

Application of a Mechanistic Model to Explore Management Strategies for Biological Control of an Agricultural Pest

 ,
,  , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Models
2.2. Crop Model and Climate Inputs
2.3. Initial Conditions
2.4. Timing of Canola Sowing and Aphid Arrival
2.5. Biological Control
2.6. Chemical and Biological Controls
3. Results
3.1. Timing of Canola Sowing and Aphid Arrival
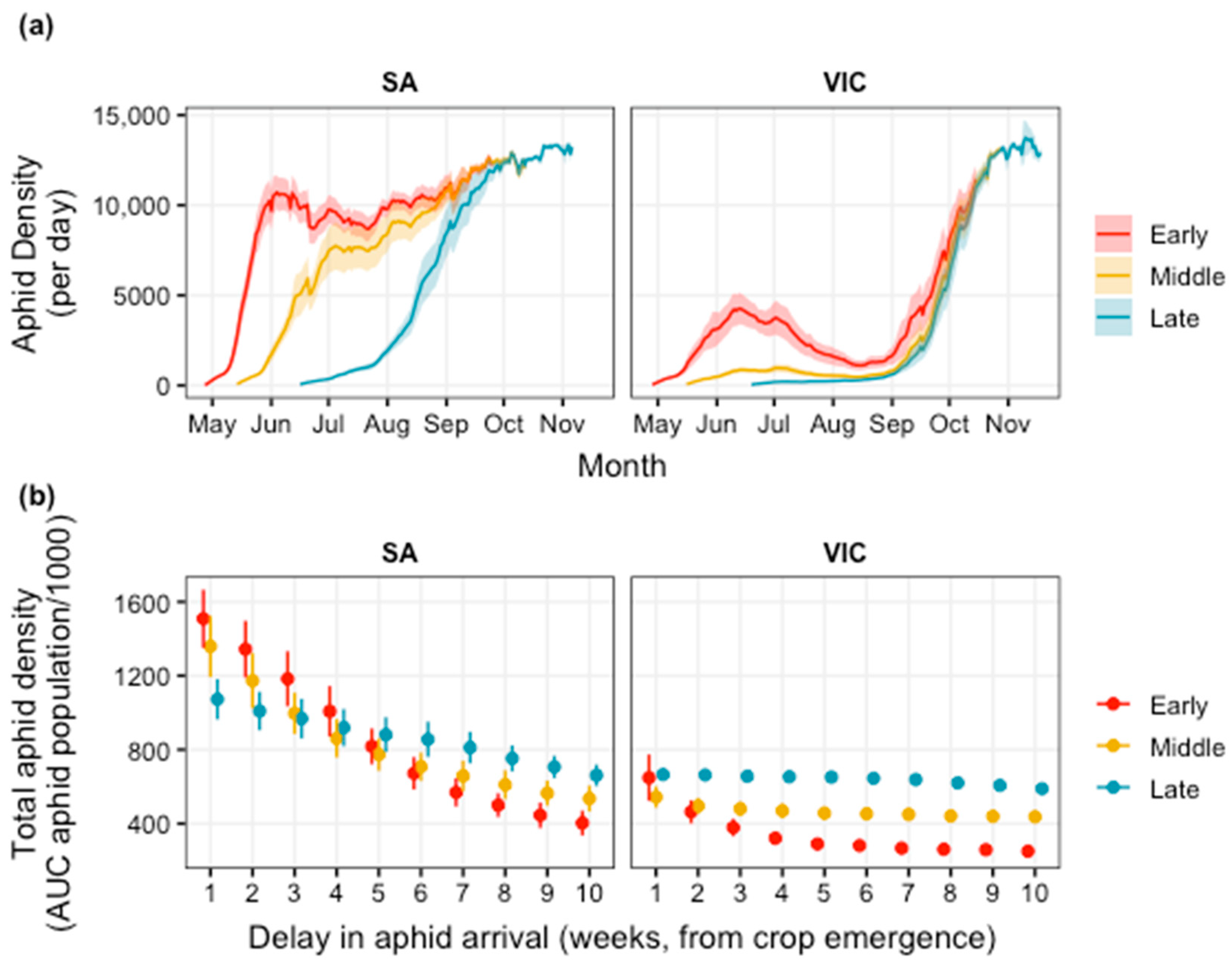
3.2. Biological Control
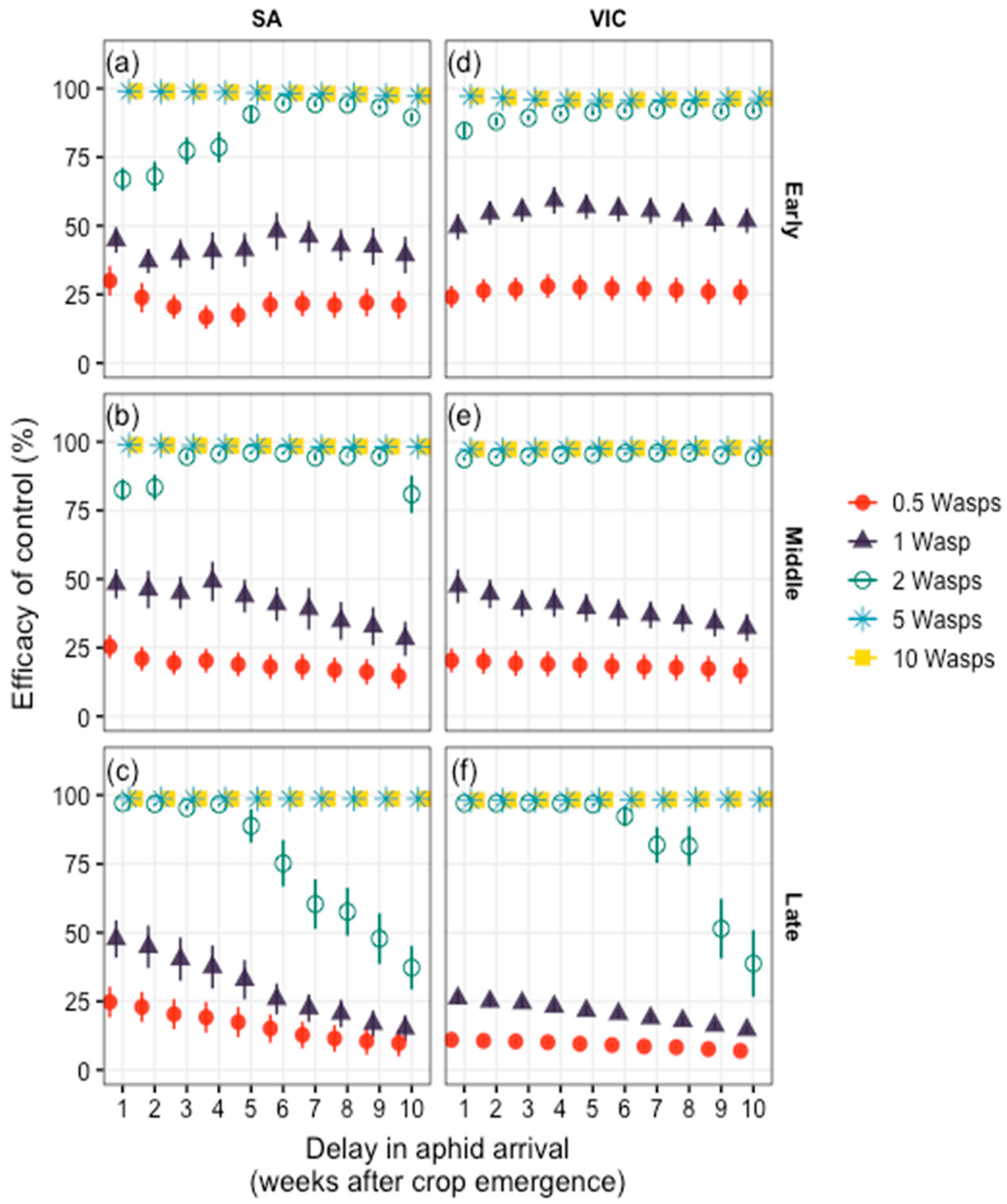
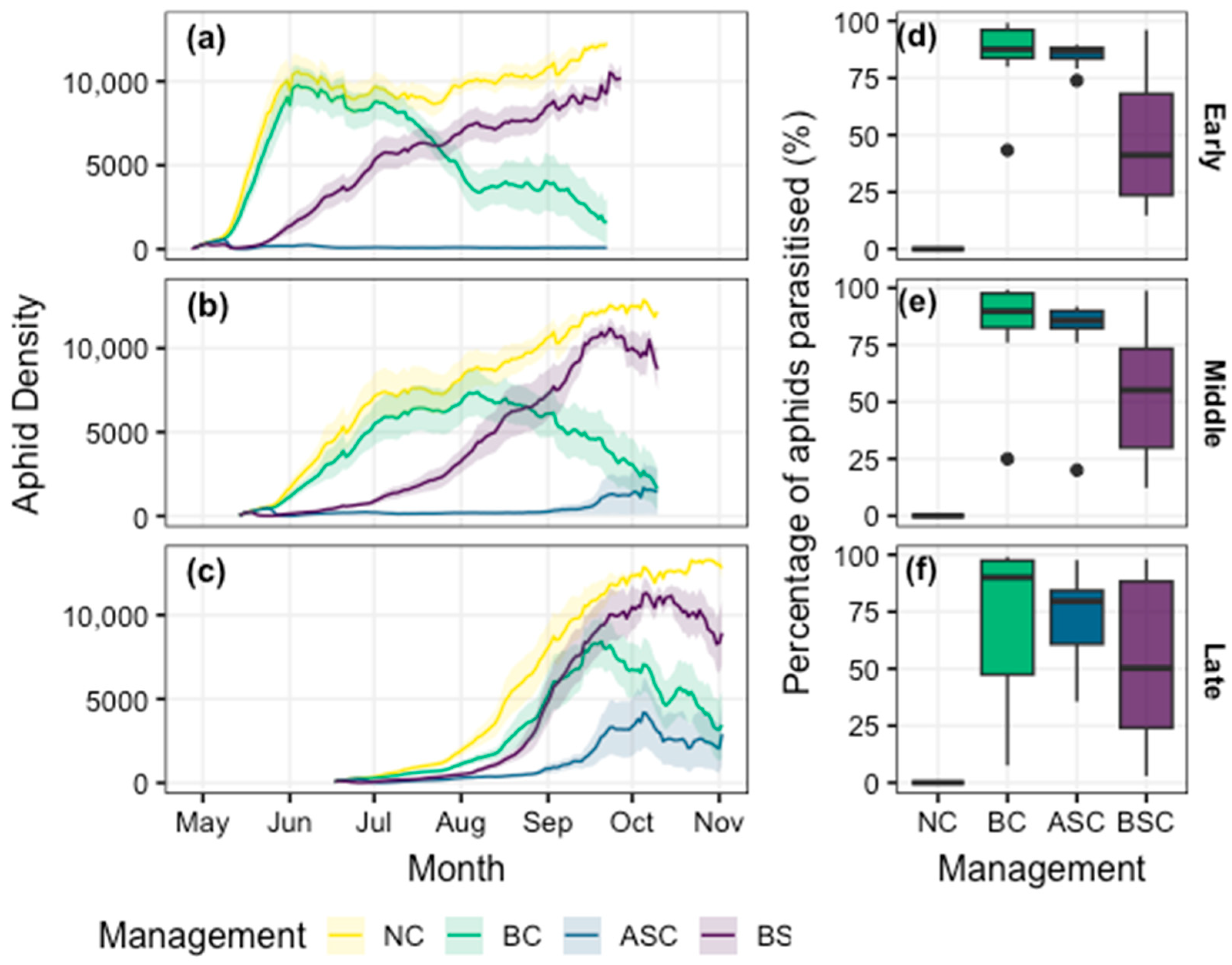
3.3. Chemical and Biological Controls
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stern, V.; Smith, R.; van den Bosch, R.; Hagen, K. The integration of chemical and biological control of the spotted alfalfa aphid: The integrated control concept. Hilgardia 1959, 29, 81–101. [Google Scholar] [CrossRef]
- Zalucki, M.P.; Adamson, D.; Furlong, M.J. The future of IPM: Whither or wither? Aust. J. Entomol. 2009, 48, 85–96. [Google Scholar] [CrossRef]
- Hill, M.P.; Macfadyen, S.; Nash, M.A. Broad spectrum pesticide application alters natural enemy communities and may facilitate secondary pest outbreaks. PeerJ 2017, 5, e4179. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, D. Pesticides and pest control. In Integrated Pest Management: Innovation-Development Process; Peshin, R., Dhawa, A.K., Eds.; Springer: Dordrecht, Germany, 2009; Volume 1, pp. 83–87. [Google Scholar]
