
Pan-Genome-Wide Identification and Transcriptome-Wide Analysis of ZIP Genes in Cucumber

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Identification of ZIP Genes in C. sativus
2.2. Motif Analysis and Gene Structure
2.3. Phylogenetic Analysis and Multiple Sequence Alignment
2.4. Gene Duplication Analysis and Genome Distribution
2.5. Analysis of Promoter Regions of CsZIP Genes
2.6. Analysis of Transcriptome Data
2.7. Plant Materials and Growth Conditions
2.8. RNA isolation and Real-Time PCR Analysis
3. Results
3.1. Identification of CsZIP Genes from C. sativus Pan-Genome
3.2. Characterization of CsZIP Genes from Chinese Long 9930
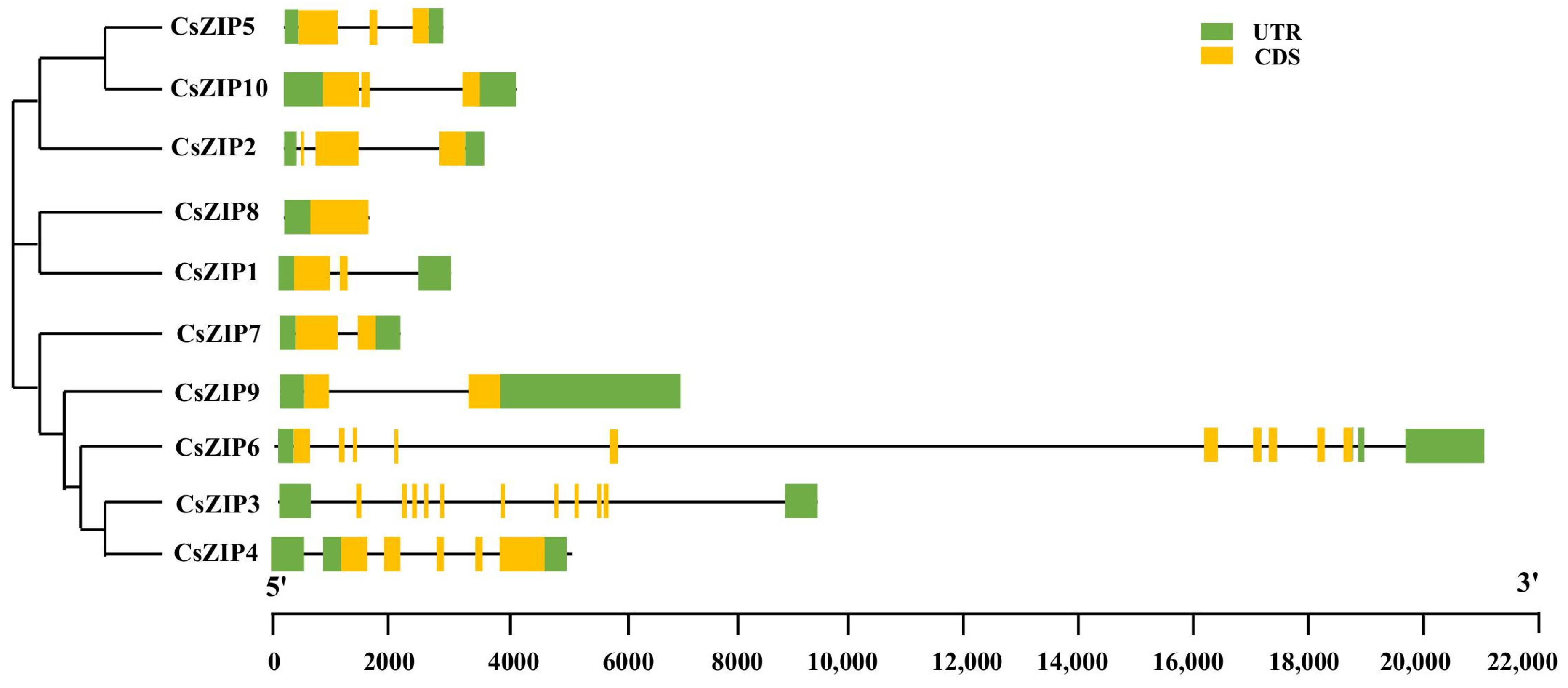
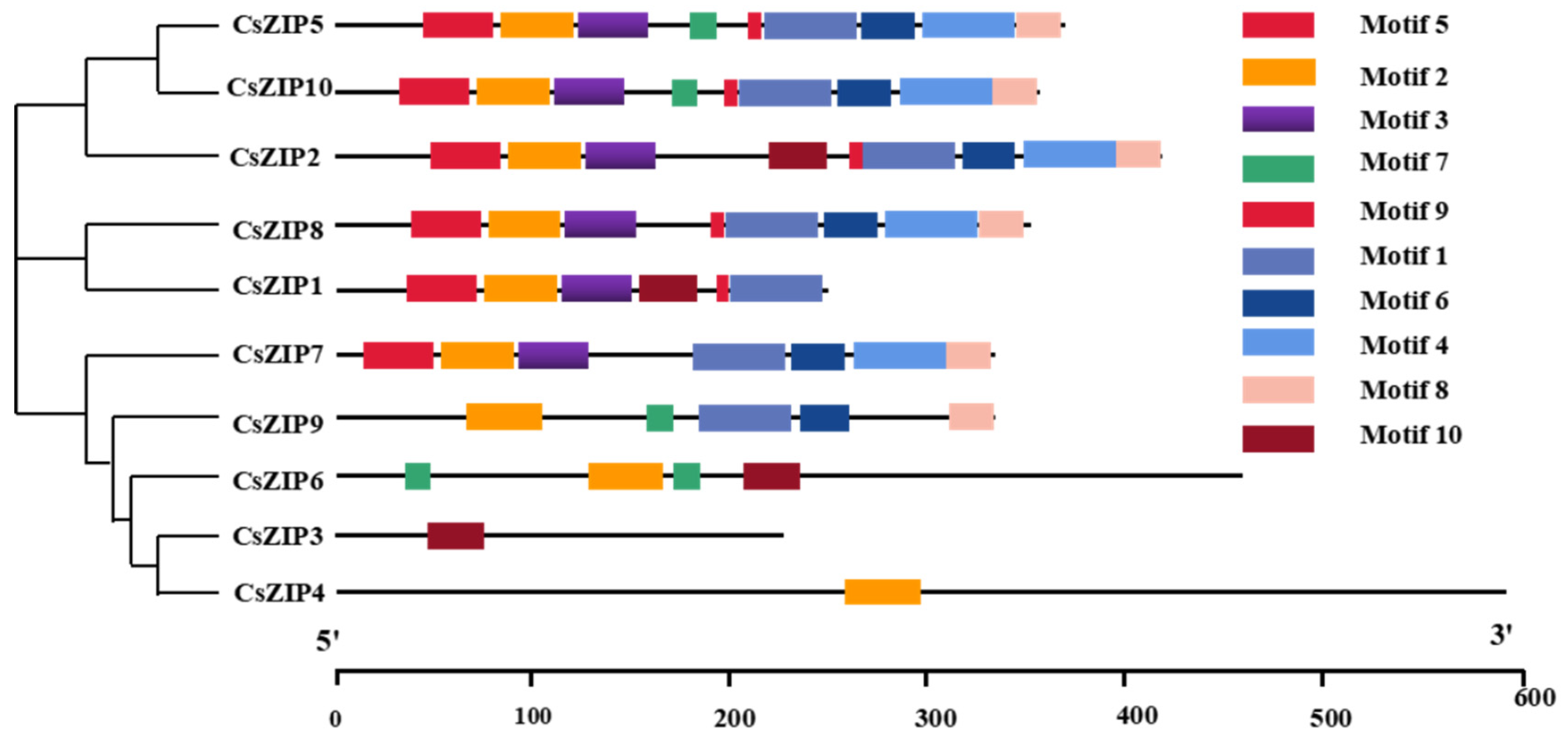
3.3. Analysis of Phylogenetic Relationship and Gene Structure and Protein Motif
3.4. Multiple Sequence Alignment
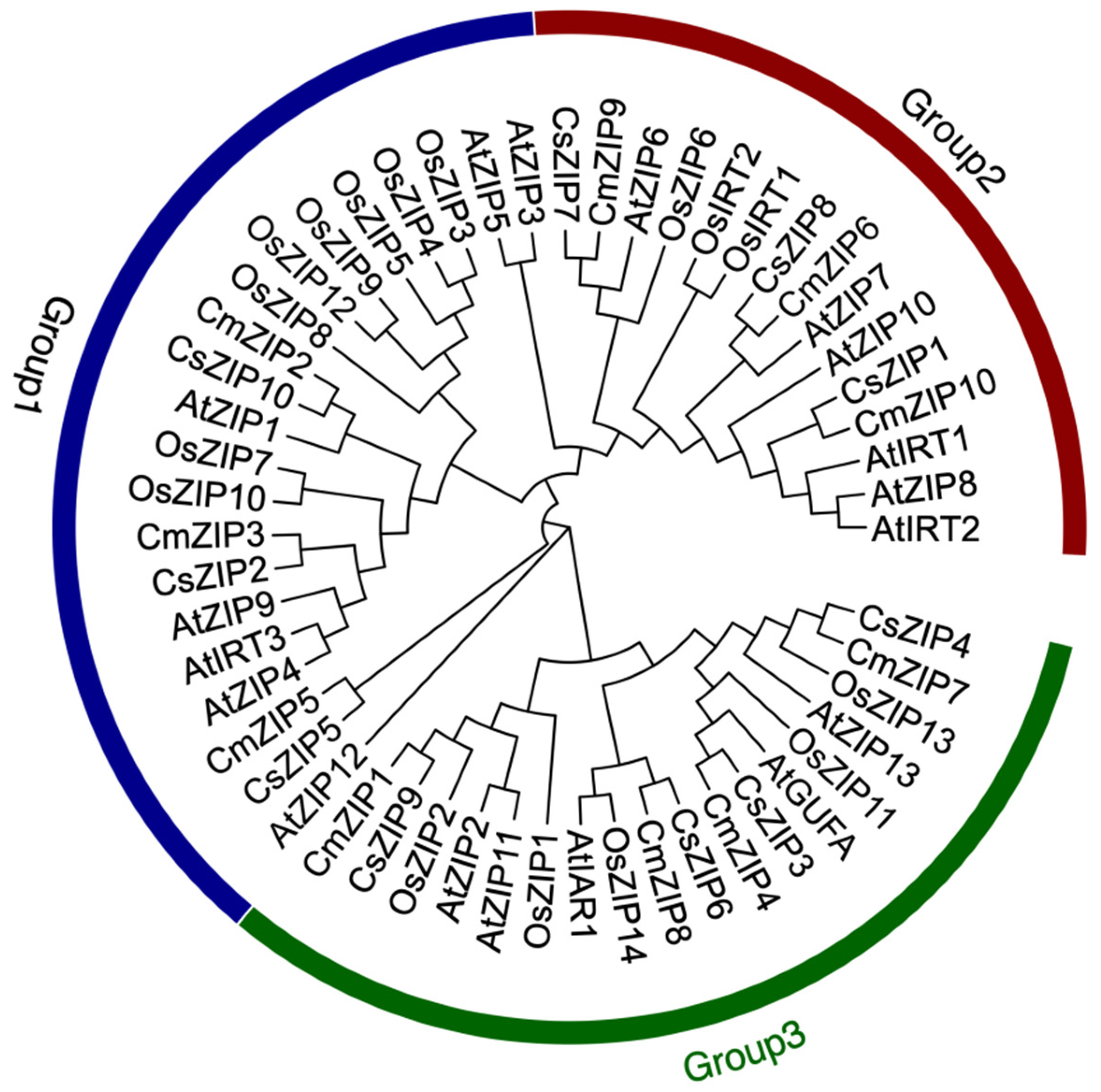
3.5. Phylogenetic Analysis among Different Species
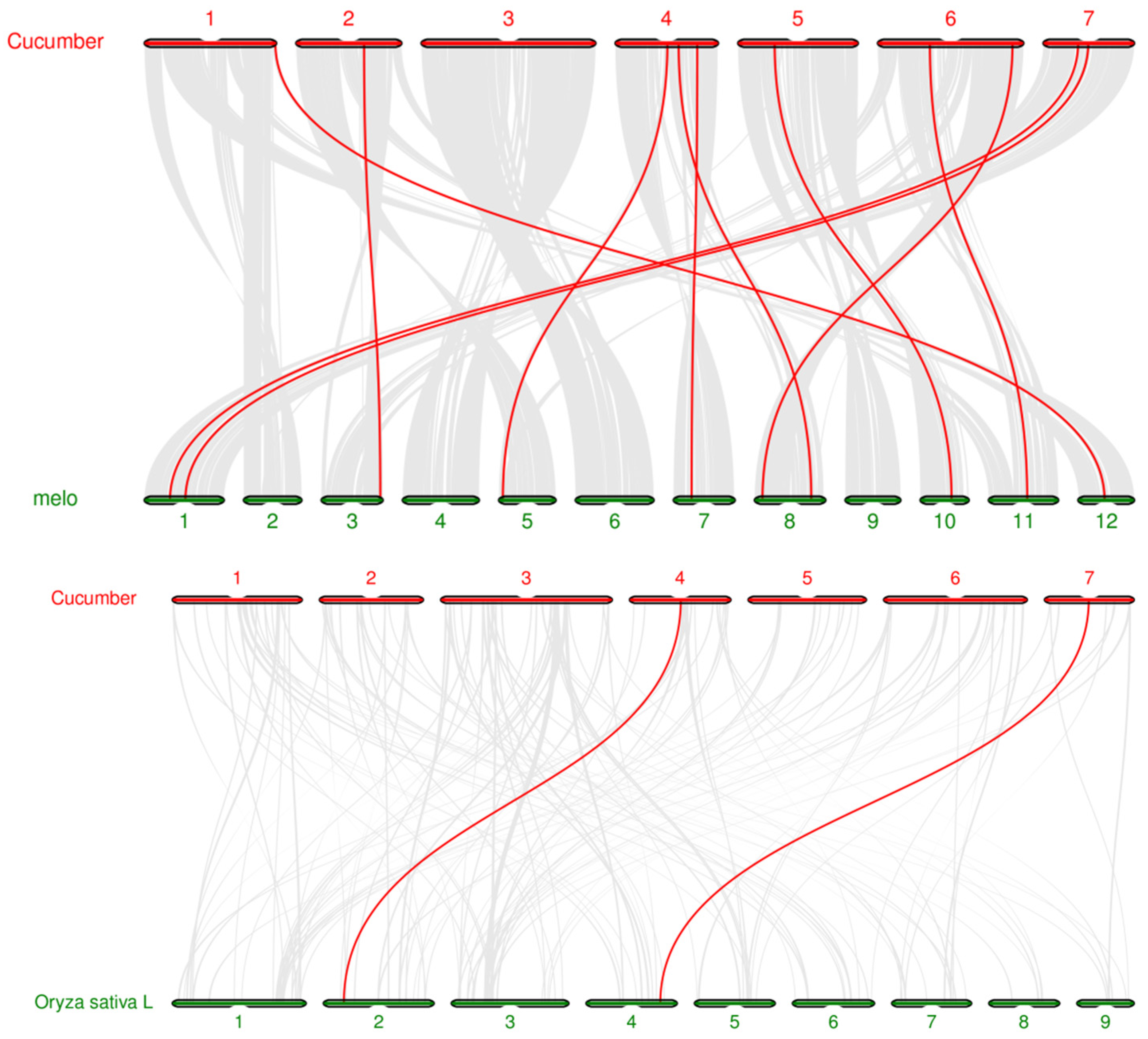
3.6. Chromosome Localization and Collinearity Analysis
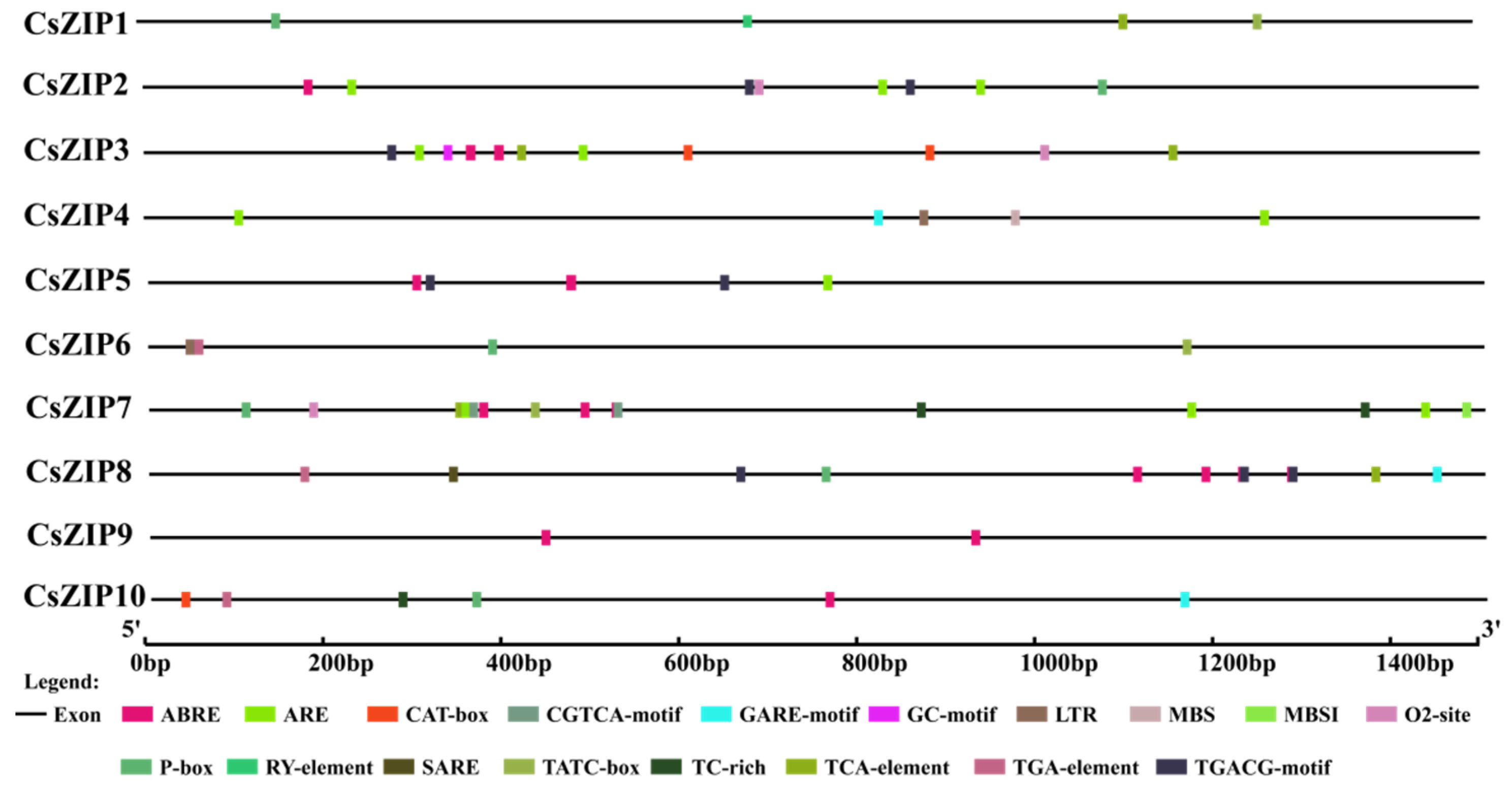
3.7. Analysis of Cis-Acting Elements in Promoter Region of CsZIP Genes
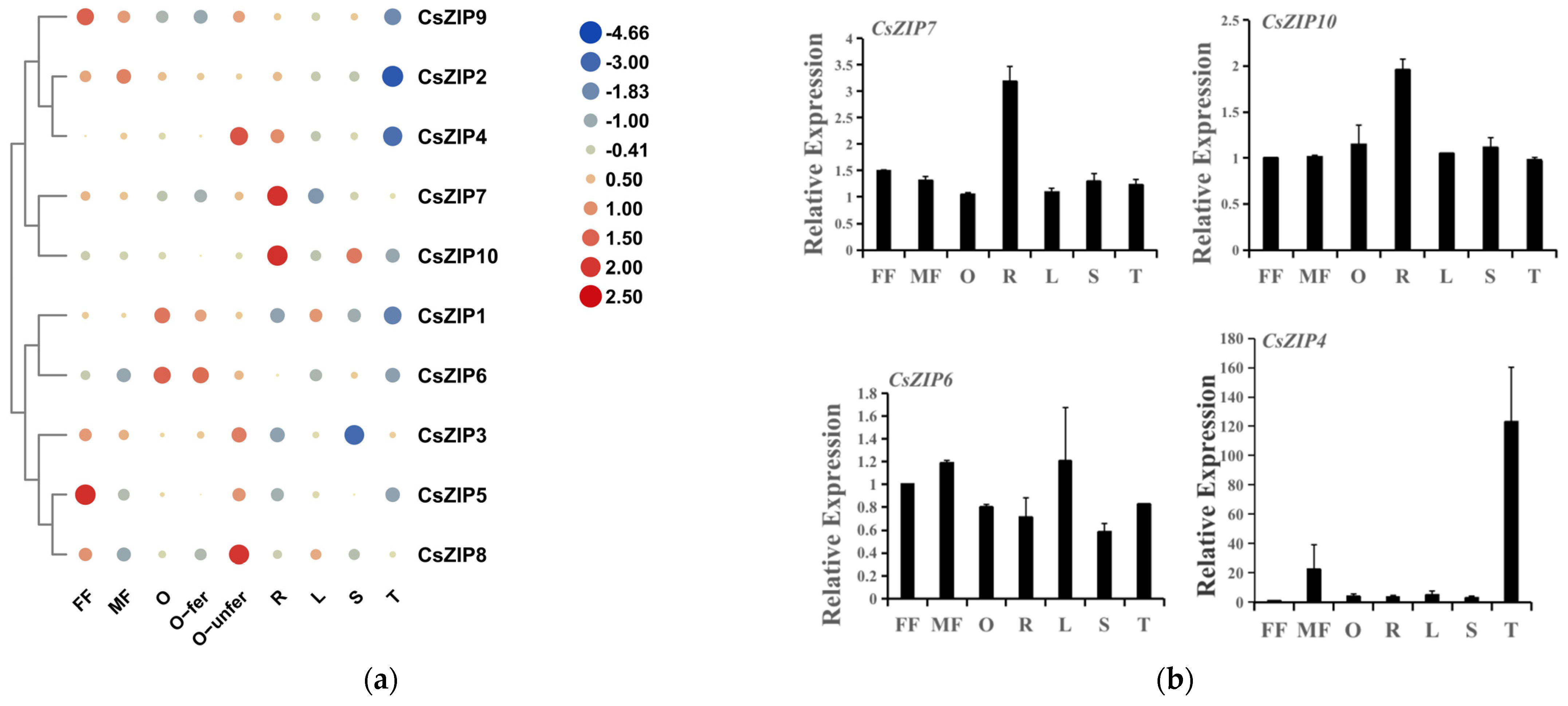
3.8. Expression of CsZIP Genes in Different Tissues
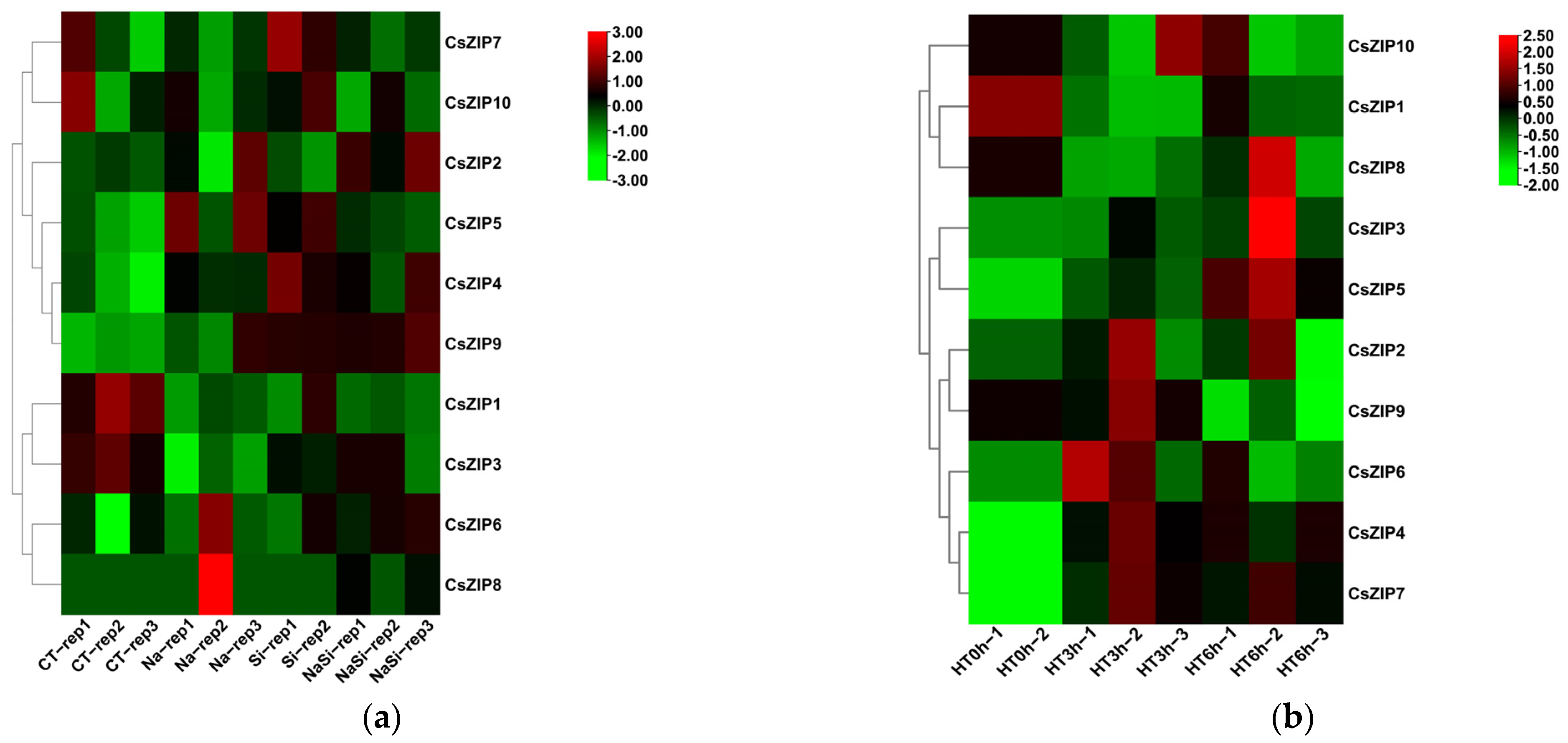
3.9. Expression Profiles of CsZIP Genes under Abiotic and Biotic Stresses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grotz, N.; Guerinot, M.L. Molecular aspects of Cu, Fe and Zn homeostasis in plants. Biochim. Biophys. Acta 2006, 1763, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Guerinot, M.L. The ZIP family of metal transporters. Biochim. Biophys. Acta 2000, 1465, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Gaither, L.A.; Eide, D.J. Eukaryotic zinc transporters and their regulation. Biometals 2001, 14, 251–270. [Google Scholar] [CrossRef]
- Nishida, S.; Tsuzuki, C.; Kato, A.; Aisu, A.; Yoshida, J.; Mizuno, T. AtIRT1, the primary iron uptake transporter in the root, mediates excess nickel accumulation in Arabidopsis thaliana. Plant Cell Physiol. 2011, 52, 1433–1442. [Google Scholar] [CrossRef] [PubMed]
