
Genome-Wide Identification and Expression Analysis of the CesA/Csl Gene Superfamily in Alfalfa (Medicago sativa L.)

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material Preparation
2.2. Identification and Characterization of Cellulose Synthase Genes
2.3. Phylogenetic, Chromosomal Location and Synteny Analysis
2.4. Gene Structure and Motif Composition Analysis
2.5. Promoter Cis-Acting Element Analysis
2.6. Transcriptome Data Analysis
2.7. qRT-PCR
3. Results
3.1. Basic Information on the CesA/Csl Genes in Alfalfa
3.2. Phylogenetic Analysis
3.3. Motifs and Gene Structure
3.4. Promoter Analysis
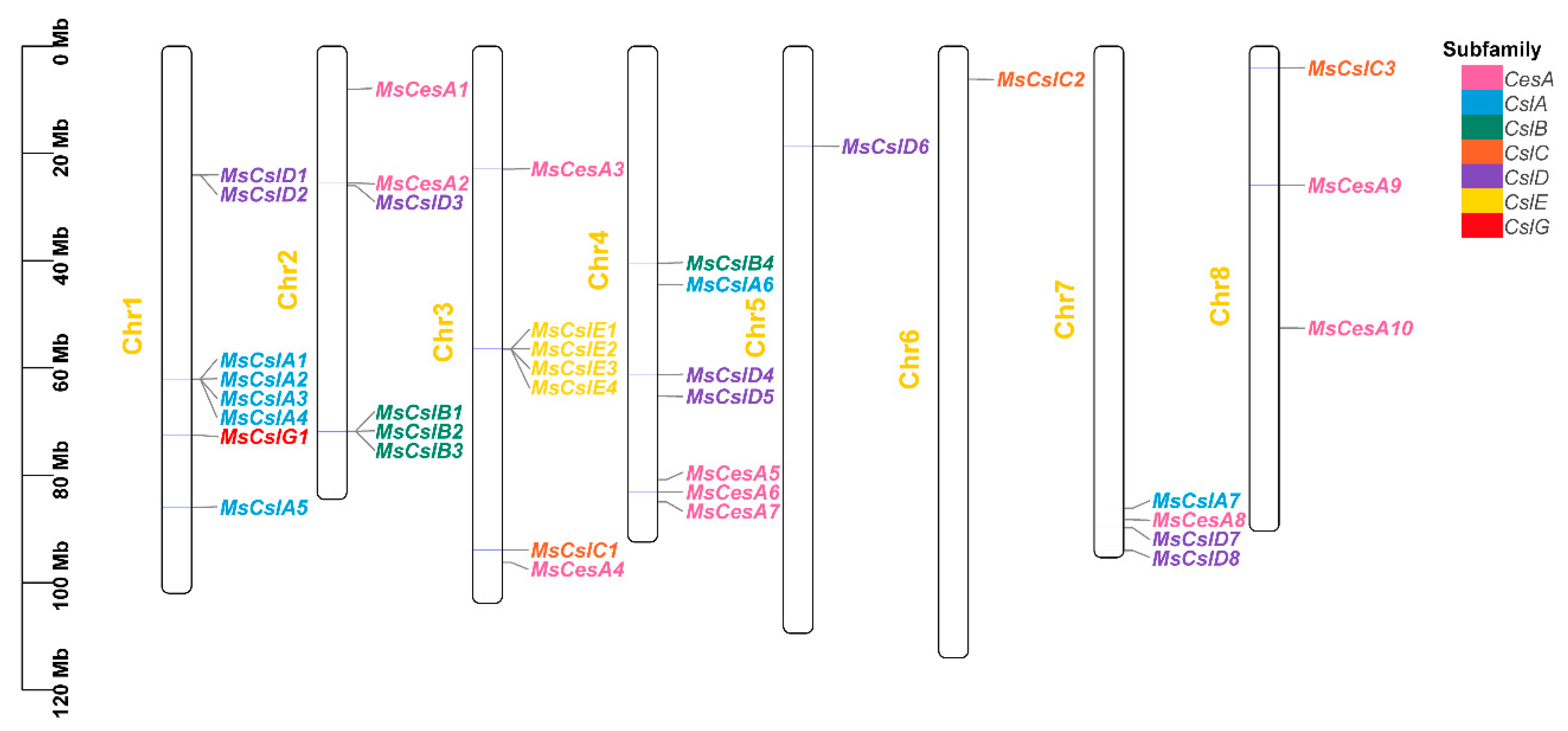
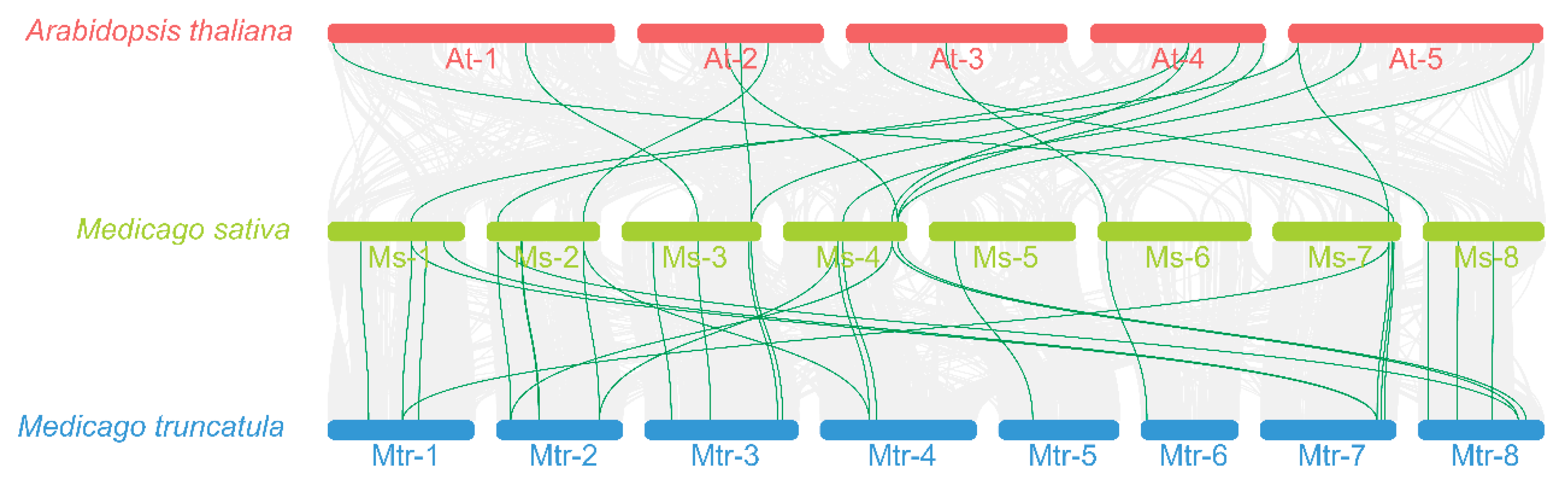
3.5. Collinearity Analysis
3.6. Analysis of the Expression of CesA/Csl Genes in Various Tissues
3.7. Expression Analysis of CesA/Csls under Abiotic Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, B.; Gao, Y.; Zhang, L.; Zhou, Y. The Plant Cell Wall: Biosynthesis, Construction, and Functions. J. Integr. Plant Biol. 2021, 63, 251–272. [Google Scholar] [CrossRef] [PubMed]
- Little, A.; Schwerdt, J.G.; Shirley, N.J.; Khor, S.F.; Neumann, K.; O’Donovan, L.A.; Lahnstein, J.; Collins, H.M.; Henderson, M.; Fincher, G.B.; et al. Revised Phylogeny of the Cellulose Synthase Gene Superfamily: Insights into Cell Wall Evolution. Plant Physiol. 2018, 177, 1124–1141. [Google Scholar] [CrossRef] [PubMed]
- Turner, S.; Kumar, M. Cellulose Synthase Complex Organization and Cellulose Microfibril Structure. Philos. Trans. A Math. Phys. Eng. Sci. 2018, 376, 20170048. [Google Scholar] [CrossRef] [PubMed]
- Polko, J.K.; Kieber, J.J. The Regulation of Cellulose Biosynthesis in Plants. Plant Cell 2019, 31, 282–296. [Google Scholar] [CrossRef] [PubMed]
- Rongpipi, S.; Ye, D.; Gomez, E.D.; Gomez, E.W. Progress and Opportunities in the Characterization of Cellulose—An Important Regulator of Cell Wall Growth and Mechanics. Front. Plant Sci. 2018, 9, 1894. [Google Scholar] [CrossRef] [PubMed]
- Pear, J.R.; Kawagoe, Y.; Schreckengost, W.E.; Delmer, D.P.; Stalker, D.M. Higher Plants Contain Homologs of the Bacterial CelA Genes Encoding the Catalytic Subunit of Cellulose Synthase. Proc. Natl. Acad. Sci. USA 1996, 93, 12637–12642. [Google Scholar] [CrossRef] [PubMed]