- Horne, P.A.; Page, J.; Nicholson, C. When will integrated pest management strategies be adopted? Example of the development and implementation of integrated pest management strategies in cropping systems in Victoria. Anim. Prod. Sci. 2008, 48, 1601–1607. [Google Scholar] [CrossRef]
- Micic, S.; Hoffmann, A.A.; Strickland, G.; Weeks, A.R.; Bellati, J.; Henry, K.; Nash, M.A.; Umina, P.A. Pests of germinating grain crops in southern Australia: An overview of their biology and management options. Aust. J. Exp. Agric. 2008, 48, 1560–1573. [Google Scholar] [CrossRef]
- Macfadyen, S.; Moradi-Vajargah, M.; Umina, P.A.; Hoffmann, A.; Nash, M.; Holloway, J.; Severtson, D.; Hill, M.; Van Helden, M.; Barton, M. Identifying critical research gaps that limit control options for invertebrate pests in Australian grain production systems. Austral Entomol. 2019, 58, 9–26. [Google Scholar] [CrossRef]
- McDonald, G.; Umina, P.A.; Macfadyen, S.; Mangano, P.; Hoffmann, A.A. Predicting the timing of first generation egg hatch for the pest redlegged earth mite Halotydeus destructor (Acari: Penthaleidae). Exp. Appl. Acar. 2015, 65, 259–276. [Google Scholar] [CrossRef]
- Schliephake, E.; Graichen, K.; Rabenstein, F. Investigations on the vector transmission of the Beet mild yellowing virus (BMYV) and the Turnip yellows virus (TuYV). J. Plant Dis. Protect. 2000, 107, 81–87. [Google Scholar]