- Krausko, M.; Labajová, M.; Peterková, D.; Jásik, J. Specific expression of AtIRT1 in phloem companion cells suggests its role in iron translocation in aboveground plant organs. Plant Signal. Behav. 2021, 16, 1925020. [Google Scholar] [CrossRef] [PubMed]
- Vert, G.; Barberon, M.; Zelazny, E.; Séguéla, M.; Briat, J.F.; Curie, C. Arabidopsis IRT2 cooperates with the high-affinity iron uptake system to maintain iron homeostasis in root epidermal cells. Planta 2009, 229, 1171–1179. [Google Scholar] [CrossRef]
- Milner, M.J.; Seamon, J.; Craft, E.; Kochian, L.V. Transport properties of members of the ZIP family in plants and their role in Zn and Mn homeostasis. J. Exp. Bot. 2013, 64, 369–381. [Google Scholar] [CrossRef]
- Gaitán-Solís, E.; Taylor, N.J.; Siritunga, D.; Stevens, W.; Schachtman, D.P. Overexpression of the transporters AtZIP1 and AtMTP1 in cassava changes zinc accumulation and partitioning. Front. Plant Sci. 2015, 6, 492. [Google Scholar] [CrossRef]
- Ramegowda, Y.; Venkategowda, R.; Jagadish, P. Expression of a rice Zn transporter, OsZIP1, increases Zn concentration in tobacco and finger millet transgenic plants. Plant Biotechnol. Rep. 2013, 7, 309–319. [Google Scholar] [CrossRef]
- Van de Mortel, J.E.; Almar Villanueva, L.; Schat, H.; Kwekkeboom, J.; Coughlan, S.; Moerland, P.D.; Ver Loren van Themaat, E.; Koornneef, M.; Aarts, M.G. Large expression differences in genes for iron and zinc homeostasis, stress response, and lignin biosynthesis distinguish roots of Arabidopsis thaliana and the related metal hyperaccumulator Thlaspi caerulescens. Plant Physiol. 2006, 142, 1127–1147. [Google Scholar] [CrossRef]
- Itai, R.N.; Ogo, Y.; Kobayashi, T.; Nakanishi, H.; Nishizawa, N.K. rice genes involved in phytosiderophore biosynthesis are synchronously regulated during the early stages of iron deficiency in roots. Rice 2013, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Ishimaru, Y.; Suzuki, M.; Tsukamoto, T.; Suzuki, K.; Nakazono, M.; Kobayashi, T.; Wada, Y.; Watanabe, S.; Matsuhashi, S.; Takahashi, M.; et al. Rice plants take up iron as an Fe3+-phytosiderophore and as Fe2+. Plant J. 2006, 45, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; An, G. Over-expression of OsIRT1 leads to increased iron and zinc accumulations in rice. Plant Cell Environ. 