
Nectar Production and Three Main Sugars in Nectar of Salvia pratensis and Salvia glutinosa in Correlation with Abiotic Factors


Abstract
1. Introduction
2. Materials and Methods
2.1. Researched Plant Species
2.2. Sampling Locations
2.3. Nectar Sampling
2.3.1. Flower Protection
2.3.2. Nectar Collection
2.3.3. Nectar Quantity
2.3.4. Measurement of Abiotic Factors
2.4. Nectar Analysis
2.4.1. Sample and Standard Solutions
2.4.2. Sugar Analysis
2.5. Statistical Analysis
3. Results
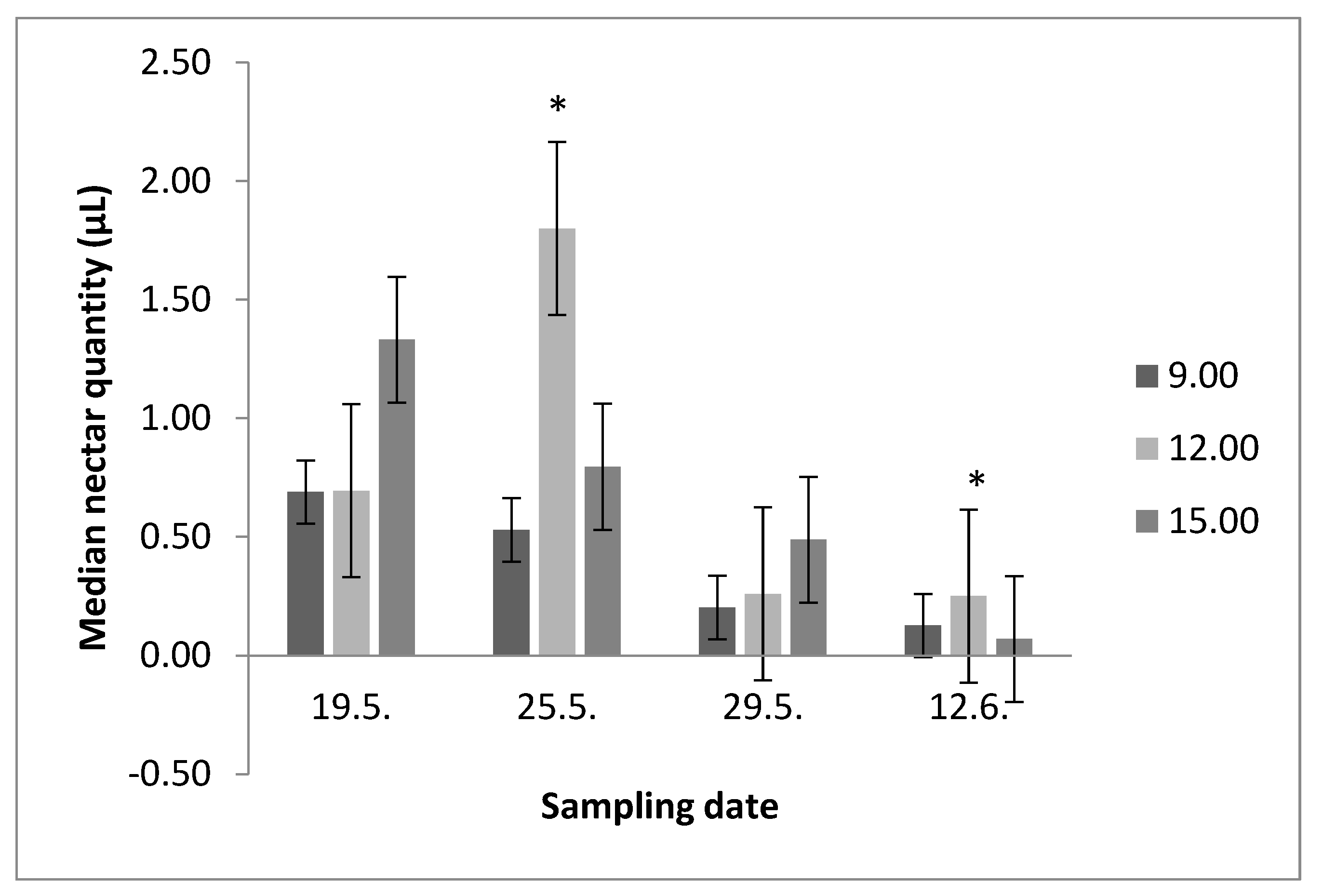
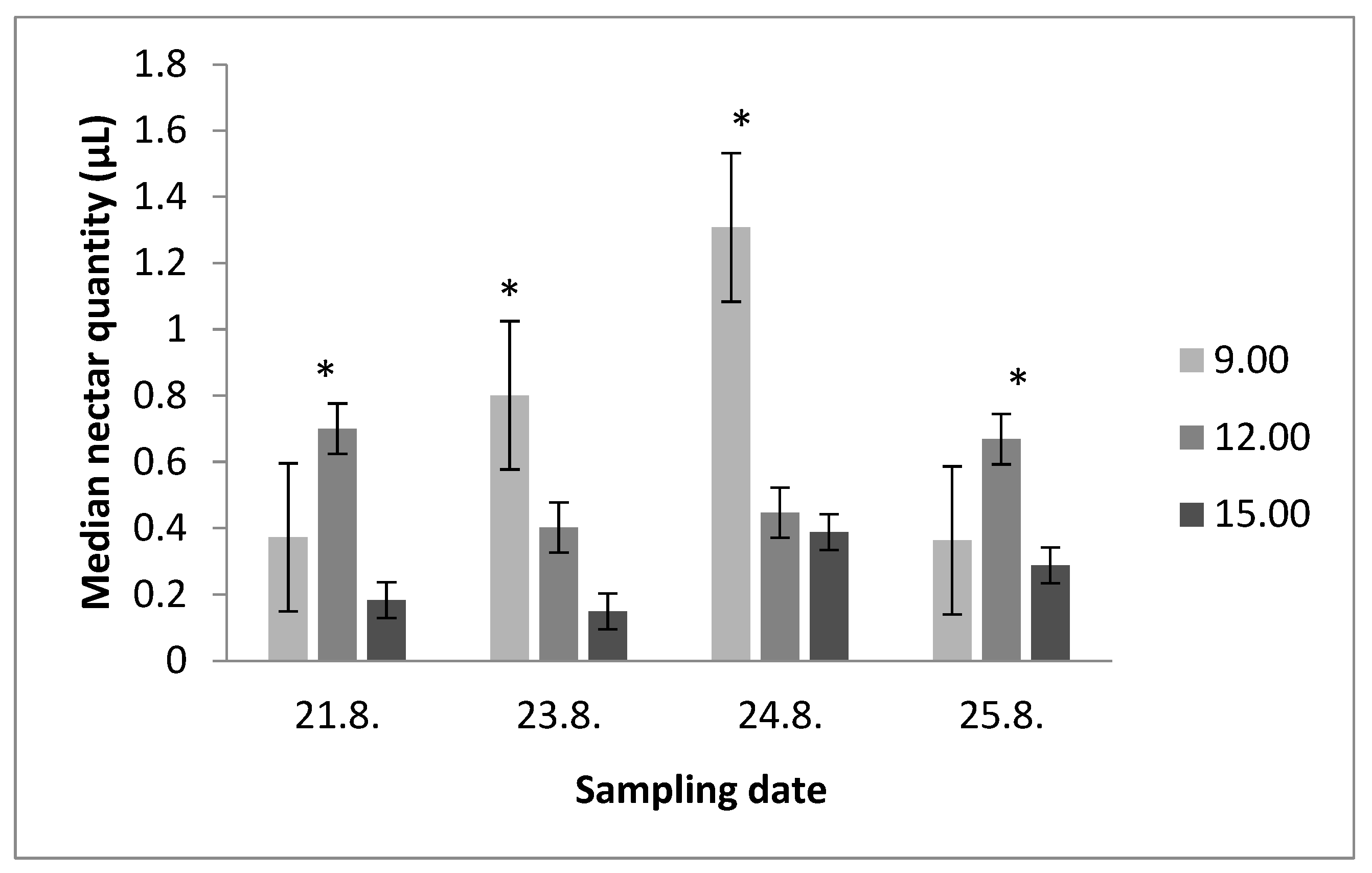
3.1. Nectar Secretion throughout the Day
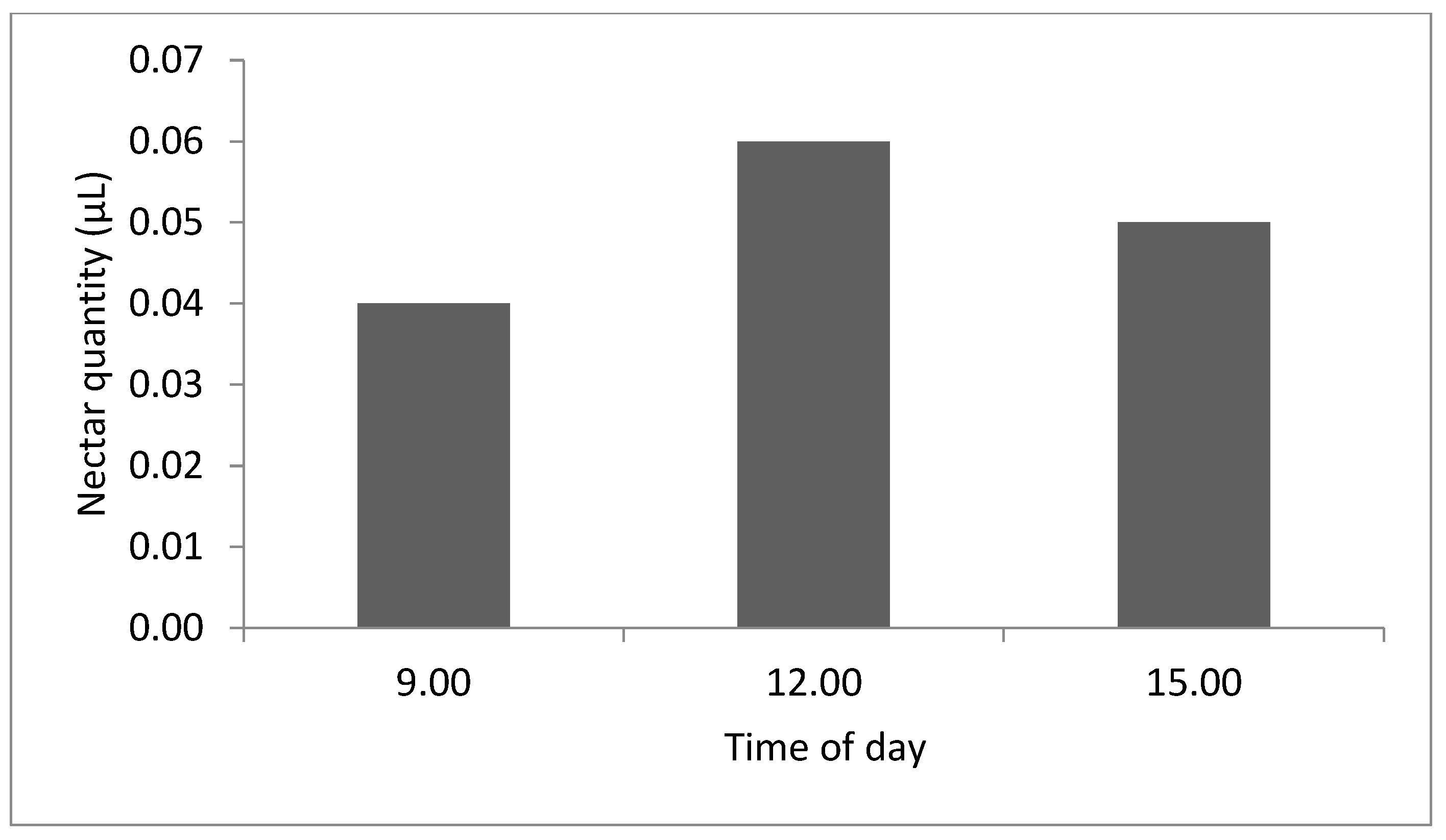
3.2. Average Nectar Production throughout the Day
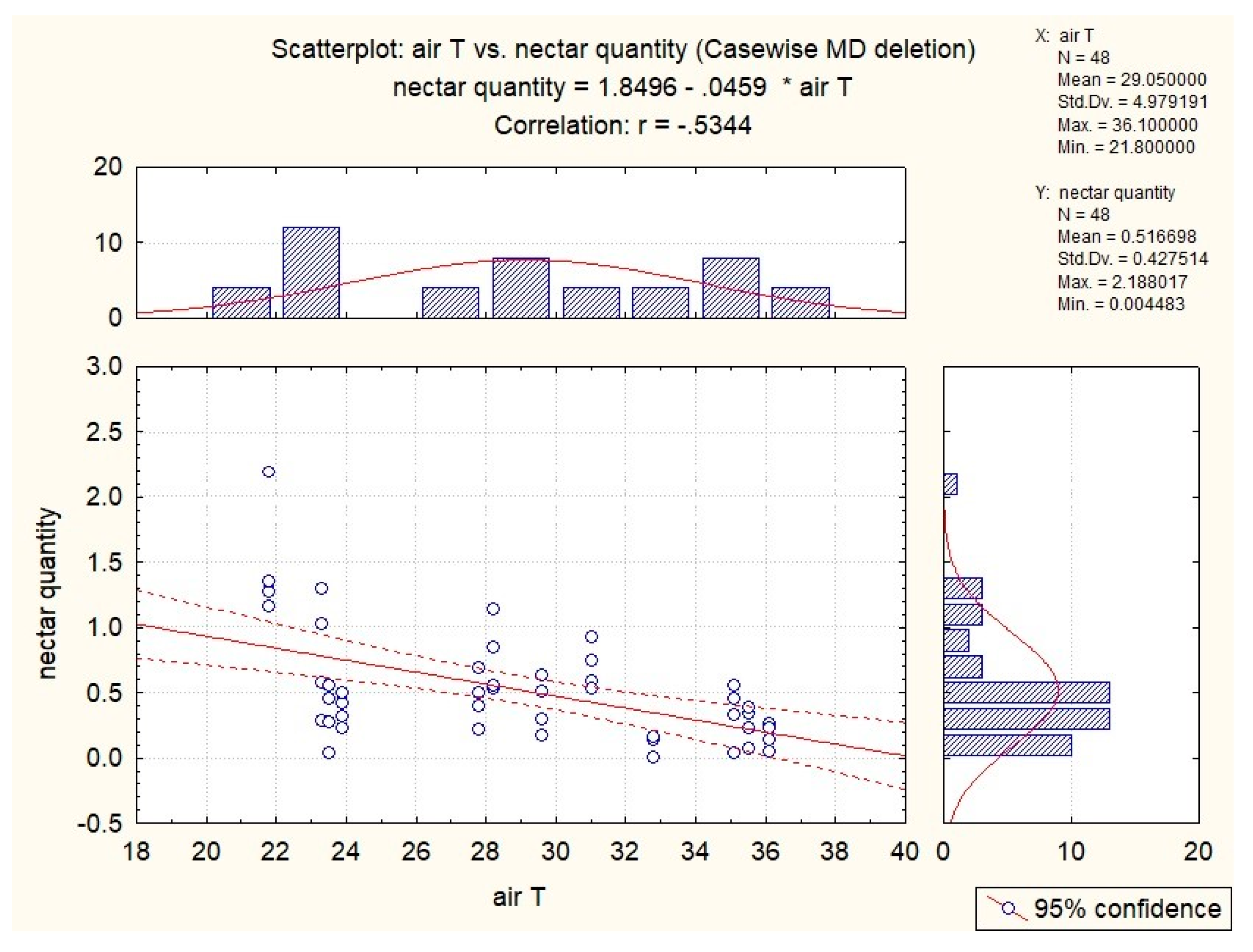
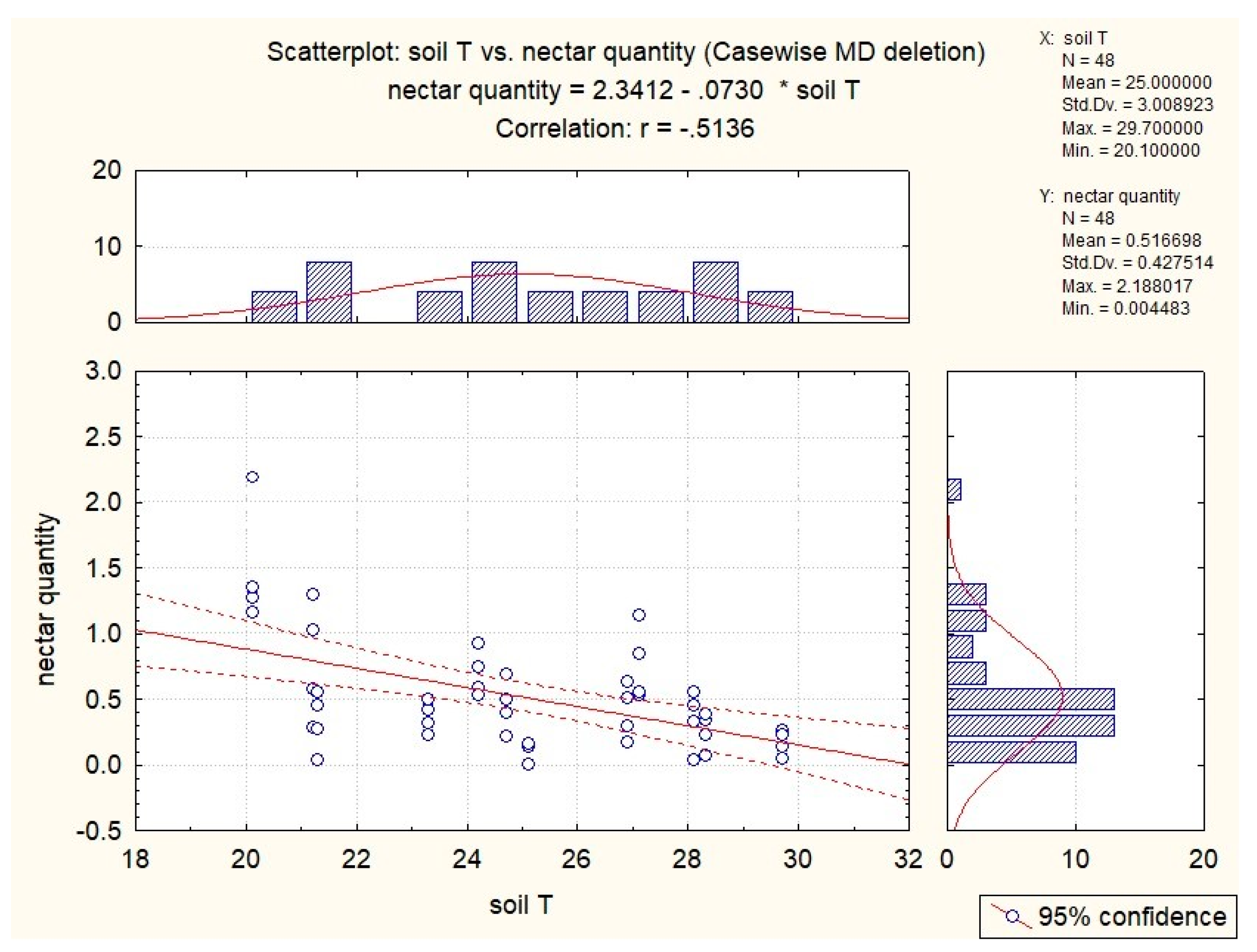
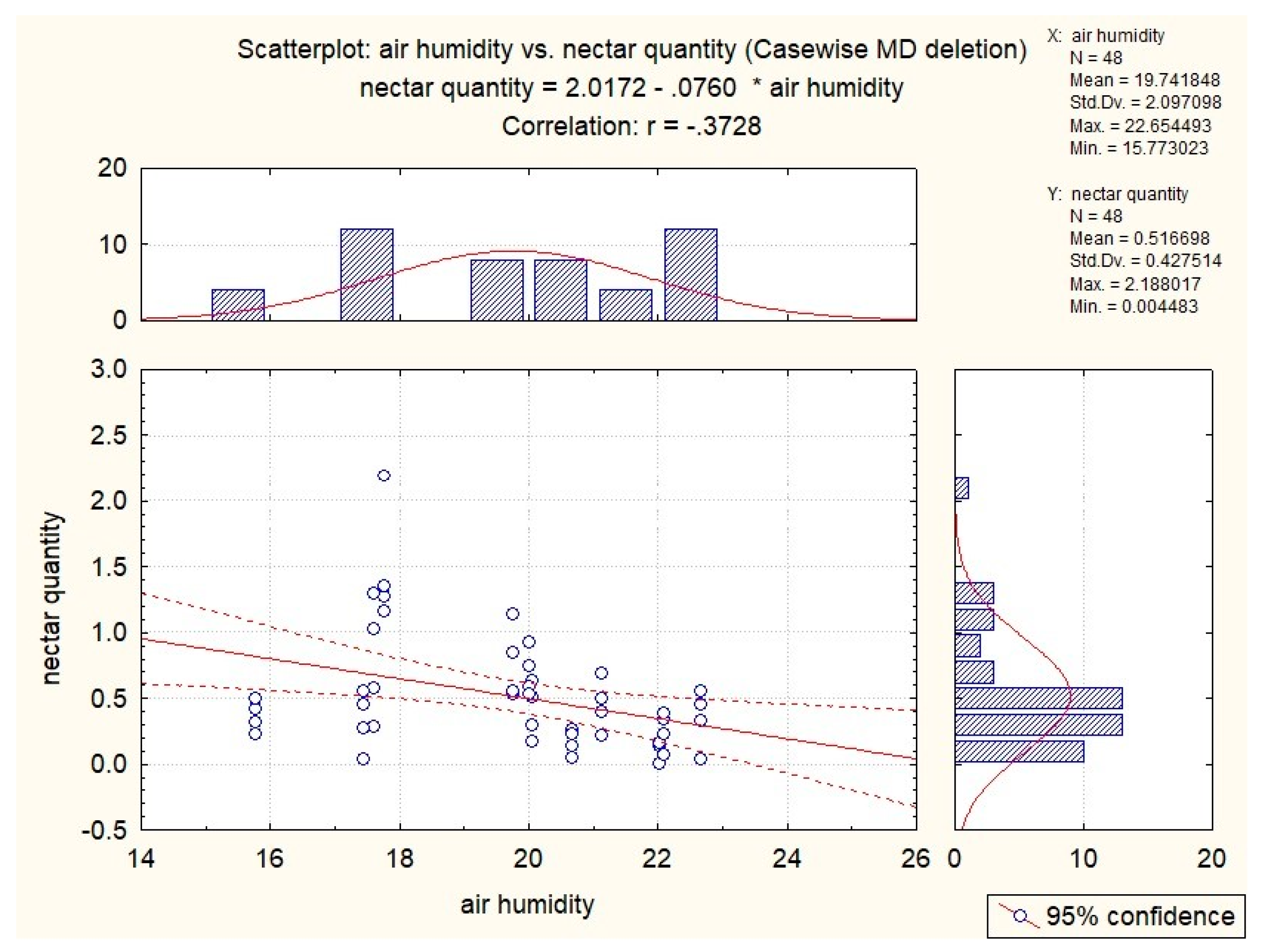
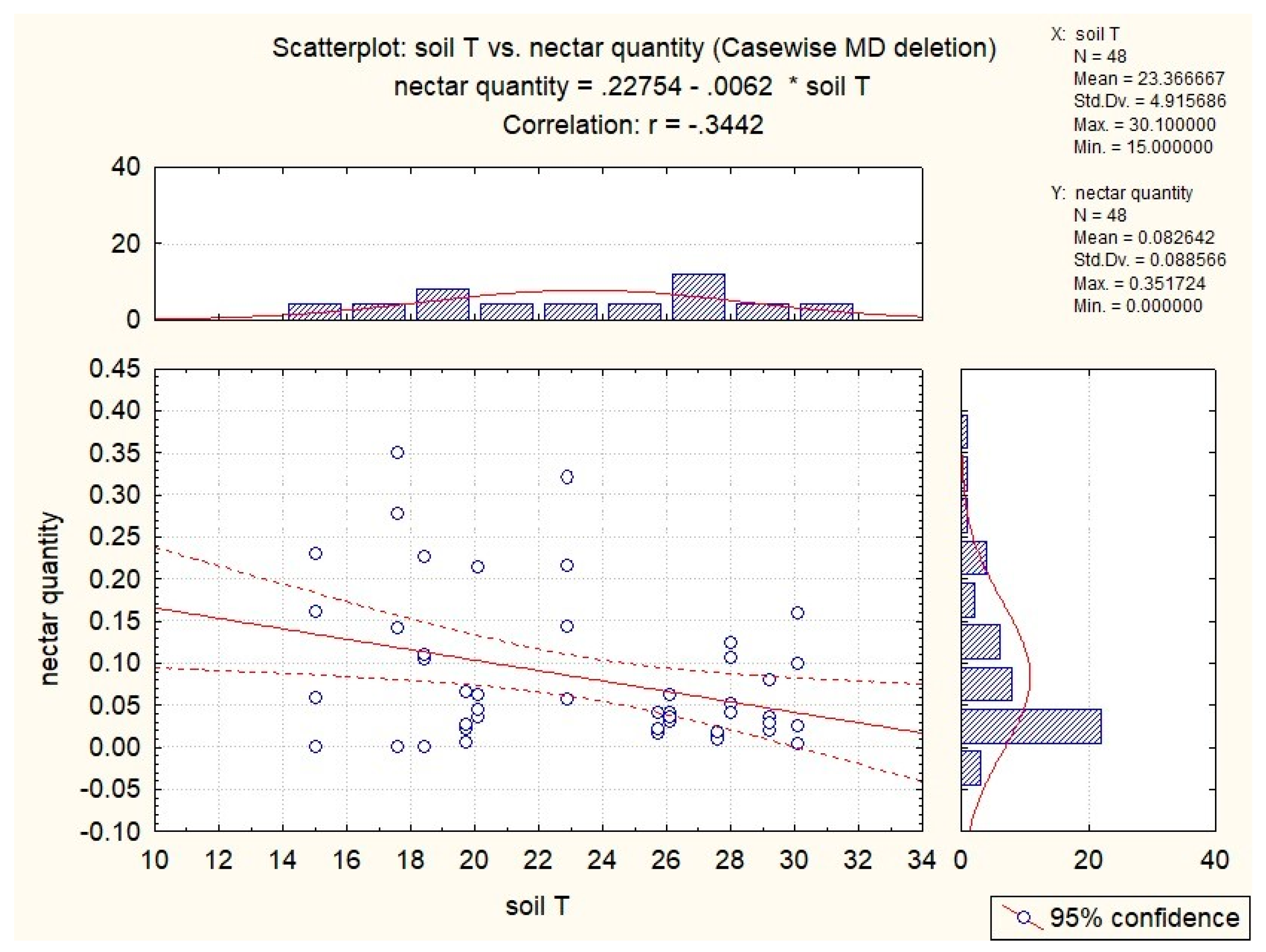
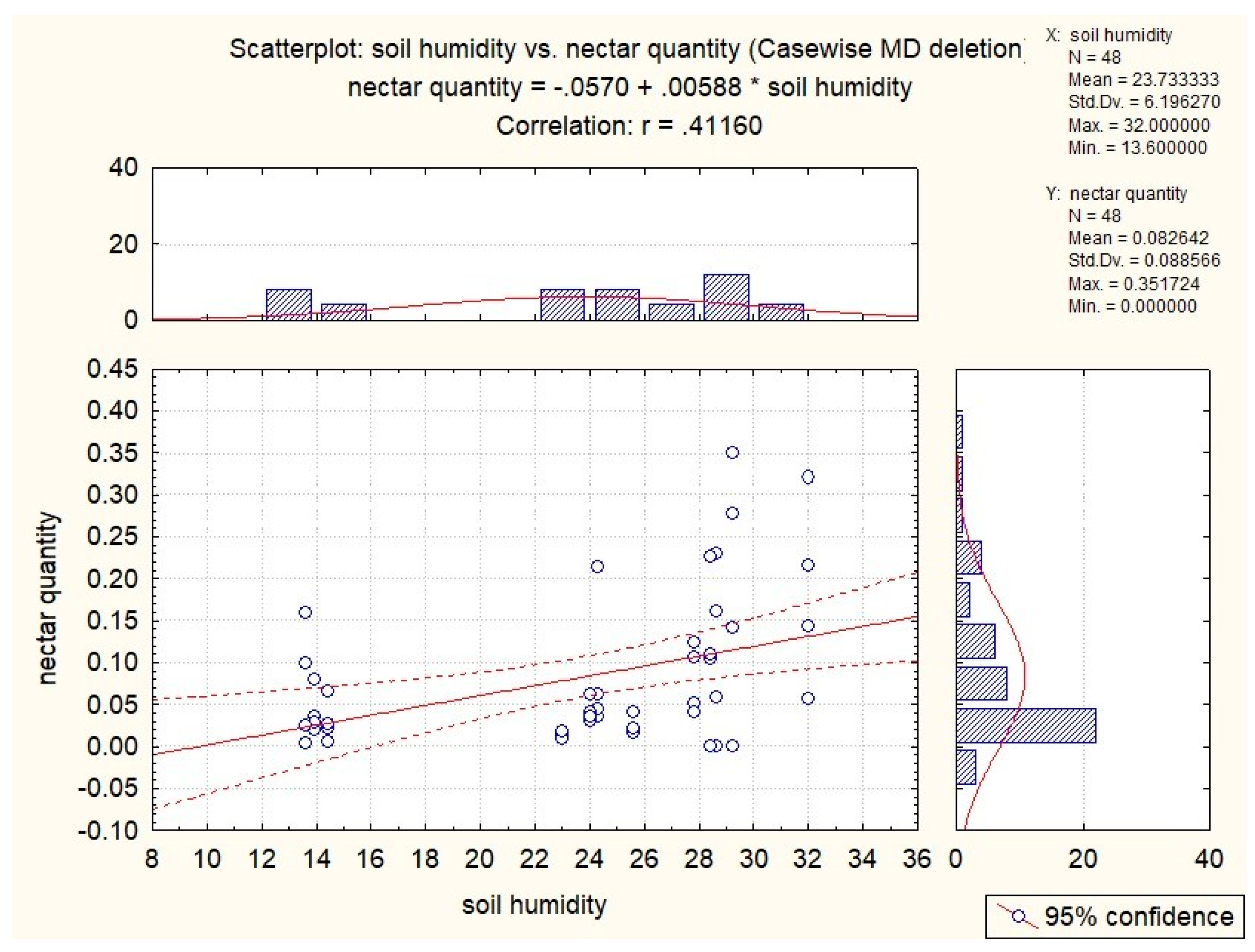
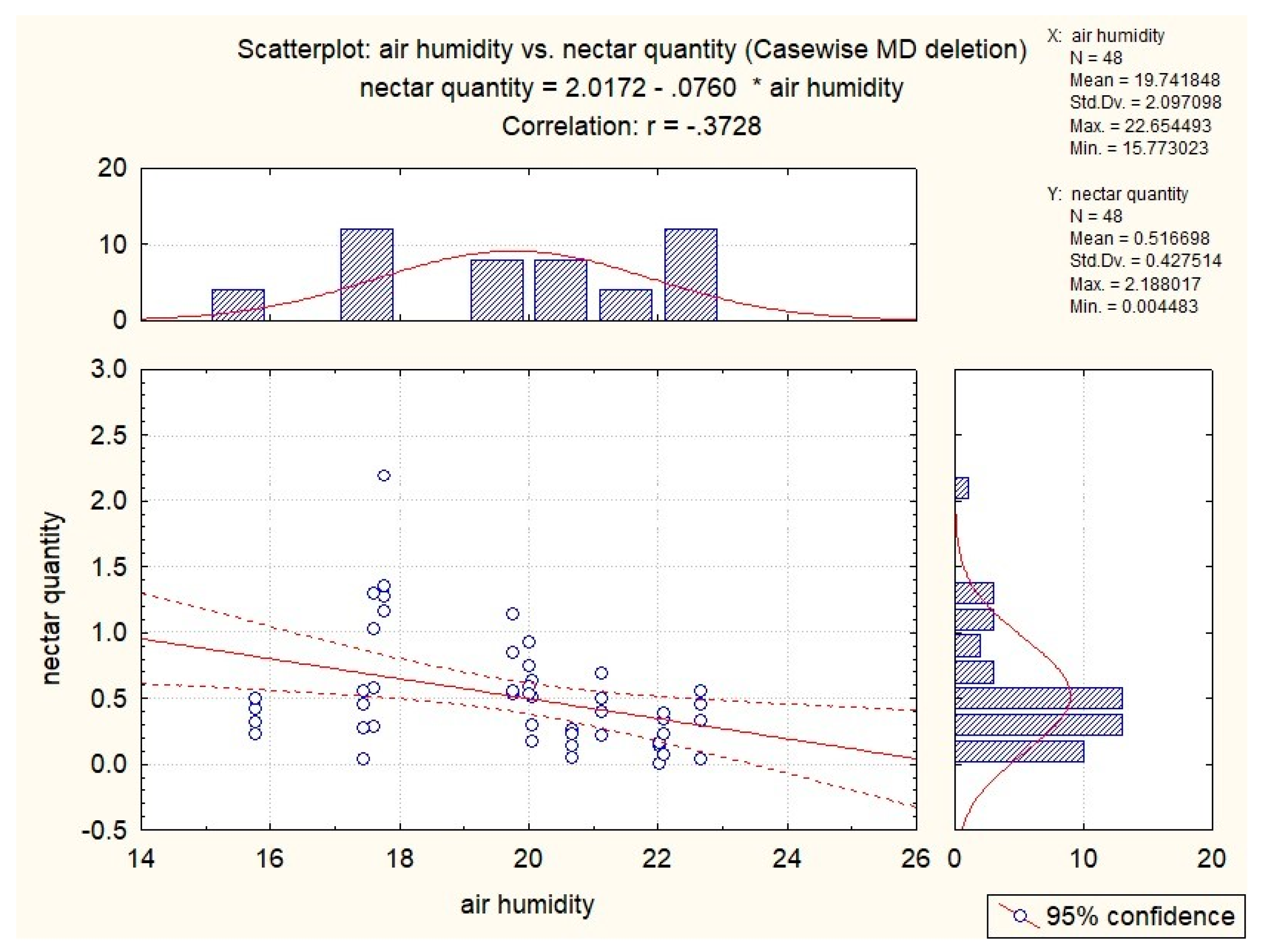
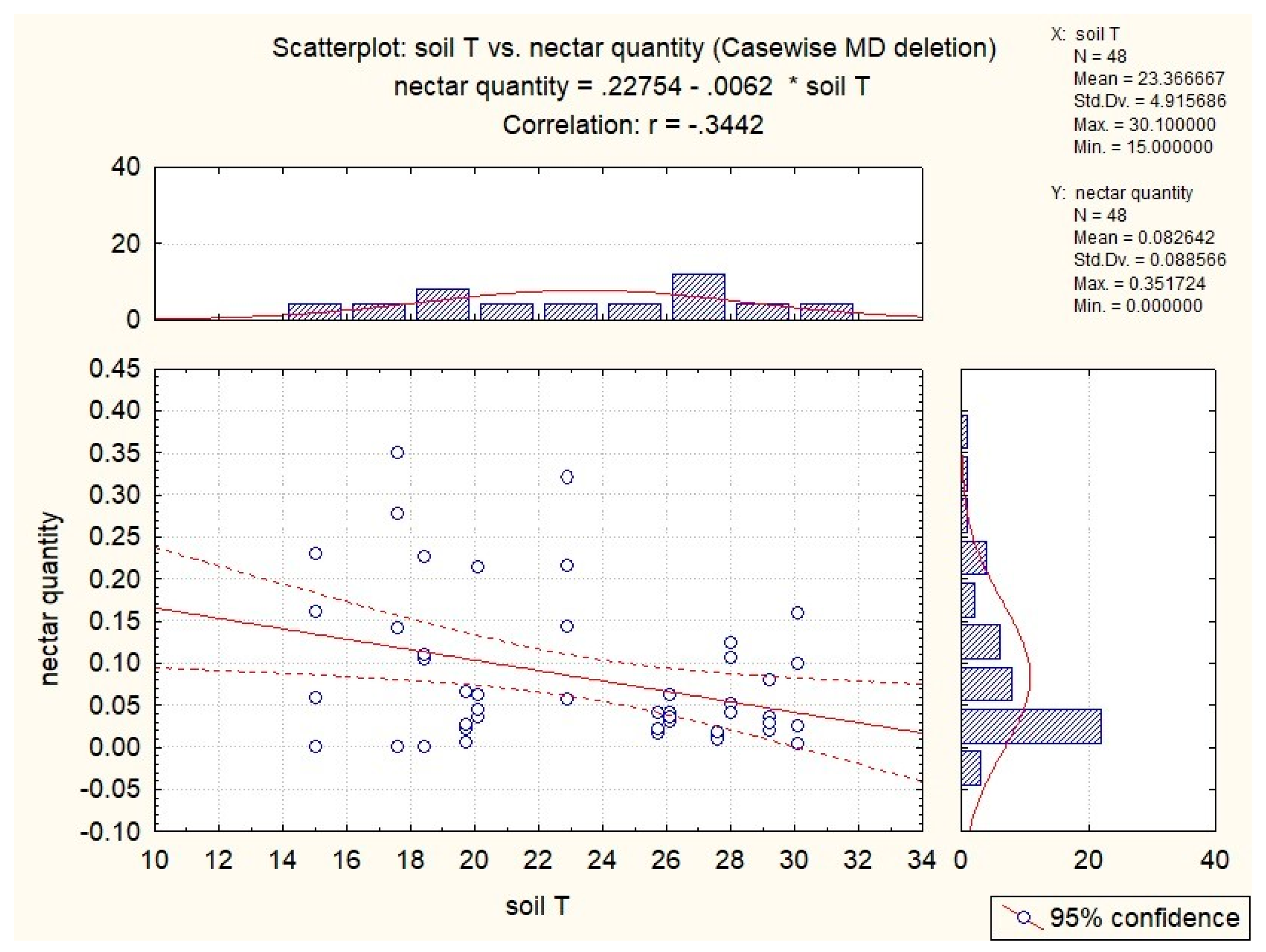
3.3. Influence of Abiotic Factors on Nectar Production
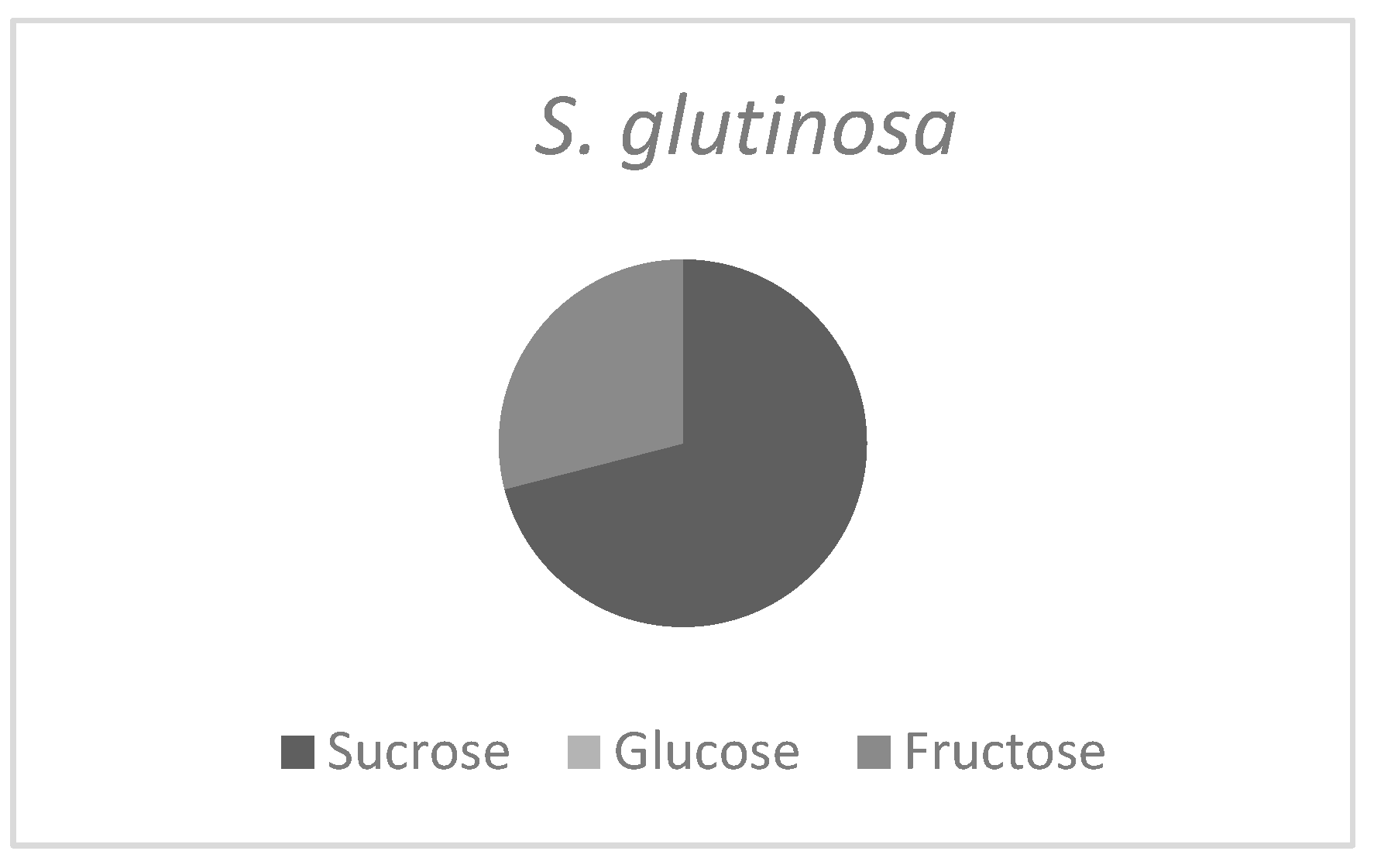
3.4. Ratio of Three Main Sugars in Salvia Nectar Samples
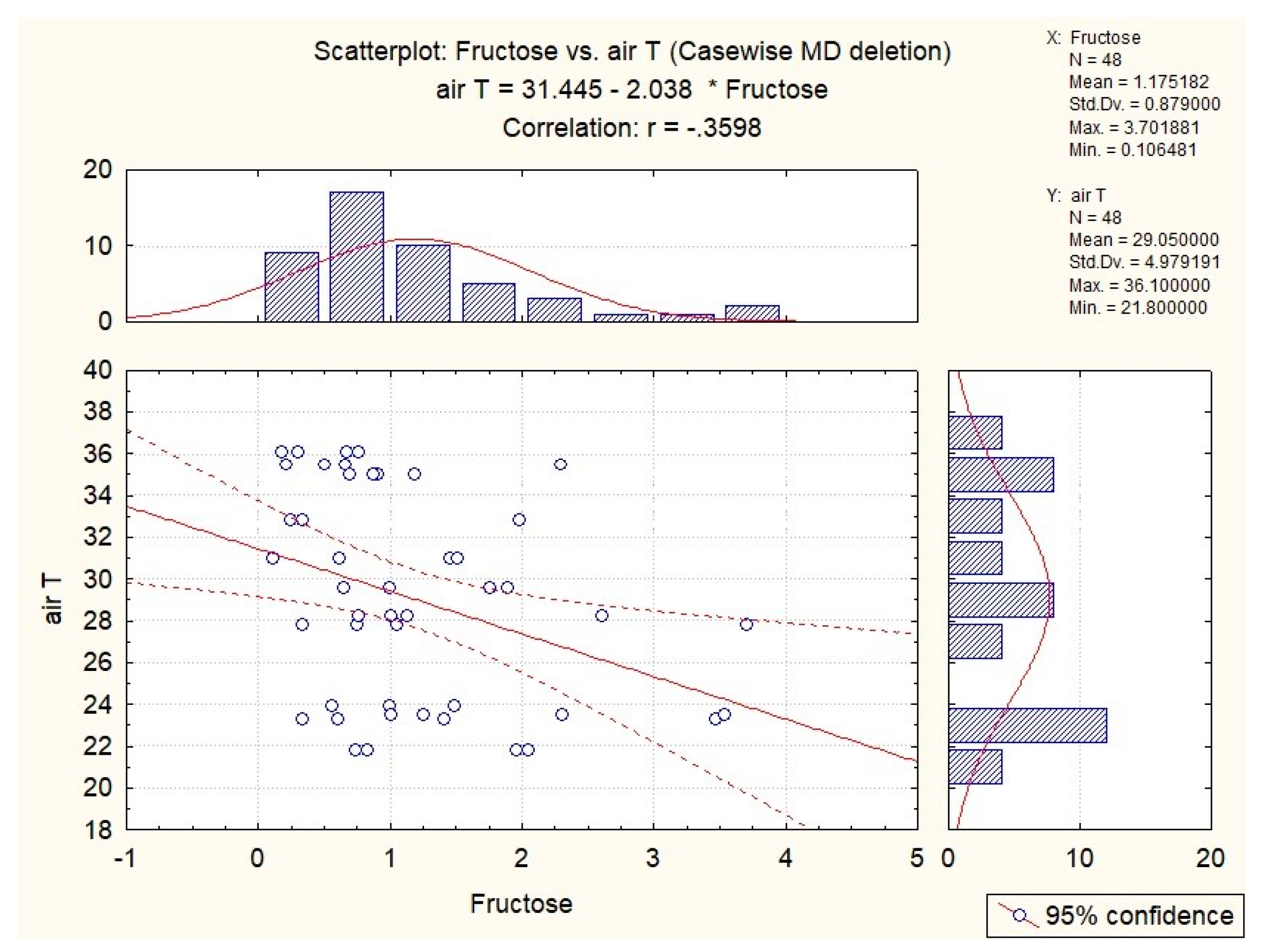
3.5. Influence of Abiotic Factors on Sugar Concentrations in S. pratensis Nectar
3.6. Influence of Abiotic Factors on Sugar Concentrations in S. glutinosa Nectar
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Buchmann, S.L.; Naphan, G.P. The Forgotten Pollinators; Island Press: Washington, DC, USA, 1997; 292p. [Google Scholar]
- Saunders, M.E. Insect pollinators collect pollen from wind-pollinated plants: Implications for pollination ecology and sustainable agriculture. Insect Conserv. Divers. 2017, 11, 13–31. [Google Scholar] [CrossRef]
