
Impact of Drought, Heat, Excess Light, and Salinity on Coffee Production: Strategies for Mitigating Stress Through Plant Breeding and Nutrition

 , , ,
, , ,  ,
,  and
and
Abstract
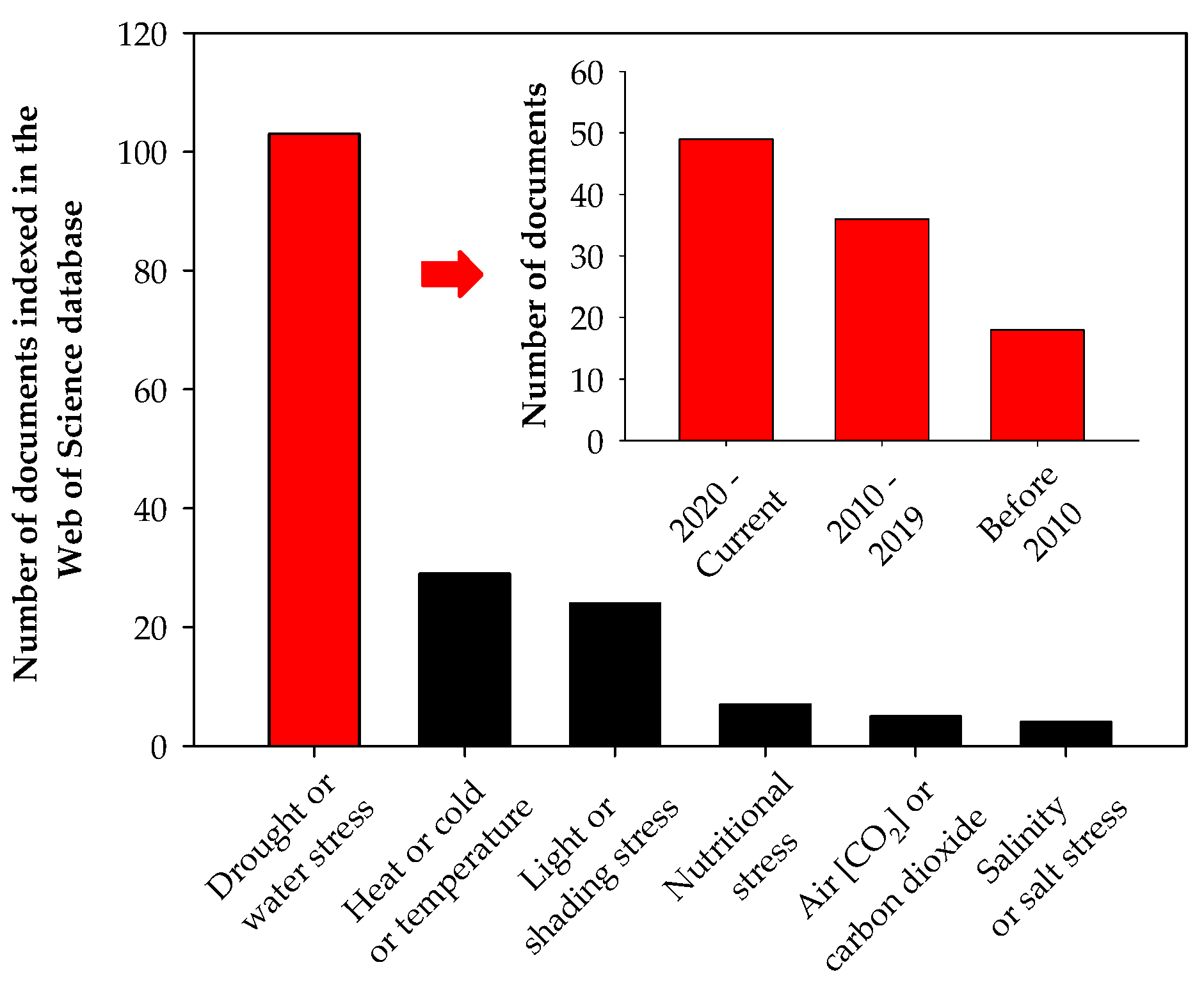
1. Introduction
2. Abiotic Stress Factors and Their Relationship with Coffee Plant Growth
2.1. Drought Stress in Coffee
2.2. Heat and Excess Light Stress in Coffee
2.3. Salinity Stress in Coffee
3. Strategies to Improve Coffee Plant Tolerance to Abiotic Stress
3.1. Plant Breeding Towards Abiotic Stress Tolerance
3.2. Plant Nutrition as a Tool to Improve Coffee Tolerance to Abiotic Stress
4. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- USDA—United States Department of Agriculture. Brazil: Coffee Annual. 2023. Available online: https://fas.usda.gov/data/brazil-coffee-annual-8 (accessed on 16 July 2024).
- WMO—World Meteorological Organization. State of the Global Climate 2023. WMO-No. 1347. 2024. Available online: https://library.wmo.int/idurl/4/68835 (accessed on 21 October 2024).