- Arioli, T.; Peng, L.; Betzner, A.S.; Burn, J.; Wittke, W.; Herth, W.; Camilleri, C.; Höfte, H.; Plazinski, J.; Birch, R.; et al. Molecular Analysis of Cellulose Biosynthesis in Arabidopsis. Science 1998, 279, 717–720. [Google Scholar] [CrossRef]
- Popper, Z.A.; Michel, G.; Hervé, C.; Domozych, D.S.; Willats, W.G.T.; Tuohy, M.G.; Kloareg, B.; Stengel, D.B. Evolution and Diversity of Plant Cell Walls: From Algae to Flowering Plants. Annu. Rev. Plant Biol. 2011, 62, 567–590. [Google Scholar] [CrossRef]
- Nixon, B.T.; Mansouri, K.; Singh, A.; Du, J.; Davis, J.K.; Lee, J.-G.; Slabaugh, E.; Vandavasi, V.G.; O’Neill, H.; Roberts, E.M.; et al. Comparative Structural and Computational Analysis Supports Eighteen Cellulose Synthases in the Plant Cellulose Synthesis Complex. Sci. Rep. 2016, 6, 28696. [Google Scholar] [CrossRef]
- Duncombe, S.G.; Chethan, S.G.; Anderson, C.T. Super-Resolution Imaging Illuminates New Dynamic Behaviors of Cellulose Synthase. Plant Cell 2022, 34, 273–286. [Google Scholar] [CrossRef]
- Kurek, I.; Kawagoe, Y.; Jacob-Wilk, D.; Doblin, M.; Delmer, D. Dimerization of Cotton Fiber Cellulose Synthase Catalytic Subunits Occurs via Oxidation of the Zinc-Binding Domains. Proc. Natl. Acad. Sci. USA 2002, 99, 11109–11114. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Dhugga, K.S.; Gill, K.; Singh, J. Novel Structural and Functional Motifs in Cellulose Synthase (CesA) Genes of Bread Wheat (Triticum Aestivum, L.). PLoS ONE 2016, 11, e0147046. [Google Scholar] [CrossRef] [PubMed]
- Pancaldi, F.; van Loo, E.N.; Schranz, M.E.; Trindade, L.M. Genomic Architecture and Evolution of the Cellulose Synthase Gene Superfamily as Revealed by Phylogenomic Analysis. Front. Plant Sci. 2022, 13, 870818. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.G.; Howells, R.M.; Huttly, A.K.; Vickers, K.; Turner, S.R. Interactions among Three Distinct CesA Proteins Essential for Cellulose Synthesis. Proc. Natl. Acad. Sci. USA 2003, 100, 1450–1455. [Google Scholar] [CrossRef]
