
Effect of Ultraviolet B Radiation on the Biosynthesis of Carotenoids in Foxtail Millet Grains

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. UV-B Radiation Treatment and Sample Collection
2.3. Color Determination and Extraction of Total Carotenoids from Foxtail Millet Grains
2.4. Analysis of Lutein and Zeaxanthin Using High-Performance Liquid Chromatography (HPLC)
2.5. Transcriptome Sequencing and Data Analysis
2.6. RNA Extraction and qRT-PCR Assay
2.7. Swiss-Prot, GO, and KEGG Pathway Annotations
2.8. Statistical Analysis
3. Results
3.1. Low-Intensity UV-B Lightened the Millet Color
3.2. Low Doses of UV-B Reduced the Carotenoid Content of Millet
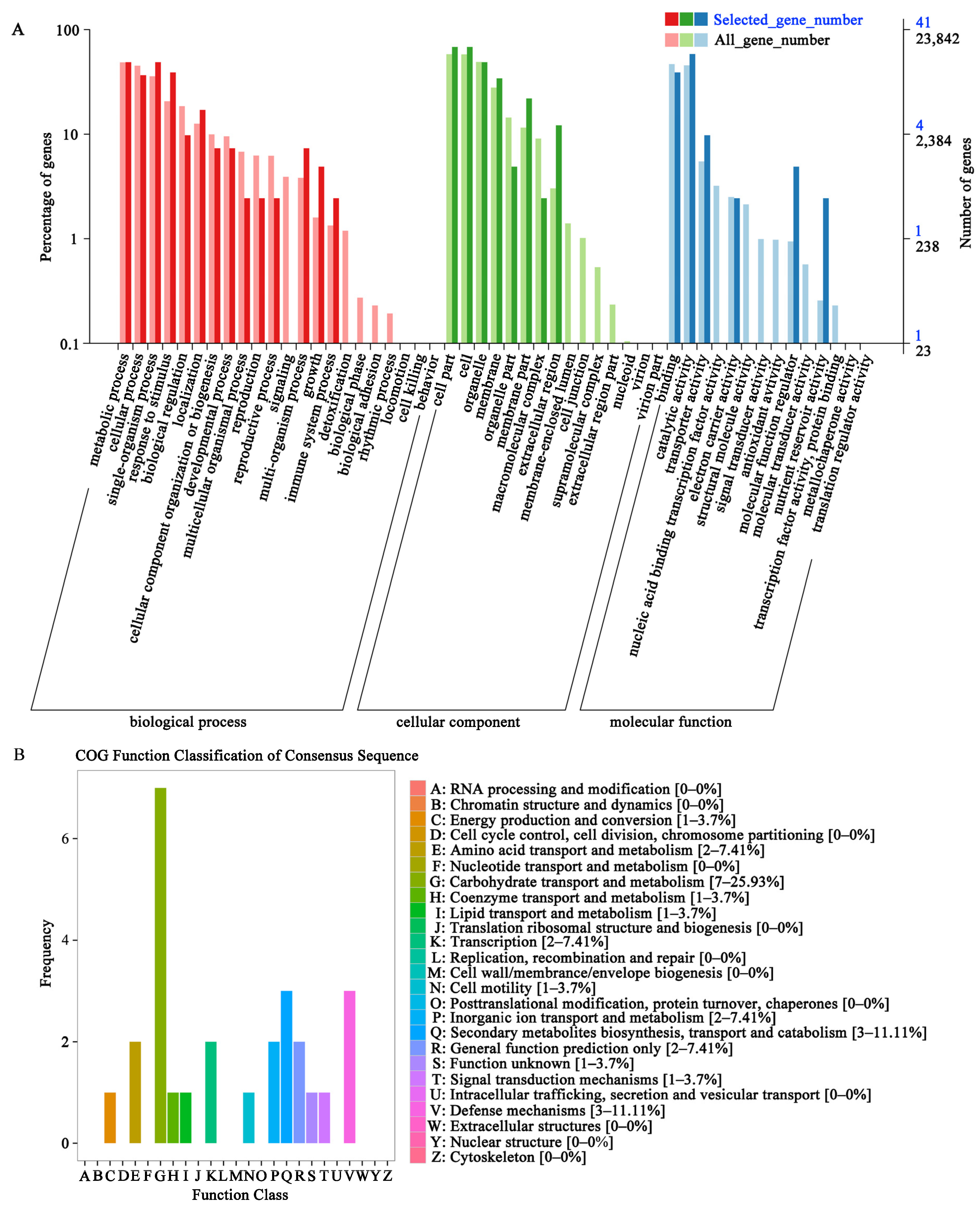
3.3. Transcriptome Analysis of Foxtail Millet Exposed to the Different UV-B Radiation Intensity
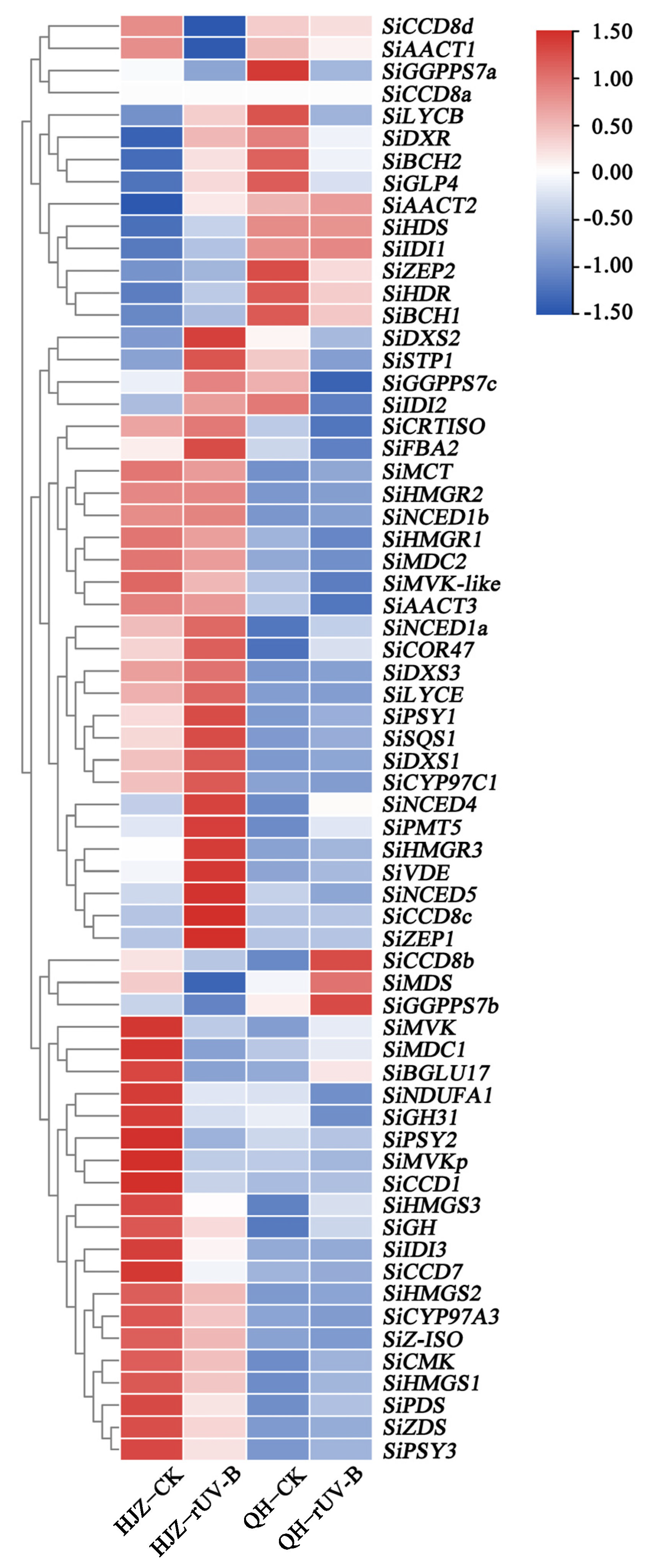
3.4. Expression Profiling of Carotenoid and Starch Genes Based on Transcriptome Data
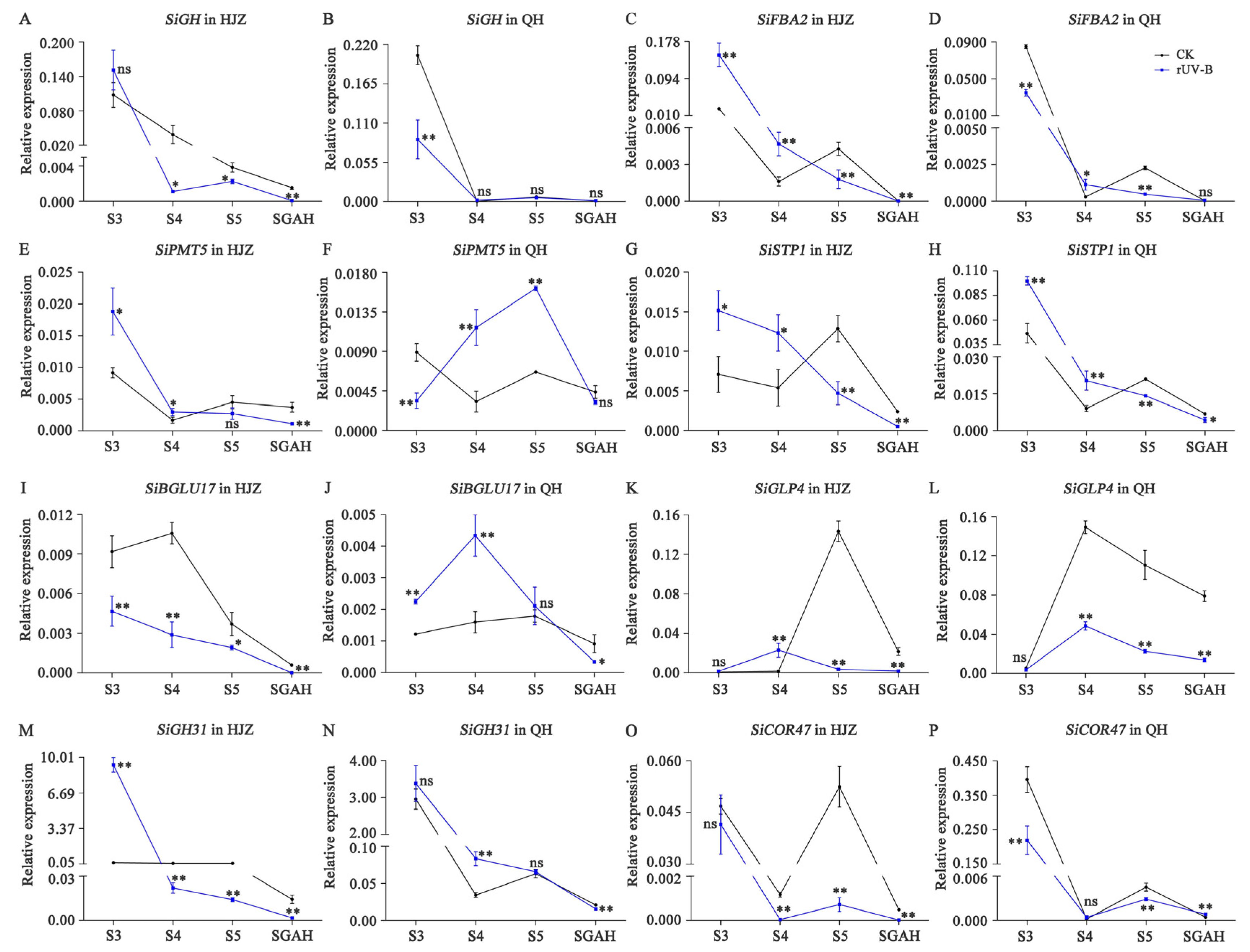
3.5. Expression Changes of Carotenoid and Starch Metabolism Genes at Four Stages of Grain Maturation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andrady, A.; Aucamp, P.J.; Bais, A.F.; Ballare, C.L.; Bjorn, L.O.; Bornman, J.F.; Caldwell, M.; Callaghan, T.; Cullen, A.P.; Erickson, D.J.; et al. Environmental effects of ozone depletion and its interactions with climate change: Progress report, 2004. Photochem. Photobiol. Sci. 2005, 4, 177–184. [Google Scholar]
- Shen, J.; Jiang, C.Q.; Yan, Y.F.; Liu, B.R.; Zu, C.L. Effect of increased UV-B radiation on carotenoid accumulation and total antioxidant capacity in tobacco (Nicotiana tabacum L.) leaves. Genet. Mol. Res. 2017, 16, gmr16018438. [Google Scholar] [CrossRef]
- Clayton, W.A.; Albert, N.W.; Thrimawithana, A.H.; McGhie, T.K.; Deroles, S.C.; Schwinn, K.E.; Warren, B.A.; McLachlan, A.R.G.; Bowman, J.L.; Jordan, B.R.; et al. UVR8-mediated induction of flavonoid biosynthesis for UVB tolerance is conserved between the liverwort Marchantia polymorpha and flowering plants. Plant J. 2018, 96, 503–517. [Google Scholar] [CrossRef]
