Biodiversity and Vegetation Succession in Vineyards, Moravia (Czech Republic)

 , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Charactersistics of the Vineyards
2.2. Method of Vegetation Assessment
2.3. Statistical Data Evaluation
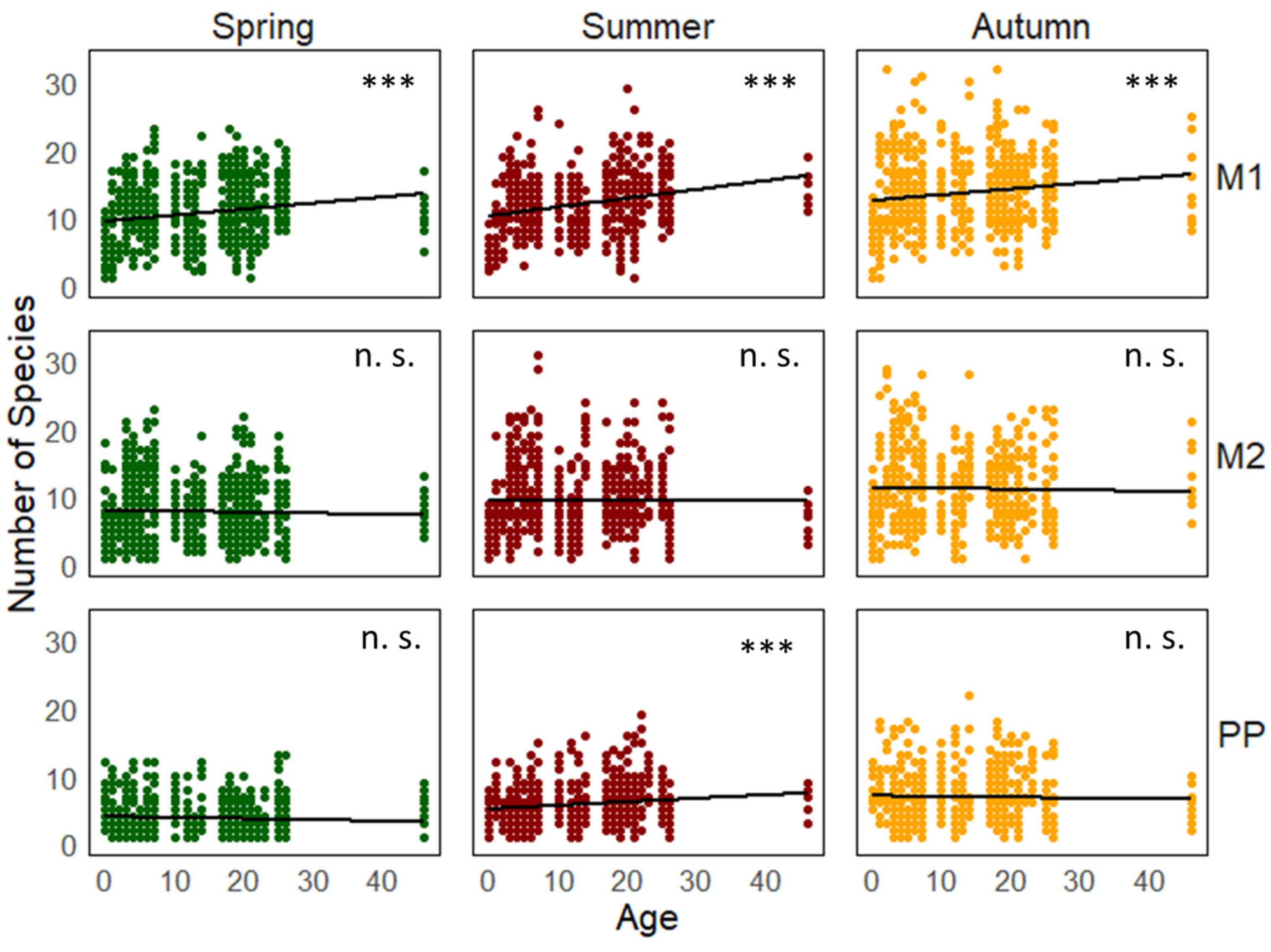
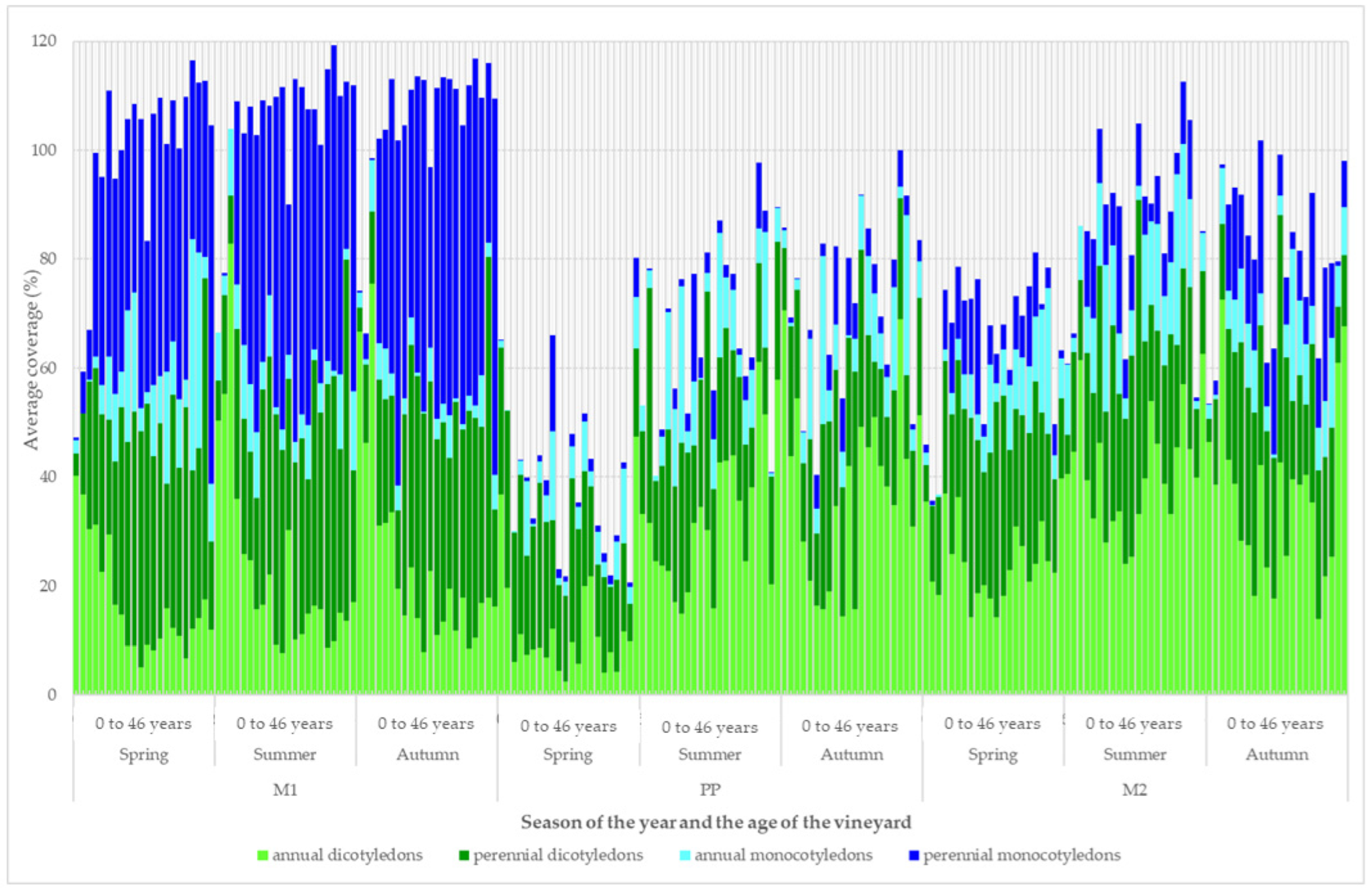
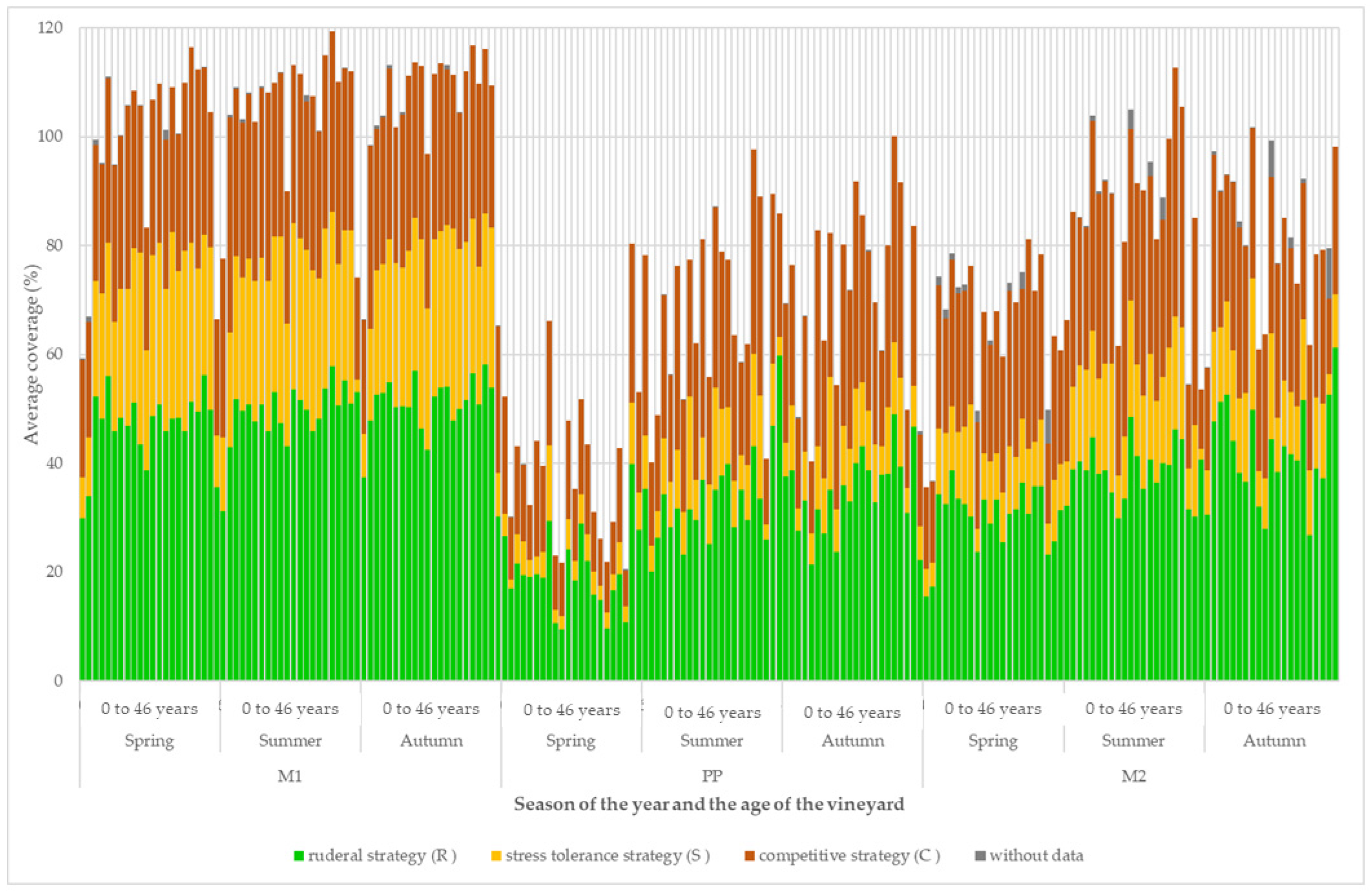
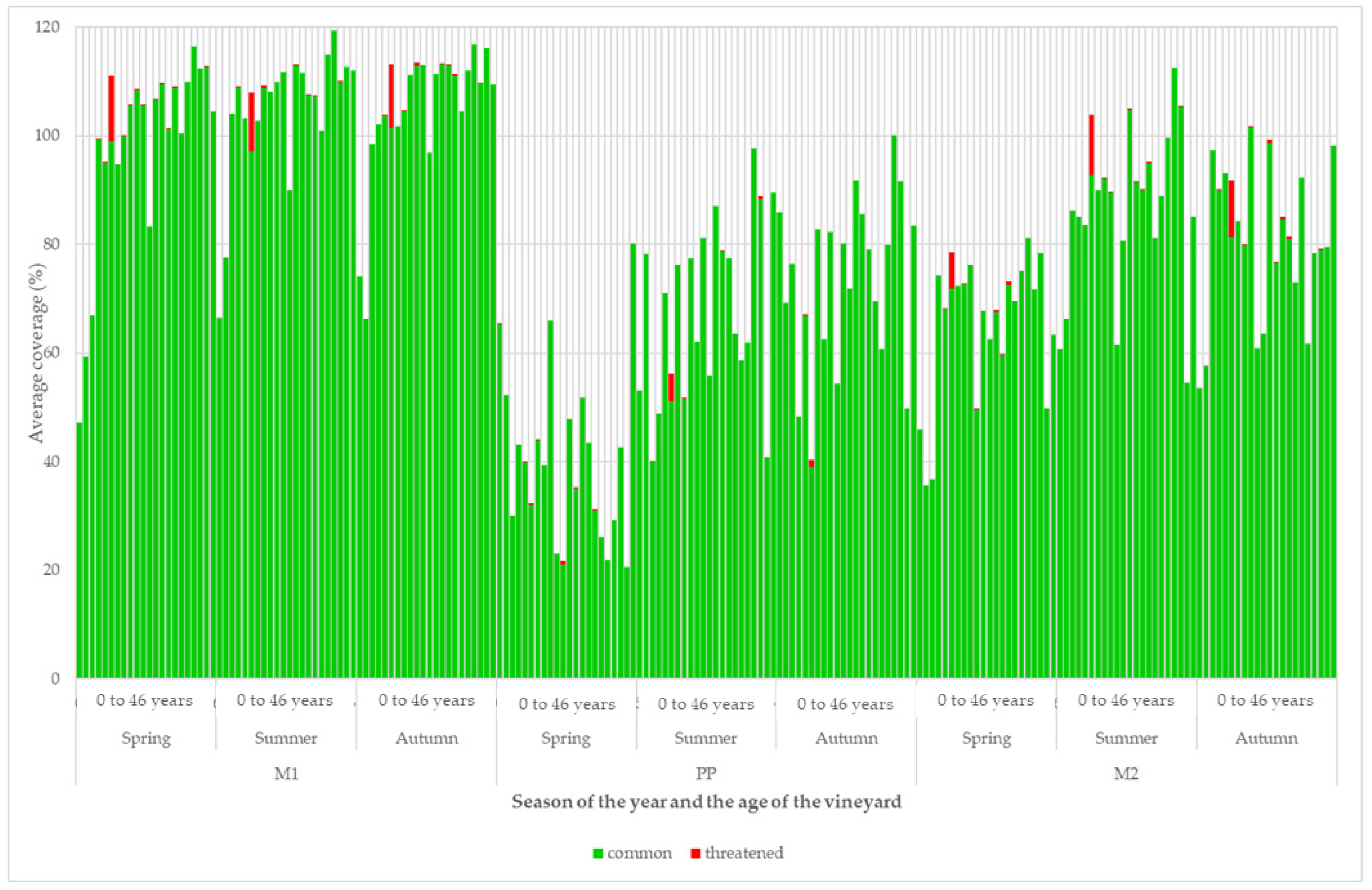
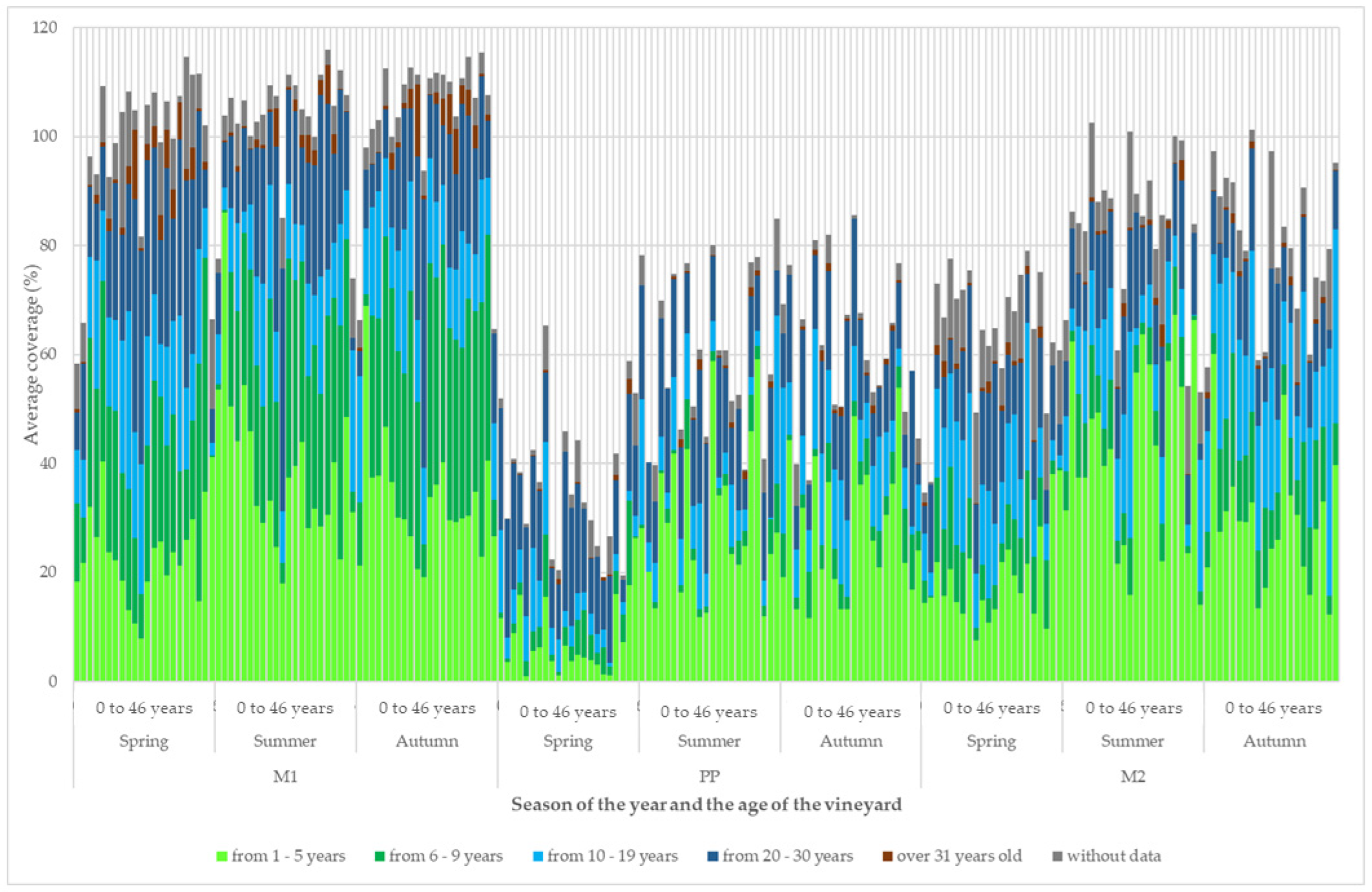
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Authorship | Family | Biological Properties | Functional Groups | Groups on Native and Invasion Status | Ecological Strategy | Treatened Species | Successional Age Optimum | ||
|---|---|---|---|---|---|---|---|---|---|---|
| (C) | (S) | (R) | ||||||||
| Agrimonia eupatoria | L. | Rosaceae | perennial dicotyledons | entomophilous species | native | 47.8 | 36.7 | 15.5 | common | 32 |
| Achillea millefolium | L. | Asteraceae | perennial dicotyledons | entomophilous species | native | 22 | 58.8 | 19.2 | common | 23 |
| Ailanthus altissima | (Mill.) Swingle | Simaroubaceae | perennial dicotyledons | deep-rooting species | invasive neophytes | 74.9 | 21.3 | 3.8 | common | 27 |
| Alcea biennis | Winterl | Malvaceae | perennial dicotyledons | entomophilous species | native | 0 | 0 | 0 | C2b | without data |
| Amaranthus powellii | S. Watson | Amaranthaceae | annual dicotyledons | annual weeds | invasive neophytes | 57.4 | 20.4 | 22.2 | common | 1.5 |
| Amaranthus retroflexus | L. | Amaranthaceae | annual dicotyledons | annual weeds | invasive neophytes | 52.1 | 23.7 | 24.2 | common | 1.5 |
| Anagallis arvensis | L. | Primulaceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 9.7 | 0 | 90.3 | common | 5 |
| Anagallis foemina | Mill. | Primulaceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 16.7 | 9.9 | 73.6 | C3 | 4 |
| Anthemis arvensis | L. | Asteraceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 21.3 | 0 | 78.4 | common | 10 |
| Anthyllis vulneraria | L. | Fabaceae | perennial dicotyledons | legumes | native | 51.6 | 4.6 | 43.8 | common | 18 |
| Apera spicaventi | (L.) P. Beauv. | Poaceae | annual monocotyledons | species with an anti-erosion effect | naturalized archaeophytes | 24.7 | 42.6 | 32.7 | common | 3 |
| Agrimonia eupatoria | L. | Rosaceae | perennial dicotyledons | entomophilous species | native | 47.8 | 36.7 | 15.5 | common | 32 |
| Arctium lappa | L. | Asteraceae | perennial dicotyledons | entomophilous species | naturalized archaeophytes | 79.1 | 7.9 | 13 | common | 8.5 |
| Arctium tomentosum | Mill. | Asteraceae | perennial dicotyledons | entomophilous species | naturalized archaeophytes | 79.1 | 7.9 | 13 | common | 8 |
| Arrhenatherum elatius | (L.) J. Presl et C. Presl | Poaceae | perennial monocotyledons | species with an anti-erosion effect | invasive archaeophytes | 23.1 | 40.3 | 36.5 | common | 25 |
| Artemisia absinthium | L. | Asteraceae | perennial dicotyledons | entomophilous species | naturalized archaeophytes | 36.9 | 21.9 | 41.2 | common | 8 |
| Artemisia vulgaris | L. | Asteraceae | perennial dicotyledons | entomophilous species | native | 41.6 | 29.6 | 28.8 | common | 10 |
| Atriplex patula | L. | Chenopodiaceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 53.9 | 0 | 46.1 | common | 4.5 |
| Atriplex sagittata | Borkh. | Chenopodiaceae | annual dicotyledons | annual weeds | invasive archaeophytes | 38.3 | 29 | 32.8 | common | 5 |
| Avena fatua | L. | Poaceae | annual monocotyledons | species with an anti-erosion effect | naturalized archaeophytes | 48.8 | 25.5 | 25.8 | common | 4 |
| Berteroa incana | (L.) DC. | Brassicaceae | perennial dicotyledons | entomophilous species | naturalized archaeophytes | 9.9 | 52.1 | 38 | common | 16.5 |
| Brassicca napus subsp. napus | L. | Brassicaceae | annual dicotyledons | crops | casual archaeophytes | 58 | 0.3 | 41.8 | common | without data |
| Bromus hordeaceus | L. | Poaceae | annual monocotyledons | species with an anti-erosion effect | naturalized archaeophytes | 11.6 | 47.2 | 41.2 | common | 12 |
| Calamagrostis epigejos | (L.) Roth | Poaceae | perennial monocotyledons | species with an anti-erosion effect | native | 40.8 | 51.2 | 8 | common | 16 |
| Camelina microcarpa | DC. | Brassicaceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 33.6 | 0 | 66.4 | common | 6.5 |
| Camelina sativa | (L.) Crantz | Brassicaceae | annual dicotyledons | crops | casual archaeophytes | 33.6 | 0 | 66.4 | common | without data |
| Capsella bursa-pastoris | (L.) Medik. | Brassicaceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 43.4 | 0 | 56.6 | common | 2 |
| Carduus acanthoides | L. | Asteraceae | perennial dicotyledons | entomophilous species | naturalized archaeophytes | 71.5 | 0 | 28.5 | common | 8 |
| Carlina vulgaris | L. | Asteraceae | annual dicotyledons | entomophilous species | native | 27.8 | 0 | 72.3 | common | 4.5 |
| Centaurea jacea | L. | Asteraceae | perennial dicotyledons | entomophilous species | native | 39.2 | 24.4 | 36.5 | common | 25 |
| Cerastium holosteoides | (Spenn.) Möschl | Caryophyllaceae | annual dicotyledons | entomophilous species | native | 0 | 9.7 | 90.3 | common | 15 |
| Cichorium intybus | L. | Asteraceae | perennial dicotyledons | entomophilous species | naturalized archaeophytes | 74.4 | 0 | 25.6 | common | 11 |
| Cirsium arvense | (L.) Scop. | Asteraceae | perennial dicotyledons | deep-rooting species | invasive archaeophytes | 80.8 | 0 | 19.2 | common | 10 |
| Consolida regalis | S. F. Gray | Ranunculaceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 20.4 | 36.9 | 42.7 | common | 4 |
| Convolvulus arvensis | L. | Convolvulaceae | perennial dicotyledons | deep-rooting species | naturalized archaeophytes | 36 | 4.7 | 59.3 | common | 22 |
| Conyza canadensis | (L.) Cronquist | Asteraceae | annual dicotyledons | annual weeds | invasive neophytes | 35.4 | 9.6 | 55.1 | common | 5 |
| Crataegus laevigata | (Poir.) DC. | Rosaceae | perennial dicotyledons | deep-rooting species | native | 18 | 56.9 | 25.1 | common | 5 |
| Crepis biennis | L. | Asteraceae | perennial dicotyledons | entomophilous species | native | 58.8 | 0 | 41.2 | common | 12.5 |