- Knuth, P. Handbook of Flower Pollination Based upon Herman’s Muller Work, the Fertilisation of Flowers by Insects; The Clarendon Press: Oxford, UK, 1906; 383p. [Google Scholar]
- Nepi, M.; Grasso, D.A.; Mancuso, S. Nectar in plant–insect mutualistic relationships: From food reward to partner manipulation. Front. Plant Sci. 2018, 9, 10–63. [Google Scholar] [CrossRef] [PubMed]
- Kulloli, S.K.; Chandore, A.; Aitawade, M.M. Nectar dynamics and pollination studies in three species of Lamiaceae. Curr. Sci. 2011, 100, 509–516. [Google Scholar]
- Roy, R.; Schmitt, A.J.; Thomas, J.B.; Carter, C.J. Rewiew: Nectar biology: From molecules to ecosystems. Plant Sci. 2017, 262, 148–164. [Google Scholar] [CrossRef] [PubMed]
- Nicolson, S.W.; Thornburg, R.W. Nectar chemistry. In Nectaries and Nectar; Nicolson, S., Nepi, M., Pacini, E., Eds.; Springer: New York, NY, USA, 2007; pp. 215–263. [Google Scholar]
- Ballantyne, G.; Wilmer, P. Nectar theft and floral ant-repellence: A link between nectar volume and ant-repellent traits? PLoS ONE 2012, 7, e43869. [Google Scholar] [CrossRef] [PubMed]
- Pacini, E.; Nicolson, S.W. Introduction. In Nectaries and Nectar; Nicolson, S., Nepi, M., Pacini, E., Eds.; Springer: New York, NY, USA, 2007. [Google Scholar]
- Baker, G.; Baker, I. Amino-acids in nectar and their evolutionary significance. Nature 1973, 241, 543–545. [Google Scholar] [CrossRef]
- Nepi, M. Nectary structure and ultrastructure. In Nectaries and Nectar; Nicolson, S., Nepi, M., Pacini, E., Eds.; Springer: New York, NY, USA, 2007. [Google Scholar]
- Nikolić, T. Morfologija biljaka—Razvoj, građa i uloga biljnih tkiva, organa i organskih sustava; Alfa d.d.: Zagreb, Croatia, 2017. [Google Scholar]