- Stracke, R.; Favory, J.J.; Gruber, H.; Bartelniewoehner, L.; Bartels, S.; Binkert, M.; Funk, M.; Weisshaar, B.; Ulm, R. The Arabidopsis bZIP transcription factor HY5 regulates expression of the PFG1/MYB12 gene in response to light and ultraviolet-B radiation. Plant Cell Environ. 2010, 33, 88–103. [Google Scholar] [PubMed]
- Liang, T.; Shi, C.; Peng, Y.; Tan, H.; Xin, P.; Yang, Y.; Wang, F.; Li, X.; Chu, J.; Huang, J.; et al. Brassinosteroid-activated BRI1-EMS-SUPPRESSOR 1 inhibits flavonoid biosynthesis and coordinates growth and UV-B stress responses in plants. Plant Cell 2020, 32, 3224–3239. [Google Scholar] [CrossRef] [PubMed]
- Rius, S.P.; Emiliani, J.; Casati, P. P1 epigenetic regulation in leaves of high altitude maize landraces: Effect of UV-B radiation. Front. Plant Sci. 2016, 7, 523. [Google Scholar] [CrossRef] [PubMed]
- Rius, S.P.; Grotewold, E.; Casati, P. Analysis of the P1 promoter in response to UV-B radiation in allelic variants of high-altitude maize. BMC Plant Biol. 2012, 12, 92. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Wang, P.; Yang, R.; Gu, Z. Effects of UV-B radiation on the isoflavone accumulation and physiological-biochemical changes of soybean during germination: Physiological-biochemical change of germinated soybean induced by UV-B. Food Chem. 2018, 250, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Dillon, F.M.; Tejedor, M.D.; Ilina, N.; Chludil, H.D.; Mithofer, A.; Pagano, E.A.; Zavala, J.A. Solar UV-B radiation and ethylene play a key role in modulating effective defenses against Anticarsia gemmatalis larvae in field-grown soybean. Plant Cell Environ. 2018, 41, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Mao, B.; Wang, Y.; Zhao, T.H.; Tian, R.R.; Wang, W.; Ye, J.S. Combined effects of elevated O3 concentrations and enhanced UV-B radiation of the biometric and biochemical properties of soybean roots. Front Plant Sci. 2017, 8, 1568. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.H.; Wang, L.J.; Li, S.H.; Duan, W.; Loescher, W.; Liang, Z.C. The effects of UV-B radiation on photosynthesis in relation to Photosystem II photochemistry, thermal dissipation and antioxidant defenses in winter wheat (Triticum aestivum L.) seedlings at different growth temperatures. Funct. Plant Biol. 2007, 34, 907–917. [Google Scholar] [CrossRef]
- Chen, H.; Gong, Y.; Han, R. Cadmium telluride quantum dots (CdTe-QDs) and enhanced ultraviolet-B (UV-B) radiation trigger antioxidant enzyme metabolism and programmed cell death in wheat seedlings. PLoS ONE 2014, 9, e110400. [Google Scholar] [CrossRef]
- Faseela, P.; Puthur, J.T. Intraspecific variation in sensitivity of high yielding rice varieties towards UV-B radiation. Physiol. Mol. Biol. Pla. 2019, 25, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wu, S. Traditional maintenance and multiplication of foxtail millet (Setaria italica (L.) P. Beauv.) landraces in China. Euphytica 1996, 87, 33–38. [Google Scholar] [CrossRef]
- Aidoo, M.K.; Bdolach, E.; Fait, A.; Lazarovitch, N.; Rachmilevitch, S. Tolerance to high soil temperature in foxtail millet (Setaria italica L.) is related to shoot and root growth and metabolism. Plant Physiol. Biochem. 2016, 106, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Doust, A.N.; Kellogg, E.A.; Devos, K.M.; Bennetzen, J.L. Foxtail millet: A sequence-driven grass model system. Plant Physiol. 2009, 149, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Liu, J.; Cheng, L.; Zhang, Y.Y.; Hou, S.Y.; Sun, Z.X.; Li, H.Y.; Han, Y.H. Carotenoid composition and expression of biosynthetic genes in yellow and white foxtail millet [Setaria italica (L.) Beauv]. J. Cereal Sci. 2019, 85, 84–90. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, J.; Qie, Q.; Yang, Y.; Hou, S.; Wang, X.; Li, X.; Han, Y. Comparative analysis of flavonoid metabolites in foxtail millet (Setaria italica) with different eating quality. Life 2021, 11, 578. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Kuijer, H.N.J.; Al-Babili, S. Carotenoid biofortification of crops in the CRISPR era. Trends Biotechnol. 2020, 39, 857–860. [Google Scholar] [CrossRef] [PubMed]