- Koutouleas, A.; Sarzynski, T.; Bordeaux, M.; Bosselmann, A.S.; Campa, C.; Etienne, H.; Turreira-García, N.; Rigal, C.; Vaast, P.; Ramalho, J.C.; et al. Shaded-Coffee: A Nature-Based Strategy for Coffee Production under Climate Change? A Review. Front. Sustain. Food Syst. 2022, 6, 877476. [Google Scholar] [CrossRef]
- DaMatta, F.M.; Avila, R.T.; Cardoso, A.A.; Martins, S.C.V.; Ramalho, J.C. Physiological and agronomic performance of the coffee crop in the context of climate change and global warming: A Review. J. Agric. Food Chem. 2018, 66, 5264–5274. [Google Scholar] [CrossRef] [PubMed]
- Magrach, A.; Ghazoul, J. Climate and pest-driven geographic shifts in global coffee production: Implications for forest cover, biodiversity and carbon storage. PLoS ONE 2015, 10, e0133071. [Google Scholar] [CrossRef] [PubMed]
- Rigal, C.; Xu, J.; Vaast, P. Young shade trees improve soil quality in intensively managed coffee systems recently converted to agroforestry in Yunnan Province, China. Plant Soil 2020, 453, 119–137. [Google Scholar] [CrossRef]
- Braga, G.B.; Imbuzeiro, H.M.A.; Pires, G.F.; Oliveira, L.R.D.; Barbosa, R.A.; Vilela, K.D.F. Frost risk and rural insurance in Brazil. Rev. Bras. Meteorol. 2021, 36, 703–711. [Google Scholar] [CrossRef]
- Jayakumar, M.; Rajavel, M.; Surendran, U.; Gopinath, G.; Ramamoorthy, K. Impact of climate variability on coffee yield in India—With a micro-level case study using long-term coffee yield data of humid tropical Kerala. Clim. Change 2017, 145, 335–349. [Google Scholar] [CrossRef]
- Lima, R.B.; dos Santos, T.B.; Vieira, L.G.E.; Ferrarese, M.L.L.; Ferrarese-Filho, O.; Donatti, L.; Boeger, M.R.T.; Petkowicz, C.L.O. Heat stress causes alterations in the cell-wall polymers and anatomy of coffee leaves (Coffea arabica L.). Carbohydr. Polym. 2013, 93, 135–143. [Google Scholar] [CrossRef]
- Avila, R.T.; Almeida, W.L.; Costa, L.C.; Machado, K.L.G.; Barbosa, M.L.; Souza, R.P.B.; Martino, P.B.; Juárez, M.A.T.; Marçal, D.M.S.; Martins, S.C.V.; et al. Elevated air [CO2] improves photosynthetic performance and alters biomass accumulation and partitioning in drought-stressed coffee plants. Environ. Exp. Bot. 2020, 177, 104137. [Google Scholar] [CrossRef]
- dos Santos, T.B.; da Silva Ferreira, M.F.; Marques, I.; Oliveira, S.C.; Zaidan, I.R.; Oliveira, M.G.; Rodrigues, W.P.; Ribas, A.F.; Guyot, R.; Ramalho, J.C.; et al. Current challenges and genomic advances towards the development resilient coffee genotypes to abiotic stresses. In Genomic Designing for Abiotic Stress Resistant Technical Crops; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2022; pp. 41–69. [Google Scholar]
- Kumari, V.V.; Banerjee, P.; Verma, V.C.; Sukumaran, S.; Chandran, M.A.S.; Gopinath, K.A.; Venkatesh, G.; Yadav, S.K.; Singh, V.K.; Awasthi, N.K. Plant nutrition: An Effective Way to alleviate abiotic stress in agricultural crops. Int. J. Mol. Sci. 2022, 23, 8519. [Google Scholar] [CrossRef]
- Sharma, M.; Kumar, P.; Verma, V.; Sharma, R.; Bhargava, B.; Irfan, M. Understanding plant stress memory response for abiotic stress resilience: Molecular insights and prospects. Plant Physiol. Biochem. 2022, 179, 10–24. [Google Scholar] [CrossRef]
- Menezes-Silva, P.E.; Sanglard, L.M.; Ávila, R.T.; Morais, L.E.; Martins, S.C.; Nobres, P.; Patreze, C.M.; Ferreira, M.A.; Araújo, W.L.; Fernie, A.R. Photosynthetic and metabolic acclimation to repeated drought events play key roles in drought tolerance in coffee. J. Exp. Bot. 2017, 68, 4309–4322. [Google Scholar] [CrossRef] [PubMed]
- Kabbadj, A.; Makoudi, B.; Mouradi, M.; Pauly, N.; Frendo, P.; Ghoulam, C. Physiological and biochemical responses involved in water deficit tolerance of nitrogen-fixing Vicia faba. PLoS ONE 2017, 12, e0190284. [Google Scholar] [CrossRef] [PubMed]
- Salehi-Lisar, S.Y.; Bakhshayeshan-Agdam, H. Drought Stress in Plants: Causes, Consequences, and Tolerance; Hossain, M., Wani, S., Bhattacharjee, S., Burritt, D., Tran, L.S., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; Volume 1, pp. 1–16. [Google Scholar]
- Gimenez, C.; Gallardo, M.; Thompson, R.B. Plant water relations. In Encyclopedia of Soils in the Environment; Hillel, D., Ed.; Elsevier: Oxford, UK, 2005; pp. 231–238. [Google Scholar]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Somasundaram, R.; Panneerselvam, R. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Bhargava, S.; Sawant, K. Drought stress adaptation: Metabolic adjustment and regulation of gene expression. Plant Breed. 2013, 132, 21–32. [Google Scholar] [CrossRef]
- DaMatta, F.M.; Maestri, M.; Barros, R.S.; Regazzi, A.J. Water relations of coffee leaves (Coffea arabica and C. canephora) in response to drought. J. Hortic. Sci.-India 1993, 68, 741–746. [Google Scholar] [CrossRef]
- Lima, A.A.; Santos, I.S.; Torres, M.E.L.; Cardon, C.H.; Caldeira, C.F.; Lima, R.R.; Davies, W.J.; Dodd, I.C.; Chalfun-Junior, A. Drought and re-watering modify ethylene production and sensitivity, and are associated with coffee anthesis. Environ. Exp. Bot. 2021, 181, 104289. [Google Scholar] [CrossRef]
- Cavatte, P.C.; Oliveira, A.A.G.; Morais, L.E.; Martins, S.C.V.; Sanglard, L.M.V.P.; DaMatta, F.M. Could shading reduce the negative impacts of drought on coffee? A morphophysiological analysis. Physiol. Plant. 2012, 144, 111–122. [Google Scholar] [CrossRef]
- DaMatta, F.M.; Maestri, M.; Barros, R.S. Phothosynthetic performance of two coffee species under drought. Photosynthetica 1997, 34, 257–264. [Google Scholar] [CrossRef]
- de Andrade, L.I.F.; Linhares, P.C.A.; da Fonseca, T.M.; Silva, A.A.; Santos, J.P.; Pereira, M.P.; Silva, V.A.; Marchiori, P.E.R. Photosynthetic efficiency and root plasticity promote drought tolerance in coffee genotypes. Acta Physiol. Plant. 2022, 44, 109. [Google Scholar] [CrossRef]
- Chekol, H.; Warkineh, B.; Shimber, T.; Mierek-Adamska, A.; Dąbrowska, G.B.; Degu, A. Drought Stress Responses in Arabica Coffee Genotypes: Physiological and Metabolic Insights. Plants 2024, 13, 828. [Google Scholar] [CrossRef] [PubMed]