2009, 32, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, H.; Ogawa, I.; Ishimaru, Y.; Mori, S.; Nishizawa, N.K. Iron deficiency enhances cadmium uptake and translocation mediated by the Fe2+ transporters OsIRT1 and OsIRT2 in rice. Soil. Sci. Plant Nutr. 2006, 52, 464–469. [Google Scholar] [CrossRef]
- Ishimaru, Y.; Kim, S.; Tsukamoto, T.; Oki, H.; Kobayashi, T.; Watanabe, S.; Matsuhashi, S.; Takahashi, M.; Nakanishi, H.; Mori, S.; et al. Mutational reconstructed ferric chelate reductase confers enhanced tolerance in rice to iron deficiency in calcareous soil. Proc. Natl. Acad. Sci. USA 2007, 104, 7373–7378. [Google Scholar] [CrossRef]
- Liu, X.S.; Feng, S.J.; Zhang, B.Q.; Wang, M.Q.; Cao, H.W.; Rono, J.K.; Chen, X.; Yang, Z.M. OsZIP1 functions as a metal efflux transporter limiting excess zinc, copper and cadmium accumulation in rice. BMC Plant Biol. 2019, 19, 283. [Google Scholar] [CrossRef]
- Shimaru, Y.; Masuda, H.; Suzuki, M.; Bashir, K.; Takahashi, M.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Overexpression of the OsZIP4 zinc transporter confers disarrangement of zinc distribution in rice plants. J. Exp. Bot. 2007, 58, 2909–2915. [Google Scholar] [CrossRef]
- Lee, S.; Jeong, H.J.; Kim, S.A.; Lee, J.; Guerinot, M.L.; An, G. OsZIP5 is a plasma membrane zinc transporter in rice. Plant Mol. Biol. 2010, 73, 507–517. [Google Scholar] [CrossRef]
- Lee, S.; Kim, S.A.; Lee, J.; Guerinot, M.L.; An, G. Zinc deficiency-inducible OsZIP8 encodes a plasma membrane-localized zinc transporter in rice. Mol. Cells 2010, 29, 551–558. [Google Scholar] [CrossRef]
- Wu, X.; Zhu, Z.B.; Chen, J.H.; Huang, Y.F.; Liu, Z.L.; Zou, J.W.; Chen, Y.H.; Su, N.N.; Cui, J. Transcriptome analysis revealed pivotal transporters involved in the reduction of cadmium accumulation in pak choi (Brassica chinensis L.) by exogenous hydrogen-rich water. Chemosphere 2019, 216, 684–697. [Google Scholar] [CrossRef]