- Jansen, M.; Hectors, K.; O’Brien, N.M.; Guisez, Y.; Potters, G. Plant stress and human health: Do human consumers benefit from UV-B acclimated crops. Plant Sci. 2008, 175, 449–458. [Google Scholar] [CrossRef]
- Middleton, E.M.; Teramura, A.H. The role of flavonol glycosides and carotenoids in protecting soybean from ultraviolet-B damage. Plant Physiol. 1993, 103, 741–752. [Google Scholar] [CrossRef]
- Li, L.; Yuan, H.; Zeng, Y.; Xu, Q. Plastids and carotenoid accumulation. Sub-Cell. Biochem. 2016, 79, 273–293. [Google Scholar]
- Sun, T.; Yuan, H.; Cao, H.; Yazdani, M.; Tadmor, Y.; Li, L. Carotenoid metabolism in plants: The role of plastids. Mol. Plant. 2018, 11, 58–74. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Rao, S.; Zhou, X.; Li, L. Plant carotenoids: Recent advances and future perspectives. Mol. Hortic. 2022, 2, 3. [Google Scholar] [CrossRef]
- Vranova, E.; Coman, D.; Gruissem, W. Network analysis of the MVA and MEP pathways for isoprenoid synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef]
- Cazzonelli, C.I.; Pogson, B.J. Source to sink: Regulation of carotenoid biosynthesis in plants. Trends Plant Sci. 2010, 15, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid metabolism in plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef]
- Sandmann, G. Diversity and origin of carotenoid biosynthesis: Its history of coevolution towards plant photosynthesis. New Phytol. 2021, 232, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Harjes, C.E.; Rocheford, T.R.; Bai, L.; Brutnell, T.P.; Kandianis, C.B.; Sowinski, S.G.; Stapleton, A.E.; Vallabhaneni, R.; Williams, M.; Wurtzel, E.T.; et al. Natural genetic variation in lycopene epsilon cyclase tapped for maize biofortification. Science 2008, 319, 330–333. [Google Scholar] [CrossRef]
- Ahrazem, O.; Gomez-Gomez, L.; Rodrigo, M.J.; Avalos, J.; Limon, M.C. Carotenoid cleavage oxygenases from microbes and photosynthetic organisms: Features and functions. Int. J. Mol. Sci. 2016, 17, 1781. [Google Scholar] [CrossRef]