- Chekol, H.; Bezuayehu, Y.; Warkineh, B.; Shimber, T.; Mierek-Adamska, A.; Dąbrowska, G.B.; Degu, A. Unraveling Drought Tolerance and Sensitivity in Coffee Genotypes: Insights fromSeed Traits, Germination, and Growth-Physiological Responses. Agriculture 2023, 13, 1754. [Google Scholar] [CrossRef]
- Aman, M.; Worku, M.; Shimbir, T.; Astatkie, T. Root traits and biomass production of drought-resistant and drought-sensitive arabica coffee varieties growing under contrasting watering regimes. Agrosyst. Geosci. Environ. 2024, 7, e20488. [Google Scholar] [CrossRef]
- Praxedes, S.C.; DaMatta, F.M.; Loureiro, M.E.; Ferrão, M.A.G.; Cordeiro, A.T. Effects of long-term soil drought on photosynthesis and carbohydrate metabolism in mature robusta coffee (Coffea canephora Pierre var. kouillou) leaves. Environ. Exp. Bot. 2006, 56, 263–273. [Google Scholar] [CrossRef]
- Hassan, M.U.; Chattha, M.U.; Khan, I.; Chattha, M.B.; Barbanti, L.; Aamer, M.; Iqbal, M.M.; Nawaz, M.; Mahmood, A.; Ali, A.; et al. Heat stress in cultivated plants: Nature, impact, mechanisms, and mitigation strategies—A review. Plant Biosyst. 2021, 155, 211–234. [Google Scholar] [CrossRef]
- Carvalho, M.H.C. Drought stress and reactive oxygen species. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef]
- de Guedes, F.A.F.; Nobres, P.; Ferreira, D.C.R.; Menezes-Silva, P.E.; Ribeiro-Alves, M.; Correa, R.L.; DaMatta, F.M.; Alves-Ferreira, M. Transcriptional memory contributes to drought tolerance in coffee (Coffea canephora) plants. Environ. Exp. Bot. 2018, 147, 220–233. [Google Scholar] [CrossRef]
- Woodward, F.I. Stomatal numbers are sensitive to increases in CO2 from preindustrial levels. Nature 1987, 327, 617–618. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions: Photosynthesis and stomatal conductance responses to rising [CO2]. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef]
- Rakocevic, M.; Ribeiro, R.V.; Marchiori, P.E.R.; Filizola, H.F.; Batista, E.R. Structural and functional changes in coffee trees after 4 years under free air CO2 enrichment. Ann. Bot. 2018, 121, 1065–1078. [Google Scholar] [CrossRef]
- Leakey, A.D.B.; Ainsworth, E.A.; Bernacchi, C.J.; Rogers, A.; Long, S.P.; Ort, D.R. Elevated CO2 effects on plant carbon, nitrogen, and water relations: Six important lessons from FACE. J. Exp. Bot. 2009, 60, 2859–2876. [Google Scholar] [CrossRef] [PubMed]
- Sicher, R.C.; Timlin, D.; Bailey, B. Responses of growth and primary metabolism of water-stressed barley roots to rehydration. J. Plant Physiol. 2012, 169, 686–695. [Google Scholar] [CrossRef]
- Li, N.; Euring, D.; Cha, J.Y.; Lin, Z.; Lu, M.; Huang, L.J.; Kim, W.Y. Plant hormone-mediated regulation of heat tolerance in response to global climate change. Front. Plant Sci. 2021, 11, 2318. [Google Scholar] [CrossRef]
- Gerganova, M.; Popova, A.V.; Stanoeva, D.; Velitchkova, M. Tomato plants acclimate better to elevated temperature and high light than to treatment with each factor separately. Plant Physiol. Biochem. 2016, 104, 234–241. [Google Scholar] [CrossRef]
- Martins, S.C.V.; Araújo, W.L.; Tohge, T.; Fernie, A.R.; DaMatta, F.M. In high-light-acclimated coffee plants the metabolic machinery is adjusted to avoid oxidative stress rather than to benefit from extra light enhancement in photosynthetic yield. PLoS ONE 2014, 9, e94862. [Google Scholar] [CrossRef]
- Roda, N.d.M.; Branchi, B.A.; Longo, R.M.; Pontin, J.; Abreu, D.P.d.; Santos, P.R.d.; Campostrini, E. The Advantages of Using Kaolin-Based Particle Films to Improve Coffee Production in the Minas Gerais Cerrado Biome. Sustainability 2022, 14, 4485. [Google Scholar] [CrossRef]
- de Oliveira, R.R.; Ribeiro, T.H.C.; Cardon, C.H.; Fedenia, L.; Maia, V.A.; Barbosa, B.C.F.; Caldeira, C.F.; Klein, P.E.; Chalfun-Junior, A. Elevated Temperatures Impose Transcriptional Constraints and Elicit Intraspecific Differences Between Coffee Genotypes. Front. Plant Sci. 2020, 11, 1113. [Google Scholar] [CrossRef]
- Rodrigues, W.P.; Silva, J.R.; Ferreira, L.S.; Machado Filho, J.A.; Figueiredo, F.A.M.M.A.; Ferraz, T.M.; Bernado, W.P.; Bezerra, L.B.S.; Abreu, D.P.; Cespom, L.; et al. Stomatal and photochemical limitations of photosynthesis in coffee (Coffea spp.) plants subjected to elevated temperatures. Crop Pasture Sci. 2018, 69, 317–325. [Google Scholar] [CrossRef]
- Vilas-Boas, T.; Duarte, A.A.; Della Torre, F.; Lovato, M.B.; Lemos-Filho, J.P. Does acclimation in distinct light conditions determine differences in the photosynthetic heat tolerance of coffee plants? Plant Biol. 2023, 25, 1101–1108. [Google Scholar] [CrossRef]
- Thioune, E.-H.; McCarthy, J.; Gallagher, T.; Osborne, B. A humidity shock leads to rapid, temperature dependent changes in coffee leaf physiology and gene expression. Tree Physiol. 2017, 37, 367–379. [Google Scholar]
- Liu, X.G.; Wan, M.D.; Wu, H.; Yang, Q.L. Photosynthetic response and use of water and light of Arabica coffee leaf under different irrigation and light levels. Oxid. Commun. 2016, 39, 873–883. [Google Scholar]
- Pompelli, M.F.; Martins, S.C.V.; Antunes, W.C.; Chaves, A.R.M.; DaMatta, F.M. Photosynthesis and photoprotection in coffee leaves is affected by nitrogen and light availabilities in winter conditions. J. Plant Physiol. 2010, 167, 1052–1060. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-López, N.F.; Cavatte, P.C.; Silva, P.E.M.; Martins, S.C.V.; Morais, L.E.; Medina, E.F.; DaMatta, F.M. Physiological and biochemical abilities of robusta coffee leaves for acclimation to cope with temporal changes in light availability. Physiol. Plant. 2013, 149, 4555. [Google Scholar] [CrossRef]
- FAO—Food and Agriculture Organization of the United Nations. Soil Letters Salt-Affected Soils Are a Global Issue. Intergovernmental Technical Panel on Soils. 2021. Available online: https://openknowledge.fao.org/server/api/core/bitstreams/8b5c687f-5a9b-4034-a4e2-0457066c1ae3/content (accessed on 16 July 2024).