- Guo, J.; Li, K.; Zhang, X.; Huang, H.; Huang, F.; Zhang, L.; Wang, Y.; Li, T.; Yu, H. Genetic properties of cadmium translocation from straw to brown rice in low-grain cadmium rice line. Ecotoxicol. Environ. Saf. 2019, 182, 109422. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Li, D.; Du, X.; Xia, S.; Liu, C.; Shi, G. Comparative transcriptome analysis reveals key cadmium transport-related genes in roots of two pak choi (Brassica rapa L. ssp. chinensis) cultivars. BMC Genom. 2017, 18, 587. [Google Scholar] [CrossRef] [PubMed]
- Bienert, S.; Waterhouse, A.; Beer, T.A.; Tauriello, G.; Studer, G.; Bordoli, L.; Schwede, T. The SWISS-MODEL Repository-new features and functionality. Nucleic Acids Res. 2017, 45, D313–D319. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Liu, C.; Cao, B.; Qin, M.; Long, D.; Xiang, Z.; Zhao, A. Genome-Wide Identification and Characterization of Four Gene Families Putatively Involved in Cadmium Uptake, Translocation and Sequestration in Mulberry. Front. Plant Sci. 2018, 9, 879. [Google Scholar] [CrossRef]
- Palusińska, M.; Barabasz, A.; Kozak, K.; Papierniak, A.; Maślińska, K.; Antosiewicz, D.M. Zn/Cd status-dependent accumulation of Zn and Cd in root parts in tobacco is accompanied by specific expression of ZIP genes. BMC Plant Biol. 2020, 20, 37. [Google Scholar] [CrossRef] [PubMed]
- Alamholo, M.; Tarinejad, A. Molecular mechanism of drought stress tolerance in barley (Hordeum vulgare L.) via a combined analysis of the transcriptome data. Czech J. Genet. Plant Breed. 2023, 59, 76–94. [Google Scholar] [CrossRef]
- Wang, L.; Xu, Z.; Yin, W.; Xu, K.; Wang, S.; Shang, Q.; Sa, W.; Liang, J.; Wang, L. Genome-wide analysis of the Thaumatin-like gene family in Qingke (Hordeum vulgare L. var. nudum) uncovers candidates involved in plant defense against biotic and abiotic stresses. Front. Plant Sci. 2022, 13, 912296. [Google Scholar] [CrossRef]
- Zheng, S.; Dai, H.; Meng, Q.; Huang, R.; Tong, H.; Yuan, L. Identification and expression analysis of the ZRT, IRT-like protein (ZIP) gene family in Camellia sinensis (L.) O. Kuntze. Plant Physiol. Biochem. 2022, 172, 87–100. [Google Scholar] [CrossRef]
- Si, Z.; Qiao, Y.; Zhang, K.; Ji, Z.; Han, J. Characterization of Nucleotide Binding -Site-Encoding Genes in Sweetpotato, Ipomoea batatas (L.) Lam., and Their Response to Biotic and Abiotic Stresses. Cytogenet. Genome Res. 2021, 161, 257–271. [Google Scholar] [CrossRef]
