
Identifying Plant Functional Traits of Weeds in Fields Planted with Glyphosate-Tolerant Maize for Preferable Weed Management Practices


Abstract
1. Introduction
2. Results
2.1. Indicator Weed Species
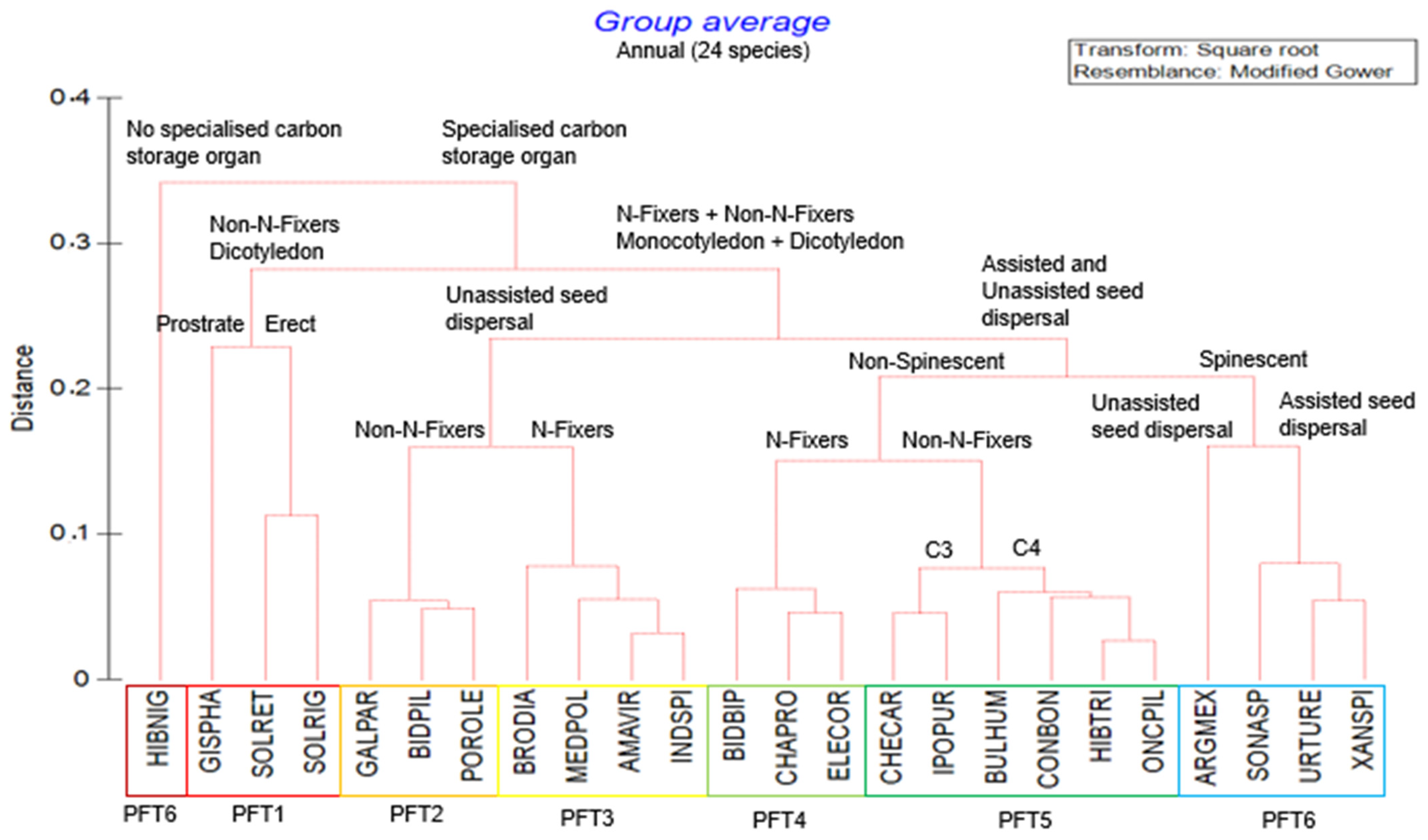
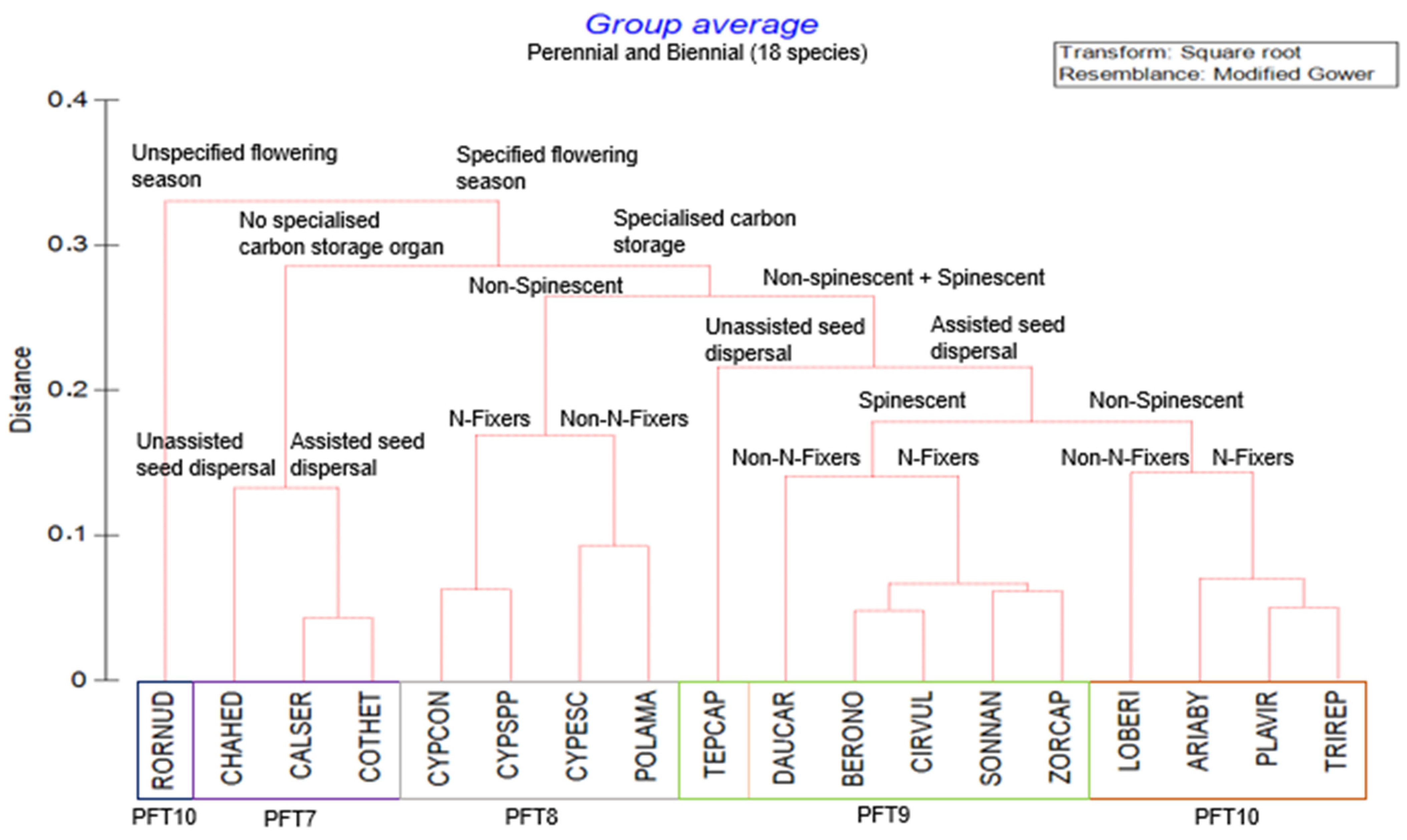
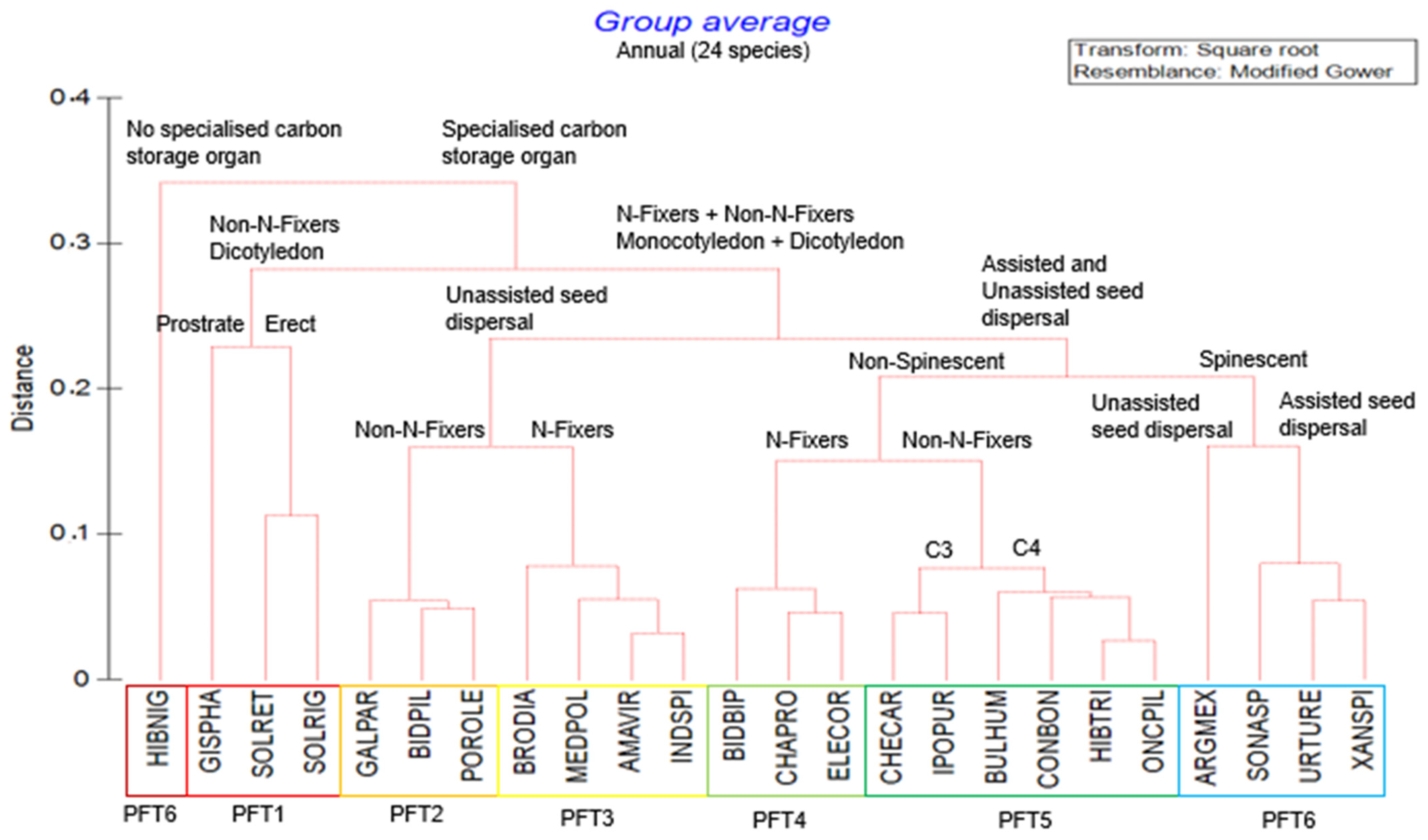
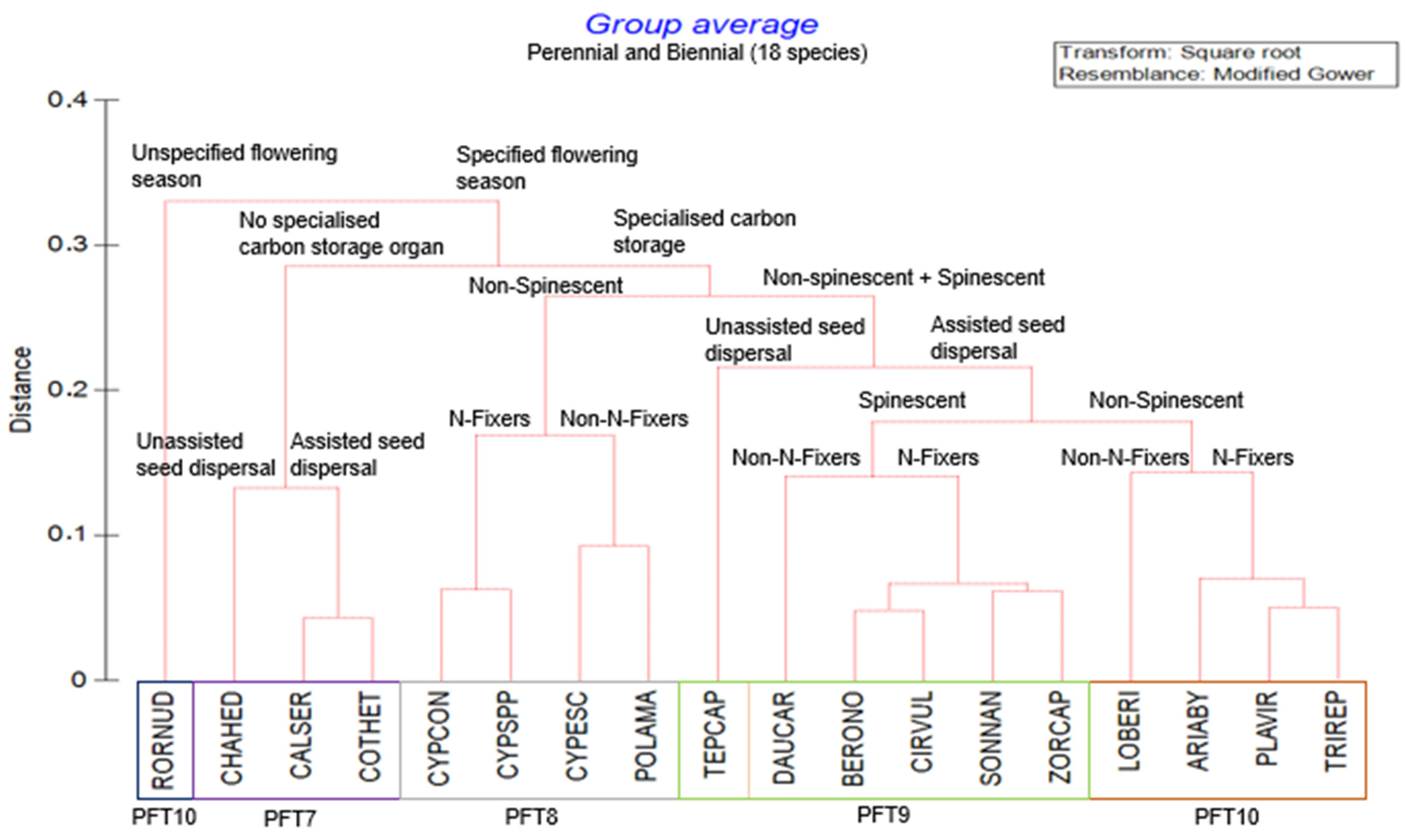
2.2. Plant Functional Types
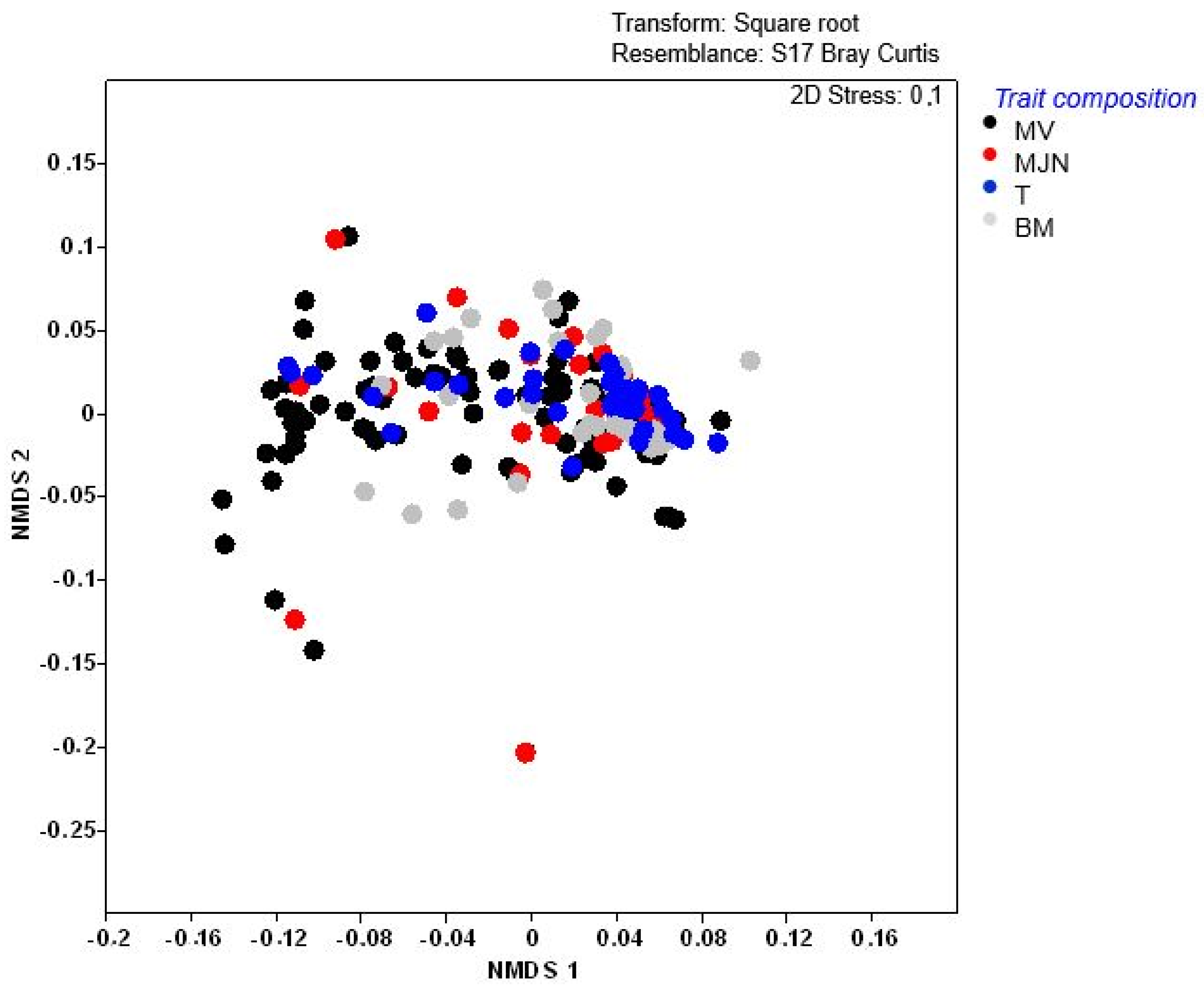
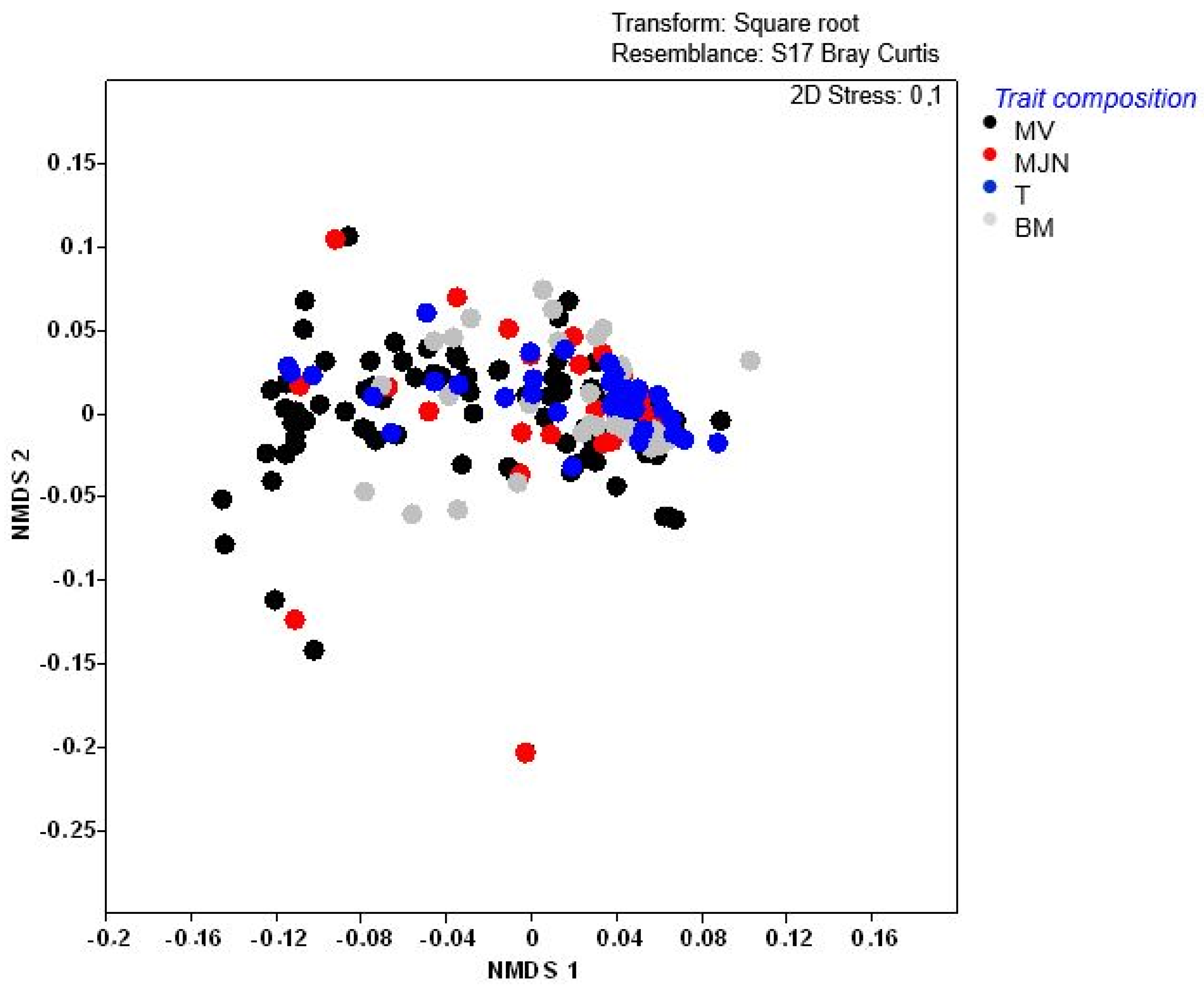
2.3. Weed Trait Assemblages
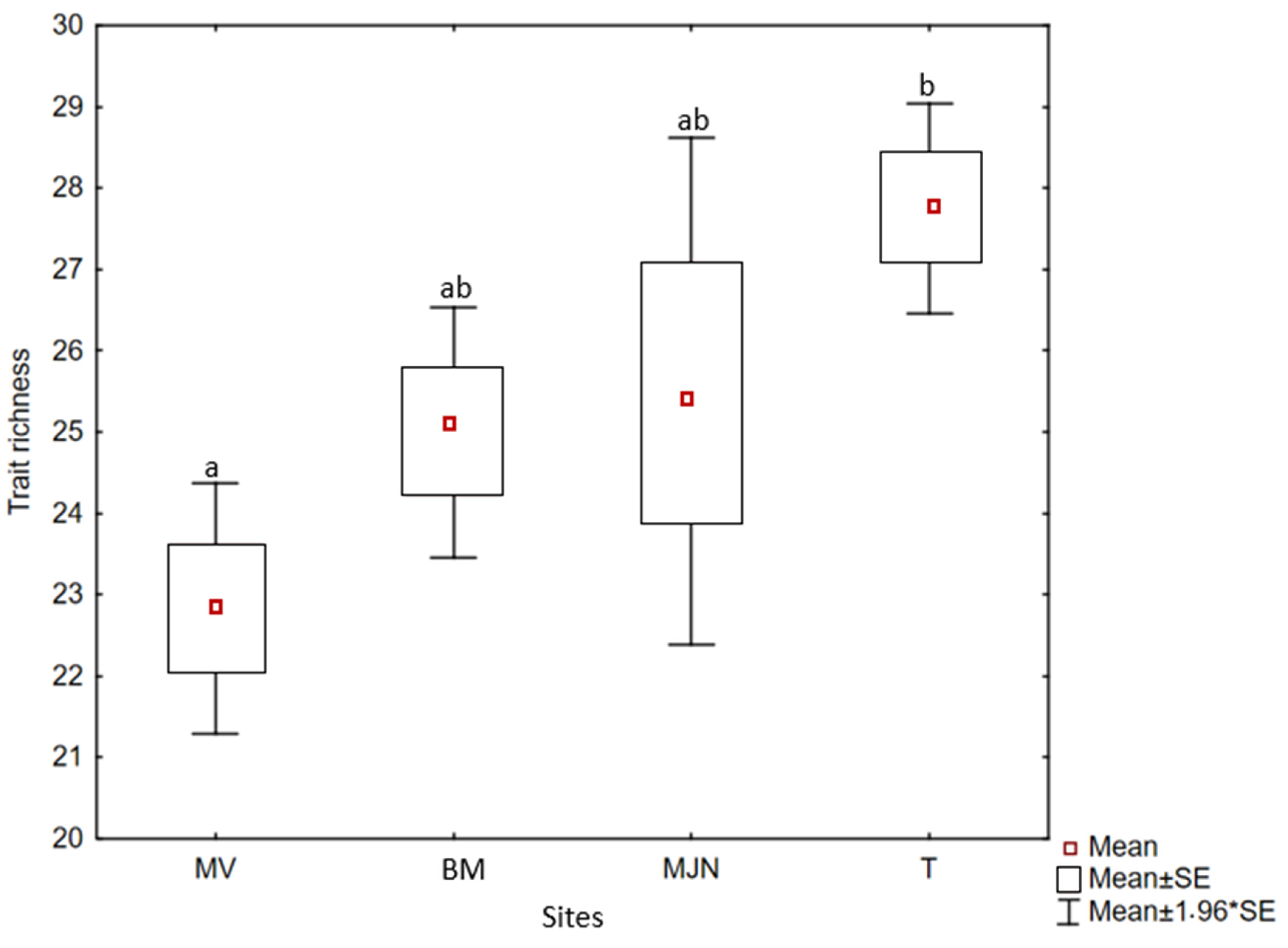
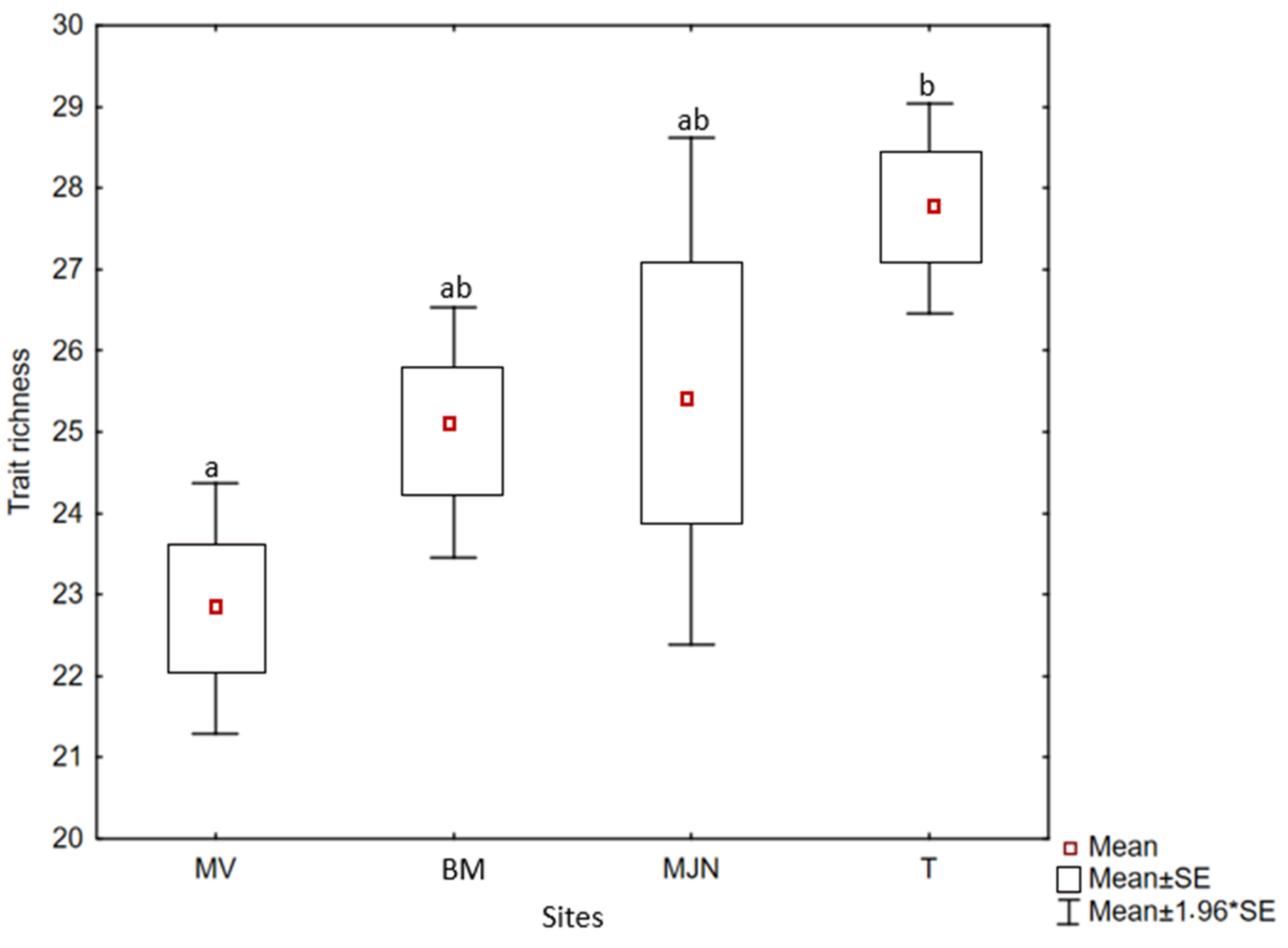
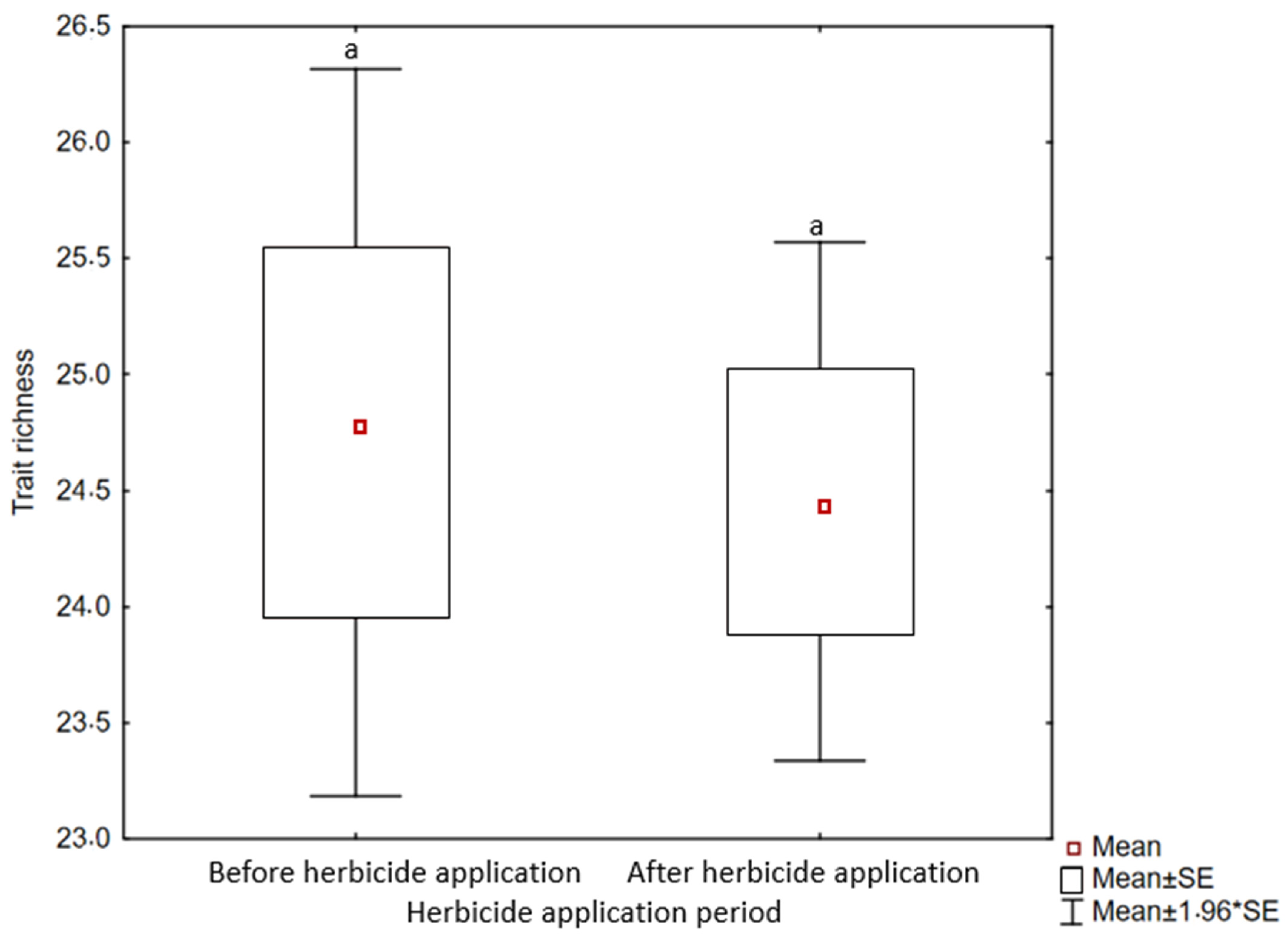
2.4. Weed Trait Richness
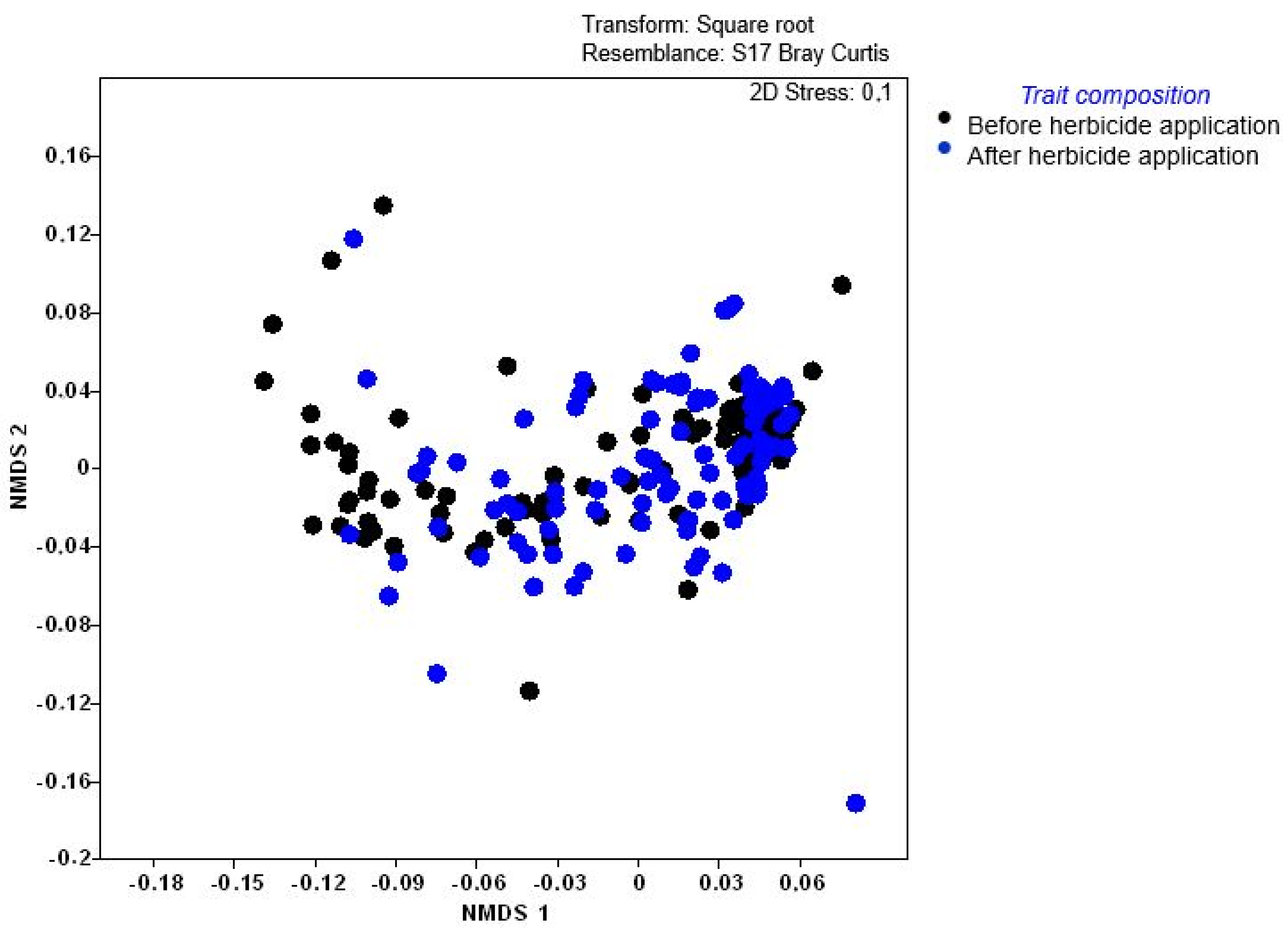
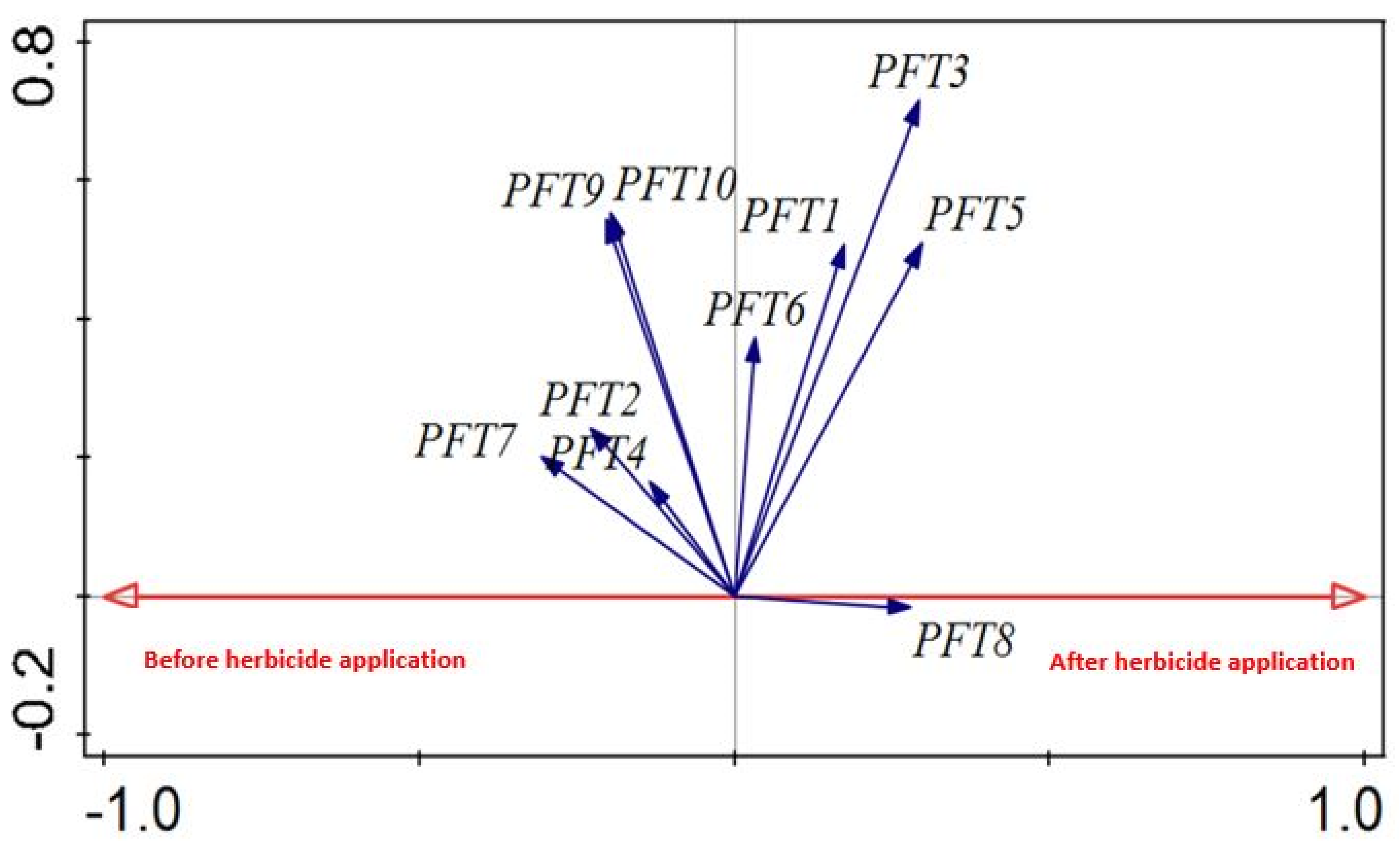
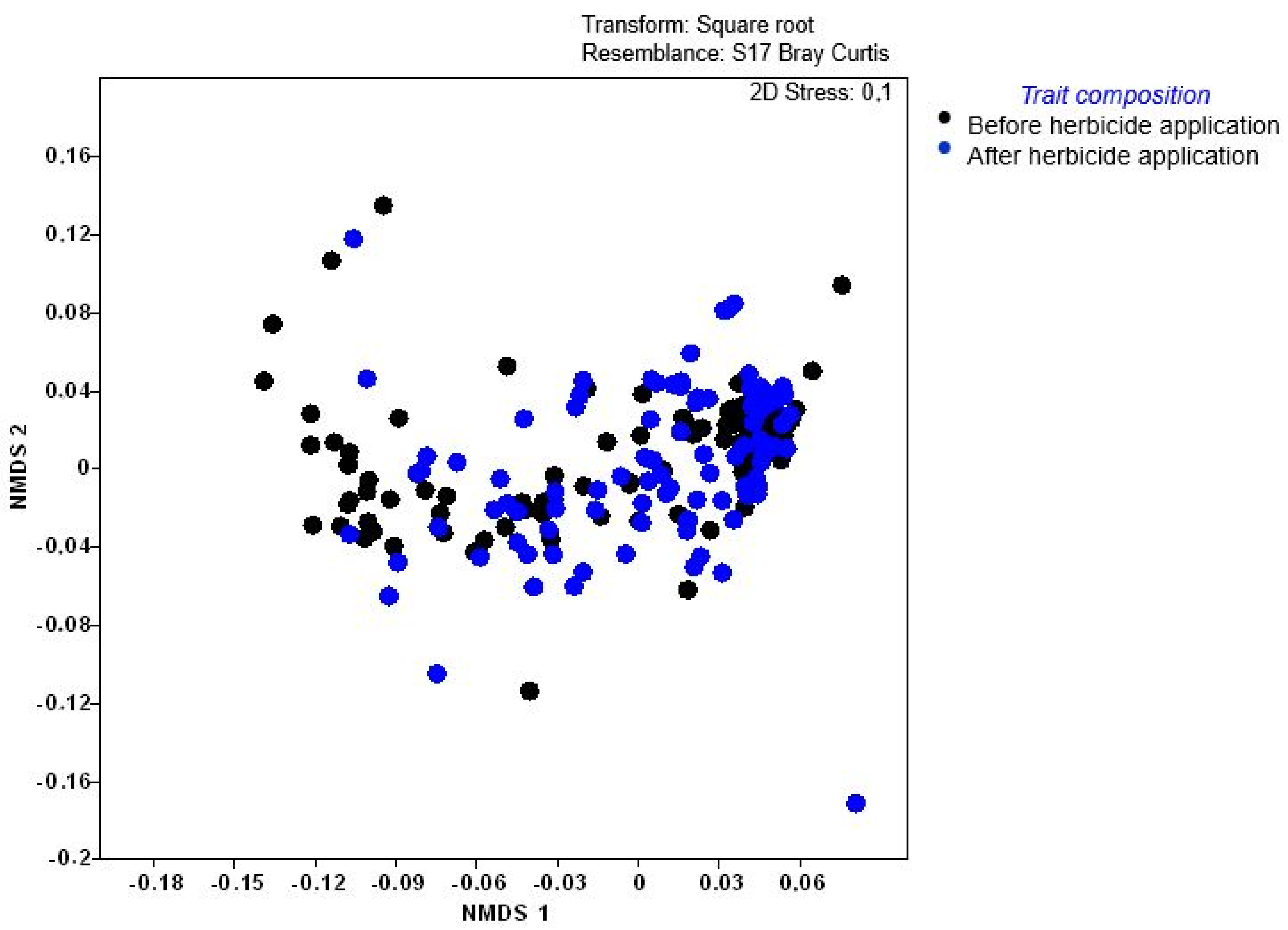
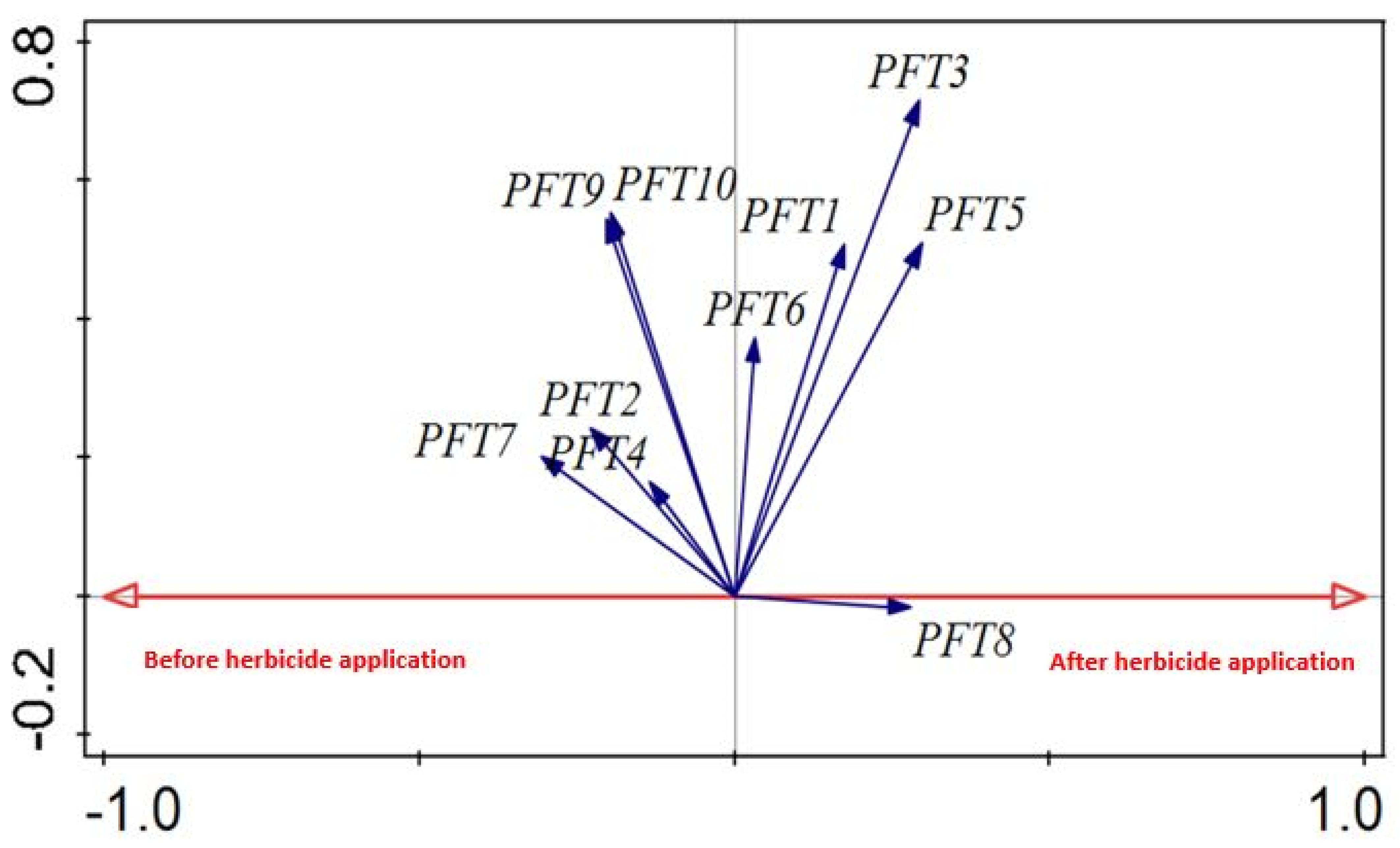
2.5. Weed Trait Responses to Disturbance
3. Discussion
3.1. Weed Traits Composition and Richness
3.2. Plant Functional Types
3.3. Herbicide-Resistant Weeds
4. Materials and Methods
4.1. Generated Plant List of Weed Species