- Majeed, A.; Muhammad, Z. Salinity: A major agricultural problem—Causes, impacts on crop productivity and management strategies. In Plant Abiotic Stress Tolerance; Hasanuzzaman, M., Hakeem, K., Nahar, K., Alharby, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 83–99. [Google Scholar]
- Zaman, M.; Shahid, S.A.; Heng, L. Guideline for Salinity Assessment, Mitigation and Adaptation Using Nuclear and Related Techniques; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Chhabra, R. Classification of salt-affected soils. Arid Land Res. Manag. 2004, 19, 61–79. [Google Scholar] [CrossRef]
- Yadav, S.; Irfan, M.; Ahmad, A.; Hayat, S. Causes of salinity and plant manifestations to salt stress: A review. J. Environ. Biol. 2011, 32, 667–685. [Google Scholar]
- Hnilickova, H.; Hnilicka, F.; Martinkova, J.; Kraus, K. Effects of Salt Stress on Water Status; Photosynthesis and Chlorophyll Fluorescence of Rocket. Plant Soil Environ. 2017, 63, 362–367. [Google Scholar] [CrossRef]
- Mbarki, S.; Sytar, O.; Cerda, A.; Zivcak, M.; Rastogi, A.; He, X.; Zoghlami, A.; Abdelly, C.; Brestic, M. Strategies to mitigate the salt stress effects on photosynthetic apparatus and productivity of crop plants. In Salinity Responses and Tolerance in Plants; Kumar, V., Wani, S., Suprasanna, P., Tran, L.S., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; Volume 1, pp. 85–136. [Google Scholar]
- Lima, R.B.; dos Santos, T.B.; Vieira, L.G.E.; Ferrarese, M.D.L.L.; Ferrarese-Filho, O.; Donatti, L.; Petkowicz, C.L.O. Salt stress alters the cell wall polysaccharides and anatomy of coffee (Coffea arabica L.) leaf cells. Carbohydr. Polym. 2014, 112, 686–694. [Google Scholar] [CrossRef]
- Oyebamiji, Y.O.; Adigun, B.A.; Shamsudin, N.A.A.; Ikmal, A.M.; Salisu, M.A.; Malike, F.A.; Lateef, A.A. Recent Advancements in Mitigating Abiotic Stresses in Crops. Horticulturae 2024, 10, 156. [Google Scholar] [CrossRef]
- Lehmann, J.; Amonette, J.E.; Roberts, K. Role of biochar in mitigation of climate change. In Handbook of Climate Change and Agroecosystems; Hillel, D., Rosenzweig, C., Eds.; Imperial College Press: London, UK, 2010; Volume 1, pp. 343–363. [Google Scholar]
- Jeffery, S.; Verheijen, F.G.; Kammann, C.; Abalos, D. Biochar effects on methane emissions from soils: A meta-analysis. Soil Biol. Biochem. 2016, 101, 251–258. [Google Scholar] [CrossRef]
- Amoakwah, E.; Frimpong, K.A.; Okae-Anti, D.; Arthur, E. Soil water retention, air flow and pore structure characteristics after corn cob biochar application to a tropical sandy loam. Geoderma 2017, 307, 189–197. [Google Scholar] [CrossRef]
- Sorrenti, G.; Masiello, C.A.; Dugan, B.; Toselli, M. Biochar physico-chemical properties as affected by environmental exposure. Sci. Total Environ. 2016, 563–564, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Zulfiqar, F.; Wei, X.; Shaukat, N.; Chen, J.; Raza, A.; Younis, A.; Nafees, M.; Abideen, Z.; Zaid, A.; Latif, N.; et al. Effects of biochar and biochar-compost mix on growth, performance and physiological responses of potted Alpinia zerumbet. Sustainability 2021, 13, 11226. [Google Scholar] [CrossRef]
- Ahmad, M.; Lee, S.S.; Lee, S.E.; Al-Wabel, M.I.; Tsang, D.C.W.; Ok, Y.S. Biochar-induced changes in soil properties affected immobilization/mobilization of metals/metalloids in contaminated soils. J. Soils Sediments 2017, 17, 717–730. [Google Scholar] [CrossRef]
- Reyes-Herrera, D.F.; Sánchez-Reinoso, A.D.; Lombardini, L.; Restrepo-Díaz, H. Physiological Responses of Coffee (Coffea arabica L.) Plants to Biochar Application under Water Deficit Conditions. Not. Bot. Horti Agrobot. Cluj-Napoca 2023, 51, 12873. [Google Scholar] [CrossRef]
- Wan, J.; Zhang, P.; Wang, R.; Sun, L.; Ju, Q.; Xu, J. Comparative physiological responses and transcriptome analysis reveal the roles of melatonin and serotonin in regulating growth and metabolism in Arabidopsis. BMC Plant Biol. 2018, 18, 362. [Google Scholar] [CrossRef]
- Zhao, C.; Nawaz, G.; Cao, Q.; Xu, T. Melatonin is a potential target for improving horticultural crop resistance to abiotic stress. Sci. Hortic. 2022, 291, 110560. [Google Scholar] [CrossRef]
- Ayyaz, A.; Shahzadi, A.K.; Fatima, S.; Yasin, G.; Zafar, Z.U.; Athar, H.U.R.; Farooq, M.A. Uncovering the role of melatonin in plant stress tolerance. Theor. Exp. Plant Physiol. 2022, 34, 335–346. [Google Scholar] [CrossRef]
- Weeda, S.; Zhang, N.; Zhao, X.; Ndip, G.; Guo, Y.; Buck, G.A.; Fu, C.; Ren, S. Arabidopsis transcriptome analysis reveals key roles of melatonin in plant defense systems. PLoS ONE 2014, 9, e93462. [Google Scholar] [CrossRef]
- Wei, Y.; Zeng, H.; Hu, W.; Chen, L.; He, C.; Shi, H. Comparative transcriptional profiling of melatonin synthesis and catabolic genes indicates the possible role of melatonin in developmental and stress responses in rice. Front. Plant Sci. 2016, 7, 676. [Google Scholar] [CrossRef]
- Ye, J.; Wang, S.; Deng, X.; Yin, L.; Xiong, B.; Wang, X. Melatonin increased maize (Zea mays L.) seedling drought tolerance by alleviating drought-induced photosynthetic inhibition and oxidative damage. Acta Physiol. Plant. 2016, 38, 48. [Google Scholar]
- Campos, C.N.; Ávila, R.G.; Souza, K.R.D.; Azevedo, L.M.; Alves, J.D. Melatonin reduces oxidative stress and promotes drought tolerance in young Coffea arabica L. plants. Agric. Water Manag. 2019, 211, 37–47. [Google Scholar] [CrossRef]