- Song, X.; Li, E.; Song, H.; Du, G.; Li, S.; Zhu, H.; Chen, G.; Zhao, C.; Qiao, L.; Wang, J.; et al. Genome-wide identification and characterization of nonspecific lipid transfer protein (nsLTP) genes in Arachis duranensis. Genomics 2020, 112, 4332–4341. [Google Scholar] [CrossRef]
- Vert, G.; Grotz, N.; Dédaldéchamp, F.; Gaymard, F.; Guerinot, M.L.; Briat, J.F.; Curie, C. IRT1, an Arabidopsis transporter essential for iron uptake from the soil and for plant growth. Plant Cell 2002, 14, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Akther, M.S.; Das, U.; Tahura, S.; Prity, S.A.; Islam, M.; Kabir, A.H. Regulation of Zn uptake and redox status confers Zn deficiency tolerance in tomato. Sci. Hortic. 2020, 273, 109624. [Google Scholar] [CrossRef]
- Kabir, A.H.; Tahura, S.; Elseehy, M.M.; El-Shehawi, A.M. Molecular characterization of Fe-acquisition genes causing decreased Fe uptake and photosynthetic inefficiency in Fe-deficient sunflower. Sci. Rep. 2021, 11, 5537. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, C.; Duan, L.; Luan, Q.; Li, J.; Yang, A.; Qi, X.; Ren, Z. CsMYB60 is a key regulator of flavonols and proanthocyanidans that determine the colour of fruit spines in cucumber. J. Exp. Bot. 2019, 70, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Chou, K.C.; Shen, H.B. Cell-PLoc: A package of Web servers for predicting subcellular localization of proteins in various organisms. Nat. Protoc. 2008, 3, 153–162. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef]
- Li, L.; Stoeckert, C.J., Jr.; Roos, D.S. OrthoMCL: Identification of ortholog groups for eukaryotic genomes. Genome Res. 2003, 13, 2178–2189. [Google Scholar] [CrossRef]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef]
- Koch, M.A.; Haubold, B.; Mitchell-Olds, T. Comparative evolutionary analysis of chalcone synthase and alcohol dehydrogenase loci in Arabidopsis, Arabis, and related genera (Brassicaceae). Mol. Biol. Evol. 2000, 17, 1483–1498. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Li, H.; Wang, S.; Chai, S.; Yang, Z.; Zhang, Q.; Xin, H.; Xu, Y.; Lin, S.; Chen, X.; Yao, Z.; et al. Graph-based pan-genome reveals structural and sequence variations related to agronomic traits and domestication in cucumber. Nat. Commun. 2022, 13, 682. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, N.; Zhang, Z.; Shi, G. Genome-Wide Identification and Expression Profile Reveal Potential Roles of Peanut ZIP Family Genes in Zinc/Iron-Deficiency. Toler. Plants 2022, 11, 786. [Google Scholar] [CrossRef]
- Eng, B.H.; Guerinot, M.L.; Eide, D.; Saier, M.H., Jr. Sequence analyses and phylogenetic characterization of the ZIP family of metal ion transport proteins. J. Membr. Biol. 1998, 166, 1–7. [Google Scholar] [CrossRef]
- Fu, X.Z.; Zhou, X.; Xing, F.; Ling, L.L.; Chun, C.P.; Cao, L.; Aarts, M.G.M.; Peng, L.Z. Genome-Wide Identification, Cloning and Functional Analysis of the Zinc/Iron-Regulated Transporter-Like Protein (ZIP) Gene Family in Trifoliate Orange (Poncirus trifoliata L. Raf.). Front. Plant Sci. 2017, 8, 588. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, S.; Li, D. Genome-Wide Analysis of the ZRT, IRT-Like Protein (ZIP) Family and Their Responses to Metal Stress in Populus trichocarpa. Plant Mol. Biol. Rep. 2017, 35, 534–549. [Google Scholar] [CrossRef]
- Li, S.; Liu, X.; Zhou, X.; Li, Y.; Yang, W.; Chen, R. Improving Zinc and Iron Accumulation in Maize Grains Using the Zinc and Iron Transporter ZmZIP5. Plant Cell Physiol. 2019, 60, 2077–2085. [Google Scholar] [CrossRef] [PubMed]
- Cheong, J.J.; Choi, Y.D. Methyl jasmonate as a vital substance in plants. Trends Genet. 2003, 19, 409–413. [Google Scholar] [CrossRef]
- Castillo-Gonzalez, J.; Ojeda-Barrios, D.; Hernandez-Rodriguez, A.; Gonzalez-Franco, A.C.; Robles-Hernandez, L.; Lopez-Ochoa, G.R. ZINC METALLOENZYMES IN PLANTS. Interciencia 2018, 43, 242–248. [Google Scholar]
- Eriksson, S.; Böhlenius, H.; Moritz, T.; Nilsson, O. GA4 is the active gibberellin in the regulation of LEAFY transcription and Arabidopsis floral initiation. Plant Cell 2006, 18, 2172–2181. [Google Scholar] [CrossRef]