- Pacini, E.; Nepi, M.; Vesprini, J.L. Nectar biodiversity: A short review. Plant Syst. Evol. 2003, 238, 7–21. [Google Scholar] [CrossRef]
- Pozo, M.I.; Lievens, B.; Jacquemyn, H. Impact of microorganisms on nectar chemistry, pollinator attraction and plant fitness. In Nectar: Production, Chemical Composition and Benefits to Animals and Plants; Peck, R.L., Ed.; Nova Science Publishers Inc.: New York, NY, USA, 2015; pp. 1–45. [Google Scholar]
- Irwin, R.E.; Cook, D.; Richardson, L.L.; Manson, J.S.; Gardner, D.R. Secondary compounds in floral rewards of toxic rangeland plants: Impacts on pollinators. J. Agric. Food Chem. 2014, 62, 7335–7344. [Google Scholar] [CrossRef] [PubMed]
- Hagler, J.R.; Buchmann, S.L. Honeybee (Hymenoptera: Apidae) foraging responses to phenolic-rich nectars. J. Kans. Entomol. Soc. 1993, 66, 223–230. [Google Scholar]
- Palmer-Young, E.C.; Farrell, I.W.; Adler, L.S.; Milano, N.J.; Egan, P.A.; Junker, R.R.; Irwin, R.E.; Stevenson, P.C. Chemistry of floral rewards: Intra- and interspecific variability of nectar and pollen secondary metabolites across taxa. Ecol. Monogr. 2019, 89, e01335. [Google Scholar] [CrossRef]
- Chalcoff, V.R.; Aizen, M.A.; Galetto, L. Nectar concentration and composition of 26 species from the temperate forest of South America. Ann. Bot. 2006, 97, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Lanza, J.; Smith, G.C.; Sack, S.; Cash, A. Variation in nectar volume and composition of Impatiens capensis at the individual, plant, and population levels. Oecologia 1995, 102, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Leiss, K.A.; Klinkhamer, P.G.L. Genotype by environment interactions in the nectar production of Echium vulgare. Funct. Ecol. 2005, 19, 454–459. [Google Scholar] [CrossRef]