| Crepis capillaris | (L.) Wallr. | Asteraceae | perennial dicotyledons | entomophilous species | naturalized archaeophytes | 42.8 | 0 | 57.2 | common | 9.5 |
| Cynoglossum officinale | L. | Boraginaceae | perennial dicotyledons | entomophilous species | native | 58.3 | 0 | 41.7 | common | 20 |
| Dactylis glomerata | L. | Poaceae | perennial monocotyledons | species with an anti-erosion effect | native | 29 | 37.3 | 33.7 | common | 32 |
| Datura stramonium | L. | Solanaceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 71.3 | 0 | 28.7 | common | without data |
| Daucus carota | L. | Apiaceae | perennial dicotyledons | entomophilous species | native | 29.2 | 41.3 | 29.5 | common | 15 |
| Descurainia sophia | (L.) Prantl | Brassicaceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 30.2 | 10.7 | 59.1 | common | 4.5 |
| Digitaria sanguinalis | (L.) Scop. | Poaceae | annual monocotyledons | species with an anti-erosion effect | invasive archaeophytes | 17 | 3.5 | 79.5 | common | 2 |
| Echinochloa crus-galli | (L.) P. Beauv. | Poaceae | annual monocotyledons | species with an anti-erosion effect | invasive archaeophytes | 41.8 | 12.4 | 45.8 | common | 1 |
| Echinops sphaerocephalus | L. | Asteraceae | perennial dicotyledons | entomophilous species | invasive neophytes | 0 | 0 | 0 | common | 16 |
| Echium vulgare | L. | Boraginaceae | perennial dicotyledons | entomophilous species | native | 78.1 | 0 | 21.9 | common | 14.5 |
| Elymus repens | (L.) Gould | Poaceae | perennial monocotyledons | species with an anti-erosion effect | native | 33.8 | 35.9 | 30.3 | common | 8 |
| Erigeron annuus | (L.) Pers. | Asteraceae | annual dicotyledons | annual weeds | invasive neophytes | 41.3 | 13 | 45.8 | common | 30 |
| Erodium cicutarium | (L.) L‘Hér. | Geraniaceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 37.7 | 18.6 | 43.7 | common | 6 |
| Eryngium campestre | L. | Apiaceae | perennial dicotyledons | entomophilous species | native | 84.1 | 15.9 | 0 | common | 40 |
| Euphorbia esula | L. | Euphorbiaceae | perennial dicotyledons | entomophilous species | native | 5.7 | 27.8 | 66.5 | common | 19 |
| Euphorbia helioscopia | L. | Euphorbiaceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 18.9 | 18.3 | 62.3 | common | 12 |
| Fagopyrum esculentum | Moench | Polygonaceae | annual dicotyledons | crops | casual archaeophytes | 0 | 0 | 0 | common | without data |
| Falcaria vulgaris | Bernh. | Apiaceae | perennial dicotyledons | entomophilous species | native | 75.4 | 11.7 | 12.9 | common | 27.5 |
| Fallopia convolvulus | (L.) Á. Löve | Polygonaceae | annual dicotyledons | annual weeds | native | 25.6 | 22.8 | 51.6 | common | 7.5 |
| Festuca arundinacea | Schreb. | Poaceae | perennial monocotyledons | species with an anti-erosion effect | native | 49.9 | 27.4 | 22.7 | common | 1 |
| Festuca pratensis | Huds. | Poaceae | perennial monocotyledons | species with an anti-erosion effect | native | 17.4 | 42.2 | 40.3 | common | 32 |
| Festuca rubra | L. | Poaceae | perennial monocotyledons | species with an anti-erosion effect | native | 16.3 | 49.6 | 34.1 | common | 30 |
| Fragaria vesca | L. | Rosaceae | perennial dicotyledons | entomophilous species | native | 27.2 | 49.2 | 23.6 | common | 35 |
| Fumaria officinalis | L. | Fumariaceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 19.3 | 0 | 80.7 | common | 19 |
| Galeopsis tetrahit | L. | Lamiaceae | annual dicotyledons | annual weeds | native | 33.1 | 0 | 66.9 | common | 9.5 |
| Galinsoga parviflora | Cav. | Asteraceae | annual dicotyledons | annual weeds | invasive neophytes | 15.5 | 32 | 52.5 | common | 2 |
| Galium album | Mill. | Rubiaceae | perennial dicotyledons | entomophilous species | native | 4.5 | 35.4 | 60.1 | common | 26 |
| Galium aparine | L. | Rubiaceae | annual dicotyledons | annual weeds | native | 12.2 | 0 | 87.8 | common | 28 |
| Galium verum | L. | Rubiaceae | perennial dicotyledons | entomophilous species | native | 1.3 | 71.8 | 26.9 | common | 34 |
| Geranium pusillum | Burm. f. | Geraniaceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 38.9 | 18.4 | 42.7 | common | 6 |
| Geranium pyrenaicum | Burm. f. | Geraniaceae | perennial dicotyledons | entomophilous species | naturalized neophytes | 48.8 | 16.2 | 35 | common | 11 |
| Geranium robertianum | L. | Geraniaceae | annual dicotyledons | entomophilous species | native | 32 | 11.3 | 56.7 | common | 39 |
| Geum urbanum | L. | Rosaceae | perennial dicotyledons | entomophilous species | native | 31.7 | 32.8 | 35.5 | common | 40 |
| Holosteum umbellatum | L. | Caryophyllaceae | annual dicotyledons | annual weeds | native | 1.8 | 0 | 98.2 | common | 75 |
| Hordeum murinum | L. | Poaceae | annual monocotyledons | species with an anti-erosion effect | naturalized archaeophytes | 19.8 | 18 | 62.2 | common | without data |
| Humulus lupulus | L. | Cannabaceae | perennial dicotyledons | deep-rooting species | native | 47.3 | 30.1 | 22.7 | common | 35 |
| Hypericum perforatum | L. | Hypericaceae | perennial dicotyledons | entomophilous species | native | 6.7 | 52.6 | 40.7 | common | 23 |
| Inula salicina | L. | Asteraceae | perennial dicotyledons | entomophilous species | native | 21.7 | 33.5 | 44.8 | C4a | 23 |
| Chelidonium majus | L. | Papaveraceae | annual dicotyledons | annual weeds | native | 54.7 | 6.6 | 38.8 | common | 29 |
| Chenopodium album | L. | Chenopodiaceae | annual dicotyledons | annual weeds | native | 30.5 | 34.1 | 35.4 | common | 3 |
| Chenopodium hybridum | L. | Chenopodiaceae | annual dicotyledons | annual weeds | native | 43.1 | 0 | 56.9 | common | 3 |
| Chenopodium polyspermum | L. | Chenopodiaceae | annual dicotyledons | annual weeds | native | 33.8 | 1.1 | 65 | common | 2 |
| Juglans regia | L. | Juglandaceae | perennial dicotyledons | deep-rooting species | invasive archaeophytes | 52.4 | 31.3 | 16.3 | common | 35 |
| Lactuca serriola | L. | Asteraceae | annual dicotyledons | annual weeds | invasive archaeophytes | 66.3 | 12.1 | 21.6 | common | 5 |
| Lamium album | L. | Lamiaceae | perennial dicotyledons | entomophilous species | naturalized archaeophytes | 39.8 | 2.4 | 57.8 | common | 21 |
| Lamium amplexicaule | L. | Lamiaceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 20.3 | 0 | 79.7 | common | 10.5 |