- Jones, R.A.C.; Coutts, B.A. Yield-limiting potential of Beet western yellows virus in Brassica napus. Aust. J. Agric. Res. 2007, 58, 788–801. [Google Scholar] [CrossRef]
- Congdon, B.S.; Kehoe, M.A.; Filardo, F.F.; Coutts, B.A. In-field capable loop-mediated isothermal amplification detection of Turnip yellows virus in plants and its principal aphid vector Myzus persicae. J. Virol. Methods 2019, 265, 15–21. [Google Scholar] [CrossRef]
- Umina, P.A.; McDonald, G.; Maino, J.; Edwards, O.; Hoffmann, A.A. Escalating insecticide resistance in Australian grain pests: Contributing factors, industry trends and management opportunities. Pest Manag. Sci. 2019, 75, 1494–1506. [Google Scholar] [CrossRef]
- Pym, A.; Umina, P.A.; Reidy-Crofts, J.; Troczka, B.J.; Matthews, A.; Gardner, J.; Hunt, B.; van Rooyen, A.; Edwards, O.; Bass, C. Overexpression of UDP-glucuronosyltransferase and cytochrome P450 enzymes confers resistance to sulfoxaflor in field populations of the aphid, Myzus persicae. Insect Biochem. Molec. 2022, 143, 103743. [Google Scholar] [CrossRef]
- Ferreira, S.E.; Sampaio, M.V.; Silva de Oliveira, R.; Vasconcelos, H.L.D. Parasitism rate of Myzus persicae (Sulzer) by Diaeretiella rapae (McIntosh) in the presence of an alternative, resistant host. Rev. Bras. Entomol. 2018, 62, 13–18. [Google Scholar] [CrossRef]
- Ward, S.; Umina, P.A.; Parry, H.; Balfour-Cunningham, A.; Cheng, X.; Heddle, T.; Holloway, J.C.; Langley, C.; Severtson, D.; Van Helden, M.; et al. Is what you see what you get? The relationship between field observed and actual aphid parasitism rates in canola crops. Pest Manag. Sci. 2022, 78, 3596–3607. [Google Scholar] [CrossRef]