- Pacini, E.; Nepi, M. Nectar production and presentation. In Nectaries and Nectar; Nicolson, S., Nepi, M., Pacini, E., Eds.; Springer: New York, NY, USA, 2007. [Google Scholar]
- Corbet, S.A. Nectar sugar content: Estimating standing crop and secretion rate in the field. Apidologie 2003, 34, 1–10. [Google Scholar] [CrossRef]
- Morrant, D.S.; Schumann, R.; Petit, S. Field methods for sampling and storing nectar from flowers with low nectar volumes. Ann. Bot. 2008, 103, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Mačukanović Jocić, M.; Duletić-Laušević, S.; Jocic, G. Nectar production in three melliferous species of Lamiaceae in natural and experimental conditions. Acta Vet. 2004, 54, 475–487. [Google Scholar]
- Cardoza, Y.J.; Harris, G.K.; Grozinger, C.M. Effects of soil quality enhancement on pollinator-plant interactions. Psyche J. Entomol. 2012, 2012, 581458. [Google Scholar] [CrossRef]
- Corbet, S.A. Pollination and the weather. Isr. J. Bot. 1990, 39, 13–30. [Google Scholar]
- Corbet, S.A.; Willmer, P.G.; Beament, J.W.L.; Unwin, D.M.; Prys-Jones, O.E. Post-secretory determinants of sugar concentration in nectar. Plant Cell Environ. 1979, 2, 293–308. [Google Scholar] [CrossRef]
- Mačukanović-Jocić, M.; Djurdjević, L. Influence of microclimatic conditions on nectar exudation in Glechoma hirsuta. Arch. Biol. Sci. 2005, 57, 119–126. [Google Scholar] [CrossRef]
- Petanidou, T. Sugars in Mediterranean Floral Nectars: An Ecological and Evolutionary Approach. J. Chem. Ecol. 2005, 31, 1065–1087. [Google Scholar] [CrossRef]
- Búrquez, A.; Corbet, S.A. Do flowers reabsorb nectar? Funct. Ecol. 1991, 5, 369–379. [Google Scholar] [CrossRef]