| Lamium maculatum | L. | Lamiaceae | perennial dicotyledons | entomophilous species | native | 32.1 | 16.1 | 51.8 | common | 15 |
| Lamium purpureum | L. | Lamiaceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 23.2 | 0 | 76.8 | common | 25 |
| Lathyrus sativus | L. | Fabaceae | annual dicotyledons | crops | casual archaeophytes | 0 | 0 | 0 | common | without data |
| Lathyrus tuberosus | L. | Fabaceae | perennial dicotyledons | deep-rooting species | naturalized archaeophytes | 21.9 | 19.9 | 58.3 | common | 18 |
| Lepidium draba | L. | Brassicaceae | perennial dicotyledons | deep-rooting species | naturalized archaeophytes | 47.6 | 0 | 52.4 | common | 11 |
| Ligustrum vulgare | L. | Oleaceae | perennial dicotyledons | deep-rooting species | native | 27.7 | 64 | 8.3 | common | 43 |
| Linaria vulgaris | Mill. | Scrophulariaceae | perennial dicotyledons | entomophilous species | naturalized archaeophytes | 10.7 | 25 | 64.3 | common | 10.5 |
| Linum usitatissimum | L. | Linaceae | annual dicotyledons | crops | casual archaeophytes | 2.1 | 0 | 97.9 | common | without data |
| Lolium multiflorum | Lam. | Poaceae | perennial monocotyledons | species with an anti-erosion effect | naturalized neophytes | 17.4 | 38.6 | 44 | common | 1 |
| Lolium perenne | L. | Poaceae | perennial monocotyledons | species with an anti-erosion effect | native | 18.5 | 29.3 | 52.3 | common | 8 |
| Lotus corniculatus | L. | Fabaceae | perennial dicotyledons | legumes | native | 11.5 | 19.1 | 69.4 | common | 15 |
| Luzula campestris | (L.) DC. | Juncaceae | perennial monocotyledons | species with an anti-erosion effect | native | 15.7 | 47.9 | 36.4 | common | 15 |
| Malva neglecta | Wallr. | Malvaceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 11 | 33.6 | 55.5 | common | 1 |
| Matricaria discoidea | DC. | Asteraceae | annual dicotyledons | annual weeds | naturalized neophytes | 26.6 | 0 | 73.4 | common | 3 |
| Medicago lupulina | L. | Fabaceae | perennial dicotyledons | legumes | native | 22.4 | 19.9 | 57.7 | common | 13 |
| Medicago sativa | L. | Fabaceae | perennial dicotyledons | legumes | naturalized neophytes | 31 | 25.9 | 43.1 | common | 10.5 |
| Melilotus albus | Medik. | Fabaceae | perennial dicotyledons | legumes | naturalized archaeophytes | 36.5 | 0 | 63.5 | common | 18 |
| Melilotus officinalis | (L.) Pall. | Fabaceae | perennial dicotyledons | legumes | naturalized archaeophytes | 23.5 | 40.1 | 36.4 | common | 23 |
| Mercurialis annua | L. | Euphorbiaceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 24.4 | 0 | 75.6 | common | without data |
| Microthlaspi perfoliatum | (L.) F. K. Mey. | Brassicaceae | annual dicotyledons | annual weeds | native | 0 | 0 | 0 | common | 45 |
| Myosotis arvensis | (L.) Hill | Boraginaceae | annual dicotyledons | annual weeds | native | 31.7 | 0 | 68.3 | common | 25 |
| Nonea pulla | (L.) DC. | Boraginaceae | perennial dicotyledons | entomophilous species | native | 48.4 | 0 | 51.9 | C4a | 30 |
| Onobrychis viciifolia | Scop. | Fabaceae | perennial dicotyledons | legumes | naturalized neophytes | 44.1 | 31.1 | 24.7 | common | 25 |
| Onopordum acanthium | L. | Asteraceae | perennial dicotyledons | entomophilous species | naturalized archaeophytes | 61.5 | 22.7 | 15.9 | common | 75 |
| Panicum miliaceum | L. | Poaceae | annual monocotyledons | species with an anti-erosion effect | casual neophytes | 0 | 0 | 0 | common | without data |
| Papaver rhoeas | L. | Papaveraceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 55 | 0 | 45 | common | 3 |
| Parthenocissus inserta | (A. Kern.) Fritsch | Vitaceae | perennial dicotyledons | deep-rooting species | invasive neophytes | 65.9 | 15.6 | 18.5 | common | 47 |
| Petrorhagia prolifera | (L.) P. W. Ball et Heywood | Caryophyllaceae | perennial dicotyledons | entomophilous species | native | 4.4 | 51.7 | 43.9 | C4a | 7 |
| Phacelia tanacetifolia | Benth. | Hydrophyllaceae | annual dicotyledons | crops | casual neophytes | 51.6 | 0 | 48.4 | common | without data |
| Physalis alkekengi | L. | Solanaceae | perennial dicotyledons | crops | naturalized archaeophytes | 0 | 0 | 0 | common | without data |
| Pilosella aurantiaca | (L.) F. W. Schultz et Sch. Bip. | Asteraceae | perennial dicotyledons | entomophilous species | native | 42.4 | 0 | 57.6 | C3 | 20 |
| Pilosella officinarum | Vaill. | Asteraceae | perennial dicotyledons | entomophilous species | native | 9.6 | 26.3 | 64.1 | 27 | |
| Pisum sativum | L. | Fabaceae | annual dicotyledons | crops | casual archaeophytes | 0 | 0 | 0 | common | without data |
| Plantago lanceolata | L. | Plantaginaceae | perennial dicotyledons | entomophilous species | native | 61.1 | 0.9 | 38 | common | 16 |
| Plantago major | L. | Plantaginaceae | perennial dicotyledons | entomophilous species | native | 82.3 | 0 | 17.7 | common | 6 |
| Plantago media | L. | Plantaginaceae | perennial dicotyledons | entomophilous species | native | 68.2 | 0 | 31.8 | common | 30 |
| Poa annua | L. | Poaceae | annual monocotyledons | species with an anti-erosion effect | native | 6.8 | 35 | 58.2 | common | 4 |
| Poa pratensis | L. | Poaceae | perennial monocotyledons | species with an anti-erosion effect | native | 12.4 | 0 | 87.6 | common | 30 |
| Polygonum aviculare | L. | Polygonaceae | annual dicotyledons | annual weeds | native | 15.3 | 29.7 | 55 | common | 1 |
| Portulaca oleracea | L. | Portulacaceae | annual dicotyledons | annual weeds | invasive archaeophytes | 0.3 | 0 | 99.7 | common | without data |
| Potentilla argentea | L. | Rosaceae | perennial dicotyledons | entomophilous species | native | 21.3 | 50.4 | 28.3 | common | 19 |
| Potentilla reptans | L. | Rosaceae | perennial dicotyledons | entomophilous species | native | 35.3 | 29.3 | 35.4 | common | 22 |
| Quercus petraea | (Matt.) Liebl. | Fagaceae | perennial dicotyledons | deep-rooting species | native | 28.1 | 63.2 | 8.6 | common | 35 |
| Raphanus sativus | L. | Brassicaceae | annual dicotyledons | crops | casual archaeophytes | 62.1 | 0 | 37.9 | common | without data |
| Raphanus raphanistrum | L. | Brassicaceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 63.2 | 0 | 36.8 | common | 4.5 |
| Reseda lutea | L. | Resedaceae | perennial dicotyledons | entomophilous species | naturalized archaeophytes | 51 | 0 | 49.1 | common | 13 |