- Song, X.; Xu, L.; Yu, J.; Tian, P.; Hu, X.; Wang, Q.; Pan, Y. Genome-Wide Characterization of the Cellulose Synthase Gene Superfamily in Solanum Lycopersicum. Gene 2019, 688, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Desprez, T.; Juraniec, M.; Crowell, E.F.; Jouy, H.; Pochylova, Z.; Parcy, F.; Höfte, H.; Gonneau, M.; Vernhettes, S. Organization of Cellulose Synthase Complexes Involved in Primary Cell Wall Synthesis in Arabidopsis Thaliana. Proc. Natl. Acad. Sci. USA 2007, 104, 15572–15577. [Google Scholar] [CrossRef]
- Persson, S.; Paredez, A.; Carroll, A.; Palsdottir, H.; Doblin, M.; Poindexter, P.; Khitrov, N.; Auer, M.; Somerville, C.R. Genetic Evidence for Three Unique Components in Primary Cell-Wall Cellulose Synthase Complexes in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 15566–15571. [Google Scholar] [CrossRef]
- Yin, Y.; Johns, M.A.; Cao, H.; Rupani, M. A Survey of Plant and Algal Genomes and Transcriptomes Reveals New Insights into the Evolution and Function of the Cellulose Synthase Superfamily. BMC Genom. 2014, 15, 260. [Google Scholar] [CrossRef]
- Marcotuli, I.; Colasuonno, P.; Blanco, A.; Gadaleta, A. Expression Analysis of Cellulose Synthase-like Genes in Durum Wheat. Sci. Rep. 2018, 8, 15675. [Google Scholar] [CrossRef]
- Farrokhi, N.; Burton, R.A.; Brownfield, L.; Hrmova, M.; Wilson, S.M.; Bacic, A.; Fincher, G.B. Plant Cell Wall Biosynthesis: Genetic, Biochemical and Functional Genomics Approaches to the Identification of Key Genes. Plant Biotechnol. J. 2006, 4, 145–167. [Google Scholar] [CrossRef]
- Keegstra, K.; Walton, J. β-Glucans—Brewer’s Bane, Dietician’s Delight. Science 2006, 311, 1872–1873. [Google Scholar] [CrossRef] [PubMed]
- Richmond, T.A.; Somerville, C.R. The Cellulose Synthase Superfamily. Plant Physiol. 2000, 124, 495–498. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Guo, K.; Li, Y.; Tu, Y.; Hu, H.; Wang, B.; Cui, X.; Peng, L. Expression Profiling and Integrative Analysis of the CESA/CSL Superfamily in Rice. BMC Plant Biol. 2010, 10, 282. [Google Scholar] [CrossRef] [PubMed]
- Suwignyo, B.; Aristia Rini, E.; Helmiyati, S. The Profile of Tropical Alfalfa in Indonesia: A Review. Saudi J. Biol. Sci. 2023, 30, 103504. [Google Scholar] [CrossRef]
- Oba, M.; Allen, M.S. Evaluation of the Importance of the Digestibility of Neutral Detergent Fiber from Forage: Effects on Dry Matter Intake and Milk Yield of Dairy Cows. J. Dairy Sci. 1999, 82, 589–596. [Google Scholar] [CrossRef]