- Búrquez, A.; Corbet, S.A. Dynamics of production and exploitation of nectar: Lessons from Impatiens glandulifera Royle. In Nectary Biology; Bahadur, B., Ed.; Dattsons: Nagpur, India, 1998; pp. 130–152. [Google Scholar]
- Kradolfer, U.; Erhardt, A. Nectar secretion patterns in Salvia pratensis L. (Lamiaceae). Flora 1995, 190, 229–235. [Google Scholar] [CrossRef]
- Sampson, B.J.; Cane, J.H. Impact of enhanced ultraviolet-B radiation on flower, pollen, and nectar production. Am. J. Bot. 1999, 86, 108–114. [Google Scholar] [CrossRef]
- Bertazzini, M.; Forlani, G. Intraspecific variability of floral nectar volume and composition in rapeseed (Brassica napus L. var. oleifera). Front. Plant Sci. 2016, 7, 288. [Google Scholar] [CrossRef]
- Kostryco, M.; Chwil, M. Nectar abundance and nectar composition in selected Rubus idaeus L. varieties. Agriculture 2022, 12, 1132. [Google Scholar] [CrossRef]
- Percival, M.S. Types of nectar in angiosperms. New Phytol. 1961, 60, 235–281. [Google Scholar] [CrossRef]
- Schondube, J.E.; Martınez del Rio, C. Concentration-dependent sugar preferences in nectar-feeding birds: Mechanisms and consequences. Funct. Ecol. 2003, 17, 445–453. [Google Scholar] [CrossRef]
- Scheiner, R.; Page, R.E.; Erber, J. Sucrose responsiveness and behavioral plasticity in honey bees (Apis mellifera). Apidologie 2004, 35, 133–142. [Google Scholar] [CrossRef]
- Kim, W.; Gilet, T.; Bush, J.W.M. Optimal concentrations in nectar feeding. Proc. Natl. Acad. Sci. USA 2011, 108, 16618–16621. [Google Scholar] [CrossRef]
- Ravnjak, B.; Bavcon, J.; Božič, J. Avtohtone Medovite Rastline; Botanični vrt Univerze v Ljubljani, Oddelek za biologijo, Biotehniška fakulteta = University Botanic Gardens Ljubljana, Department of Biology, Biotechnical Faculty: Ljubljana, Slovenia, 2020; 285p. [Google Scholar]
