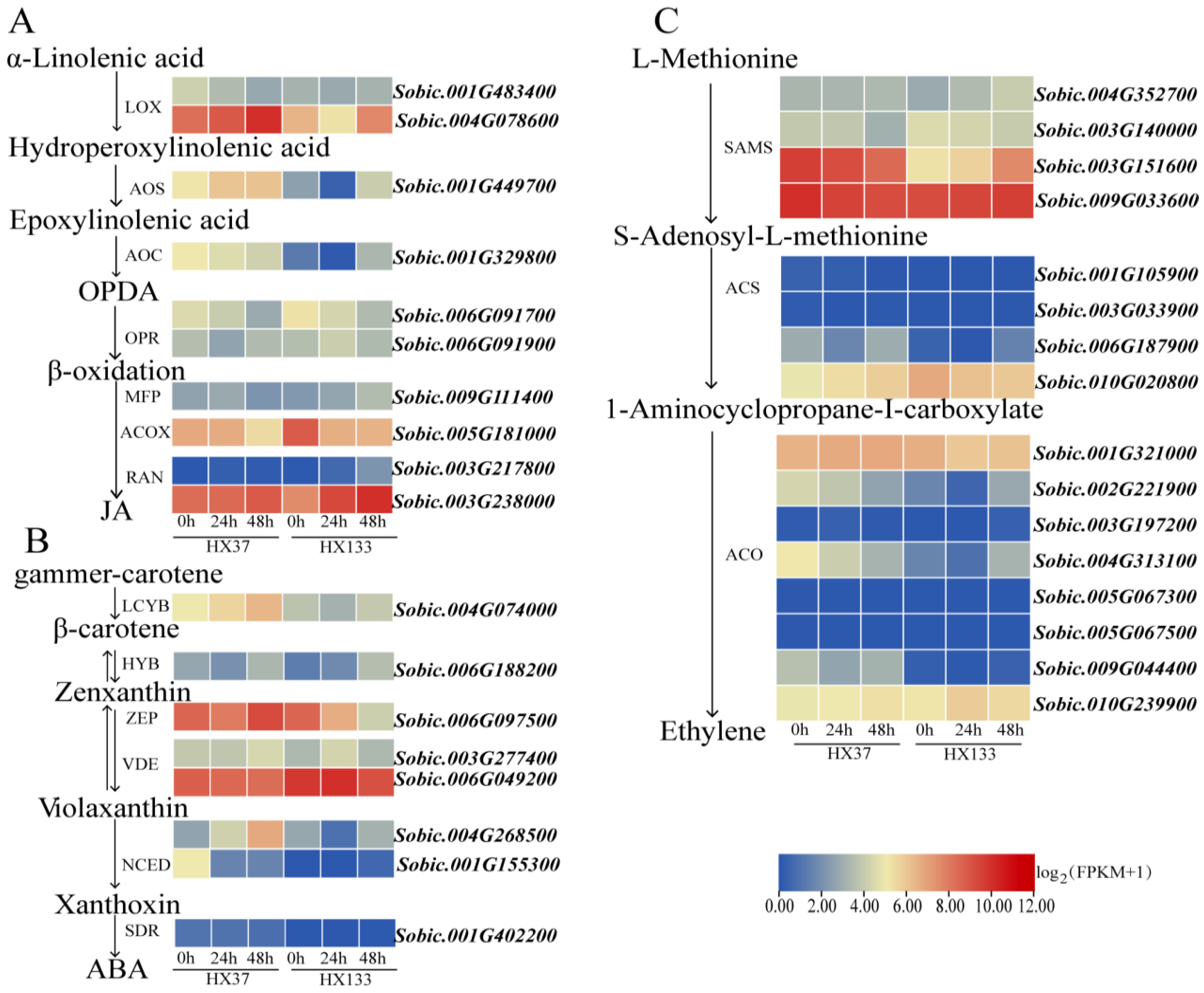
3.6.1. Jasmonic Acid Biosynthetic Pathway
3.6.2. Abscisic Acid Biosynthetic Pathway
3.6.3. Ethylene Biosynthetic Pathway
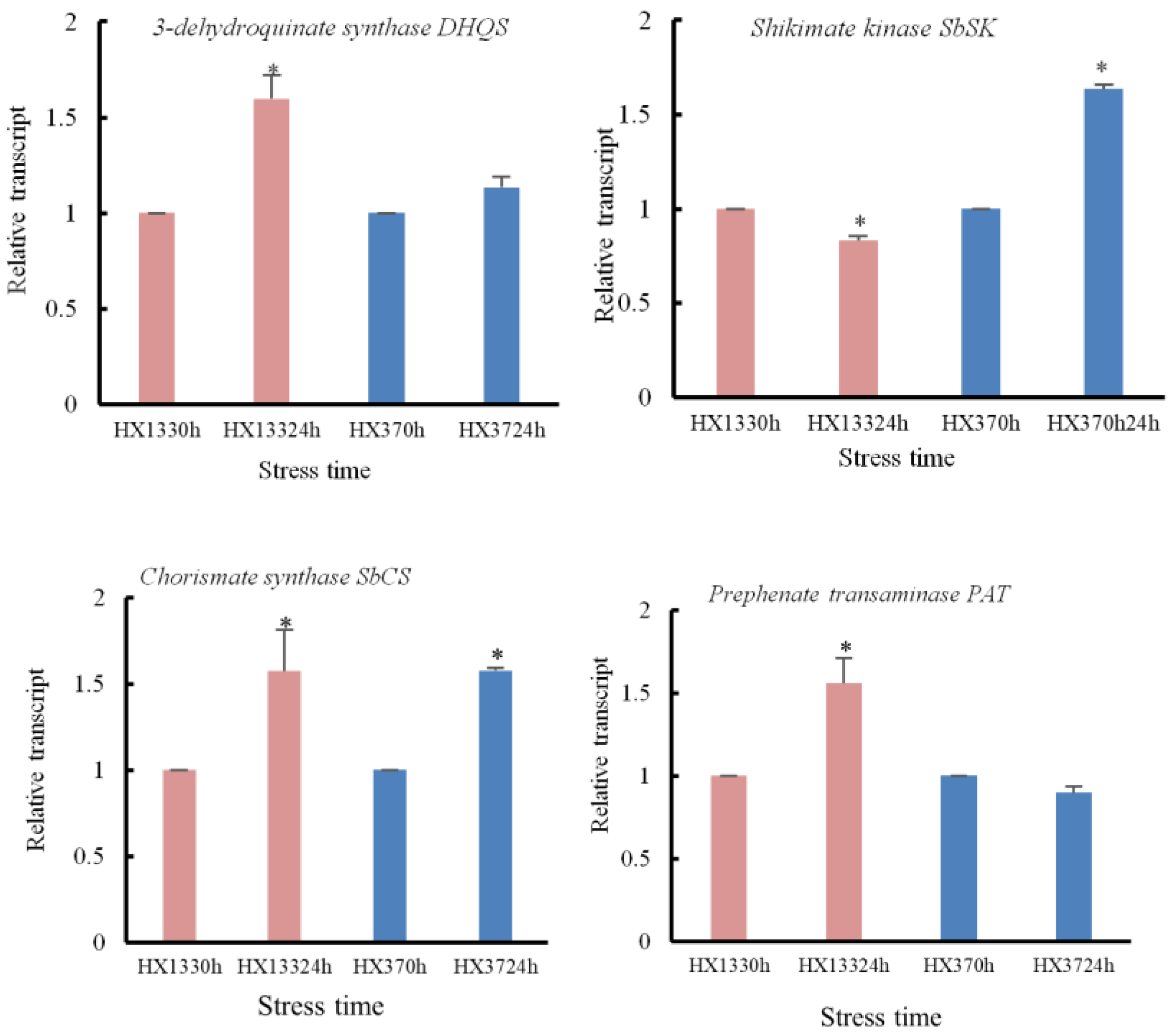
3.6.4. Shikimic Acid Pathway During Aphid Defense
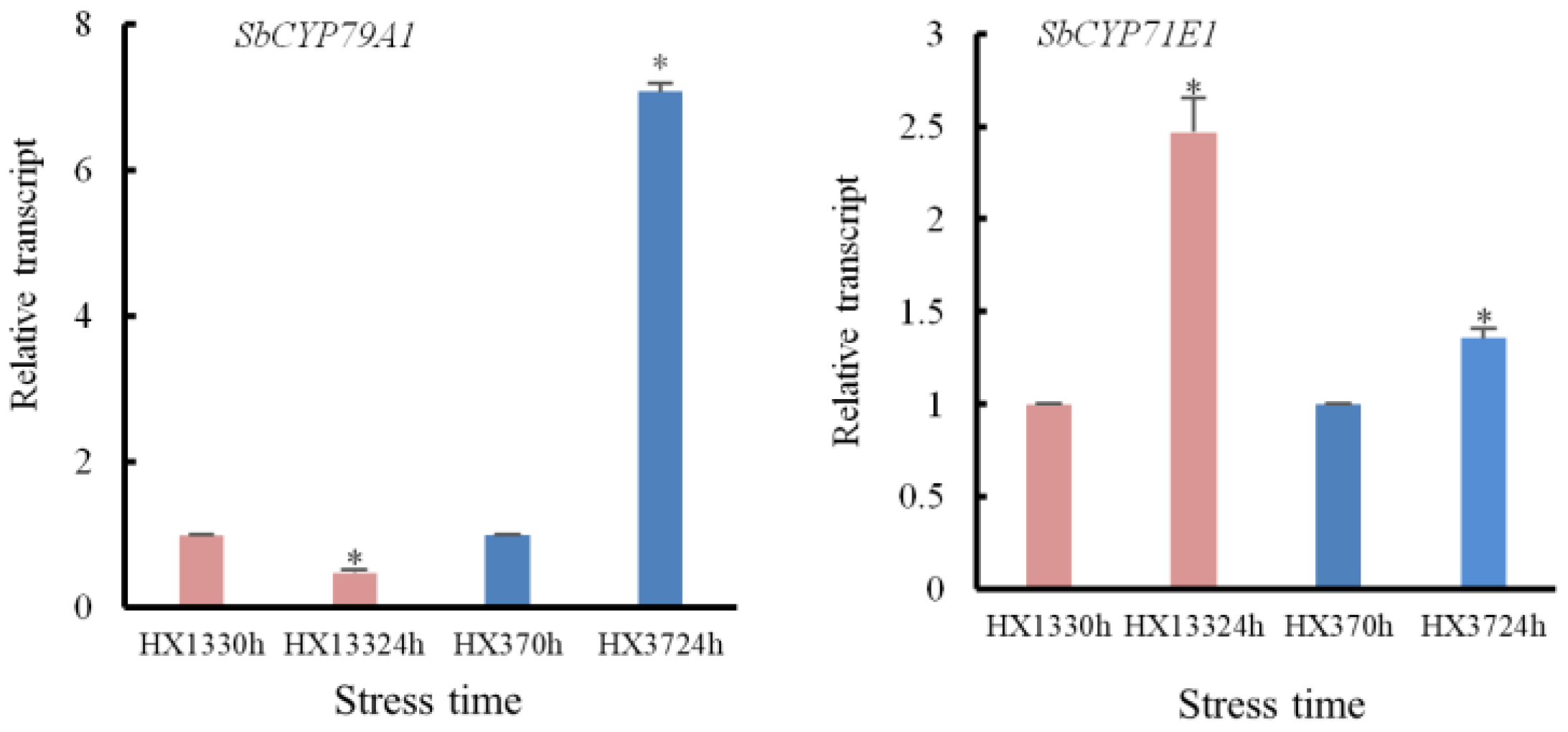
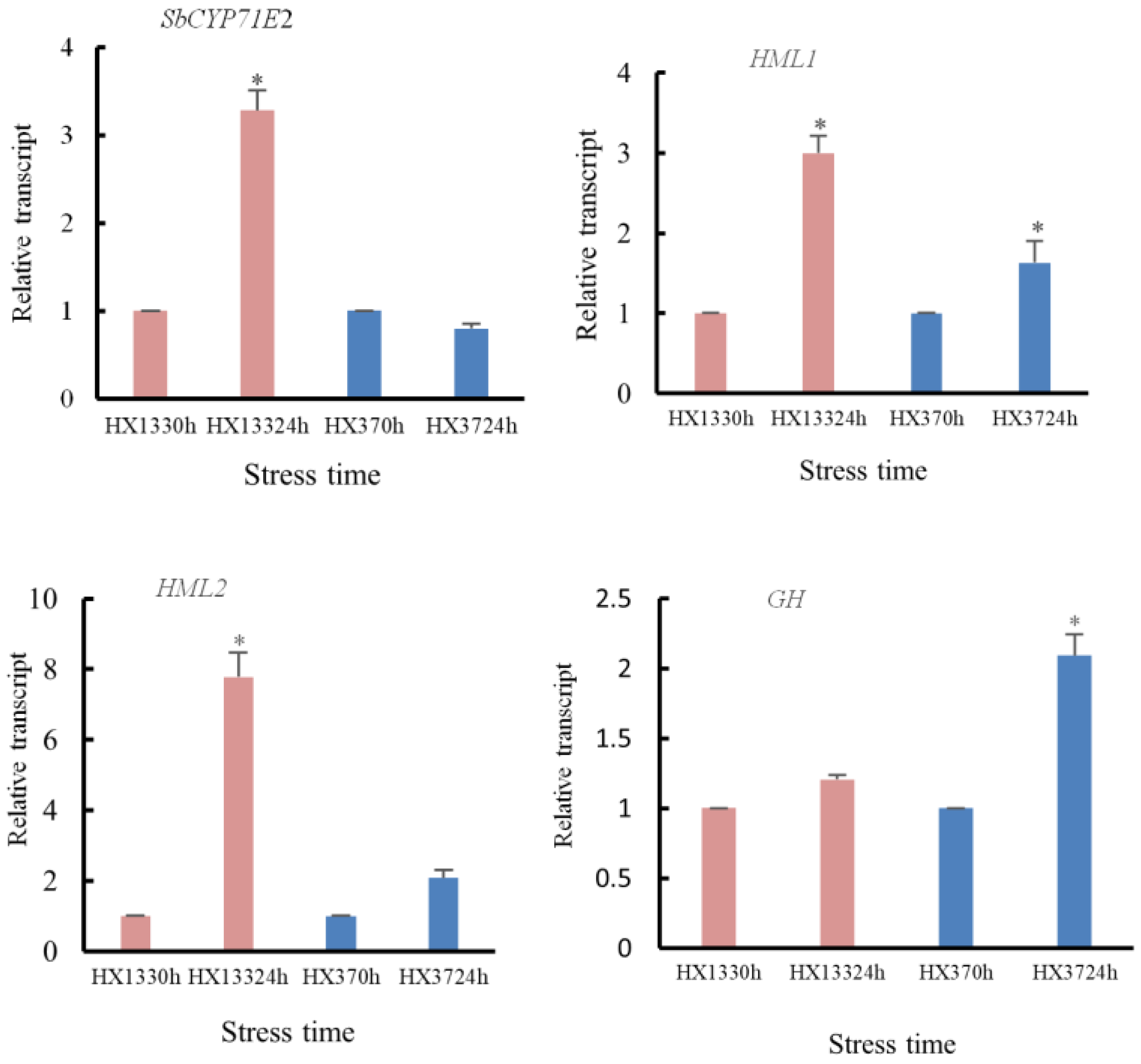
3.6.5. Dhurrin Biosynthesis and Metabolic Pathways
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| RNA-seqI | RNA Sequencing |
| qRT-PCR | Quantitative Reverse Transcription Polymerase Chain Reaction |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| DEGs | Differentially Expressed Genes |
| BP | Biological Processes |
| CC | Cellular Components |
| MF | Molecular Functions |
| hpi | Hours Post-Infestation |
| JA | Jasmonic Acid |
| ET | Ethylene |
| ABA | Abscisic Acid |
| LOX | Lipoxygenase |
| AOS | Allene Oxide Synthase |
| OPR | 12-Oxophytodienoate Reductase |
| LCYB | Lycopene Beta-Cyclase |
| ZEP | Zeaxanthin Epoxidase |
| NCED | 9-Cis-Epoxycarotenoid Dioxygenase |
| SAMS | S-Adenosyl-L-Methionine Synthase |
| ACS | ACC Synthase |
| ACO | ACC Oxidase |
| HML | Hydroxymandelonitrile Lyase |
| GH | Glycoside Hydrolase |
References
- Bowling, R.D.; Brewer, M.J.; Kerns, D.L.; Gordy, J.; Seiter, N.; Elliott, N.E.; Buntin, G.D.; Way, M.O.; Royer, T.A.; Biles, S.; et al. Sugarcane Aphid (Hemiptera: Aphididae): A New Pest on Sorghum in North America. J. Integr. Pest Manag. 2016, 7, 12. [Google Scholar] [CrossRef]
- Balbi, E.I.; Decker-Franco, C.; Szwarc, D.; Casuso, V.M.; Saluso, A.; Arneodo, J.D. Unveiling the occurrence of Melanaphis sorghi in Argentina following a major aphid outbreak. Phytoparasitica 2022, 51, 13–18. [Google Scholar] [CrossRef]
- Pekarcik, A.J.; Jacobson, A.L.; Giles, K. Evaluating Sugarcane Aphid, Melanaphis sacchari (Hemiptera: Aphididae), Population Dynamics, Feeding Injury, and Grain Yield Among Commercial Sorghum Varieties in Alabama. J. Econ. Entomol. 2021, 114, 757–768. [Google Scholar] [CrossRef]
- Peña-Martínez, R.; Lomeli-Flores, J.R.; Bujanos-Muñiz, R.; Salas-Monzón, R.; Hernández-Torres, O.E.; Marín-Jarillo, A.; Ibarra, J.E.; Vanegas-Rico, J.M.; Muñoz-Viveros, A.L. Comparative biology and life tables of sorghum aphid Melanaphis sorghi (Theobald) (Hemiptera: Aphididae) from Mexico, at different temperatures. Phytoparasitica 2024, 52, 33. [Google Scholar] [CrossRef]
- Viswanathan, R.; Nithya, K.; Vishnuvardhan, J.; Balasaravanan, S.; Kaverinathan, K.; Visalatchi, D. Sorghum (Sorghum bicolor) a new host to sugarcane yellow leaf and mosaic viruses in India. Indian Phytopathol. 2023, 76, 867–877. [Google Scholar] [CrossRef]