- Bannerman, J.A.; Roitberg, B.D. Impact of extreme and fluctuating temperatures on aphid–parasitoid dynamics. Oikos 2014, 123, 89–98. [Google Scholar] [CrossRef]
- Ro, T.H.; Long, G.E. GPA-Phenodynamics, a simulation model for the population dynamics and phenology of green peach aphid in potato: Formulation, validation, and analysis. Ecol. Model. 1999, 119, 197–209. [Google Scholar] [CrossRef]
- Trivedi, T.; Khurana, S.; Puri, S.; Bhar, L.; Mehta, S.; Jain, R.; Singh, G.; Chaudhari, S.; Mohasin, M.; Dhandapani, A. Development of forewarning system of potato aphid (Myzus persicae) on potato (Solanum tuberosum) in India. Indian J. Agric. Sci. 2002, 72, 341–345. [Google Scholar]
- Zhu, M.; Radcliffe, E.B.; Ragsdale, D.W.; MacRae, I.V.; Seeley, M.W. Low-level jet streams associated with spring aphid migration and current season spread of potato viruses in the U.S. northern Great Plains. Agric. Forest Meteorol. 2006, 138, 192–202. [Google Scholar] [CrossRef]
- Maling, T.; Diggle, A.J.; Thackray, D.J.; Siddique, K.H.M.; Jones, R.A.C. An epidemiological model for externally acquired vector-borne viruses applied to Beet western yellows virus in Brassica napus crops in a Mediterranean-type environment. Crop Pasture Sci. 2010, 61, 132–144. [Google Scholar] [CrossRef]