- Lin, S.; Medina, C.A.; Norberg, O.S.; Combs, D.; Wang, G.; Shewmaker, G.; Fransen, S.; Llewellyn, D.; Yu, L.-X. Genome-Wide Association Studies Identifying Multiple Loci Associated with Alfalfa Forage Quality. Front. Plant Sci. 2021, 12, 648192. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Du, H.; Chen, Z.; Lu, H.; Zhu, F.; Chen, H.; Meng, X.; Liu, Q.; Liu, P.; Zheng, L.; et al. The Chromosome-Level Genome Sequence of the Autotetraploid Alfalfa and Resequencing of Core Germplasms Provide Genomic Resources for Alfalfa Research. Mol. Plant 2020, 13, 1250–1261. [Google Scholar] [CrossRef]
- Eddy, S.R. Accelerated Profile HMM Searches. PLoS Comput. Biol. 2011, 7, e1002195. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.-Y. Ggtree: An r Package for Visualization and Annotation of Phylogenetic Trees with Their Covariates and Other Associated Data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- O’Rourke, J.A.; Fu, F.; Bucciarelli, B.; Yang, S.S.; Samac, D.A.; Lamb, J.F.S.; Monteros, M.J.; Graham, M.A.; Gronwald, J.W.; Krom, N.; et al. The Medicago Sativa Gene Index 1.2: A Web-Accessible Gene Expression Atlas for Investigating Expression Differences between Medicago Sativa Subspecies. BMC Genom. 2015, 16, 502. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Deng, H.; Ma, W.; Zhou, Q.; Liu, Z. Genome-Wide Identification of the MADS-Box Transcription Factor Family in Autotetraploid Cultivated Alfalfa (Medicago Sativa L.) and Expression Analysis under Abiotic Stress. BMC Genom. 2021, 22, 603. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-Based Genome Alignment and Genotyping with HISAT2 and HISAT-Genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ouyang, S.; Zhu, W.; Hamilton, J.; Lin, H.; Campbell, M.; Childs, K.; Thibaud-Nissen, F.; Malek, R.L.; Lee, Y.; Zheng, L.; et al. The TIGR Rice Genome Annotation Resource: Improvements and New Features. Nucleic Acids Res. 2007, 35, D883–D887. [Google Scholar] [CrossRef]