4.2. Functional Trait Identification and Adaptation for Herbicide Disturbance Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baker, H.G. The evolution of weeds. Annu. Rev. Ecol. Syst. 1974, 5, 1–24. [Google Scholar] [CrossRef]
- Głowacka, A. Dominant weeds in maize (Zea mays L.) cultivation and their competitiveness under conditions of various methods of weed control. Acta Agrobot. 2012, 64, 119–126. [Google Scholar] [CrossRef]
- Bazzaz, F.A. Life History of Colonizing Plants: Some Demographic, Genetic, and Physiological Features. In Ecology of Biological Invasions of North America and Hawaii; Ecological Studies Series; Mooney, H.A., Drake, J.A., Eds.; Springer: New York, NY, USA, 1986; Volume 58, pp. 96–110. [Google Scholar] [CrossRef]
- Vila-Aiub, M.M. Fitness of herbicide-resistant weeds: Current knowledge and implications for management. Plants 2019, 8, 469. [Google Scholar] [CrossRef]
- Lavorel, S.; McIntyre, S.; Landsberg, J.; Forbes, T. Plant functional classifications: From general groups to specific groups based on response to disturbance. Trends Ecol. Evol. 1997, 12, 474–478. [Google Scholar] [CrossRef]
- Lavorel, S.; Garnier, E. Predicting changes in community composition and ecosystem functioning from plant traits: Revisiting the Holy Grail. Funct. Ecol. 2002, 16, 545–556. [Google Scholar] [CrossRef]
- Díaz, S.; Lavorel, S.; McIntyre, S.; Falczuk, V.; Casanoves, F.; Milchunas, D.G.; Skarpe, C.; Rusch, G.; Sternberg, M.; Noy-Meir, I.; et al. Plant trait responses to grazing—A global synthesis. Glob. Chang. Biol. 2007, 13, 313–341. [Google Scholar] [CrossRef]
- Linstädter, A.; Schellberg, J.; Brüser, K.; Moreno García, C.A.; Oomen, R.J.; Du Preez, C.C.; Ruppert, J.C.; Ewert, F. Are there consistent grazing indicators in drylands? Testing plant functional types of various complexity in South Africa’s grassland and savanna biomes. PLoS ONE 2014, 9, e104672. [Google Scholar] [CrossRef] [PubMed]
- Kimball, S.; Funk, J.L.; Spasojevic, M.J.; Suding, K.N.; Parker, S.; Goulden, M.L. Can functional traits predict plant community response to global change? Ecosphere 2016, 7, e01602. [Google Scholar] [CrossRef]