- Beltran, J.C.; Stange, C. Apocarotenoids: A new carotenoid-derived pathway. Sub-Cell. Biochem. 2016, 79, 239–272. [Google Scholar]
- Dhar, M.K.; Mishra, S.; Bhat, A.; Chib, S.; Kaul, S. Plant carotenoid cleavage oxygenases: Structure-function relationships and role in development and metabolism. Brief. Funct. Genom. 2020, 19, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.H.; Zhu, J.; Jiang, J.G. Carotenoids biosynthesis and cleavage related genes from bacteria to plants. Crit. Rev. Food Sci. 2018, 58, 2314–2333. [Google Scholar] [CrossRef] [PubMed]
- Ablazov, A.; Mi, J.; Jamil, M.; Jia, K.P.; Wang, J.Y.; Feng, Q.; Al-Babili, S. The apocarotenoid zaxinone is a positive regulator of strigolactone and abscisic acid biosynthesis in arabidopsis roots. Front. Plant Sci. 2020, 11, 578. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Haider, I.; Jamil, M.; Fiorilli, V.; Saito, Y.; Mi, J.; Baz, L.; Kountche, B.A.; Jia, K.P.; Guo, X.; et al. The apocarotenoid metabolite zaxinone regulates growth and strigolactone biosynthesis in rice. Nat. Commun. 2019, 10, 810. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Pan, X.; Wang, R.; Xu, J.; Guo, J.; Yang, T.; Zhao, J.; Nadeem, F.; Liu, X.; Shan, H.; et al. ZmCCD10a encodes a distinct type of carotenoid cleavage dioxygenase and enhances plant tolerance to low phosphate. Plant Physiol. 2020, 184, 374–392. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Aalel, S.M.; Young, J.C.; Rabalski, I.; Hucl, P.; Fregeau-Reid, J. Identification and quantification of seed carotenoids in selected wheat species. J. Agric. Food Chem. 2007, 55, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Shen, R.; Yang, S.; Zhao, G.; Shen, Q.; Diao, X. Identification of carotenoids in foxtail millet (Setaria italica) and the effects of cooking methods on carotenoid content. J. Cereal Sci. 2015, 61, 86–93. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef] [PubMed]