- Kaur, S.; Dhugga, K.S.; Beech, R.; Singh, J. Genome-Wide Analysis of the Cellulose Synthase-like (Csl) Gene Family in Bread Wheat (Triticum Aestivum L.). BMC Plant Biol. 2017, 17, 193. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yang, T.; Dai, D.; Hu, Y.; Guo, X.; Guo, H. Evolution, Gene Expression Profiling and 3D Modeling of CSLD Proteins in Cotton. BMC Plant Biol. 2017, 17, 119. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.A.; Rehman, H.M.; Baloch, F.S.; Ijaz, B.; Ali, M.A.; Khan, I.A.; Lee, J.D.; Chung, G.; Yang, S.H. Genome and Transcriptome-Wide Analyses of Cellulose Synthase Gene Superfamily in Soybean. J. Plant Physiol. 2017, 215, 163–175. [Google Scholar] [CrossRef]
- Ermawar, R.A.; Collins, H.M.; Byrt, C.S.; Henderson, M.; O’Donovan, L.A.; Shirley, N.J.; Schwerdt, J.G.; Lahnstein, J.; Fincher, G.B.; Burton, R.A. Genetics and Physiology of Cell Wall Polysaccharides in the Model C4 Grass, Setaria Viridis spp. BMC Plant Biol. 2015, 15, 236. [Google Scholar] [CrossRef]
- Yin, Y.; Huang, J.; Xu, Y. The Cellulose Synthase Superfamily in Fully Sequenced Plants and Algae. BMC Plant Biol. 2009, 9, 99. [Google Scholar] [CrossRef] [PubMed]
- Bulone, V.; Schwerdt, J.G.; Fincher, G.B. Co-Evolution of Enzymes Involved in Plant Cell Wall Metabolism in the Grasses. Front. Plant Sci. 2019, 10, 1009. [Google Scholar] [CrossRef]
- Betts, M.J.; Guigó, R.; Agarwal, P.; Russell, R.B. Exon Structure Conservation despite Low Sequence Similarity: A Relic of Dramatic Events in Evolution? EMBO J. 2001, 20, 5354–5360. [Google Scholar] [CrossRef]
- Carmel, L.; Rogozin, I.B.; Wolf, Y.I.; Koonin, E.V. Evolutionarily Conserved Genes Preferentially Accumulate Introns. Genome Res. 2007, 17, 1045–1050. [Google Scholar] [CrossRef]
- Li, G.; Liu, X.; Liang, Y.; Zhang, Y.; Cheng, X.; Cai, Y. Genome-Wide Characterization of the Cellulose Synthase Gene Superfamily in Pyrus Bretschneideri and Reveal Its Potential Role in Stone Cell Formation. Funct. Integr. Genom. 2020, 20, 723–738. [Google Scholar] [CrossRef]
- Daras, G.; Templalexis, D.; Avgeri, F.; Tsitsekian, D.; Karamanou, K.; Rigas, S. Updating Insights into the Catalytic Domain Properties of Plant Cellulose Synthase (CesA) and Cellulose Synthase-like (Csl) Proteins. Molecules 2021, 26, 4335. [Google Scholar] [CrossRef]