- Martin, A.; Isaac, M. Plant functional traits in agroecosystems: A blueprint for research. J. Appl. Ecol. 2015, 52, 1425–1435. [Google Scholar] [CrossRef]
- Keddy, P.A. Assembly and response rules: Two goals for predictive community ecology. Int. J. Veg. Sci. 1992, 3, 157–164. [Google Scholar] [CrossRef]
- Kraft, N.J.B.; Valencia, R.; Ackerly, D.D. Functional traits and niche-based tree community assembly in an Amazonian forest. J. Sci. 2008, 322, 580–582. [Google Scholar] [CrossRef] [PubMed]
- Poorter, H.; Niinemets, Ü.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. N. Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef] [PubMed]
- Grime, J.P. Vegetation classification by reference to strategy. Nature 1974, 250, 26–31. [Google Scholar] [CrossRef]
- Muller, M.; Siebert, F.; Ntloko, B.R.; Siebert, S.J. A floristic assessment of grassland diversity loss in South Africa. Bothalia 2021, 51, 1–9. [Google Scholar] [CrossRef]
- Marshall, E.J.P.; Moonen, A.C. Field margins in northern Europe: Their functions and interactions with agriculture. Agric. Ecosyst. Environ. 2002, 89, 5–21. [Google Scholar] [CrossRef]