| Ribes aureum | Pursh | Grossulariaceae | perennial dicotyledons | deep-rooting species | casual neophytes | 31 | 58.3 | 10.8 | common | without data |
| Robinia pseudoacacia | L. | Fabaceae | perennial dicotyledons | deep-rooting species | invasive neophytes | 28.8 | 41.4 | 29.8 | common | 30 |
| Rosa canina | L. | Rosaceae | perennial dicotyledons | deep-rooting species | native | 16.6 | 50.2 | 33.2 | common | 35 |
| Rubus sect. Rubus | Kaplan et al. | Rosaceae | perennial dicotyledons | deep-rooting species | native | 46 | 39.1 | 14.9 | common | without data |
| Rumex crispus | L. | Polygonaceae | perennial dicotyledons | deep-rooting species | native | 65.1 | 0 | 35 | common | 7 |
| Rumex obtusifolius | L. | Polygonaceae | perennial dicotyledons | deep-rooting species | native | 80.6 | 0 | 19.5 | common | 7.5 |
| Salvia pratensis | L. | Lamiaceae | perennial dicotyledons | entomophilous species | native | 62.8 | 2.4 | 34.8 | common | 35 |
| Sambucus nigra | L. | Sambucaceae | perennial dicotyledons | deep-rooting species | native | 36.2 | 23.4 | 40.4 | common | 30 |
| Scabiosa ochroleuca | L. | Dipsacaceae | perennial dicotyledons | entomophilous species | native | 25.6 | 61.6 | 12.8 | common | 30 |
| Scleranthus annuus | L. | Caryophyllaceae | annual dicotyledons | annual weeds | native | 0 | 0.7 | 99.3 | common | 2 |
| Secale cereale | L. | Poaceae | annual monocotyledons | crops | casual archaeophytes | 0 | 0 | 0 | common | without data |
| Securigera varia | (L.) Lassen | Fabaceae | perennial dicotyledons | legumes | native | 39.6 | 1.6 | 58.9 | common | 25 |
| Senecio jacobaea | L. | Asteraceae | annual dicotyledons | entomophilous species | native | 65.6 | 0 | 34.4 | common | 35 |
| Senecio vulgaris | L. | Asteraceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 26.4 | 0 | 73.6 | common | 4 |
| Setaria pumila | (Poir.) Roem. et Schult. | Poaceae | annual monocotyledons | species with an anti-erosion effect | invasive archaeophytes | 38.4 | 24.7 | 37 | common | 4 |
| Setaria viridis | (L.) P. Beauv. | Poaceae | annual monocotyledons | species with an anti-erosion effect | invasive archaeophytes | 23.6 | 41.5 | 34.9 | common | 3 |
| Setaria verticillata | (L.) P. Beauv. | Poaceae | annual monocotyledons | species with an anti-erosion effect | invasive archaeophytes | 25.2 | 33.2 | 41.6 | common | 3 |
| Silene latifolia subsp. alba | (Mill.) Greuter et Burdet | Caryophyllaceae | perennial dicotyledons | entomophilous species | naturalized archaeophytes | 51.2 | 0 | 48.8 | common | 10 |
| Silene noctiflora | L. | Caryophyllaceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 49.3 | 4.8 | 45.9 | C4a | 3.5 |
| Silene vulgaris | (Moench) Garcke | Caryophyllaceae | perennial dicotyledons | entomophilous species | native | 48.1 | 0.2 | 51.7 | common | 26.5 |
| Sinapis alba | L. | Brassicaceae | annual dicotyledons | crops | casual archaeophytes | 56 | 0 | 44 | common | without data |
| Solanum nigrum | L. | Solanaceae | annual dicotyledons | annual weeds | native | 0 | 0 | 100 | common | 4 |
| Sonchus oleraceus | L. | Asteraceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 61.1 | 0 | 38.9 | common | 4 |
| Stellaria media | (L.) Vill. | Caryophyllaceae | annual dicotyledons | annual weeds | native | 5 | 0 | 95 | common | 11.5 |
| Stipa pennata | L. | Poaceae | perennial monocotyledons | species with an anti-erosion effect | native | 16.5 | 83.5 | 0 | C3 | 75 |
| Symphytum officinale | L. | Boraginaceae | perennial dicotyledons | entomophilous species | native | 68.1 | 0 | 31.9 | common | 32 |
| Tanacetum vulgare | L. | Asteraceae | perennial dicotyledons | entomophilous species | naturalized archaeophytes | 59.1 | 18.3 | 22.6 | common | 18 |
| Taraxacum sect. Taraxacum | Kirschner et al. | Asteraceae | perennial dicotyledons | entomophilous species | native | 55.4 | 0 | 44.7 | common | without data |
| Tragopogon dubius | Scop. | Asteraceae | perennial dicotyledons | entomophilous species | naturalized archaeophytes | 17.4 | 22.4 | 60.2 | common | 13 |
| Tragopogon orientalis | L. | Asteraceae | perennial dicotyledons | entomophilous species | native | 42 | 0 | 58 | common | 16 |
| Trifolium alexandrinum | L. | Fabaceae | annual dicotyledons | legumes | casual neophytes | 0 | 0 | 0 | common | without data |
| Trifolium campestre | Schreb. | Fabaceae | perennial dicotyledons | legumes | native | 9.2 | 41.7 | 49.1 | common | 14.5 |
| Trifolium incarnatum | L. | Fabaceae | annual dicotyledons | legumes | casual neophytes | 40.4 | 11.8 | 47.8 | common | without data |
| Trifolium pratense | L. | Fabaceae | perennial dicotyledons | legumes | native | 24.7 | 31.5 | 43.8 | common | 13 |
| Trifolium repens | L. | Fabaceae | perennial dicotyledons | legumes | native | 25.4 | 12.7 | 61.9 | common | 7 |
| Tripleurospermum inodorum | (L.) Sch. Bip. | Asteraceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 23.5 | 0 | 76.5 | common | 5 |
| Triticum aestivum | L. | Poaceae | annual monocotyledons | crops | casual archaeophytes | 39.3 | 22.8 | 37.9 | common | without data |
| Urtica dioica | L. | Urticaceae | perennial dicotyledons | deep-rooting species | native | 44.8 | 18.2 | 37.1 | common | 25 |
| Urtica urens | L. | Urticaceae | annual dicotyledons | annual weeds | naturalized archaeophytes | 22.1 | 32.1 | 45.8 | C3 | 25 |
| Veronica hederifolia | L. | Scrophulariaceae | annual dicotyledons | annual weeds | native | 19.3 | 0 | 80.7 | common | 4.8 |
| Vicia cracca | L. | Fabaceae | perennial dicotyledons | legumes | native | 27.3 | 28.7 | 44 | common | 30 |
| Vicia pannonica | Crantz | Fabaceae | annual dicotyledons | legumes | naturalized archaeophytes | 0 | 0 | 0 | C2t | without data |
| Viola arvensis | Murray | Violaceae | annual dicotyledons | annual weeds | native | 30.8 | 1.6 | 67.6 | common | 6 |
References
- Kleijn, D.; Rundlöf, M.; Scheper, J.; Smith, H.G.; Tscharntke, T. Does conservation on farmland contribute to halting the biodiversity decline? Trends Ecol. Evol. 2011, 26, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Pe’er, G.; Zinngrebe, Y.; Hauck, J.; Schindler, S.; Dittrich, A.; Zingg, S.; Tscharntke, T.; Oppermann, R.; Sutcliffe, L.M.; Sirami, C.; et al. Adding some green to the greening: Improving the EU’s Ecological Focus Areas for biodiversity and farmers. Conserv. Lett. 2017, 10, 517–530. [Google Scholar] [CrossRef]