- Sseremba, G.; Tongoona, P.B.; Musoli, P.; Eleblu, J.S.Y.; Melomey, L.D.; Bitalo, D.N.; Atwijukire, E.; Mulindwa, J.; Aryatwijuka, N.; Muhumuza, E.; et al. Viability of Deficit Irrigation Pre-Exposure in Adapting Robusta Coffee to Drought Stress. Agronomy 2023, 13, 674. [Google Scholar] [CrossRef]
- Silva, V.A.; Antunes, W.C.; Guimarães, B.L.S.; Paiva, R.M.C.; Silva, V.F.; Ferrão, M.A.G.; DaMatta, F.M.; Loureiro, M.E. Physiological response of Conilon coffee clone sensitive to drought grafted onto tolerant rootstock. Pesqui. Agropecu. Bras. 2010, 45, 457–464. [Google Scholar]
- Teixeira, A.L.; Souza, F.D.F.; Pereira, A.A.; Oliveira, A.C.B.D.; Rocha, R.B. Selection of arabica coffee progenies tolerant to heat stress. Cienc. Rural 2015, 45, 1228–1234. [Google Scholar] [CrossRef]
- Naik, B.J.; Kim, S.C.; Seenaiah, R.; Basha, P.A.; Song, E.Y. Coffee cultivation techniques, impact of climate change on coffee production, role of nanoparticles and molecular markers in coffee crop improvement, and challenges. J. Plant Biotechnol. 2021, 48, 207–222. [Google Scholar] [CrossRef]
- Mofatto, L.S.; Carneiro, F.D.A.; Vieira, N.G.; Duarte, K.E.; Vidal, R.O.; Alekcevetch, J.C.; Cotta, M.G.; Verdeil, J.-L.; Fabienne, L.-M.; Lartaud, M.; et al. Identification of candidate genes for drought tolerance in coffee by high-throughput sequencing in the shoot apex of different Coffea arabica cultivars. BMC Plant Biol. 2016, 16, 94. [Google Scholar] [CrossRef]
- De Carvalho, K.; Petkowicz, C.L.O.; Nagashima, G.T.; Bespalhok Filho, J.C.; Vieira, L.G.E.; Pereira, L.F.P.; Domingues, D.S. Homeologous genes involved in mannitol synthesis reveal unequal contributions in response to abiotic stress in Coffea arabica. Mol. Genet. Genom. 2014, 289, 951–963. [Google Scholar] [CrossRef]
- Atkinson, N.J.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3544. [Google Scholar] [CrossRef]
- Torres, L.F.; Reichel, T.; Déchamp, E.; de Aquino, S.O.; Duarte, K.E.; Alves, G.S.C.; Silva, A.T.; Cotta, M.G.; Costa, T.S.; Diniz, L.E.C.; et al. Expression of DREB-like genes in Coffea canephora and C. arabica subjected to various types of abiotic stress. Trop. Plant Biol. 2019, 12, 98–116. [Google Scholar] [CrossRef]
- Duque, A.S.; Almeida, A.M.; Bernardes da Silva, A.; Marques da Silva, J.; Farinha, A.P.; Santos, D.; Fevereiro, P.; Araújo, S.S. Abiotic stress responses in plants: Unraveling the complexity of genes and networks to survive. In Abiotic Stress: Plant Responses and Applications in Agriculture; Vahdati, K., Leslie, C., Eds.; INTECH Open: Rijeka, Croatia; London, UK, 2013; pp. 49–102. [Google Scholar]
- Mishra, M.K.; Slater, A. Recent Advances in the Genetic Transformation of Coffee. Biotechnol. Res. Int. 2012, 2012, 580857. [Google Scholar] [CrossRef]
- Vats, S.; Kumawat, S.; Kumar, V.; Patil, G.B.; Joshi, T.; Sonah, H.; Sharma, T.R.; Deshmukh, R. Genome editing in plants: Exploration of technological advancements and challenges. Cell 2019, 8, 1386. [Google Scholar] [CrossRef] [PubMed]
- Breitler, J.-C.; Dechamp, E.; Campa, C.; Rodrigues, L.A.Z.; Guyot, R.; Marraccini, P.; Etienne, H. CRISPR/Cas9-mediated efficient targeted mutagenesis has the potential to accelerate the domestication of Coffea canephora. Plant Cell Tissue Organ Cult. 2018, 134, 383–394. [Google Scholar] [CrossRef]
- Casarin, T.; Freitas, N.C.; Pinto, R.T.; Breitler, J.-C.; Rodrigues, L.A.Z.; Marraccini, P.; Etienne, H.; Diniz, L.E.C.; Andrade, A.C.; Paiva, L.V. Multiplex CRISPR/Cas9-mediated knockout of the phytoene desaturase gene in Coffea canephora. Sci. Rep. 2022, 12, 17270. [Google Scholar] [CrossRef]
- Waraich, E.A.; Ahmad, R.; Ashraf, M.Y.; Saifullah Ahmad, M. Improving agricultural water use efficiency by nutrient management in crop plants. Acta Agric. Scand. B Soil. Plant Sci. 2011, 61, 291–304. [Google Scholar] [CrossRef]
- He, M.; Dijkstra, F.A. Drought effect on plant nitrogen and phosphorus: A meta-analysis. New Phytol. 2014, 204, 924–931. [Google Scholar] [CrossRef]
- Silva, E.C.; Nogueira, R.J.M.C.; Silva, M.A.; Albuquerque, M.B. Drought Stress and Plant Nutrition. Plant Stress 2011, 5, 32–41. [Google Scholar]
- Martinez, H.E.P.; Souza, B.P.; Caixeta, E.T.; Carvalho, F.P.; Clemente, J.M. Water deficit changes nitrate uptake and expression of some nitrogen related genes in coffee-plants (Coffea arabica L.). Sci. Hortic. 2020, 267, 109254. [Google Scholar] [CrossRef]
- Rabêlo, F.H.S. Role of Plant Nutrition in Mitigating Biotic and Abiotic Stresses in Plants. Inform. Agron. 2024, 2, 5–22. (In Portuguese) [Google Scholar]
- Andrade, O.V.S.; Lima, J.S.; Neves, T.T.; Benevenute, P.A.N.; Santos, L.C.; Nascimento, V.L.; Guilherme, L.R.G.; Marchiori, P.E.R. The role of potassium iodate in mitigating the damages of water deficit in coffee plants. J. Soil Sci. Plant Nutr. 2024, 24, 5772–5788. [Google Scholar] [CrossRef]
- Acidri, R.; Sawai, Y.; Sugimoto, Y.; Sasagawa, D.; Masunaga, T.; Yamamoto, S.; Nishihara, E. Foliar nitrogen supply enhances the recovery of photosynthetic performance of cold-stressed coffee (Coffea arabica L.) seedlings. Photosynthetica 2020, 58, 951–960. [Google Scholar] [CrossRef]
- Nunes, M.A.; Ramalho, J.D.C.; Dias, M.A. Effect of Nitrogen Supply on the Photosynthetic Performance of Leaves from Coffee Plants Exposed to Bright Light. J. Exp. Bot. 1993, 44, 893–899. [Google Scholar] [CrossRef]
- Salamanca-Jimenez, A.; Doane, T.A.; Horwath, W.R. Performance of Coffee Seedlings as Affected by Soil Moisture and Nitrogen Application. Adv. Agron. 2016, 136, 221–244. [Google Scholar]