- Wessels, K.J.; Reyers, B.; van Jaarsveld, A.S.; Rutherford, M.C. Identification of potential conflict areas between land transformation and biodiversity conservation in north-eastern South Africa. Agric. Ecosyst. Environ. 2003, 95, 157–178. [Google Scholar] [CrossRef]
- Dale, V.; Polasky, S. Measures of the effects of agricultural practices on ecosystem services. Ecol. Econ. 2007, 64, 286–296. [Google Scholar] [CrossRef]
- Abate, T.; van Huis, A.; Ampofo, J.K.O. Pest management strategies in traditional agriculture: An African perspective. Annu. Rev. Entomol. 2000, 45, 631–659. [Google Scholar] [CrossRef]
- Banik, D.; Jha, M.K. Chapter 8: ‘Weed Management’. In Research Trends in Crop and Weed; Nand, V., Ed.; ND University of Agriculture and Technology: Kumarganj, India, 2020; p. 131. [Google Scholar]
- Tudi, M.; Daniel Ruan, H.; Wang, L.; Lyu, J.; Sadler, R.; Connell, D.; Chu, C.; Phung, D.T. Agriculture development, pesticide application and its impact on the environment. Int. J. Environ. Res. Public Health 2021, 18, 1112. [Google Scholar] [CrossRef]
- Nitin, K.S.; Masehela, T.S.; Chakravarthy, A.K.; Geerts, S. Management of pests using genetic tools in Africa. In Genetic Methods and Tools for Managing Crop Pests; Springer: Berlin/Heidelberg, Germany, 2022; pp. 303–326. [Google Scholar] [CrossRef]
- ISAAA. Global Status of Commercialized Biotech/GM Crops in 2019: Biotech Crops Drive Socio-Economic Development and Sustainable Environment in the New Frontier; ISAAA Brief No. 55; ISAAA: Ithaca, NY, USA, 2019. [Google Scholar]
- Morton, J.F. The impact of climate change on smallholder and subsistence agriculture. Proc. Natl. Acad. Sci. USA 2007, 104, 19680–19685. [Google Scholar] [CrossRef]
- Ortiz-Bobea, A.; Tack, J. Is another genetic revolution needed to offset climate change impacts for US maize yields? Environ. Res. Lett. 2018, 13, 124009. [Google Scholar] [CrossRef]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [PubMed]
- BFAP. Baseline Agricultural Outlook. Available online: https://www.sagis.org.za/BFAPBaseline-2019.pdf (accessed on 11 April 2020).