- Thompson, D.A.; Lehmler, H.-J.; Kolpin, D.W.; Hladik, M.L.; Vargo, J.D.; Schilling, K.E.; LeFevre, G.H.; Peeples, T.L.; Poch, M.C.; LaDuca, L.E. A critical review on the potential impacts of neonicotinoid insecticide use: Current knowledge of environmental fate, toxicity, and implications for human health. Environ. Sci. Process. Impacts 2020, 22, 1315–1346. [Google Scholar] [CrossRef] [PubMed]
- Guden, B.; Yol, E.; Ikten, C.; Erdurmus, C.; Uzun, B. Molecular and morphological evidence for resistance to sugarcane aphid (Melanaphis sacchari) in sweet sorghum [Sorghum bicolor (L.) Moench]. 3 Biotech 2019, 9, 245. [Google Scholar] [CrossRef]
- Haar, P.J.; Buntin, G.D.; Jacobson, A.; Pekarcik, A.; Way, M.O.; Zarrabi, A. Evaluation of Tactics for Management of Sugarcane Aphid (Hemiptera: Aphididae) in Grain Sorghum. J. Econ. Entomol. 2019, 112, 2719–2730. [Google Scholar] [CrossRef]
- Huang, Y. Improvement of crop protection against greenbug using the worldwide sorghum germplasm collection and genomics-based approaches. Plant Genet. Resour. 2011, 9, 317–320. [Google Scholar] [CrossRef]
- Paudyal, S.; Armstrong, J.S.; Giles, K.L.; Payton, M.E.; Opit, G.P.; Limaje, A. Categories of Resistance to Sugarcane Aphid (Hemiptera: Aphididae) Among Sorghum Genotypes. J. Econ. Entomol. 2019, 112, 1932–1940. [Google Scholar] [CrossRef]
- Pingault, L.; Palmer, N.A.; Koch, K.G.; Heng-Moss, T.; Bradshaw, J.D.; Seravalli, J.; Twigg, P.; Louis, J.; Sarath, G. Differential Defense Responses of Upland and Lowland Switchgrass Cultivars to a Cereal Aphid Pest. Int. J. Mol. Sci. 2020, 21, 7966. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Duan, X.; Cui, J.; Xue, W.; Zhang, Q. Differential molecular responses of aphid-sensitive and aphid-resistant sorghum lines to aphid infestation. Arthropod-Plant Interact. 2011, 6, 113–120. [Google Scholar] [CrossRef]
- Pingault, L.; Varsani, S.; Palmer, N.; Ray, S.; Williams, W.P.; Luthe, D.S.; Ali, J.G.; Sarath, G.; Louis, J. Transcriptomic and volatile signatures associated with maize defense against corn leaf aphid. BMC Plant Biol. 2021, 21, 138. [Google Scholar] [CrossRef]
- Shrestha, K.; Huang, Y. Genome-wide characterization of the sorghum JAZ gene family and their responses to phytohormone treatments and aphid infestation. Sci. Rep. 2022, 12, 3238. [Google Scholar] [CrossRef]
- Paudyal, S.; Armstrong, J.S.; Giles, K.L.; Hoback, W.; Aiken, R.; Payton, M.E. Differential responses of sorghum genotypes to sugarcane aphid feeding. Planta 2020, 252, 14. [Google Scholar] [CrossRef]
- Triplett, E.; Hayes, C.; Emendack, Y.; Longing, S.; Monclova, C.; Simpson, C.; Laza, H.E. Leaf structural traits mediating pre-existing physical innate resistance to sorghum aphid in sorghum under uninfested conditions. Planta 2023, 258, 46. [Google Scholar] [CrossRef]
- Palial, S.; Kumar, S.; Atri, C.; Sharma, S.; Banga, S.S. Antixenosis and antibiosis mechanisms of resistance to turnip aphid, Lipaphis erysimi (Kaltenbach) in Brassica juncea-fruticulosa introgression lines. J. Pest Sci. 2021, 95, 749–760. [Google Scholar] [CrossRef]