- Vinecky, F.; Davrieux, F.; Mera, A.C.; Alves, G.S.C.; Lavagnini, G.; Leroy, T.; Bonnot, F.; Rocha, O.C.; Bartholo, G.F.; Guerra, A.F.; et al. Controlled irrigation and nitrogen, phosphorous and potassium fertilization affect the biochemical composition and quality of Arabica coffee beans. J. Agric. Sci. 2017, 155, 902–918. [Google Scholar] [CrossRef]
- Ramirez-Builes, V.H.; Kusters, J.; Thiele, E.; Lopez-Ruiz, J.C. Physiological and Agronomical Response of Coffee to Different Nitrogen Forms with and without Water Stress. Plants 2024, 13, 1387. [Google Scholar] [CrossRef]
- Rocha, B.C.P.; Martinez, H.E.P.; Ribeiro, C.; Brito, D.S. Nitrogen Metabolism in Coffee Plants Subjected to Water Deficit and Nitrate Doses. Braz. Arch. Biol. Technol. 2023, 66, e23210060. [Google Scholar]
- Rakocevic, M.; Marchiori, P.E.R.; Zambrosi, F.C.B.; Machado, E.C.; Maia, A.H.N.; Ribeiro, R.V. High phosphorus supply enhances leaf gas exchange and growth of young Arabica coffee plants under water deficit. Exp. Agric. 2022, 58, e30. [Google Scholar] [CrossRef]
- Ramírez-Builes, V.H.; Küsters, J.; de Souza, T.R.; Simmes, C. Calcium Nutrition in Coffee and Its Influence on Growth, Stress Tolerance, Cations Uptake, and Productivity. Front. Agron. 2020, 2, 590892. [Google Scholar] [CrossRef]
- Silva, D.M.; Souza, K.R.D.; Boas, L.V.V.; Alves, Y.S.; Alvez, J.D. The effect of magnesium nutrition on the antioxidant response of coffee seedlings under heat stress. Sci. Hortic. 2017, 224, 115–125. [Google Scholar] [CrossRef]
- Dias, K.G.L.; Guimarães, P.T.G.; Furtini Neto, A.E.; Silveira, H.R.O.; Lacerda, J.J.J. Effect of Magnesium on Gas Exchange and Photosynthetic Efficiency of Coffee Plants Grown under Different Light Levels. Agriculture 2017, 7, 85. [Google Scholar] [CrossRef]
- Ramirez-Builes, V.H.; Küsters, J.; Thiele, E.; Leal-Varon, L.A. Boron Nutrition in Coffee Improves Drought Stress Resistance and, Together with Calcium, Improves Long-Term Productivity and Seed Composition. Agronomy 2024, 14, 474. [Google Scholar] [CrossRef]
- de Sousa, G.F.; Silva, M.A.; de Morais, E.G.; Van Opbergen, G.A.Z.; Van Opbergen, G.G.A.Z.; de Oliveira, R.R.; Amaral, D.; Brown, P.; Chalfun-Junior, A.; Guilherme, L.R.G. Selenium enhances chilling stress tolerance in coffee species by modulating nutrient, carbohydrates, and amino acids content. Front. Plant Sci. 2022, 13, 1000430. [Google Scholar] [CrossRef] [PubMed]
- Sousa, G.F.d.; Silva, M.A.; Carvalho, M.R.d.; Morais, E.G.d.; Benevenute, P.A.N.; Van Opbergen, G.A.Z.; Van Opbergen, G.G.A.Z.; Guilherme, L.R.G. Foliar Selenium Application to Reduce the Induced-Drought Stress Effects in Coffee Seedlings: Induced Priming or Alleviation Effect? Plants 2023, 12, 3026. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species/Genotype | Growth Conditions | Main Changes Induced by Drought Stress | Tolerance Mechanisms for Stress Adaptation | References |
|---|---|---|---|---|
| C. arabica Catuaí LCH 2077-2-5-86 | Leaf water potential (Ψpd) of −2.4 MPa | Increased leaf osmotic potential and reduction in the specific leaf area. | Maintenance of a high relative water content. | [21] |
| C. arabica cv. Catuaí Vermelho | Well-watered plants, water deficit plants (Ψpd from −0.2 MPa to −2.0 MPa), and re-watered plants (plants subjected to water deficit followed by irrigation) | Decreased foliar and flower bud ethylene production without changing root ethylene production. | Changes in ethylene levels and sensitivity to promote coffee anthesis. | [22] |
| C. arabica cv. Catuaí Vermelho IAC 44 | 30 and 100% of soil-available water | Decreased relative growth rates and reduced gas exchange parameters. | Increased proline concentration. | [23] |
| C. arabica Mundo Novo LCH 464-18 | Ψpd of −2.4 MPa | Increased leaf osmotic potential and reduction in the specific leaf area. | Maintenance of a high relative water content. | [21] |
| C. arabica cv. Red Catuai | Ψpd of approximately −1.5 and −2.7 MPa | Net photosynthetic rate decreased because of non-stomatal limitations. | - | [24] |
| C. arabica Red Catuaí LCH 2077-2-5-44 | Ψpd of −2.4 MPa | Increased leaf osmotic potential and reduction in the specific leaf area. | Maintenance of a high relative water content. | [21] |
| C. arabica genotype 07 (progeny from Paraíso) | Ψpd of −3.0 MPa followed by rehydration | Increase in the number of root hairs and reductions in the root cross-section area, cortex thickness, and tracheary element diameter. | Greater root plasticity, which increased hydraulic conductivity under water deficit, contributed to the restoration of photosynthetic efficiency after the return of water availability. | [25] |
| C. arabica genotype 19 (progeny from Araponga) | Ψpd of −1.3 MPa followed by rehydration | Increase in the vascular cylinder area, in number of metaxylem poles, and in number of tracheary elements. | Greater root plasticity, which increased hydraulic conductivity under water deficit, contributed to the restoration of photosynthetic efficiency after the return of water availability. | [25] |
| Several genotypes of C. arabica | Well-watered (soil moisture of 60–80% field water capacity) and drought-stressed plants (seedlings were initially fully irrigated and subsequently subjected to drought conditions by withholding water until the end of the experiment—around 300 days of coffee age). | Lower chlorophyll a and b contents. | Elevations in glucose, maltose, amino acids, and organic acids concentrations. | [26] |