- Becatti, E.; Petroni, K.; Giuntini, D.; Castagna, A.; Calvenzani, V.; Serra, G.; Mensuali-Sodi, A.; Tonelli, C.; Ranieri, A. Solar UV-B radiation influences carotenoid accumulation of tomato fruit through both ethylene-dependent and -independent mechanisms. J. Agric. Food Chem. 2009, 57, 10979–10989. [Google Scholar] [CrossRef] [PubMed]
- Mackerness, S.A.H. Plant responses to ultraviolet-B (UV-B: 280–320 nm) stress: What are the key regulators? Plant Growth Regul. 2000, 32, 27–39. [Google Scholar] [CrossRef]
- Badmus, U.O.; Crestani, G.; Cunningham, N.; Havaux, M.; Urban, O.; Jansen, M.A.K. UV radiation induces specific changes in the carotenoid profile of Arabidopsis thaliana. Biomolecules 2022, 12, 1879. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.S.; Wang, L.L.; Zhou, X.R.; Shuang, S.M.; Zhu, Z.H.; Li, N.; Li, Y.; Liu, F.; Liu, S.C.; Lu, P.; et al. Determination of protein, fat, starch, and amino acids in foxtail millet [Setaria italica (L.) Beauv.] by Fourier transform near-infrared reflectance spectroscopy. Food Sci. Biotechnol. 2013, 22, 1495–1500. [Google Scholar] [CrossRef]
- Zhang, L.Z.; Liu, R.H. Phenolic and carotenoid profiles and antiproliferative activity of foxtail millet. Food Chem. 2015, 174, 495–501. [Google Scholar] [CrossRef]
- Li, X.; He, Y.; Xie, C.; Zu, Y.; Zhan, F.; Mei, X.; Xia, Y.; Li, Y. Effects of UV-B radiation on the infectivity of Magnaporthe oryzae and rice disease-resistant physiology in Yuanyang terraces. Photochem. Photobiol. Sci. 2018, 17, 8–17. [Google Scholar] [CrossRef]
- Teramura, A.H.; Sullivan, J.H. Effects of UV-B radiation on photosynthesis and growth of terrestrial plants. Photosynth. Res. 1994, 39, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, D.J.; Tadeo, F.R.; Legaz, F.; Primo-Millo, E.; Talon, M. In vivo sucrose stimulation of colour change in citrus fruit epicarps: Interactions between nutritional and hormonal signals. Physiol. Plant. 2001, 112, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.H.; Zhou, X.S.; Rao, S.; Liu, J.P.; Li, L. Chapter Thirteen—Protein–protein interaction techniques to investigate post-translational regulation of carotenogenesis. Method Enzymol. 2022, 671, 301–325. [Google Scholar]