- Weryszko-Chmielweska, E. Ecological features of flowers including nectary structure of chosen species from Lamiaceae family. Fitoterapia 2000, 69, 191–192. [Google Scholar]
- Celep, F.; Atalay, Z.; Dikmen, F.; Doǧan, M.; Sytsma, K.J.; Claßen-Bockhoff, R. Pollination Ecology, Specialization, and Genetic Isolation in Sympatric Bee-Pollinated Salvia (Lamiaceae). Int. J. Plant Species 2020, 181, 800–811. [Google Scholar] [CrossRef]
- Drew, B. Evolution, Pollination Biology, and Species Richness of Salvia (Lamiaceae). Int. J. Plant Sci. 2020, 181, 767–769. [Google Scholar] [CrossRef]
- Martinčič, A.; Wraber, T.; Jogan, N.; Podobnik, A.; Turk, B.; Vreš, B.; Ravnik, V.; Frajman, B.; Strgulc Krajšek, S.; Trčak, B.; et al. Mala flora Slovenije: Ključ za določanje praprotnic in semenk; Tehniška založba Slovenije: Ljubljana, Slovenia, 2007; 967p. [Google Scholar]
- Bavcon, J.; Ravnjak, B. Travniška kadulja (Salvia pratensis L.) v Sloveniji; Biotehniška fakulteta: Ljubljana, Slovenia, 2014. [Google Scholar]
- Reith, M.; Baumann, G.; Claßen-Bockoff, R.; Speck, T. New Insights into the Functional Morphology of the Lever Mechanism of Salvia pratensis (Lamiaceae). Ann. Bot. 2007, 100, 393–400. [Google Scholar] [CrossRef]
- Petanidou, T.; Goethals, V.; Smets, E. Nectary structure of Labiatae in relation to their nectar secretion and characteristics in a Mediterranean shrub community: Does flowering time matter? Plant Syst. Evol. 2000, 225, 103–118. [Google Scholar] [CrossRef]
- ARSO [Agencija Republike Slovenije za Okolje]. Available online: https://www.arso.gov.si/ (accessed on 15 February 2024).