- Preciado-Rangel, P.; Reyes-Pérez, J.J.; Ramírez-Rodríguez, S.C.; Salas-Pérez, L.; Fortis-Hernández, M.; Murillo-Amador, B.; Troyo-Diéguez, E. Foliar Aspersion of Salicylic Acid Improves Phenolic and Flavonoid Compounds, and Also the Fruit Yield in Cucumber (Cucumis sativus L.). Plants 2019, 8, 44. [Google Scholar] [CrossRef] [PubMed]
- Pedas, P.; Schjoerring, J.K.; Husted, S. Identification and characterization of zinc-starvation-induced ZIP transporters from barley roots. Plant Physiol. Biochem. 2009, 47, 377–383. [Google Scholar] [CrossRef]
- Wintz, H.; Fox, T.; Wu, Y.Y.; Feng, V.; Chen, W.; Chang, H.S.; Zhu, T.; Vulpe, C. Expression profiles of Arabidopsis thaliana in mineral deficiencies reveal novel transporters involved in metal homeostasis. J. Biol. Chem. 2003, 278, 47644–47653. [Google Scholar] [CrossRef]
- Gong, Y.; Zhao, L.; Yan, F. Genome-Wide Identification and Expression Pattern Analysis of the Tomato ZIP Gene Family. Northeast. Agric. Sci. 2023, 48, 42–48+109. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Name | Accession Group | Accession Name | Accession Group |
|---|---|---|---|
| 9930 | East Asian cultivated accession | Hx117 | Indian cultivated accession |
| XTMC | East Asian cultivated accession | Hx14 | Indian cultivated accession |
| Cu2 | East Asian cultivated accession | W4 | Indian wild accession |
| Cuc37 | Eurasian cultivated accession | W8 | Indian wild accession |
| Gy14 | Eurasian cultivated accession | Cuc64 | Indian wild accession |
| 9110gt | Eurasian cultivated accession | PI183967 | Indian wild accession |
| Cuc80 | Xishuangbanna cultivated accession |
| Protein Name | 9930 | XTMC | Cu2 | Cuc80 | Cuc37 | Gy14 | 9110gt | PI183967 | Cuc64 | W4 | W8 | Hx14 | Hx117 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CsZIP1 | 249 | 350 | 350 | - | 350 | 350 | 350 | 350 | 350 | 350 | 350 | 350 | 350 |
| CsZIP2 | 417 | 417 | 417 | 417 | 417 | 441 | 417 | 417 | 417 | 417 | 417 | 408 | 417 |
| CsZIP3 | 228 | 275 | 275 | 275 | 275 | 306 | 275 | 275 | 306 | 275 | 275 | 275 | 275 |
| CsZIP4 | 594 | 594 | 639 | 594 | 594 | 594 | 594 | 594 | 594 | 594 | 594 | 594 | 594 |
| CsZIP5 | 367 | 367 | 367 | 365 | 367 | 367 | 367 | 367 | 369 | 367 | 365 | 369 | 815 |
| CsZIP6 | 463 | 463 | 463 | 463 | 463 | 407 | 463 | 463 | 463 | 463 | 463 | 463 | 463 |