- Neuville, S.; Le Ralec, A.; Outreman, Y.; Jaloux, B. The delay in arrival of the parasitoid Diaeretiella rapae influences the efficiency of cabbage aphid biological control. BioControl 2016, 61, 115–126. [Google Scholar] [CrossRef]
- Stark, J.D.; Acheampong, S. A demographic and modeling approach to determine the suitability of two hosts, Brevicoryne brassicae (Linnaeus) and Myzus persicae (Sulzer) (Heteroptera: Aphididae) of the aphid parasitoid, Diaeretiella rapae (McIntosh) (Hymenoptera: Aphidiidae). Pan-Pac. Entomol. 2007, 83, 75–77. [Google Scholar] [CrossRef]
- Bianchi, F.J.J.A.; Ives, A.R.; Shellhorn, N.A. Interactions between conventional and organic farming for biocontrol services across the landscape. Ecol. Appl. 2013, 23, 1531–1543. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Watt, M.S.; Withers, T.M.; Leriche, A.; Watson, M.C.A. process-based population dynamics model to explore target and non-target impacts of a biological control agent. Ecol. Model. 2009, 220, 2035–2050. [Google Scholar] [CrossRef]
- Sporleder, M.; Carhuapoma, P.; Juarez, H.; Gamarra, H.; Simon, R.; Kroschel, J. ILCYM-Insect Life Cycle Modeling. A Software Package for Developing Temperature-Based Insect Phenology Models with Applications for Local, Regional and Global Analysis of Insect Population and Mapping; International Potato Center: Lima, Peru, 2017. Available online: https://cgspace.cgiar.org/handle/10568/109024 (accessed on 3 March 2020).