- McKenna, M.A.; Thomson, J.D. A technique for sampling and measuring small amounts of floral nectar. Ecology 1988, 69, 1306–1307. [Google Scholar] [CrossRef]
- Herrera, C.M.; Garcia, I.M.; Perez, R. Invisible floral larcenies: Microbial communities degrade floral nectar of bumble bee-pollinated plants. Ecology 2008, 89, 2369–2376. [Google Scholar] [CrossRef] [PubMed]
- Vaudo, A.D.; Tooker, J.F.; Grozinger, C.M.; Patch, H.M. Bee nutrition and floral resource restoration. Curr. Opin. Insect. Sci. 2015, 10, 133–141. [Google Scholar] [CrossRef]
- Karbassioon, A.; Stanley, D.A. Exploring relationships between time of day and pollinator activity in the context of pesticide use. Basic Appl. Ecol. 2023, 72, 74–81. [Google Scholar] [CrossRef]
- Karbassioon, A.; Yearlsey, J.; Dirilgen, T.; Hodge, S.; Stout, J.C.; Stanley, D.A. Responses in honeybee and bumblebee activity to changes in weather conditions. Oecologia 2023, 201, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Willmer, P. Pollination and Floral Ecology; Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- Baker, H.G.; Baker, I. Floral nectar sugar constituents in relation to pollinator type. In Handbook of Experimental Pollination Ecology; Jones, C.E., Little, R.J., Van Nostrand, R., Eds.; Scientific and Academic Editions: Rosemead, CA, USA, 1983; pp. 117–141. [Google Scholar]
- Petanidou, T. Ecological and evolutionary aspects of oral nectars in Mediterranean habitats. In Nectaries and Nectar; Nicolson, S.W., Nepi, M., Pacini, E., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 343–377. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UVB (µW/cm2) | Air T (°C) | Soil T (°C) | Air AH (g/m3) | Soil RH (%) | |
|---|---|---|---|---|---|
| S. glutinosa | x | −0.53 | −0.51 | −0.37 | x |