- Batáry, P.; Dicks, L.V.; Kleijn, D.; Sutherland, W.J. The role of agri-environment schemes in conservation and environmental management. Conserv. Biol. 2015, 29, 1006–1016. [Google Scholar] [CrossRef] [PubMed]
- Paiola, A.; Assandri, G.; Brambilla, M.; Zottini, M.; Pedrini, P.; Nascimbene, J. Exploring the potential of vineyards for biodiversity conservation and delivery of biodiversity-mediated ecosystem services: A global-scale systematic review. Sci. Total Environ. 2020, 706, 135839. [Google Scholar] [CrossRef] [PubMed]
- Henle, K.; Alard, D.; Clitherow, J.; Cobb, P.; Firbank, L.; Kull, T.; McCracken, D.; Moritz, R.F.; Niemelä, J.; Rebane, M.; et al. Identifying and managing the conflicts between agriculture and biodiversity conservation in Europe—A review. Agric. Ecosyst. Environ. 2008, 124, 60–71. [Google Scholar] [CrossRef]
- Sutcliffe, L.M.; Batáry, P.; Kormann, U.; Báldi, A.; Dicks, L.V.; Herzon, I.; Kleijn, D.; Tryjanowski, P.; Apostolova, I.; Arlettaz, R.; et al. Harnessing the biodiversity value of Central and Eastern European farmland. Divers. Distrib. 2015, 21, 722–730. [Google Scholar] [CrossRef]
- Loos, J.; Von Wehrden, H. Beyond biodiversity conservation: Land sharing constitutes sustainable agriculture in European cultural landscapes. Sustainability 2018, 10, 1395. [Google Scholar] [CrossRef]
- Brugisser, O.T.; Schmidt-Entling, M.H.; Bacher, S. Effects of vineyards management on biodiversity at three trophic levels. Biol. Conserv. 2010, 143, 1521–1528. [Google Scholar] [CrossRef]
- Nascimbene, J.; Marini, L.; Paoletti, M.G. Organic farming benefits local plant diversity in vineyard farms located in intensive agricultural landscapes. Environ. Manag. 2012, 49, 1054–1060. [Google Scholar] [CrossRef]
- Rusch, A.; Beaumelle, L.; Giffard, B.; Ugaglia, A.A. Chapter Seven—Harnessing biodiversity and ecosystem services to safeguard multifunctional vineyard landscapes in a global change context. In Advances in Ecological Research; Bohan, D.A., Dumbrell, A.J., Vanbergen, A.J., Eds.; Academic Press: Cambridge, MA, USA, 2021; Volume 65, pp. 305–335. [Google Scholar] [CrossRef]
- Tscharntke, T.; Klein, A.M.; Kruess, A.; Steffan-Dewenter, I.; Thies, C. Landscape perspectives on agricultural intensification and biodiversity-ecosystem service management. Ecol. Lett. 2005, 8, 857–874. [Google Scholar] [CrossRef]
- Hall, R.M.; Penke, N.; Kriechbaum, M.; Kratschmer, S.; Jung, V.; Chollet, S.; Guernion, M.; Nicolai, A.; Burel, F.; Fertil, A.; et al. Vegetation management intensity and landscape diversity alter plant species richness, functional traits and community composition across European vineyards. Agric. Syst. 2020, 177, 102706. [Google Scholar] [CrossRef]
- Cabrera-Pérez, C.; Valencia-Gredilla, F.; Royo-Esnal, A.; Recasens, J. Organic mulches as an alternative to conventional under-vine weed management in Mediterranean irrigated vineyards. Plants 2022, 11, 2785. [Google Scholar] [CrossRef] [PubMed]
- Maixner, M.; Ahrens, U.; Seemüller, E. Detection of the German grapevine yellows (Vergilbungskrankheit) MLO in grapevine, alternative hosts and a vector by a specific PCR procedure. Eur. J. Plant Pathol. 1995, 101, 241–250. [Google Scholar] [CrossRef]
- Castillo, P.; Rapoport, H.F.; Rius, J.P.; Díaz, R.J. Suitability of weed species prevailing in Spanish vineyards as hosts for root-knot nematodes. Eur. J. Plant Pathol. 2008, 120, 43–51. [Google Scholar] [CrossRef]
- Gualandri, V.; Asquini, E.; Bianchedi, P.; Covelli, L.; Brilli, M.; Malossini, U.; Bragagna, P.; Saldarelli, P.; Si-Ammour, A. Identification of herbaceous hosts of the Grapevine Pinot gris virus (GPGV). Eur. J. Plant Pathol. 2017, 147, 21–25. [Google Scholar] [CrossRef]
- Demian, E.; Jaksa-Czotter, N.; Varallyay, E. Grapevine Pinot gris virus is present in different non-Vitis hosts. Plants 2022, 11, 1830. [Google Scholar] [CrossRef] [PubMed]
- Prosdocimi, M.; Cerdà, A.; Tarolli, P. Soil water erosion on Mediterranean vineyards: A review. Catena 2016, 141, 1–21. [Google Scholar] [CrossRef]
- Heap, I.; Duke, S.O. Overview of glyphosate-resistant weeds worldwide. Pest Manag. Sci. 2018, 74, 1040–1049. [Google Scholar] [CrossRef] [PubMed]
- Biddoccu, M.; Ferraris, S.; Opsi, F.; Cavallo, E. Long-term monitoring of soil management effects on runoff and soil erosion in sloping vineyards in Alto Monferrato (North-West Italy). Soil Res. 2016, 155, 176–189. [Google Scholar] [CrossRef]
- Garcia, L.; Celette, F.; Gary, C.; Ripoche, A.; Valdes-Gomez, H.; Metay, A. Management of service crops for the provision of ecosystem services in vineyards: A review. Agric. Ecosyst. Environ. 2018, 251, 158–170. [Google Scholar] [CrossRef]
- Donnini, S.; Tessarin, P.; Ribera-Fonseca, A.; Di Foggia, M.; Parpinello, G.P.; Rombola, A.D. Glyphosate impacts on polyphenolic composition in grapevine (Vitis vinifera L.) berries and wine. Food Chem. 2016, 213, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Zaller, J.G.; Cantelmo, C.; Dos Santos, G.; Muther, S.; Gruber, E.; Pallua, P.; Mandl, K.; Friedrich, B.; Hofstetter, I.; Schmuckenschlager, B.; et al. Herbicides in vineyards reduce grapevine root mycorrhization and alter soil microorganisms and the nutrient composition in grapevine roots, leaves, xylem sap and grape juice. Environ. Sci. Pollut. R. 2018, 25, 23215–23226. [Google Scholar] [CrossRef] [PubMed]
- Guerra, B.; Steenwerth, K. Influence of Floor Management Technique on Grapevine Growth, Disease Pressure, and Juice and Wine Composition: A Review. Am. J. Enol. Viticult. 2012, 63, 149–164. [Google Scholar] [CrossRef]
- Steenwerth, K.; Belina, K.M. Cover crops and cultivation: Impacts on soil N dynamics and microbiological function in a Mediterranean vineyard agroecosystem. Appl. Soil Ecol. 2008, 40, 370–380. [Google Scholar] [CrossRef]
- Coniberti, A.; Ferrari, V.; Disegna, E.; Petillo, M.G.; Lakso, A.N. Under-trellis cover crop and planting density to achieve vine balance in a humid climate. Sci. Hortic. 2018, 227, 65–74. [Google Scholar] [CrossRef]