- Barton, M.; Parry, H.; Ward, S.; Hoffmann, A.A.; Umina, P.A.; van Helden, M.; Macfadyen, S. Forecasting impacts of biological control under future climates: Mechanistic modelling of an aphid pest and a parasitic wasp. Ecol. Model. 2021, 457, 109679. [Google Scholar] [CrossRef]
- Moran, N.A. The evolution of aphid life cycles. Annu. Rev. Entomol. 1992, 37, 321–348. [Google Scholar] [CrossRef]
- Coutts, B.A.; Hawkes, J.R.; Jones, R.A.C. Occurrence of Beet western yellows virus and its aphid vectors in over-summering broad-leafed weeds and volunteer crop plants in the grainbelt region of south-western Australia. Aust. J. Agric. Res. 2006, 57, 975–982. [Google Scholar] [CrossRef]
- Mitsunaga, T.; Nagasaka, K.; Moriya, S. Differences in the reproductive durations of Myzus persicae (Hemiptera: Aphididae) parasitized by three aphidid parasitoids. Appl. Entomol. Zool. 2016, 51, 297–304. [Google Scholar] [CrossRef]
- Heddle, T.; Van Helden, M.; Michael Nash, M.; Muirhead, K. Parasitoid communities and interactions with Diuraphis noxia in Australian cereal production systems. BioControl 2020, 65, 571–582. [Google Scholar] [CrossRef]
- Holzworth, D.P.; Huth, N.I.; de Voil, P.G.; Zurcher, E.J.; Herrmann, N.I.; McLean, G.; Chenu, K.; van Oosterom, E.J.; Snow, V.; Murphy, C.; et al. APSIM–Evolution towards a new generation of agricultural systems simulation. Environ. Model. Softw. 2014, 62, 327–350. [Google Scholar] [CrossRef]
- Robertson, M.J.; Lilley, J.M. Simulation of growth, development and yield of canola (Brassica napus) in APSIM. Crop Pasture Sci. 2016, 67, 332–344. [Google Scholar] [CrossRef]
- Jeffrey, S.J.; Carter, J.O.; Moodie, K.B.; Beswick, A.R. Using spatial interpolation to construct a comprehensive archive of Australian climate data. Environ. Model. Softw. 2001, 16, 309–330. [Google Scholar] [CrossRef]
- GRDC (Grain Research and Development Corporation). Victorian Winter Crop Growing Guide. Department of Economic Development, Jobs, Transport and Resources, Victoria. 2023. Available online: https://grdc.com.au/resources-and-publications/all-publications/nvt-crop-sowing-guides/vic-tas-crop-sowing-guide (accessed on 10 January 2023).
- Rogers, D. Random search and insect population models. J. Anim. Ecol. 1972, 41, 369–383. [Google Scholar] [CrossRef]
- APVMA (Australian Pesticides and Veterinary Medicines Authority). APVMA Public Chemical Registration Information System Database. Available online: https://portal.apvma.gov.au/pubcris. (accessed on 21 October 2023).
- Abo El-Ghar, G.E.S.; El-Sayed, A.E.M. Long-term effects of insecticides on Diaeretiella rapae (M’Intosh), a parasite of the cabbage aphid. Pestic. Sci. 1992, 36, 109–114. [Google Scholar] [CrossRef]
- Abo El-Ghar, G.E.S.; El-Sayed, A.E.M. Impact of two synthetic pyrethroids and methomyl on management of the Cabbage Aphid, Brevicoryne brassicae (L.) and its Associated Parasitoid, Diaeretiella rapae (M’Intosh). Pestic. Sci. 1989, 25, 35–41. [Google Scholar] [CrossRef]
- Kirkland, L.S.; Chirgwin, E.; Ward, S.E.; Congdon, B.S.; van Rooyen, A.; Umina, P.A. P450-mediated resistance in Myzus persicae (Sulzer) (Hemiptera: Aphididae) reduces the efficacy of neonicotinoid seed treatments in Brassica napus. Pest Manag. Sci. 2023, 79, 1851–1859. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023. Available online: https://www.R-project.org/ (accessed on 1 July 2023).