- Knoll, J.E.; Uchimiya, M.; Harris-Shultz, K. Juice chemical properties of 24 sorghum cultivars under varying levels of sugarcane aphid (Melanaphis sacchari) infestation. Arthropod-Plant Interact. 2021, 15, 707–719. [Google Scholar] [CrossRef]
- Balakrishnan, D.; Bateman, N.; Kariyat, R.R. Rice physical defenses and their role against insect herbivores. Planta 2024, 259, 110. [Google Scholar] [CrossRef]
- Cardona, J.B.; Grover, S.; Busta, L.; Sattler, S.E.; Louis, J. Sorghum cuticular waxes influence host plant selection by aphids. Planta 2022, 257, 22. [Google Scholar] [CrossRef]
- Hu, X.; Hereward, J.P.; Wang, D.; Yang, Q.; Wang, Y. A host-specialized aphid lineage helps another conspecific lineage utilize a new host by disrupting the plant defenses. J. Pest Sci. 2023, 97, 1525–1540. [Google Scholar] [CrossRef]
- Bezerra, R.H.S.; Sousa-Souto, L.; Santana, A.E.G.; Ambrogi, B.G. Indirect plant defenses: Volatile organic compounds and extrafloral nectar. Arthropod-Plant Interact. 2021, 15, 467–489. [Google Scholar] [CrossRef]
- Armstrong, J.S.; Rooney, W.L.; Peterson, G.C.; Villenueva, R.T.; Brewer, M.J.; Sekula-Ortiz, D. Sugarcane Aphid (Hemiptera: Aphididae): Host Range and Sorghum Resistance Including Cross-Resistance from Greenbug Sources. J. Econ. Entomol. 2015, 108, 576–582. [Google Scholar] [CrossRef]
- Poosapati, S.; Poretsky, E.; Dressano, K.; Ruiz, M.; Vazquez, A.; Sandoval, E.; Estrada-Cardenas, A.; Duggal, S.; Lim, J.-H.; Morris, G.; et al. A sorghum genome-wide association study (GWAS) identifies a WRKY transcription factor as a candidate gene underlying sugarcane aphid (Melanaphis sacchari) resistance. Planta 2022, 255, 37. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, G.; Wang, X.; Su, Y.; Diao, H.; Sun, D.; Shang, J.; Li, Y.; Guo, Y.; Qiu, L. Identification and characterization of resistance to soybean aphid in 22 highly resistant soybean accessions. Euphytica 2022, 218, 141. [Google Scholar] [CrossRef]
- Zhang, H.; Huang, J.; Huang, Y. Identification and characterization of plant resistance genes (R genes) in sorghum and their involvement in plant defense against aphids. Plant Growth Regul. 2022, 96, 443–461. [Google Scholar] [CrossRef]
- Tzin, V.; Fernandez-Pozo, N.; Richter, A.; Schmelz, E.A.; Schoettner, M.; Schafer, M.; Ahern, K.R.; Meihls, L.N.; Kaur, H.; Huffaker, A.; et al. Dynamic Maize Responses to Aphid Feeding Are Revealed by a Time Series of Transcriptomic and Metabolomic Assays. Plant Physiol. 2015, 169, 1727–1743. [Google Scholar] [CrossRef]