- Ragasová, L.; Kopta, T.; Winkler, J.; Pokluda, R. The Current Stage of Greening Vegetation in Selected Wine-Regions of South Moravian Region (Czech Republic). Agronomy 2019, 9, 541. [Google Scholar] [CrossRef]
- Schmid, A.; Weibel, F. Das sandwich system—ein Verfahren zur herbizidfreien Baumstreifenbewirtschaftung? [The sandwich system, a procedure for herbicide free in-row weed control?]. Obstbau 2000, 25, 214–217. [Google Scholar]
- Mia, M.J.; Furmanczyk, E.M.; Golian, J.; Kwiatkowska, J.; Malusá, E.; Neri, D. Living Mulch with Selected Herbs for Soil Management in Organic Apple Orchards. Horticulturae 2021, 7, 59. [Google Scholar] [CrossRef]
- Mia, M.J.; Massetani, F.; Murri, G.; Neri, D. Sustainable alternatives to chemicals for weed control in the orchard—A review. Hortic. Sci. 2020, 47, 1–12. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015, 1, e1500052. [Google Scholar] [CrossRef]
- Körner, K.; Jeltsch, F. Detecting general plant functional type responses in fragmented landscapes using spatially-explicit simulations. Ecol. Model. 2008, 210, 87–300. [Google Scholar] [CrossRef]
- Murphy, H.T.; Lovett-Doust, J. Context and connectivity in plant metapopulations and landscape mosaics: Does the matrix matter? Oikos 2004, 105, 3–14. [Google Scholar] [CrossRef]
- Ries, L.; Sisk, T.D. A Predictive model of edge effects. Ecology 2004, 85, 2917–2926. [Google Scholar] [CrossRef]
- Fernández-Lázaro, D.; Mielgo-Ayuso, J.; Seco Calvo, J.; Córdova Martínez, A.; Caballero García, A.; Fernandez-Lazaro, C.I. Modulation of Exercise-Induced Muscle Damage, Inflammation, and Oxidative Markers by Curcumin Supplementation in a Physically Active Population: A Systematic Review. Nutrients 2020, 15, 501. [Google Scholar] [CrossRef] [PubMed]
- Legrand, A.; Cornez, L.; Samkari, W.; Mazzella, J.M.; Venisse, A.; Boccio, V.; Auribault, K.; Keren, B.; Benistan, K.; Germain, D.P.; et al. Mutation spectrum in the ABCC6 gene and genotype-phenotype correlations in a French cohort with pseudoxanthoma elasticum. Genet. Med. 2017, 19, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Dener, E.; Ovadia, O.; Shemesh, H.; Altman, A.; Chen, S.C.; Giladi, I. Direct and indirect effects of fragmentation on seed dispersal traits in a fragmented agricultural landscape. Agric. Ecosyst. Environ. 2021, 309, 107273. [Google Scholar] [CrossRef]
- Fahrig, L. Ecological Responses to Habitat Fragmentation Per Se. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 1–23. [Google Scholar] [CrossRef]
- Ibanez, I.; Katz, D.S.W.; Peltier, D.; Wolf, S.M.; Barrie, B.T.C. Assessing the integrated effects of landscape fragmentation on plants and plant communities: The challenge of multiprocess-multiresponse dynamics. J. Ecol. 2014, 102, 882–895. [Google Scholar] [CrossRef]
- Sanguankeo, P.P.; León, R.G. Weed management practices determine plant and arthropod diversity and seed predation in vineyards. Weed Res. 2011, 51, 404–412. [Google Scholar] [CrossRef]
- Arlettaz, R.; Maurer, M.L.; Mosimann-Kampe, P.; Nusslé, S.; Abadi, F.; Braunisch, V.; Schaub, M. New vineyard cultivation practices create patchy ground vegetation, favouring Woodlarks. J. Ornithol. 2012, 153, 229–238. [Google Scholar] [CrossRef]
- Fernandez-Mena, H.; Frey, H.; Celette, F.; Garcia, L.; Barkaoui, K.; Hossard, L.; Naulleau, A.; Metral, R.; Gary, C.; Metay, A. Spatial and temporal diversity of service plant management strategies across vineyards in the south of France. Analysis through the Coverage Index. Eur. J. Agron. 2021, 123, 126191. [Google Scholar] [CrossRef]
- Geldenhuys, M.; Gaigher, R.; Pryke, J.S.; Samways, M.J. Diverse herbaceous cover crops promote vineyard arthropod diversity across different management regimes. Agric. Ecosyst. Environ. 2021, 307, 107222. [Google Scholar] [CrossRef]
- Candiago, S.; Winkler, K.J.; Giombini, V.; Giupponi, C.; Egarter Vigl, L. An ecosystem service approach to the study of vineyard landscapes in the context of climate change: A review. Sustain. Sci. 2023, 18, 997–1013. [Google Scholar] [CrossRef]
- Winkler, K.J.; Viers, J.H.; Nicholas, K.A. Assessing ecosystem services and multifunctionality for vineyard systems. Front. Environ. Sci. 2017, 5, 15. [Google Scholar] [CrossRef]
- Winter, S.; Bauer, T.; Strauss, P.; Kratschmer, S.; Paredes, D.; Popescu, D.; Landa, B.; Guzmán, G.; Gómez, J.A.; Guernion, M.; et al. Effects of vegetation management intensity on biodiversity and ecosystem services in vineyards: A meta-analysis. J. Appl. Ecol. 2018, 55, 2484–2495. [Google Scholar] [CrossRef] [PubMed]
- Quevedo, L.A.; Silva, R.A.; Godoy, R.; Jansen, K.; Matos, M.B.; Tavares Pinheiro, K.A.; Pinheiro, R.T. The impact of maternal post-partum depression on the language development of children at 12 months. Child Care Health Dev. 2012, 38, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Diaz, A.; Shin, D.S.; Moreno, B.H.; Saco, J.; Escuin-Ordinas, H.; Rodriguez, G.A.; Zaretsky, J.M.; Sun, L.; Hugo, W.; Wang, X.; et al. Interferon Receptor Signaling Pathways Regulating PD-L1 and PD-L2 Expression. Cell Rep. 2017, 19, 1189–1201. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Colmenero, M.; Bienes, R.; Eldridge, D.J.; Marques, M.J. Vegetation cover reduces erosion and enhances soil organic carbon in a vineyard in the central Spain. Catena 2013, 104, 153–160. [Google Scholar] [CrossRef]
- Sáenz-Romo, M.G.; Veas-Bernal, A.; Martínez-García, H.; Campos-Herrera, R.; Ibáñez-Pascual, S.; Martínez-Villar, E.; Pérez-Moreno, I.; Marco-Mancebón, V.S. Ground cover management in a Mediterranean vineyard: Impact on insect abundance and diversity. Agric. Ecosyst. Environ. 2019, 283, 106571. [Google Scholar] [CrossRef]
- Altieri, M.A.; Ponti, L.; Nicholls, C.I. Manipulating vineyard biodiversity for improved insect pest management: Case studies from northern California. Int. J. Biodivers. Sci. Manag. 2005, 1, 191–203. [Google Scholar] [CrossRef]
- Wilson, C.; Lukowicz, R.; Merchant, S.; Valquier-Flynn, H.; Caballero, J.; Sandoval, J.; Okuom, M.; Huber, C.; Brooks, T.D.; Wilson, E.; et al. Quantitative and Qualitative Assessment Methods for Biofilm Growth: A Mini-review. Res. Rev. J. Eng. Technol. 2017, 6. Available online: http://www.rroij.com/open-access/quantitative-and-qualitative-assessment-methods-for-biofilm-growth-a-minireview-.pdf (accessed on 20 December 2023).