- D’Andrea, L.; Simon-Moya, M.; Llorente, B.; Llamas, E.; Marro, M.; Loza-Alvarez, P.; Li, L.; Rodriguez-Concepcion, M. Interference with Clp protease impairs carotenoid accumulation during tomato fruit ripening. J. Exp. Bot. 2018, 69, 1557–1568. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Developmental Stages | Treatment | L* | a* | b* |
|---|---|---|---|---|
| S3 | HJZ-CK | 61.45 ± 1.372 | 5.20 ± 0.265 | 40.49 ± 0.268 |
| HJZ-rUV-B | 61.06 ± 0.996 | 3.72 ± 0.109 ** | 35.95 ± 0.353 ** | |
| QH-CK | 65.31 ± 0.265 | 4.26 ± 0.433 | 35.68 ± 0.194 | |
| QH-rUV-B | 62.34 ± 0.989 * | 3.44 ± 0.147 | 34.48 ± 1.062 | |
| S4 | HJZ-CK | 55.56 ± 1.235 | 8.12 ± 0.100 | 43.39 ± 0.908 |
| HJZ-rUV-B | 54.22 ± 1.615 | 11.34 ± 0.791 * | 37.88 ± 0.869 * | |
| QH-CK | 55.26 ± 0.472 | 9.55 ± 0.296 | 41.69 ± 0.118 | |
| QH-rUV-B | 61.74 ± 0.160 ** | 5.80 ± 0.468 ** | 37.18 ± 0.159 ** | |
| S5 | HJZ-CK | 53.71 ± 0.403 | 10.67 ± 0.245 | 42.98 ± 0.369 |
| HJZ-rUV-B | 55.39 ± 0.488 | 10.70 ± 0.044 | 42.28 ± 0.103 | |
| QH-CK | 56.55 ± 0.607 | 9.04 ± 0.204 | 40.24 ± 0.294 | |
| QH-rUV-B | 55.66 ± 1.531 | 8.96 ± 0.511 | 38.78 ± 0.303 * | |
| SGAH | HJZ-CK | 57.01 ± 0.322 | 9.89 ± 0.053 | 44.99 ± 0.215 |
| HJZ-rUV-B | 54.96 ± 0.240 | 9.32 ± 0.019 | 43.29 ± 0.115 | |
| QH-CK | 53.69 ± 1.010 | 9.30 ± 0.280 | 41.97 ± 0.322 | |
| QH-rUV-B | 49.99 ± 0.392 * | 9.58 ± 0.234 | 40.40 ± 0.286 * |
| Developmental Stages | Treatment | Content of Total Carotenoid (TCC, mg/kg DW) | Lutein Content (LC, mg/kg DW) | Zeaxanthin Content (ZC, mg/kg DW) | (LC + ZC)/TCC(%) |
|---|---|---|---|---|---|
| S3 | HJZ-CK | 27.87 ± 3.239 | 5.49 ± 1.012 | 2.75 ± 0.277 | 29.54% |
| HJZ-rUV-B | 21.75 ± 0.874 | 4.52 ± 0.546 * | 3.61 ± 1.243 | 37.39% | |
| QH-CK | 13.14 ± 0.335 | 3.50 ± 0.783 | 2.63 ± 0.079 | 46.70% | |