- Wang, T.; Wang, K.; Wang, C.; Zhao, Y.; Tao, Z.; Li, J.; Wang, L.; Shi, J.; Huang, S.; Xie, C.; et al. Combining quantitative trait locus mapping with multiomics profiling reveals genetic control of corn leaf aphid (Rhopalosiphum maidis) resistance in maize. J. Exp. Bot. 2023, 74, 3749–3764. [Google Scholar] [CrossRef]
- Batyrshina, Z.S.; Yaakov, B.; Shavit, R.; Singh, A.; Tzin, V. Comparative transcriptomic and metabolic analysis of wild and domesticated wheat genotypes reveals differences in chemical and physical defense responses against aphids. BMC Plant Biol. 2020, 20, 19. [Google Scholar] [CrossRef]
- Reddy, S.K.; Weng, Y.; Rudd, J.C.; Akhunova, A.; Liu, S. Transcriptomics of induced defense responses to greenbug aphid feeding in near isogenic wheat lines. Plant Sci. 2013, 212, 26–36. [Google Scholar] [CrossRef]
- Kiani, M.; Szczepaniec, A. Effects of sugarcane aphid herbivory on transcriptional responses of resistant and susceptible sorghum. BMC Genom. 2018, 19, 774. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Sun, L.; Jiao, B.; Zhao, P.; Ma, C.; Gao, J.; Zhou, S. Evaluation of aphid resistance on different rose cultivars and transcriptome analysis in response to aphid infestation. BMC Genom. 2024, 25, 232. [Google Scholar] [CrossRef]
- Lee, S.; Cassone, B.J.; Wijeratne, A.; Jun, T.-H.; Michel, A.P.; Mian, M.A.R. Transcriptomic dynamics in soybean near-isogenic lines differing in alleles for an aphid resistance gene, following infestation by soybean aphid biotype 2. BMC Genom. 2017, 18, 472. [Google Scholar] [CrossRef]
- MacWilliams, J.R.; Nabity, P.D.; Mauck, K.E.; Kaloshian, I. Transcriptome analysis of aphid-resistant and susceptible near isogenic lines reveals candidate resistance genes in cowpea (Vigna unguiculata). BMC Plant Biol. 2023, 23, 22. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Yang, Y.; Feng, P.; Ma, Q.; Su, Q.; Wang, X.; Zhang, J. Transcriptomic profiling of cotton leaves in response to cotton aphid damage. Acta Physiol. Plant. 2022, 44, 98. [Google Scholar] [CrossRef]
- Zhao, X.-R.; Zhao, D.-T.; Zhang, L.-Y.; Chang, J.-H.; Cui, J.-H. Combining transcriptome and metabolome analysis to understand the response of sorghum to Melanaphis sacchari. BMC Plant Biol. 2024, 24, 529. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhao, S.; Han, Y.; Shao, Y.; Dong, Z.; Gao, Y.; Zhang, K.; Liu, X.; Li, D.; Chang, J.; et al. Efficient and fine mapping of RMES1 conferring resistance to sorghum aphid Melanaphis sacchari. Mol. Breed. 2013, 31, 777–784. [Google Scholar] [CrossRef]
- Zhang, J.; Li, W.; Lv, P.; Yang, X.; Xu, W.; Ni, X.; Feng, H.; Zhao, G.; Pu, M.; Zhou, S.; et al. Whole-genome resequencing and transcriptome analysis provide insights on aphid-resistant quantitative trait loci/genes in Sorghum bicolor. Plant Breed. 2021, 140, 618–629. [Google Scholar] [CrossRef]