- Sáenz-Romo, M.G.; Veas-Bernal, A.; Martínez-García, H.; Ibáñez-Pascual, S.; Martínez-Villar, E.; Campos-Herrera, R.; Marco-Mancebón, V.S.; Pérez-Moreno, I. Effects of Ground Cover Management on Insect Predators and Pests in a Mediterranean Vineyard. Insects 2019, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Zanettin, G.; Bullo, A.; Pozzebon, A.; Burgio, G.; Duso, C. Influence of Vineyard inter-row groundcover vegetation management on arthropod assemblages in the Vineyards of North-Eastern Italy. Insects 2021, 12, 349. [Google Scholar] [CrossRef]
- CGS. Geological Map of the Czech Republic, 1:50,000; Czech Geological Society: Prague, Czech Republic, 2018; Available online: https://mapy.geology.cz/geocr50/ (accessed on 8 November 2022).
- CGS. Map of Soil Types of the Czech Republic, 1:50,000; Czech Geological Society: Prague, Czech Republic, 2017; Available online: https://mapy.geology.cz/pudy/ (accessed on 8 November 2022).
- Culek, M. (Ed.) Biogeographical Division of the Czech Republic (Biogeografické Členění České Republiky), 1st ed.; Enigma: Prague, Czech Republic, 1996; p. 347. (In Czech) [Google Scholar]
- IUSS Working Group WRB. World Reference Base for Soil Resources. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, 4th ed.; International Union of Soil Sciences (IUSS): Vienna, Austria, 2022; 236p, Available online: https://www.isric.org/sites/default/files/WRB_fourth_edition_2022-12-18.pdf (accessed on 20 December 2023).
- Biondi, E. Phytosociology today: Methodological and conceptual evolution. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2011, 145, 19–29. [Google Scholar] [CrossRef]
- Kaplan, Z.; Danihelka, J.; Chrtek, J.; Kirschner, J.; Kubát, K.; Štech, M.; Štěpánek, J. (Eds.) Key to the Flora of the Czech Republic [Klíč ke květeně České Republiky], 2nd ed.; Academia: Prague, Czech Republic, 2019; p. 1168. (In Czech) [Google Scholar]
- Chytrý, M.; Danihelka, J.; Kaplan, Z.; Wild, J.; Holubová, D.; Novotný, P.; Řezníčková, M.; Rohn, M.; Dřevojan, P.; Grulich, V.; et al. Pladias Database of the Czech Flora and Vegetation. Preslia 2021, 93, 1–87. [Google Scholar] [CrossRef]
- Dřevojan, P.; Čeplová, N.; Štěpánková, P.; Axmanová, I. Life Form. 2022. Available online: www.floraveg.eu (accessed on 20 December 2023).
- Tyler, T.; Herbertsson, L.; Olofsson, J.; Olsson, P.A. Ecological indicator and traits values for swedish vascular plants. Ecol. Indic. 2021, 120, 106923. [Google Scholar] [CrossRef]
- Pyšek, P.; Sádlo, J.; Chrtek, J., Jr.; Chytrý, M.; Kaplan, Z.; Pergl, J.; Pokorná, A.; Axmanová, I.; Čuda, J.; Doležal, J.; et al. Catalogue of alien plants of the Czech Republic (3rd edition): Species richness, status, distributions, habitats, regional invasion levels, introduction pathways and impacts. Preslia 2022, 94, 447–577. [Google Scholar] [CrossRef]
- Grime, J.P. Vegetation classification by reference to strategies. Nature 1974, 250, 26–31. [Google Scholar] [CrossRef]
- Grime, J.P. Plant Strategies and Vegetation Processes; Wiley: Chichester, UK, 1979. [Google Scholar]
- Guo, W.Y.; Pierce, S. Life Strategy [Životní Strategie]. 2019. Available online: www.pladias.cz (accessed on 20 December 2023). (In Czech).
- Grulich, V. The Red List of vascular plants of the Czech Republic [Červený seznam cévnatých rostlin ČR]. Příroda 2017, 35, 75–132. (In Czech) [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 20 December 2023).
- El Titi, A. Implications of Soil tillage for Weed Communities. In Soil Tillage in Agroecosystems; El Titi, A., Ed.; CRC Press: Boca Raton, FL, USA, 2003; pp. 147–185. [Google Scholar] [CrossRef]
- Kudsk, P.; Streibig, J.C. Herbicides—A two-edged sword. Weed Res. 2003, 43, 90–102. [Google Scholar] [CrossRef]
- Freemark, K.; Boutin, C. Impacts of agricultural herbicide use on terrestrial wildlife intemperate landscapes: A review withspecial reference to NorthAmerica. Agric. Ecosyst. Environ. 1995, 52, 67–91. [Google Scholar] [CrossRef]
- Wardle, D.A.; Yeates, G.W.; Watson, R.N.; Nicholson, K.S. The detritus foodweb and the diversity of soil fauna as indicators of disturbance regimes in agroecosystems. Plant Soil 1995, 179, 35–43. [Google Scholar] [CrossRef]
- El Jaouhari, M.; Damour, G.; Tixier, P.; Coulis, M. Glyphosate reduces the biodiversity of soil macrofauna and benefits exotic over native species in a tropical agroecosystem. Basic Appl. Ecol. 2023, 73, 18–26. [Google Scholar] [CrossRef]
- Winkler, J.; Ricica, T.; Hubacíková, V.; Koda, E.; Vaverková, M.D.; Havel, L.; Zółtowski, M. Water Protection Zones—Impacts on Weed Vegetation of Arable Soil. Water 2023, 15, 3161. [Google Scholar] [CrossRef]
- Chan, K.Y.; Munro, K. Evaluating mustard extracts for earthworm sampling. Pedobiology 2001, 45, 272–278. [Google Scholar] [CrossRef]
- Emmerling, C. Response of earthworm communities to different types of soil tillage. Appl. Soil Ecol. 2001, 17, 91–96. [Google Scholar] [CrossRef]
- Pommeresche, R.; Loes, A.K. Relations between agronomic practice and earthworms in Norvegian arable soils. (Global Science Books). Dyn. Soil Dyn. Plant 2009, 3, 129–142. [Google Scholar]
- Winkler, J.; Mazur, Ł.; Smékalová, M.; Podlasek, A.; Hurajová, E.; Koda, E.; Jiroušek, M.; Jakimiuk, A.; Vaverková, M.D. Influence of land use on plant community composition in Vysocina Region grasslands, Czech Republic. Environ. Prot. Eng. 2022, 48, 21–33. [Google Scholar] [CrossRef]
- Kazakou, E.; Fried, G.; Richarte, J.; Gimenez, O.; Violle, C.; Metay, A. A plant trait-based response-and-effect framework to assess vineyard inter-row soil management. Bot. Lett. 2016, 163, 373–388. [Google Scholar] [CrossRef]
- Storkey, J.; Meyer, S.; Still, K.S.; Leuschner, C. The impact of agricultural intensification and land-use change on the European arable flora. Proc. R. Soc. B Biol. Sci. 2012, 279, 1421–1429. [Google Scholar] [CrossRef]
- Marshall, E.J.P.; Brown, V.K.; Boatman, N.D.; Lutman, P.J.W.; Squire, G.R.; Ward, L.K. The role of weeds in supporting biological diversity within crop fields. Weed Res. 2003, 43, 77–89. [Google Scholar] [CrossRef]
- Ragasová, L.; Kopta, T.; Winkler, J.; Šefrová, H.; Pokluda, R. The Effect of the Proportion of Adjacent Non-Crop Vegetation on Plant and Invertebrate Diversity in the Vineyards of the South Moravian Region. Agronomy 2021, 11, 1073. [Google Scholar] [CrossRef]
- Hurajová, E.; Martínez Barroco, P.; Havel, L.; Děkanovský, I.; Winkler, J. Relationship Between Vegetation Succession and Earthworm Population in Vineyards. J. Ecol. Eng. 2024, 25, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Winkler, J.; Ježová, M.; Punčochář, R.; Hurajová, E.; Martínez Barroso, P.; Kopta, T.; Semerádová, D.; Vaverková, M.D. Fire Hazard: Undesirable Ecosystem Function of Orchard Vegetation. Fire 2023, 6, 25. [Google Scholar] [CrossRef]