- Masehela, T.S.; Rhodes, J.I.; Groenewald, H.; Poole, C.J.; Van den Berg, J.; Gouse, M.; Skowno, A.L.; Barros, E.; Seymour, C.L.; Mandivenyi, W.G.; et al. An Initial Assessment of Impacts on Biodiversity from GMOs Released into the Environment in South Africa; South African National Biodiversity Institute: Pretoria, South Africa, 2021. [Google Scholar]
- Ofosu, R.; Agyemang, E.; Marton, A.; Pasztor, G.; Taller, J.; Kazinczi, G. Herbicide resistance: Managing weeds in a changing world. Agronomy 2023, 13, 1595. [Google Scholar] [CrossRef]
- Botha, M.; Siebert, S.J.; Van den Berg, J.; Ellis, S.; Dreber, N. Plant functional types differ between the grassland and savanna biomes along an agro-ecosystem disturbance gradient in South Africa. S. Afr. J. Bot. 2017, 113, 308–317. [Google Scholar] [CrossRef]
- Fried, G.; Chauvel, B.; Munoz, F.; Reboud, X. Which traits make weeds more successful in maize crops? Insights from a three-decade monitoring in France. Plants 2019, 9, 40. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchman, N.; Gurvich, D.E.; Reich, P.B.; Ter Steege, H.; Morgan, H.D.; Van der Heijden, M.G.A.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef]
- Martínková, J.; Klimeš, A.; Puy, J.; Klimešová, J. Response of clonal versus non clonal herbs to disturbance: Different strategies revealed. Perspect. Plant Ecol. Evol. Syst. 2020, 44, 125529. [Google Scholar] [CrossRef]
- Crayn, D.M.; Smith, J.A.C.; Winter, K. Carbon-isotope ratios and photosynthetic pathways in the neotropical family Rapateaceae. Plant Biol. 2001, 3, 569–576. [Google Scholar] [CrossRef]
- Díaz, S.; Cabido, M. Vive la difference: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Liira, J.; Schmidt, T.; Aavik, T.; Arens, P.; Augenstein, I.; Bailey, D.; Billeter, R.; Bukáče, R.; Burel, F.; Blust, G. Plant functional group composition and large-scale species richness in European agricultural landscapes. J. Veg. Sci. 2008, 19, 3–14. [Google Scholar] [CrossRef]
- Wratten, S.; Gillespie, M.; Decourtye, A.; Mader, E.; Desneux, N. Pollinator habitat enhancement: Benefits to other ecosystem services. Agric. Ecosyst. Environ. 2012, 159, 112–122. [Google Scholar] [CrossRef]
- Nasim, G.; Shabbir, A. Invasive weed species—A threat to sustainable agriculture. In Crop Production for Agricultural Improvement; Springer: Berlin/Heidelberg, Germany, 2012; pp. 523–556. [Google Scholar] [CrossRef]
- Shabani, F.; Ahmadi, M.; Kumar, L.; Solhjouy-fard, S.; Tehrany, M.S.; Shabani, F.; Kalantar, B.; Esmaeili, A. Invasive weed species’ threats to global biodiversity: Future scenarios of changes in the number of invasive species in a changing climate. Ecol. Indic. 2020, 116, 106436. [Google Scholar] [CrossRef]
- Liu, X.; Trogisch, S.; He, J.S.; Niklaus, P.A.; Bruelheide, H. Tree species richness increases ecosystem carbon storage in subtropical forests. Proc. Royal Soc. 2018, 285, 20181240. [Google Scholar] [CrossRef]
- Carrick, P.; Forsythe, K. The species composition-ecosystem function relationship: A global meta-analysis using data from intact and recovering ecosystems. PLoS ONE 2020, 15, e0236550. [Google Scholar] [CrossRef]
- Liu, X.; Swenson, N.G.; Lin, D.; Mi, X.; Umaña, M.N. Linking individual-level functional traits to tree growth in a subtropical forest. Ecology 2016, 97, 2396–2405. [Google Scholar] [CrossRef]
- Hejda, M.; Štajerová, K.; Pergl, J.; Pyšek, P. Impacts of dominant plant species on trait composition of communities: Comparison between the native and invaded ranges. Ecosphere 2019, 10, e02880. [Google Scholar] [CrossRef]
- Kraft, N.; Godoy, O.; Levine, J. Plant functional traits and the multidimensional nature of species coexistence. Proc. Natl. Acad. Sci. USA 2015, 112, 797–802. [Google Scholar] [CrossRef]
- Sharma, N.; Rayamajhi, M. Different aspects of weed management in maize (Zea mays L.): A brief review. Adv. Agric. 2022, 2022, 7960175. [Google Scholar] [CrossRef]
- Gage, K.; Krausz, R.; Walters, S. Emerging challenges for weed management in herbicide-resistant crops. Agriculture 2019, 9, 180. [Google Scholar] [CrossRef]
- Loddo, D.; Scarabe, L.; Sattin, M.; Pederzoli, A.; Morsiani, C.; Canestrale, R.; Tommasini, M.G. Combination of herbicide band application and inter-row cultivation provides sustainable weed control in maize. Agronomy 2019, 10, 20. [Google Scholar] [CrossRef]
- Silva, M.R.; Galon, L.; Rosseto, E.R.O.; Silva, A.F.; Favretto, E.L.; Brunetto, L.; Gallin, A.; Silva, A.M.L.; Tonin, R.J. Weed management in glyphosate-resistant maize. Arq. Do Inst. Biológico 2020, 87, e0862019. [Google Scholar] [CrossRef]
- Vencill, W.; Nichols, R.; Webster, T.; Soteres, J.; Mallory-Smith, C.; Burgos, N. Herbicide Resistance: Toward an understanding of resistance development and the impact of herbicide-resistant crops. Weed Sci. 2012, 60, 2–30. [Google Scholar] [CrossRef]
- Franzén, M.; Dieker, P.; Schrader, J.; Helm, A. Rapid plant colonization of the forelands of a vanishing glacier is strongly associated with species traits. Arct. Antarct. Alp. Res. 2019, 51, 366–378. [Google Scholar] [CrossRef]
- Guerra, J.G.; Cabello, F.; Quintanilla, C.F.; Pena, J.M.; Dorado, J. Plant functional diversity is affected by weed management through processes of trait convergence and divergence. Front. Plant Sci. 2022, 13, 993051. [Google Scholar] [CrossRef] [PubMed]
- Kraehmer, H.; Laber, B.; Rosinger, C.; Schulz, A. Herbicides as weed control agents: State of the art: I. Weed control research and SAFENER technology: The path to modern agriculture. Plant Physiol. 2014, 166, 1119–1131. [Google Scholar] [CrossRef]
- Espig, M.; Henwood, R.J. The social foundations for re-solving herbicide resistance in Canterbury, New Zealand. PLoS ONE 2023, 18, e0286515. [Google Scholar] [CrossRef] [PubMed]
- Nichols, V.; Verhulst, N.; Cox, R.; Govaerts, B. Weed dynamics and conservation agriculture principles: A review. Field Crops Res. 2015, 183, 56–68. [Google Scholar] [CrossRef]
- Owen, M.D. Diverse approaches to herbicide-resistant weed management. Weed Sci. 2016, 64, 570–584. [Google Scholar] [CrossRef]
- Colbach, N. The functional role of the soil seed bank in agricultural ecosystems. In Seeds: The Ecology of Regeneration in Plant Communities; CABI: Wallingford, UK, 2013; pp. 235–262. [Google Scholar] [CrossRef]
- Siebert, F.; Van Staden, N.; Komape, D.M.; Swemmer, A.M.; Siebert, S.J. Effects of land-use on herbaceous vegetation in a semi-arid Mopaneveld savanna. Bothalia 2021, 51, 1–26. [Google Scholar] [CrossRef]
- Shen, Y.; Yu, S.; Lian, J.; Shen, H.; Cao, H.; Lu, H.; Ye, W. Tree aboveground carbon storage correlates with environmental gradients and functional diversity in a tropical forest. Sci. Rep. 2016, 6, 25304. [Google Scholar] [CrossRef]
- Heijting, S.; Van Der Werf, W.; Kropff, M.J. Seed dispersal by forage harvester and rigid-tine cultivator in maize. Weed Res. 2009, 49, 153–163. [Google Scholar] [CrossRef]
- Tainton, N.M. Veld Management in South Africa; University of Natal Press: Pietermaritzburg, South Africa, 1999. [Google Scholar]
- Petit, A.N.; Fontaine, F.; Vatsa, P.; Clément, C.; Vaillant, G.N. Fungicide impacts on photosynthesis in crop plants. Photosynth. Res. 2012, 111, 315–326. [Google Scholar] [CrossRef]
- Siebert, F.; Klem, J.; Van Coller, H. Forb community responses to an extensive drought in two contrasting land-use types of a semi-arid Lowveld savanna. Afr. J. Range Forage Sci. 2020, 37, 53–64. [Google Scholar] [CrossRef]
- Barrett, S.H. Crop mimicry in weeds. Econ. Bot. 1983, 37, 255–282. [Google Scholar] [CrossRef]
- Heap, I. Global perspective of herbicide resistant. Pest Manag. Sci. 2013, 70, 1306–1315. [Google Scholar] [CrossRef]
- Charbonneau, A.; Tack, D.; Lale, A.; Goldston, J.; Caple, M.; Conner, E.; Barazani, O.; Ziffer-Berger, J.; Dworkin, I.; Conner, J.K. Weed evolution: Genetic differentiation among wild, weedy, and crop radish. Evol. Appl. 2018, 11, 1964–1974. [Google Scholar] [CrossRef] [PubMed]
- Lososová, Z.; Chytry, M.; Kuhn, I. Plant attributes determining the regional abundance of weeds on central European arable land. J. Biogeogr. 2008, 35, 177–187. [Google Scholar] [CrossRef]
- Chipomho, J.; Mupeti, S.; Chipomho, C.; Mashavakure, N.; Mashingaidze, A.B. Evaluation of a pre-formulated post-emergence herbicide mixture of topramezone and dicamba on annual weeds and Bermuda grass in maize in a sub-tropical agro-ecology. Heliyon 2019, 5, e01712. [Google Scholar] [CrossRef]
- Bogale, A. Reducing weed impacts and yield losses by application of herbicides in summer-grown maize. Int. J. Agric. Sci. Food Technol. 2023, 9, 49–53. [Google Scholar] [CrossRef]