- Shrestha, K.; Huang, J.; Yan, L.; Doust, A.N.; Huang, Y. Integrated transcriptomic and pathway analyses of sorghum plants revealed the molecular mechanisms of host defense against aphids. Front. Plant Sci. 2024, 15, 1324085. [Google Scholar] [CrossRef]
- Tetreault, H.M.; Grover, S.; Scully, E.D.; Gries, T.; Palmer, N.A.; Sarath, G.; Louis, J.; Sattler, S.E. Global Responses of Resistant and Susceptible Sorghum (Sorghum bicolor) to Sugarcane Aphid (Melanaphis sacchari). Front. Plant Sci. 2019, 10, 145. [Google Scholar] [CrossRef]
- Du, J.-L.; Wu, D.-G.; Li, J.-Q.; Zhan, Q.-W.; Huang, S.-C.; Huang, B.-H.; Wang, X. Effects of aphid disoperation on photosynthetic performance and agronomic traits of different sorghum varieties. Pak. J. Bot. 2021, 53, 2275–2285. [Google Scholar] [CrossRef]
- Zou, K.; Liu, Y.; Wang, T.; Guan, M.; Li, X.; Li, J.; Yu, H.; Wu, D.; Du, J. Genomic Identification of Callose Synthase (CalS) Gene Family in Sorghum (Sorghum bicolor) and Comparative In Silico Expression Analysis under Aphid (Melanaphis sacchari) Infestation. Agronomy 2024, 14, 1393. [Google Scholar] [CrossRef]
- Serba, D.D.; Meng, X.; Schnable, J.; Bashir, E.; Michaud, J.P.; Vara Prasad, P.V.; Perumal, R. Comparative Transcriptome Analysis Reveals Genetic Mechanisms of Sugarcane Aphid Resistance in Grain Sorghum. Int. J. Mol. Sci. 2021, 22, 7129. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-Seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef]
- Mccormick, R.F.; Truong, S.K.; Sreedasyam, A.; Jenkins, J.; Shu, S.; Sims, D.; Kennedy, M.; Amirebrahimi, M.; Weers, B.D.; Mckinley, B. The Sorghum bicolor reference genome: Improved assembly, gene annotations, a transcriptome atlas, and signatures of genome organization. Plant J. 2018, 93, 338–354. [Google Scholar] [CrossRef]
- Zou, K.; Wang, T.; Guan, M.; Liu, Y.; Li, J.; Liu, Y.; Du, J.; Wu, D. Identification and Evaluation of qRT-PCR Reference Genes in Melanaphis sacchari. Insects 2024, 15, 522. [Google Scholar] [CrossRef]
- Maeda, H.; Dudareva, N. The Shikimate Pathway and Aromatic Amino Acid Biosynthesis in Plants. Annu. Rev. Plant Biol. 2012, 63, 73. [Google Scholar] [CrossRef]
- Fakhrah, S.; Bano, N.; Sarvendra, K.; Lone, R.A.; Nayak, S.P.; Kumari, A.; Rout, P.K.; Mohanty, C.S. Elucidating the Secondary Metabolite Biosynthesis Networks in Underutilized Tree Bean (Parkia timoriana) Through Integrated Metabolomic and Transcriptomic Approaches. Appl. Biochem. 2025, 197, 3521–3547. [Google Scholar] [CrossRef]
- Myrans, H.; Vandegeer, R.K.; Henry, R.J.; Gleadow, R.M. Nitrogen availability and allocation in sorghum and its wild relatives: Divergent roles for cyanogenic glucosides. J. Plant Physiol. 2021, 258–259, 153393. [Google Scholar] [CrossRef]
- English, B.; Quinn, A.A.; Warren, C.R.; Gleadow, R.M.; Myrans, H. Do cyanogenic glucosides help sorghum manage a fluctuating nitrogen supply? Funct. Plant Biol. 2025, 52, FP24343. [Google Scholar] [CrossRef] [PubMed]