- Snyder, W.E.; Ives, A.R. Interactions between specialist and generalist natural enemies: Parasitoids, predators, and pea aphid biocontrol. Ecology 2003, 84, 91–107. [Google Scholar] [CrossRef]
- McDougall, R.; Mata, L.; Ward, S.; Hoffmann, A.; Umina, P.A. Assessing the sub-lethal impacts of insecticides on aphid parasitoids through laboratory-based studies. Austral. Entomol. 2023; in revision. [Google Scholar]
- Ward, S.; Hoffmann, A.A.; van Helden, M.; Umina, P.A. The effects of insecticide seed treatments on the parasitism and predation of green peach aphids, Myzus persicae (Homoptera: Aphididae). J. Econ. Entomol. 2023, accepted. [Google Scholar]
- Calvo-Agudo, M.; Dregni, J.; González-Cabrera, J.; Dicke, M.; Heimpel, G.E.; Tena, A. Neonicotinoids from coated seeds toxic for honeydew-feeding biological control agents. Environ. Pollut. 2021, 289, 117813. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Management Scenario | Location | TOS | Aphid Arrivals | Wasp Density | Control Scenarios | No. Scenarios |
|---|---|---|---|---|---|---|
| Seasonal patterns in aphid population dynamics (Figure 2) (When and where is intervention required?) | SA Victoria | Early Middle Late | Weekly, from day of crop emergence, for 10 weeks | 0 | No control | 60 |
| Biological control (Figure 3) (How many wasps are required to suppress aphid population growth?) | SA Victoria | Early Middle Late | Weekly, from day of crop emergence, for 10 weeks | (0), 0.5, 1, 2, 5, 10 | No control/ Wasp control | 360 |
| Chemical and biological controls (Figure 4) (Can chemicals hinder or facilitate biological control?) | SA | Early Middle Late | Day of crop emergence | (0), 1 | No control/ Wasp control/Aphid-specific/Broad-spectrum | 12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barton, M.G.; Parry, H.; Umina, P.A.; Binns, M.R.; Heddle, T.; Hoffmann, A.A.; Holloway, J.; Severtson, D.; Van Helden, M.; Ward, S.; et al. Application of a Mechanistic Model to Explore Management Strategies for Biological Control of an Agricultural Pest. Agriculture 2024, 14, 150. https://doi.org/10.3390/agriculture14010150
Barton MG, Parry H, Umina PA, Binns MR, Heddle T, Hoffmann AA, Holloway J, Severtson D, Van Helden M, Ward S, et al. Application of a Mechanistic Model to Explore Management Strategies for Biological Control of an Agricultural Pest. Agriculture. 2024; 14(1):150. https://doi.org/10.3390/agriculture14010150
Chicago/Turabian StyleBarton, Madeleine G., Hazel Parry, Paul A. Umina, Matthew R. Binns, Thomas Heddle, Ary A. Hoffmann, Joanne Holloway, Dustin Severtson, Maarten Van Helden, Samantha Ward, and et al. 2024. "Application of a Mechanistic Model to Explore Management Strategies for Biological Control of an Agricultural Pest" Agriculture 14, no. 1: 150. https://doi.org/10.3390/agriculture14010150
APA StyleBarton, M. G., Parry, H., Umina, P. A., Binns, M. R., Heddle, T., Hoffmann, A. A., Holloway, J., Severtson, D., Van Helden, M., Ward, S., Wood, R., & Macfadyen, S. (2024). Application of a Mechanistic Model to Explore Management Strategies for Biological Control of an Agricultural Pest. Agriculture, 14(1), 150. https://doi.org/10.3390/agriculture14010150