| QH-rUV-B | 12.58 ± 0.857 | 2.32 ± 0.068 * | 1.22 ± 0.073 ** | 28.10% | |
| S4 | HJZ-CK | 19.29 ± 0.574 | 5.09 ± 0.312 | 2.70 ± 0.133 | 40.37% |
| HJZ-rUV-B | 19.00 ± 1.953 | 4.86 ± 0.624 | 2.53 ± 0.450 | 38.90% | |
| QH-CK | 13.95 ± 1.077 | 5.55 ± 0.219 | 3.16 ± 0.338 | 62.40% | |
| QH-rUV-B | 12.86 ± 1.279 | 4.09 ± 0.199 ** | 2.27 ± 0.079 * | 49.44% | |
| S5 | HJZ-CK | 17.63 ± 1.056 | 5.64 ± 0.197 | 3.18 ± 0.223 | 50.04% |
| HJZ-rUV-B | 16.94 ± 0.358 | 5.54 ± 0.543 | 2.95 ± 0.381 | 50.06% | |
| QH-CK | 14.04 ± 1.440 | 4.57 ± 0.264 | 2.74 ± 0.242 | 52.11% | |
| QH-rUV-B | 13.95 ± 2.148 | 3.48 ± 2.254 * | 2.89 ± 0.075 | 45.66% | |
| SGAH | HJZ-CK | 12.66 ± 0.490 | 7.84 ± 1.057 | 4.36 ± 0.644 | 96.36% |
| HJZ-rUV-B | 11.99 ± 0.469 * | 6.83 ± 0.889 | 3.73 ± 0.685 | 88.04% | |
| QH-CK | 10.03 ± 0.449 | 6.12 ± 0.578 | 3.42 ± 0.433 | 95.12% | |
| QH-rUV-B | 8.77 ± 0.527 | 4.92 ± 0.954 | 3.53 ± 0.762 | 96.44% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huo, Y.; Pang, X.; Feng, H.; Hao, R.; Zhang, R.; Wang, Y.; Wang, H.; Zhang, B.; Han, Y. Effect of Ultraviolet B Radiation on the Biosynthesis of Carotenoids in Foxtail Millet Grains. Agriculture 2024, 14, 289. https://doi.org/10.3390/agriculture14020289
Huo Y, Pang X, Feng H, Hao R, Zhang R, Wang Y, Wang H, Zhang B, Han Y. Effect of Ultraviolet B Radiation on the Biosynthesis of Carotenoids in Foxtail Millet Grains. Agriculture. 2024; 14(2):289. https://doi.org/10.3390/agriculture14020289
Chicago/Turabian StyleHuo, Yiqiong, Xiangdong Pang, Huixia Feng, Ruili Hao, Ruijie Zhang, Yushen Wang, Haigang Wang, Bin Zhang, and Yuanhuai Han. 2024. "Effect of Ultraviolet B Radiation on the Biosynthesis of Carotenoids in Foxtail Millet Grains" Agriculture 14, no. 2: 289. https://doi.org/10.3390/agriculture14020289
APA StyleHuo, Y., Pang, X., Feng, H., Hao, R., Zhang, R., Wang, Y., Wang, H., Zhang, B., & Han, Y. (2024). Effect of Ultraviolet B Radiation on the Biosynthesis of Carotenoids in Foxtail Millet Grains. Agriculture, 14(2), 289. https://doi.org/10.3390/agriculture14020289