| Several genotypes of C. arabica | Daily irrigation | Tolerant genotypes showed quicker and more complete germination compared to the sensitive group. | Higher moisture content, higher seed surface area-to-volume ratio, and higher coefficient of velocity of germination, coefficient of variation in germination time, and germination index in the tolerant genotypes compared to sensitive genotypes. | [27] |
| Several genotypes of C. arabica | Well-watered (continued watering) and water deficit plants (30 days by withholding watering followed by re-watering for 15 days) | Reduction in biomass accompanied by a higher tap root diameter, lateral root number, root volume, specific root length, and root–mass ratio in the drought-tolerant genotypes. | - | [28] |
| C. canephora var. kouillou | Ψpd of approximately −2.0 and −3.0 MPa | Drought led to sharper decreases in stomatal conductance than in photosynthesis, which was accompanied by significant declines in the internal-to-ambient CO2 concentration ratio. Starch decreased remarkably. | Better protection against oxidative stress and maintenance of SPS (EC 2.4.1.14) activity with rising leaf water deficit. This might be associated with assimilate export, which would allow for additional root growth under limited water supply. | [29] |
| C. canephora var. kouillou | Ψpd of approximately −1.5 and −2.7 MPa | Net photosynthetic rate decreased mainly as a consequence of stomatal limitations. | Maintained carbon gain at the expense of water conservation. | [24] |
| C. canephora cv. Robusta | Ψpd of −2.4 MPa | Increased leaf osmotic potential and reduction in the specific leaf area. | Maintenance of a high relative water content and increase in the maximum bulk modulus of elasticity. | [21] |
| Plant Species/Genotype | Growth Conditions | Main Changes Induced by Heat and Excess Light Stress | Tolerance Mechanisms for Stress Adaptation | References |
|---|---|---|---|---|
| Heat stress | ||||
| C. arabica cv. Acauã | 23/19 °C or 30/26 °C (day/night temperatures) | Higher leaf transpiration. | Adjustment in plant hormone signal transduction, carbon metabolism, and carbohydrate metabolism. | [42] |
| C. arabica cv. Catuaí Amarelo IAC 62 | Adequate temperature (spring) and high temperature (summer) | Exposure to summer conditions decreased photosynthetic rates, stomatal conductance, and stomatal density, and increased intrinsic water use efficiency compared with spring values. | Maintenance of Fv/Fm, electron transport rates, and membrane leakage values. | [43] |
| C. arabica cv. Catuaí Amarelo | Control plants (15 °C) and stressed plants (50 °C) | 50% reduction in Fv/Fm. | Larger leaves and a lower fraction of leaf area occupied by stomatal pores. | [44] |
| C. arabica cv. Catuaí IAC 144 | 23/19 °C or 30/26 °C (day/night temperatures) | Higher leaf transpiration. | Adjustment in carbohydrate metabolism. | [42] |
| C. arabica cv. IAPAR-59 | Control plants (24 °C) and stressed plants (37 °C) | The primary monolignol contents increased after the heat stress. Structural alterations in palisade cells and ultrastructural damage in chloroplasts were also observed. | Cell wall re-establishment through biosynthesis or assembly of major cell wall compounds. | [9] |
| C. arabica cv. Mundo Novo | Control plants (15 °C) and stressed plants (50 °C) | 50% reduction in Fv/Fm. | Larger leaves and a lower fraction of leaf area occupied by stomatal pores. | [44] |
| C. canephora cv. Conilon | Control plants (15 °C) and stressed plants (50 °C) | 50% reduction in Fv/Fm. | Larger leaves and a lower fraction of leaf area occupied by stomatal pores. | [44] |
| C. canephora cv. Conilon Emcapa 8111 Clone 02 | Adequate temperature (spring) and high temperature (summer) | - | Stomatal insensitivity to the imposed conditions, maintaining both stomatal conductance and photosynthetic rates constant. | [43] |
| C. canephora var. FRT07 | Relative humidity (RH) of 30% and 27 °C, RH of 30% and 35 °C, and RH of 30% and 42 °C | At 27 and 35 °C, upper leaves showed decreases in Fv/Fm compared with lower leaves, while at 42 °C, both upper and lower leaves were equally affected. | Induction of genes involved with abscisic acid signaling, such as CcNCED3, at 42 °C. | [45] |
| Light stress | ||||
| C. arabica | Natural light (100% transmittance), 50% transmittance, and 18% transparency | Increasing shade levels increased total chlorophyll and chlorophyll b content, maximum net photosynthetic rate, and stomatal conductance, but it significantly decreased the daily average transpiration rate. | - | [46] |
| C. arabica cv. Catuaí Vermelho IAC 44 | Full (100%) sunlight conditions or low light (10% full sunlight) in a shade environment | High energetic coast to cope with excessive light. | Increases in the antioxidant capacity associated with enhanced consumption of reducing equivalents. | [40] |
| C. arabica cv Red Catuaí IAC 44 | Full sunlight and shade (50%) conditions | Low net carbon assimilation rate. | Increased antheraxanthin and zeaxanthin pools linked to increased thermal dissipation of absorbed light. | [47] |
| C. canephora | Bushes shaded in the morning, bushes under full sunlight, and bushes shaded in the afternoon | Changes in gas exchange measurements and in the activities of antioxidant enzymes [SOD (EC 1.15.1.1), APX (EC 1.11.1.11), CAT (EC 1.11.1.6), and GR (EC 1.6.4.2)]. | Higher mass-based light-saturated photosynthetic rates, photosynthetic nitrogen use efficiencies, and long-term water use efficiencies. | [48] |
| Elements | Plant Species/Genotype | Dose and Application Mode | Stress | Changes Induced by Element Application | References |