- Darbani, B.; Motawia, M.S.; Olsen, C.E.; Nour-Eldin, H.H.; Møller, B.L.; Rook, F. The biosynthetic gene cluster for the cyanogenic glucoside dhurrin in Sorghum bicolor contains its co-expressed vacuolar MATE transporter. Sci. Rep. 2016, 6, 37079. [Google Scholar] [CrossRef] [PubMed]
- Gleadow, R.M.; Mckinley, B.A.; Blomstedt, C.K.; Lamb, A.C.; Mller, B.L.; Mullet, J.E. Regulation of dhurrin pathway gene expression during Sorghumbicolor development. Planta 2021, 254, 119. [Google Scholar] [CrossRef]
- Ku, Y.-S.; Sintaha, M.; Cheung, M.-Y.; Lam, H.-M. Plant Hormone Signaling Crosstalks between Biotic and Abiotic Stress Responses. Int. J. Mol. Sci. 2018, 19, 3206. [Google Scholar] [CrossRef] [PubMed]
- Onkokesung, N.; Baldwin, I.T.; Gális, I. The role of jasmonic acid and ethylene crosstalk in direct defense ofNicotiana attenuataplants against chewing herbivores. Plant Signal. Behav. 2014, 5, 1305–1307. [Google Scholar] [CrossRef]
- Shiu, S.-H.; Sewelam, N.; Kazan, K.; Thomas-Hall, S.R.; Kidd, B.N.; Manners, J.M.; Schenk, P.M. Ethylene Response Factor 6 Is a Regulator of Reactive Oxygen Species Signaling in Arabidopsis. PLoS ONE 2013, 8, e70289. [Google Scholar]
- Guo, H.; Sun, Y.; Peng, X.; Wang, Q.; Harris, M.; Ge, F. Up-regulation of abscisic acid signaling pathway facilitates aphid xylem absorption and osmoregulation under drought stress. J. Exp. Bot. 2016, 67, 681–693. [Google Scholar] [CrossRef]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2008, 69, 473–488. [Google Scholar] [CrossRef]
- Wang, Y.; Mostafa, S.; Zeng, W.; Jin, B. Function and Mechanism of Jasmonic Acid in Plant Responses to Abiotic and Biotic Stresses. Int. J. Mol. Sci. 2021, 22, 8568. [Google Scholar] [CrossRef]
- Li, A.; Zhou, M.; Wei, D.; Chen, H.; You, C.; Lin, J. Transcriptome Profiling Reveals the Negative Regulation of Multiple Plant Hormone Signaling Pathways Elicited by Overexpression of C-Repeat Binding Factors. Front. Plant Sci. 2017, 8, 1647. [Google Scholar] [CrossRef]
- Yang, J.; Duan, G.; Li, C.; Liu, L.; Han, G.; Zhang, Y.; Wang, C. The crosstalks between jasmonic acid and other plant hormone signaling highlight the involvement of jasmonic acid as a core component in plant response to biotic and abiotic stresses. Front. Plant Sci. 2019, 10, 1349. [Google Scholar] [CrossRef]
- Mao, Y.-B.; Liu, Y.-Q.; Chen, D.-Y.; Chen, F.-Y.; Fang, X.; Hong, G.-J.; Wang, L.-J.; Wang, J.-W.; Chen, X.-Y. Jasmonate response decay and defense metabolite accumulation contributes to age-regulated dynamics of plant insect resistance. Nat. Commun. 2017, 8, 13925. [Google Scholar] [CrossRef]