| S. pratensis | x | x | −0.34 | x | 0.41 |
| UVB (µW/cm2) | Air T (°C) | Soil T (°C) | Air AH (g/m3) | Soil RH (%) | |
|---|---|---|---|---|---|
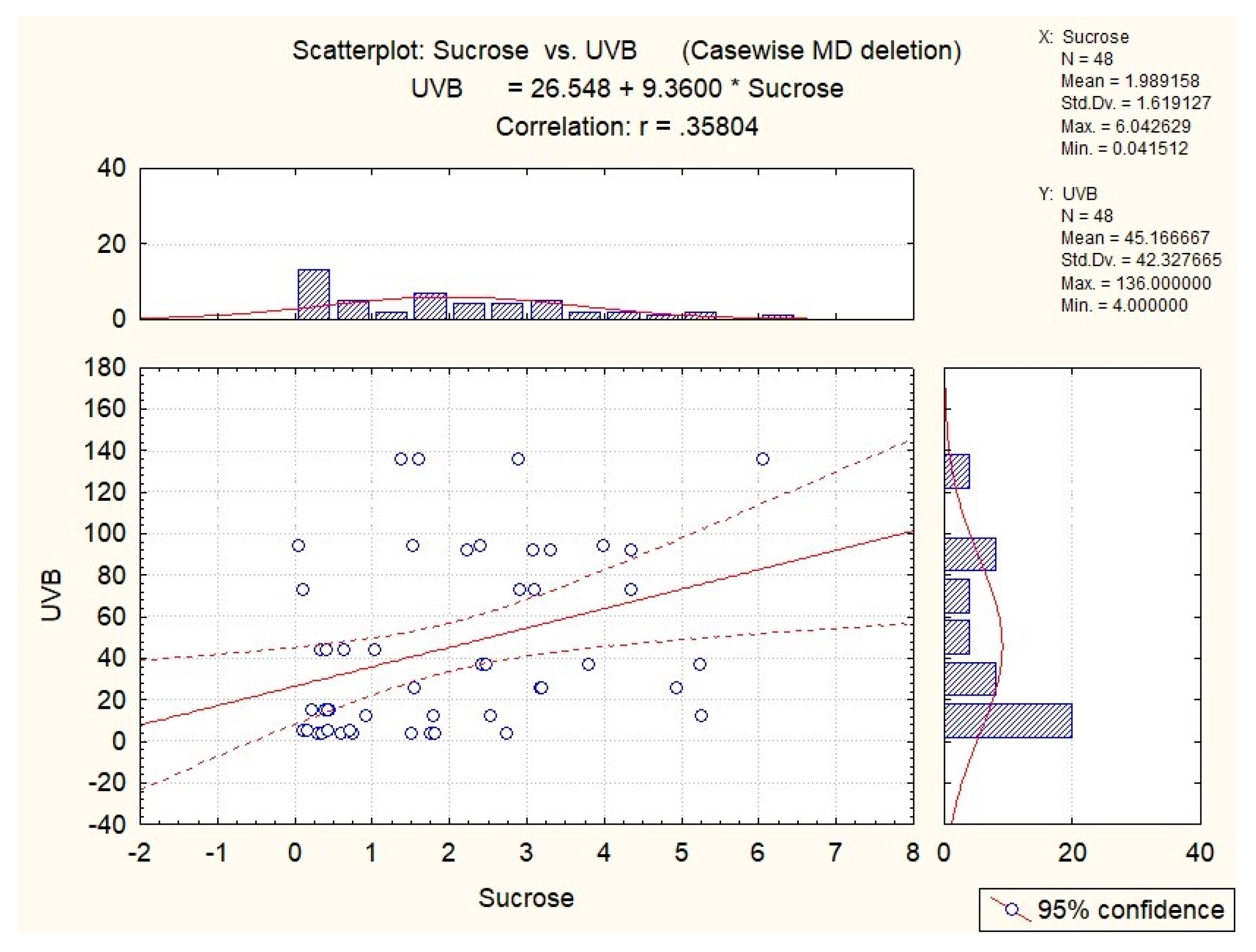
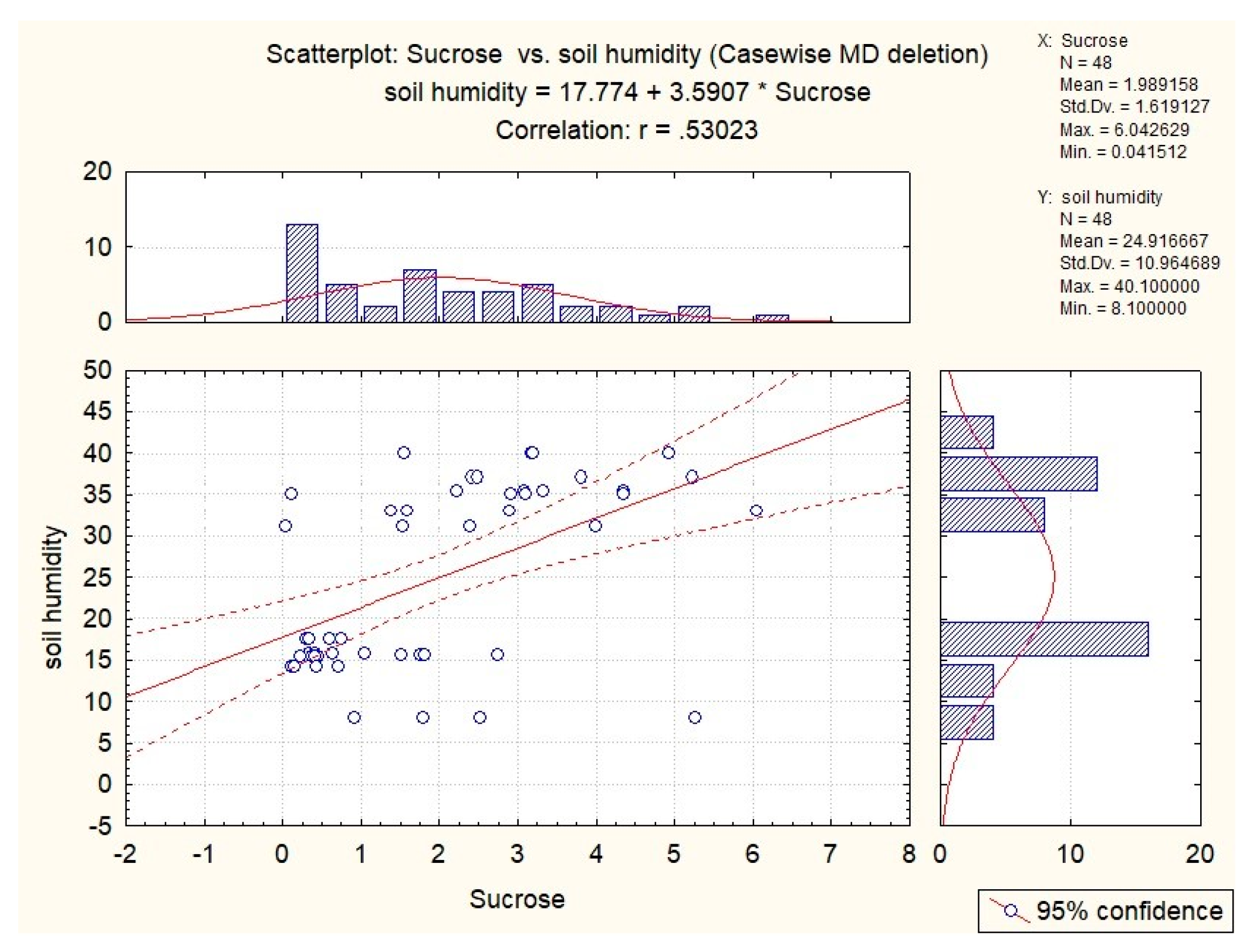
| sucrose | 0.35 | 0.31 | x | x | x |
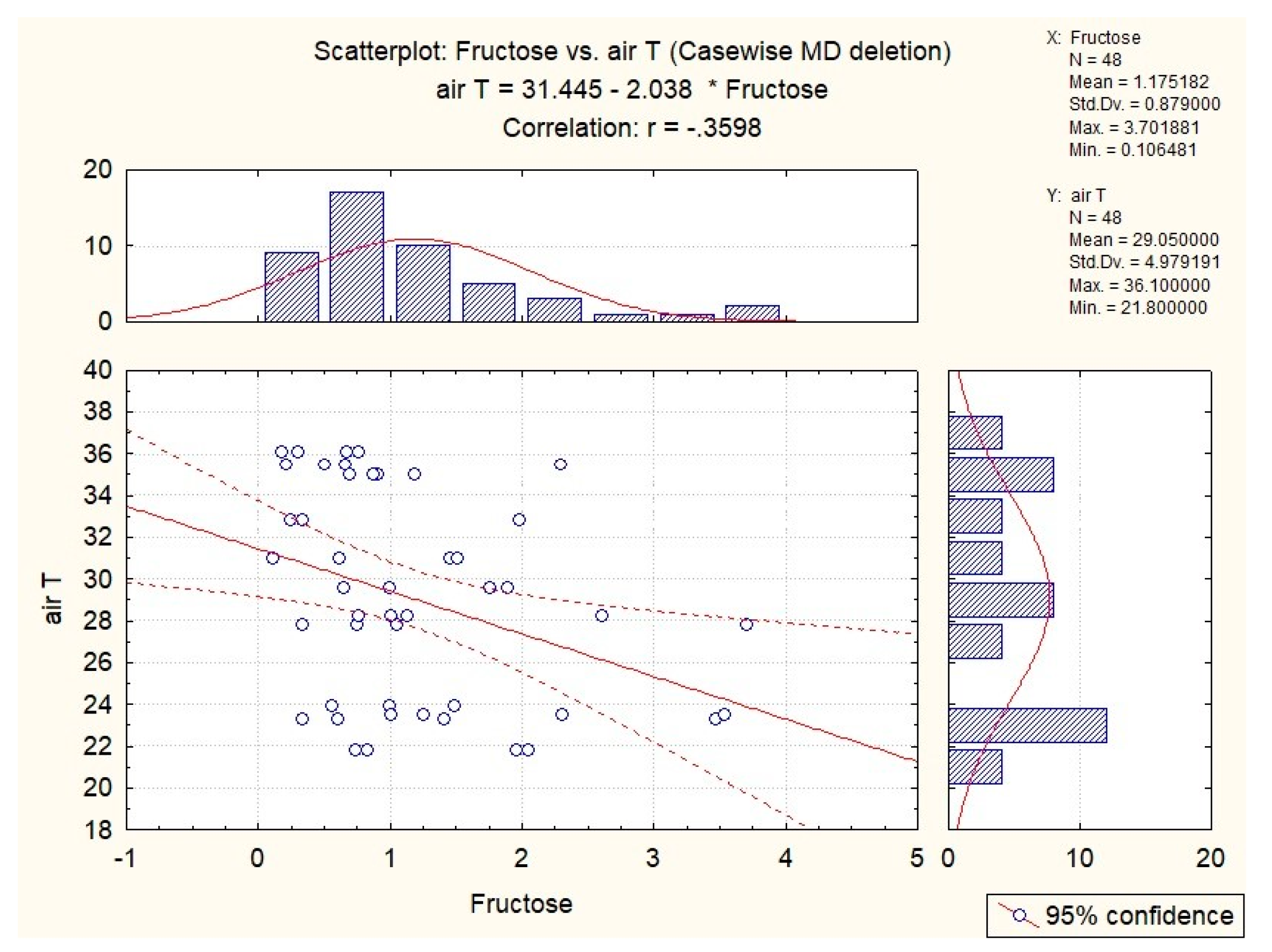
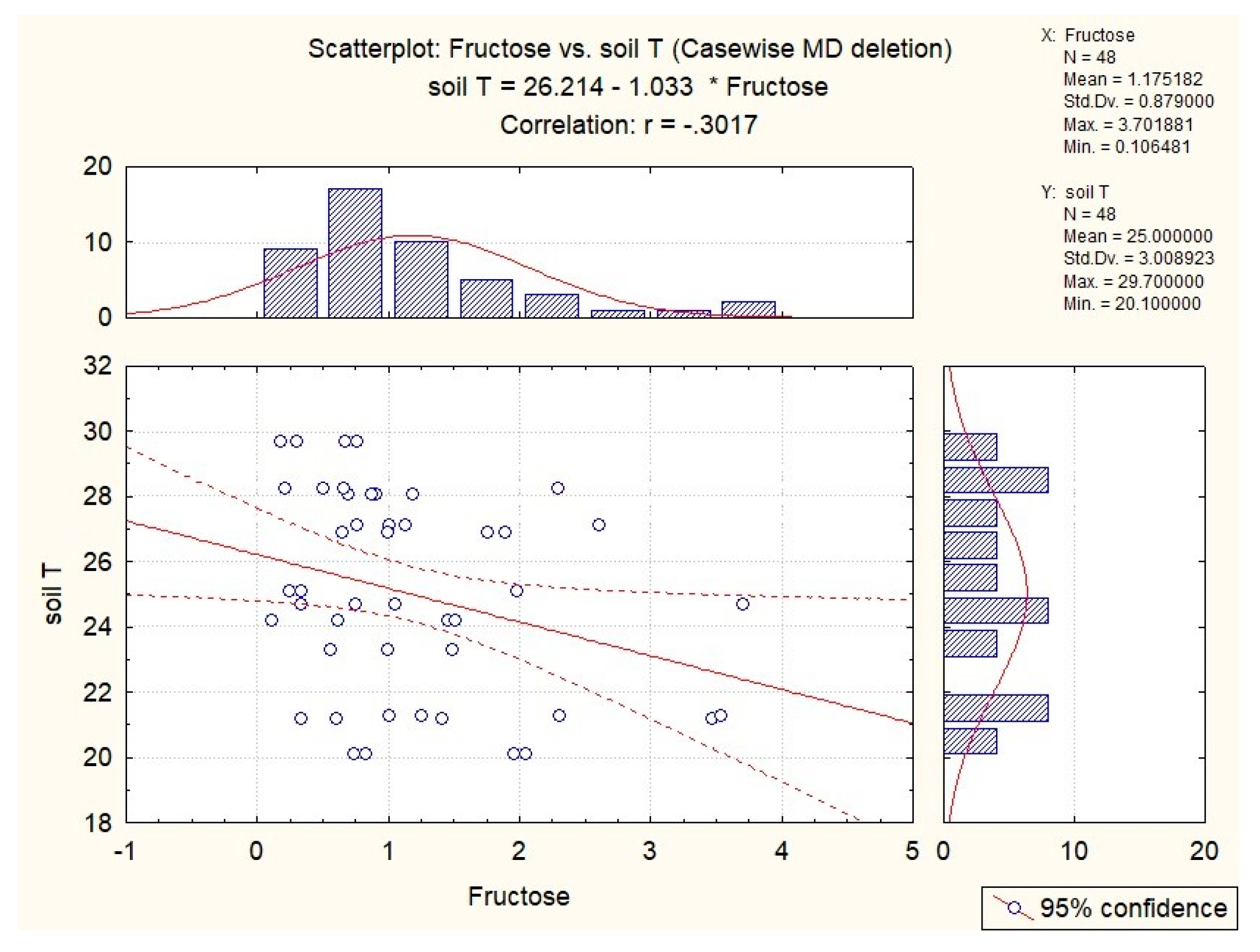
| fructose | x | x | x | x | x |
| glucose | x | x | x | x | x |
| UVB (µW/cm2) | Air T (°C) | Soil T (°C) | Air AH (g/m3) | Soil RH (%) | |
|---|---|---|---|---|---|
| sucrose | 0.36 | x | x | x | 0.53 |
| fructose | x | −0.36 | −0.30 | x | x |
| glucose | x | x | x | x | x |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malovrh, K.; Ravnjak, B.; Bavcon, J.; Križman, M. Nectar Production and Three Main Sugars in Nectar of Salvia pratensis and Salvia glutinosa in Correlation with Abiotic Factors. Agriculture 2024, 14, 668. https://doi.org/10.3390/agriculture14050668
Malovrh K, Ravnjak B, Bavcon J, Križman M. Nectar Production and Three Main Sugars in Nectar of Salvia pratensis and Salvia glutinosa in Correlation with Abiotic Factors. Agriculture. 2024; 14(5):668. https://doi.org/10.3390/agriculture14050668
Chicago/Turabian StyleMalovrh, Katja, Blanka Ravnjak, Jože Bavcon, and Mitja Križman. 2024. "Nectar Production and Three Main Sugars in Nectar of Salvia pratensis and Salvia glutinosa in Correlation with Abiotic Factors" Agriculture 14, no. 5: 668. https://doi.org/10.3390/agriculture14050668
APA StyleMalovrh, K., Ravnjak, B., Bavcon, J., & Križman, M. (2024). Nectar Production and Three Main Sugars in Nectar of Salvia pratensis and Salvia glutinosa in Correlation with Abiotic Factors. Agriculture, 14(5), 668. https://doi.org/10.3390/agriculture14050668