|---|---|---|---|---|---|
| N | C. arabica | 0, 5, 10, 20, and 40 mM N applied via foliar | Cold | Increased nitrogen concentration and photosynthetic pigments, recovery of the photosynthetic apparatus and increased net carbon assimilation rate, increased efficiency of PSII, and decreased 5-caffeoylquinic acid, mangiferin, trigonelline, and caffeine contents in the coffee beans. | [91] |
| C. arabica cv. Catuaí | 0, 1, and 2 mM NH4NO3 applied via soil | High and low light | Decreased reduction in leaf area, increase in net photosynthesis and stomatal conductance, and increase in protein and chlorophyll contents. | [92] | |
| C. arabica cv. line CX 2720 | 0, 0.1, 0.2, and 0.4 g/plant N applied via soil | Water deficit | Increased nitrogen content, water use efficiency, foliar area, chlorophyll and carotenoids contents, and shoot biomass production. | [93] | |
| C. arabica cv. Red Catuaí IAC 44 | 0 and 23 mM N applied via nutrient solution | High and low light | Increased amino acids and total protein contents under low light, increase in chlorophyll a and b and carotenoids contents, and increased APX activity under low and high light. | [47] | |
| C. arabica cv. Rubi | 0, 100, 250, 500, and 800 kg N ha−1 applied as urea via soil | Water availability | Increased caffeine content in the coffee beans. | [94] | |
| C. arabica cv. Topazio MG 1190 | From 21.4 to 28.6 mM N (100% NH4+-N/0% NO3−-N, 75% NH4+-N/25% NO3−-N, 50% NH4+-N/50% NO3−-N, 25% NH4+-N/75% NO3−-N, and 0% NH4+-N/100% NO3−-N) applied via nutrient solution | Water deficit | Nitrogen ratios of 50% NH4+-N/50% NO3−-N, and 25% NH4+-N/75% NO3−-N showed better responses to water stress, by increasing photosynthesis, chlorophyll content, nitrogen and cation uptake, and coffee yield. | [95] | |
| Many genotypes of C. arabica | 2.8 and 7.0 mmol L−1 NO3− applied via nutrient solution | Water deficit | Good nitrogen nutrition increased amino acids and proteins in the leaves of coffee. | [96] | |
| P | C. arabica cv. Ouro Verde | 345 and 690 mg P2O5 dm−3 soil | Water deficit | Higher phosphorus concentration, photosynthesis, and plant growth. | [97] |
| K | C. arabica cv. Rubi | 0, 100, 250, 500, and 800 kg K2O ha−1 applied via soil | Water availability | Increased lipid and chlorogenic acid in the coffee beans. | [94] |
| Ca | C. arabica | 0, 70, 140, and 210 mg of soluble Ca+2 L−1 applied via roots | Drought | Increased photosynthesis and biomass accumulation. | [98] |
| C. arabica | 8, 40, and 80 mg of soluble Ca+2 L−1 applied via roots | Heat | Increased photosynthesis and thicker leaves, denser epidermis, and larger, more compact, and better-structured palisade parenchyma. | [98] | |
| Mg | C. arabica cv. Catuaí 144 | 0 or 1 mmol L−1 of MgSO4 applied via nutrient solution | Heat | Lower production of hydrogen peroxide and poor lipid peroxidation and protein denaturation in the plant leaves. | [99] |
| C. arabica cv. Mundo Novo IAC 379/19 | 0, 48, 96, 192, and 384 mg Mg2+ L−1 applied via nutrient solution | Low light | Increased CO2 assimilation rate and water use efficiency, and reduced leaf transpiration. | [100] | |
| B | C. arabica var. Cenicafé 1 | 0 and 12.2 mg of B per plant applied via soil as calcium nitrate+B | Drought | Increased boron concentration and biomass production, and decreased mainly, oleic, linoleic, and linolenic acids contents in the green coffee beans. | [101] |
| Se | C. arabica cv. Arara | 0 and 80 mg Se L−1 applied as Na2SeO4 via foliar | Chilling | Increased CAT activity during stress in the plant leaves. | [102] |
| C. arabica cv. Catuaí | Foliar spray with 5 mL of Se solution at 80 mg L−1 Se + 0.5% of mineral oil (+Se) or foliar spray of 0.5% of mineral oil (-Se), using Na2SeO4 | Drought | Higher CAT, APX, and SOD activities in the plant leaves. | [103] | |
| C. canephora clone 31 | 0 and 80 mg Se L−1 applied as Na2SeO4 via foliar | Chilling | Increased starch, amino acids, and proline contents, and increased nitrogen content in the leaves during plant stress. | [102] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borgo, L.; Rabêlo, F.H.S.; Marchiori, P.E.R.; Guilherme, L.R.G.; Guerra-Guimarães, L.; Resende, M.L.V.d. Impact of Drought, Heat, Excess Light, and Salinity on Coffee Production: Strategies for Mitigating Stress Through Plant Breeding and Nutrition. Agriculture 2025, 15, 9. https://doi.org/10.3390/agriculture15010009
Borgo L, Rabêlo FHS, Marchiori PER, Guilherme LRG, Guerra-Guimarães L, Resende MLVd. Impact of Drought, Heat, Excess Light, and Salinity on Coffee Production: Strategies for Mitigating Stress Through Plant Breeding and Nutrition. Agriculture. 2025; 15(1):9. https://doi.org/10.3390/agriculture15010009
Chicago/Turabian StyleBorgo, Lucélia, Flávio Henrique Silveira Rabêlo, Paulo Eduardo Ribeiro Marchiori, Luiz Roberto Guimarães Guilherme, Leonor Guerra-Guimarães, and Mário Lúcio Vilela de Resende. 2025. "Impact of Drought, Heat, Excess Light, and Salinity on Coffee Production: Strategies for Mitigating Stress Through Plant Breeding and Nutrition" Agriculture 15, no. 1: 9. https://doi.org/10.3390/agriculture15010009
APA StyleBorgo, L., Rabêlo, F. H. S., Marchiori, P. E. R., Guilherme, L. R. G., Guerra-Guimarães, L., & Resende, M. L. V. d. (2025). Impact of Drought, Heat, Excess Light, and Salinity on Coffee Production: Strategies for Mitigating Stress Through Plant Breeding and Nutrition. Agriculture, 15(1), 9. https://doi.org/10.3390/agriculture15010009