| CsZIP7 | 335 | 335 | 335 | 335 | 335 | 335 | 335 | 335 | 335 | 302 | 335 | 335 | 346 |
| CsZIP8 | 349 | 349 | 349 | 349 | 349 | 349 | 349 | 349 | 349 | 349 | 349 | 349 | 1096 |
| CsZIP9 | 334 | 334 | 334 | 334 | 334 | 337 | 334 | - | 338 | 334 | 334 | 334 | 337 |
| CsZIP10 | 354 | 354 | 354 | 354 | 354 | 354 | 354 | 354 | 354 | 354 | 354 | 354 | 354 |
| Gene | Introns | Chromosomal Site | Gene Length | Protein Length | Protein MW (kDa) | Isoelectric Point | Subcellular Location Predicted |
|---|---|---|---|---|---|---|---|
| CsZIP1 | 2 | 1 | 2955 | 249 | 27.2 | 6.88 | Cell membrane |
| CsZIP2 | 3 | 2 | 3491 | 417 | 44.5 | 6.09 | Chloroplast |
| CsZIP3 | 9 | 4 | 9406 | 228 | 24.6 | 7.02 | Cell membrane |
| CsZIP4 | 4 | 4 | 5063 | 594 | 62.0 | 6.27 | Cell membrane |
| CsZIP5 | 2 | 4 | 2777 | 367 | 38.8 | 5.98 | Cell membrane |
| CsZIP6 | 10 | 5 | 21,173 | 463 | 49.9 | 5.97 | Cell membrane |
| CsZIP7 | 1 | 6 | 2038 | 334 | 35.7 | 5.73 | Chloroplast |
| CsZIP8 | 0 | 6 | 1490 | 348 | 38.0 | 5.58 | Cell membrane |
| CsZIP9 | 2 | 7 | 7007 | 334 | 35.9 | 6.02 | Cell membrane |
| CsZIP10 | 2 | 7 | 4076 | 354 | 37.2 | 8.53 | Cell membrane |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Yin, M.; Han, J.; Wang, X.; Chang, J.; Ren, Z.; Wang, L. Pan-Genome-Wide Identification and Transcriptome-Wide Analysis of ZIP Genes in Cucumber. Agriculture 2024, 14, 133. https://doi.org/10.3390/agriculture14010133
Wang Z, Yin M, Han J, Wang X, Chang J, Ren Z, Wang L. Pan-Genome-Wide Identification and Transcriptome-Wide Analysis of ZIP Genes in Cucumber. Agriculture. 2024; 14(1):133. https://doi.org/10.3390/agriculture14010133
Chicago/Turabian StyleWang, Zimo, Mengmeng Yin, Jing Han, Xuehua Wang, Jingshu Chang, Zhonghai Ren, and Lina Wang. 2024. "Pan-Genome-Wide Identification and Transcriptome-Wide Analysis of ZIP Genes in Cucumber" Agriculture 14, no. 1: 133. https://doi.org/10.3390/agriculture14010133
APA StyleWang, Z., Yin, M., Han, J., Wang, X., Chang, J., Ren, Z., & Wang, L. (2024). Pan-Genome-Wide Identification and Transcriptome-Wide Analysis of ZIP Genes in Cucumber. Agriculture, 14(1), 133. https://doi.org/10.3390/agriculture14010133