- Louis, J.; Basu, S.; Varsani, S.; Castano-Duque, L.; Jiang, V.; Williams, W.P.; Felton, G.W.; Luthe, D.S. Ethylene Contributes to maize insect resistance1-Mediated Maize Defense against the Phloem Sap-Sucking Corn Leaf Aphid. Plant Physiol. 2015, 169, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Ton, J.; Flors, V.; Mauch-Mani, B. The multifaceted role of ABA in disease resistance. Trends Plant Sci. 2009, 14, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Ullah, C.; Schmidt, A.; Reichelt, M.; Tsai, C.J.; Gershenzon, J. Lack of antagonism between salicylic acid and jasmonate signalling pathways in poplar. New Phytol. 2022, 235, 701–717. [Google Scholar] [CrossRef] [PubMed]
- Gleadow, R.M.; Møller, B.L. Cyanogenic Glycosides: Synthesis, Physiology, and Phenotypic Plasticity. Annu. Rev. Plant Biol. 2014, 65, 155–185. [Google Scholar] [CrossRef]
- Gamir, J.; Sánchez-Bel, P.; Flors, V. Molecular and physiological stages of priming: How plants prepare for environmental challenges. Plant Cell Rep. 2014, 33, 1935–1949. [Google Scholar] [CrossRef]
- Dangol, A.; Shavit, R.; Yaakov, B.; Strickler, S.R.; Jander, G.; Tzin, V. Characterizing serotonin biosynthesis in Setaria viridis leaves and its effect on aphids. Plant Mol. Biol. 2022, 109, 533–549. [Google Scholar] [CrossRef]
- Gruss, S.M.; Ghaste, M.; Widhalm, J.R.; Tuinstra, M.R. Seedling growth and fall armyworm feeding preference influenced by dhurrin production in sorghum. Theor. Appl. Genet. 2022, 135, 1037–1047. [Google Scholar] [CrossRef]
- Laursen, T.; Møller, B.L.; Bassard, J.-E. Plasticity of specialized metabolism as mediated by dynamic metabolons. Trends Plant Sci. 2015, 20, 20–32. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sorghum Line | Pathway | Upregulate or Downregulated | Enrichment Factor | Qvalue | Gene Number |
|---|---|---|---|---|---|
| HX370h vs. HX3724h | Photosynthesis-antenna proteins | Down | 16.2644 | 0.00006 | 6 |
| Sphingolipid metabolism | Down | 7.9431 | 0.0006 | 7 | |
| Carotenoid biosynthesis | Down | 6.7768 | 0.0158 | 5 | |
| Phenylpropanoid biosynthesis | Down | 2.4224 | 0.0272 | 14 | |
| MAPK signaling pathway in plants | Down | 2.9572 | 0.0272 | 10 | |
| Glutathione metabolism | Up | 7.6607 | 0.0028 | 6 | |
| HX370h vs. HX3748h | Starch and sucrose metabolism | Down | 2.9929 | 0.000006 | 29 |
| Galactose metabolism | Down | 3.0706 | 0.0210 | 12 | |
| MAPK signaling pathway in plants | Down | 2.1510 | 0.0390 | 19 | |
| Plant–pathogen interaction | Down | 2.0220 | 0.0390 | 21 | |
| Sphingolipid metabolism | Down | 3.4753 | 0.0390 | 8 | |
| Amino sugar and nucleotide sugar metabolism | Down | 2.1147 | 0.0390 | 18 | |
| Alanine, aspartate, and glutamate metabolism | Down | 2.8986 | 0.0469 | 9 | |
| Photosynthesis | Up | 8.1321 | 0.000004 | 11 | |
| Photosynthesis-antenna proteins | Up | 10.8429 | 0.0004 | 6 | |
| Monoterpenoid biosynthesis | Up | 7.3928 | 0.0117 | 5 | |
| Benzoxazinoid biosynthesis | Up | 21.6858 | 0.0545 | 2 | |