- Kwinda, M.; Siebert, S.J.; Van Coller, H.; Masehela, T.S. Composition and diversity patterns of weeds in herbicide tolerant maize fields and margins in the Eastern Cape, South Africa. Biodiversitas 2023, 24, 399–414. [Google Scholar] [CrossRef]
- Dufrene, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Aronson, M.F.; Handel, S.N.; Clemants, S.E. Fruit type, life form and origin determine the success of woody plant invaders in an urban landscape. Biol. Invasions. 2007, 9, 465–475. [Google Scholar] [CrossRef]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Kent, M.; Coker, P. Vegetation Description and Analysis. A Practical Approach; John Wiley & Sons: New York, NY, USA, 1994. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E: Plymouth, MA, USA, 2006; Available online: http://primer-e.com (accessed on 4 May 2020).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Functional Types | Life History | Species Names | Functional Traits Description (Supplementary Table S2) |
|---|---|---|---|
| PFT1 | Annual |
| Annual, dicotyledonous, prostrate and erect-growing, non-nitrogen-fixing weeds with specialised carbon storage organs. |
| PFT2 | Annual |
| Annual, monocotyledonous and dicotyledonous, non-nitrogen-fixing weeds with unassisted seed dispersal and specialised carbon storage organs. |
| PFT3 | Annual |
| Annual, monocotyledonous and dicotyledonous, nitrogen-fixing weeds with unassisted seed dispersal and specialised carbon storage organs. |
| PFT4 | Annual |
| Annual, monocotyledonous and dicotyledonous, nitrogen-fixing and non-spinescent weeds following an assisted seed dispersal and specialised carbon storage organs. |
| PFT5 | Annual |
| Annual, monocotyledonous and dicotyledonous, non-nitrogen-fixing and non-spinescent weeds following an assisted seed dispersal and specialised carbon storage organs with a C3 and C4 photosynthetic pathway. |
| PFT6 | Annual |
| Annual, monocotyledonous and dicotyledonous, nitrogen- and non-nitrogen-fixing spinescent weeds with assisted and unassisted seed dispersal following a specialised carbon storage organ. |
| PFT7 | Perennial/Biennial |
| Perennial and biennial, assisted and unassisted seed dispersal weed species with specified flowering season and no specialised carbon storage organs. |
| PFT8 | Perennial/Biennial |
| Perennial and biennial, nitrogen- and non-nitrogen-fixing weed species with non-spinescent, specified flowering season, and specialised carbon storage organs. |
| PFT9 | Perennial/Biennial |
| Perennial and biennial, nitrogen- and non-nitrogen-fixing weeds following non-spinescent and spinescent with assisted seed dispersal, specified flowering season, and specialised carbon storage organs. |
| PFT10 | Perennial/Biennial |
| Perennial and biennial, nitrogen- and non-nitrogen-fixing non-spinescent with assisted seed dispersal following specified flowering season with specialised carbon storage organs. |
| Statistic | Axis 1 | Axis 2 |
|---|---|---|
| Eigenvalues | 0.06 | 0.21 |
| Explained variation (% cumulative) | 5.8 | 27.19 |
| Pseudo-canonical correlation | 0.61 | <0.005 |
| Variable | Mean | t-Value | Df | p | Valid N | Std.Dev. | F-Ratio Variance | p Variance |
|---|---|---|---|---|---|---|---|---|
| PFT1-Pre | 1.394 | −2.601 | 206 | 0.009 | 104 | 7.683 | 30.517 | <0.01 |
| PFT1-Post | 12.394 | 104 | 42.446 | |||||
| PFT2-Pre | 10.125 | 2.663 | 206 | 0.008 | 104 | 34.077 | 54.299 | <0.01 |
| PFT2-Post | 1.144 | 104 | 4.625 | |||||
| PFT3-Pre | 40.134 | −2.385 | 206 | 0.017 | 104 | 108.438 | 6.503 | <0.01 |
| PFT3-Post | 109.615 | 104 | 276.539 | |||||
| PFT4-Pre | 11.471 | 1.567 | 206 | 0.118 | 104 | 42.59 | 2.806 | <0.01 |
| PFT4-Post | 3.846 | 25.42 | ||||||
| PFT5-Pre | 48.278 | −2.922 | 206 | 0.003 | 104 | 100.933 | 10.346 | <0.01 |
| PFT5-Post | 145.721 | 324.660 | ||||||
| PFT6-Pre | 19.615 | 0.421 | 206 | 0.674 | 104 | 74.676 | 1.699 | <0.01 |
| PFT6-Post | 15.730 | 57.281 | ||||||
| PFT7-Pre | 62.932 | 1.155 | 206 | 0.249 | 104 | 169.276 | 1.058 | 0.773 |
| PFT7-Post | 35.413 | 174.168 | ||||||
| PFT8-Pre | 17.865 | −1.819 | 206 | 0.070 | 104 | 63.501 | 1.093 | 0.650 |
| PFT8-Post | 34.259 | 66.409 | ||||||
| PFT9-Pre | 44.326 | 0.673 | 206 | 0.501 | 104 | 74.706 | 1.659 | <0.01 |
| PFT9-Post | 36.278 | 96.237 | ||||||
| PFT10-Pre | 33.231 | 1.507 | 206 | 0.133 | 104 | 108.620 | 6.831 | <0.01 |
| PFT10-Post | 16.038 | 41.557 |
| Plant Functional Type (PFT) | Increase/Decrease | Significant Difference p < 0.05 |
|---|---|---|
| PFT1 | Increase | √ |
| PFT2 | Decrease | √ |
| PFT3 | Increase | √ |
| PFT4 | Increase | X |
| PFT5 | Increase | √ |
| PFT6 | Decrease | X |
| PFT7 | Decrease | X |
| PFT8 | Increase | X |
| PFT9 | Decrease | X |
| PFT10 | Decrease | X |
| Scientific Name | Average Sum | Ave Sum Pre | Ave Sum Post | Herbicide-Resistant | PFT | Traits | ||
|---|---|---|---|---|---|---|---|---|
| Carbon Storage | Nitrogen-Fixing Ability | Life Span | ||||||
| Amaranthus viridis | 62.77 | 33.61 | 91.93 | √ | 3 | Specialised storage organ | N-Fixer | Annual |
| Bromus diandrus | 10.70 | 6.31 | 15.08 | √ | 3 | Specialised storage organ | N-Fixer | Annual |
| Bulbostylis humilis | 16.69 | 9.95 | 23.44 | √ | 5 | Specialised storage organ | Non-N-Fixer | Annual |
| Chenopodium carinatum | 36.83 | 10.94 | 62.73 | √ | 5 | Specialised storage organ | Non-N-Fixer | Annual |
| Cotula heterocarpa | 20.14 | 40.20 | 0.09 | × | 7 | Specialised storage organ | N-Fixer | Perennial |
| Cyperus spp. | 25.36 | 17.33 | 33.39 | √ | 8 | Specialised storage organ | N-Fixer | Perennial |
| Euphorbia prostrata | 22.38 | 16.44 | 28.32 | √ | 4 | Specialised storage organ | N-Fixer | Annual |
| Ipomoea purpurea | 32.59 | 12.14 | 53.04 | √ | 5 | Specialised storage organ | Non-N-Fixer | Annual |
| Sonchus asper | 13.67 | 16.86 | 10.48 | × | 6 | Specialised storage organ | Non-N-Fixer | Annual |
| Tephrosia capensis | 16.60 | 7.75 | 24.45 | √ | 9 | Specialised storage organ | Non-N-Fixer | Perennial |
| Seasons | Pre-Emergence | Post-Emergence | Selective (S)/Non-Selective (NS) | Measurement/Use Per Hectare (ha)–200 L of Water Mixing Ratio Used Per ha | ||
|---|---|---|---|---|---|---|
| Active Ingredient | Trade Name | Active Ingredient | Trade Name | |||
| 2017/18 | Primagram gold | Primagram gold | Primagram gold | Primagram gold | S | 2.5 L/ha |
| 2,4-D Amine | 2-4-D Amine 480 SL | 2,4-D Amine | 2-4-D Amine 480 SL | S | * 2–5.4 L/ha | |
| Halosulfuron | Halo 750 WDG | Halosulfuron | Halo 750 WDG | S | 50 g/ha | |
| Glyphosate | PowerMax | Glyphosate | PowerMax | NS | 3 L/ha | |
| Sorgomil | Sorgomil Gold 600 SC | Sorgomil | Sorgomil Gold 600 SC | S | 1.4 L/ha | |
| 2018/19 | Alpha-cypermethrin | Alpha-thrin 100 SC | Alpha-cypermethrin | Alpha-thrin 100 SC | Insecticide | 200 mL/ha |
| Metolachlor | Metolachlor 915 EC | Metolachlor | Metalochlor (^) EC | S | ^ 1–2 L/ha | |
| Glyphosate | PowerMax | Glyphosate | PowerMax | NS | 3 L/ha | |
| Mesotrione | Cantron 480 SC | Halosulfuron | Halo 750 WDG (50g water soluble bag) | S | 50 g/ha | |
| Terbuthylazine | Terbuweed 600 WDG | S | * 2–4 L/ha | |||
| 2,4-D Amine | 2,4-D Amine 480 SL | S | * 2–5.4 L/ha | |||
| Adjuvant | Villa 51 | Wetting agent | 12 mL/ha | |||
| 2019/20 | Alpha-cypermethrin | Alpha-thrin 100 SC | Post-emergence herbicides were not applied | Insecticide | 200 mL/ha | |
| Metolachlor | Metolachlor 915 EC | S | ^ 1–2 L/ha | |||
| Mesotrione | Cantron 480 SC | S | ** 210–260 mL/ha | |||
| Glyphosate | PowerMax | NS | 3 L/ha | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwinda, M.; Siebert, S.J.; Van Coller, H.; Masehela, T.S. Identifying Plant Functional Traits of Weeds in Fields Planted with Glyphosate-Tolerant Maize for Preferable Weed Management Practices. Agriculture 2024, 14, 223. https://doi.org/10.3390/agriculture14020223
Kwinda M, Siebert SJ, Van Coller H, Masehela TS. Identifying Plant Functional Traits of Weeds in Fields Planted with Glyphosate-Tolerant Maize for Preferable Weed Management Practices. Agriculture. 2024; 14(2):223. https://doi.org/10.3390/agriculture14020223
Chicago/Turabian StyleKwinda, Murendeni, Stefan John Siebert, Helga Van Coller, and Tlou Samuel Masehela. 2024. "Identifying Plant Functional Traits of Weeds in Fields Planted with Glyphosate-Tolerant Maize for Preferable Weed Management Practices" Agriculture 14, no. 2: 223. https://doi.org/10.3390/agriculture14020223
APA StyleKwinda, M., Siebert, S. J., Van Coller, H., & Masehela, T. S. (2024). Identifying Plant Functional Traits of Weeds in Fields Planted with Glyphosate-Tolerant Maize for Preferable Weed Management Practices. Agriculture, 14(2), 223. https://doi.org/10.3390/agriculture14020223