- Tilman, D.; Isbell, F.; Cowles, J.M. Biodiversity and ecosystem functioning. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 471–493. [Google Scholar] [CrossRef]
- Guerra, J.G.; Cabello, F.; Fernández-Quintanilla, C.; Peña, J.M.; Dorado, J. How weed management influence plant community composition, taxonomic diversity and crop yield: A long-term study in a Mediterranean vineyard. Agric. Ecosyst. Environ. 2022, 326, 107816. [Google Scholar] [CrossRef]
- Winkler, J.; Koda, E.; Červenková, J.; Děkanovský, I.; Nowysz, A.; Mazur, Ł.; Jakimiuk, A.; Vaverková, M.D. Green space in an extremely exposed part of the city center “Aorta of Warsaw”-Case study of the urban lawn. Urban Ecosyst. 2023, 26, 1225–1238. [Google Scholar] [CrossRef]
- Winkler, J.; Vaverková, M.D.; Havel, L. Anthropogenic life strategy of plants. Anthr. Rev. 2023, 10, 455–462. [Google Scholar] [CrossRef]
- Poni, S.; Gatti, M.; Palliotti, A.; Dai, Z.W.; Duchene, E.; Truong, T.T.; Ferrara, G.; Matarrese, A.M.S.; Gallotta, A.; Bellincontro, A.; et al. Grapevine quality: A multiple-choice issue. Sci. Hortic. 2018, 234, 445–462. [Google Scholar] [CrossRef]
- van Leeuwen, C.; Roby, J.P.; de Resseguier, L. Soil-related terroir factors: A review. OENO One 2018, 52, 173–188. [Google Scholar] [CrossRef]
- Griesser, M.; Steiner, M.; Pingel, M.; Uzman, D.; Preda, C.; Giffard, B.; Tolle, P.; Memedemin, D.; Forneck, A.; Reineke, A.; et al. General trends of different inter-row vegetation management affecting vine vigor and grape quality across european 445 vineyards. Agric. Ecosyst. Environ. 2022, 338, 108073. [Google Scholar] [CrossRef]
- Boinot, S.; Fried, G.; Storkey, J.; Metcalfe, H.; Barkaoui, K.; Lauri, P.E.; Mézière, D. Alley cropping agroforestry systems: Reservoirs for weeds or refugia for plant diversity? Agric. Ecosyst. Environ. 2019, 284, 106584. [Google Scholar] [CrossRef]
- Mahaut, L.; Cheptou, P.O.; Fried, G.; Munoz, F.; Storkey, J.; Vasseur, F.; Violle, C.; Bretagnolle, F. Weeds: Against the Rules? Trends Plant Sci. 2020, 25, 1107–1116. [Google Scholar] [CrossRef]




| Municipality | Vine Line | Year | GPS | Soil Type | Area (ha) |
|---|---|---|---|---|---|
| Horní Dunajovice | Frédy | 1995 | 48°56′36.942″ N, 16°10′37.543″ E | chernozem | 2.35 |
| 2000 | 48°56′31.549″ N, 16°11′6.951″ E | cambisol | 7.32 | ||
| 2002 | 48°56′50.739″ N, 16°10′42.269″ E | chernozem | 15.85 | ||
| 2009 | 48°56′37.083″ N, 16°10′55.425″ E | chernozem, cambisol | 4.38 | ||
| 2016 | 48°56′45.850″ N, 16°10′39.508″ E | chernozem | 1.9 | ||
| 2018 | 48°56′41.433″ N, 16°10′38.154″ E | chernozem | 2.25 | ||
| 2021 | 48°56′38.423″ N, 16°10′38.040″ E | chernozem | 3.03 | ||
| Horní Dunajovice | Stará hora | 1995 | 48°57′7.324″ N, 16°10′27.686″ E | chernozem, cambisol | 4.13 |
| 2000 | 48°57′1.878″ N, 16°10′51.851″ E | chernozem, cambisol | 14.35 | ||
| 2020 | 48°57′5.815″ N, 16°10′34.300″ E | chernozem | 9.07 | ||
| 2021 | 48°57′3.176″ N, 16°10′46.596″ E | chernozem | 3.97 | ||
| Hostěradice | Volné pole | 1972 | 48°56′47.298″ N, 16°17′18.419″ E | chernozem, cambisol | 10.23 |
| 2002 | 48°56′59.237″ N, 16°17′12.561″ E | chernozem | 17.09 | ||
| 2003 | 48°56′55.483″ N, 16°17′20.826″ E | chernozem | 8.54 | ||
| 2014 | 48°57′25.162″ N, 16°17′17.557″ E | cambisol | 15.22 | ||
| 2015 | 48°57′36.249″ N, 16°17′15.639″ E | cambisol | 7.66 | ||
| 2016 | 48°57′20.646″ N, 16°17′10.774″ E | cambisol | 19.65 | ||
| 2017 | 48°57′4.665″ N, 16°16′57.111″ E | chernozem, cambisol | 15.05 | ||
| 2018 | 48°56′47.907″ N, 16°17′33.096″ E | chernozem | 4.63 | ||
| 2020 | 48°56′49.408″ N, 16°17′40.762″ E | chernozem | 1.66 | ||
| 2021 | 48°56′59.473″ N, 16°17′39.740″ E | chernozem | 4.54 | ||
| Miroslav | U vinohradu | 2003 | 48°56′33.258″ N, 16°18′4.029″ E | chernozem | 4.37 |
| 2004 | 48°56′35.288″ N, 16°17′52.471″ E | chernozem, cambisol | 2.69 | ||
| 2007 | 48°56′42.228″ N, 16°17′59.485″ E | chernozem | 2.79 | ||
| 2014 | 48°56′34.780″ N, 16°17′56.997″ E | chernozem | 1.22 | ||
| 2019 | 48°56′40.443″ N, 16°17′51.188″ E | chernozem | 1.58 | ||
| Miroslav | Weinperky I | 1996 | 48°55′55.163″ N, 16°18′56.924″ E | chernozem, cambisol | 9.14 |
| 2011 | 48°56′23.077″ N, 16°19′11.841″ E | chernozem, cambisol | 5.89 | ||
| 2014 | 48°56′15.517″ N, 16°19′1.103″ E | cambisol | 2.4 | ||
| 2015 | 48°56′9.683″ N, 16°19′2.032″ E | chernozem | 5.98 | ||
| 2017 | 48°56′11.229″ N, 16°18′54.074″ E | cambisol | 2.44 | ||
| Miroslav | Weinperky II | 1996 | 48°56′22.790″ N, 16°18′25.823″ E | chernozem | 2.35 |
| 1998 | 48°56′20.679″ N, 16°18′18.850″ E | chernozem | 8.63 | ||
| 1999 | 48°55′57.286″ N, 16°18′19.773″ E | cambisol | 3.91 | ||
| 2000 | 48°56′2.005″ N, 16°18′18.074″ E | cambisol | 3.91 | ||
| 2001 | 48°56′13.216″ N, 16°18′32.461″ E | cambisol | 8.63 | ||
| 2002 | 48°56′24.941″ N, 16°18′9.886″ E | cambisol | 2.48 | ||
| 48°56′5.050″ N, 16°18′30.125″ E | |||||
| 2003 | 48°56′18.939″ N, 16°18′21.503″ E | chernozem | 8.63 | ||
| 2004 | 48°56′23.500″ N, 16°18′22.001″ E | chernozem | 1.96 | ||
| 2008 | 48°56′6.471″ N, 16°18′21.396″ E | cambisol | 4.96 | ||
| Miroslavské Knínice | Stará hora | 2001 | 48°58′26.916″ N, 16°19′39.700″ E | chernozem, cambisol | 5.35 |
| Zolos | 2011 | 48°58′37.253″ N, 16°20′0.871″ E | cambisol | 0.23 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hurajová, E.; Martínez Barroso, P.; Děkanovský, I.; Lumbantobing, Y.R.; Jiroušek, M.; Mugutdinov, A.; Havel, L.; Winkler, J. Biodiversity and Vegetation Succession in Vineyards, Moravia (Czech Republic). Agriculture 2024, 14, 1036. https://doi.org/10.3390/agriculture14071036
Hurajová E, Martínez Barroso P, Děkanovský I, Lumbantobing YR, Jiroušek M, Mugutdinov A, Havel L, Winkler J. Biodiversity and Vegetation Succession in Vineyards, Moravia (Czech Republic). Agriculture. 2024; 14(7):1036. https://doi.org/10.3390/agriculture14071036
Chicago/Turabian StyleHurajová, Erika, Petra Martínez Barroso, Igor Děkanovský, Yentriani Rumeta Lumbantobing, Martin Jiroušek, Amir Mugutdinov, Ladislav Havel, and Jan Winkler. 2024. "Biodiversity and Vegetation Succession in Vineyards, Moravia (Czech Republic)" Agriculture 14, no. 7: 1036. https://doi.org/10.3390/agriculture14071036
APA StyleHurajová, E., Martínez Barroso, P., Děkanovský, I., Lumbantobing, Y. R., Jiroušek, M., Mugutdinov, A., Havel, L., & Winkler, J. (2024). Biodiversity and Vegetation Succession in Vineyards, Moravia (Czech Republic). Agriculture, 14(7), 1036. https://doi.org/10.3390/agriculture14071036