| HX1330h vs. HX13324h | Galactose metabolism | Down | 4.0352 | 0.0006 | 14 |
| Plant hormone signal transduction | Down | 2.0509 | 0.0125 | 27 | |
| Glutathione metabolism | Down | 2.5014 | 0.0128 | 17 | |
| Phenylalanine, tyrosine, and tryptophan biosynthesis | Up | 3.9260 | 0.0036 | 12 | |
| Porphyrin and chlorophyll metabolism | Up | 3.3322 | 0.0188 | 11 | |
| Pentose phosphate pathway | Up | 3.1158 | 0.0188 | 12 | |
| Starch and sucrose metabolism | Up | 2.0786 | 0.0188 | 23 | |
| Carbon fixation in photosynthetic organisms | Up | 2.5446 | 0.0225 | 14 | |
| Aminoacyl-tRNA biosynthesis | Up | 2.7263 | 0.0230 | 12 | |
| Ribosome | Up | 1.6905 | 0.0247 | 34 | |
| Ubiquinone and other terpenoid-quinone biosynthesis | Up | 2.7263 | 0.0433 | 10 | |
| Steroid biosynthesis | Up | 2.5706 | 0.0433 | 11 | |
| Monobactam biosynthesis | Up | 5.9485 | 0.0433 | 4 | |
| Fatty acid biosynthesis | Up | 2.6817 | 0.0433 | 10 | |
| Carotenoid biosynthesis | Up | 3.1807 | 0.0500 | 7 | |
| Cutin, suberine, and wax biosynthesis | Up | 2.6768 | 0.0500 | 9 | |
| HX1330h vs. HX13348h | Valine, leucine, and isoleucine degradation | Down | 7.0750 | 0.0121 | 6 |
| Other glycan degradation | Down | 10.7809 | 0.0150 | 4 | |
| Circadian rhythmin plants | Down | 5.9578 | 0.0150 | 6 | |
| alpha-Linolenic acid metabolism | Down | 5.1454 | 0.0464 | 5 | |
| Propanoate metabolism | Down | 5.9578 | 0.0589 | 4 | |
| Base excision repair | Up | 4.5999 | 0.0100 | 9 | |
| Pentose phosphate pathway | Up | 3.5777 | 0.0356 | 9 | |
| Carbon fixation in photosynthetic organisms | Up | 3.0609 | 0.0356 | 11 | |
| DNA replication | Up | 3.0273 | 0.0356 | 11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guan, M.; Du, J.; Li, J.; Wang, T.; Sun, L.; Wang, Y.; Wu, D. Transcriptome Analysis of Resistant and Susceptible Sorghum Lines to the Sorghum Aphid (Melanaphis sacchari (Zehntner)). Agriculture 2025, 15, 1502. https://doi.org/10.3390/agriculture15141502
Guan M, Du J, Li J, Wang T, Sun L, Wang Y, Wu D. Transcriptome Analysis of Resistant and Susceptible Sorghum Lines to the Sorghum Aphid (Melanaphis sacchari (Zehntner)). Agriculture. 2025; 15(14):1502. https://doi.org/10.3390/agriculture15141502
Chicago/Turabian StyleGuan, Minghui, Junli Du, Jieqin Li, Tonghan Wang, Lu Sun, Yongfei Wang, and Degong Wu. 2025. "Transcriptome Analysis of Resistant and Susceptible Sorghum Lines to the Sorghum Aphid (Melanaphis sacchari (Zehntner))" Agriculture 15, no. 14: 1502. https://doi.org/10.3390/agriculture15141502
APA StyleGuan, M., Du, J., Li, J., Wang, T., Sun, L., Wang, Y., & Wu, D. (2025). Transcriptome Analysis of Resistant and Susceptible Sorghum Lines to the Sorghum Aphid (Melanaphis sacchari (Zehntner)). Agriculture, 15(14), 1502. https://doi.org/10.3390/agriculture15141502