- Wang, J.; Li, J.; Lin, W.; Deng, B.; Lin, L.; Lv, X.; Hu, Q.; Liu, K.; Fatima, M.; He, B.; et al. Genome-Wide Identification and Adaptive Evolution of CesA/Csl Superfamily among Species with Different Life Forms in Orchidaceae. Front. Plant Sci. 2022, 13, 994679. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Garcia, C.M.; Finer, J.J. Identification and Validation of Promoters and Cis-Acting Regulatory Elements. Plant Sci. 2014, 217–218, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Favery, B.; Ryan, E.; Foreman, J.; Linstead, P.; Boudonck, K.; Steer, M.; Shaw, P.; Dolan, L. KOJAK Encodes a Cellulose Synthase-like Protein Required for Root Hair Cell Morphogenesis in Arabidopsis. Genes Dev. 2001, 15, 79–89. [Google Scholar] [CrossRef]
- Wang, X.; Cnops, G.; Vanderhaeghen, R.; De Block, S.; Van Montagu, M.; Van Lijsebettens, M. AtCSLD3, a Cellulose Synthase-like Gene Important for Root Hair Growth in Arabidopsis. Plant Physiol. 2001, 126, 575–586. [Google Scholar] [CrossRef] [PubMed]
- Bernal, A.J.; Jensen, J.K.; Harholt, J.; Sørensen, S.; Moller, I.; Blaukopf, C.; Johansen, B.; de Lotto, R.; Pauly, M.; Scheller, H.V.; et al. Disruption of ATCSLD5 Results in Reduced Growth, Reduced Xylan and Homogalacturonan Synthase Activity and Altered Xylan Occurrence in Arabidopsis. Plant J. 2007, 52, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.M.; Park, S.H.; Je, B.I.; Park, S.H.; Park, S.J.; Piao, H.L.; Eun, M.Y.; Dolan, L.; Han, C. OsCSLD1, a Cellulose Synthase-like D1 Gene, Is Required for Root Hair Morphogenesis in Rice. Plant Physiol. 2007, 143, 1220–1230. [Google Scholar] [CrossRef]
- Zhu, J.; Lee, B.-H.; Dellinger, M.; Cui, X.; Zhang, C.; Wu, S.; Nothnagel, E.A.; Zhu, J.-K. A Cellulose Synthase-like Protein Is Required for Osmotic Stress Tolerance in Arabidopsis. Plant J. 2010, 63, 128–140. [Google Scholar] [CrossRef]
- Zhao, H.; Li, Z.; Wang, Y.; Wang, J.; Xiao, M.; Liu, H.; Quan, R.; Zhang, H.; Huang, R.; Zhu, L.; et al. Cellulose Synthase-like Protein OsCSLD4 Plays an Important Role in the Response of Rice to Salt Stress by Mediating Abscisic Acid Biosynthesis to Regulate Osmotic Stress Tolerance. Plant Biotechnol. J. 2022, 20, 468–484. [Google Scholar] [CrossRef]
- Li, S.; Zhang, L.; Wang, Y.; Xu, F.; Liu, M.; Lin, P.; Ren, S.; Ma, R.; Guo, Y.-D. Knockdown of a Cellulose Synthase Gene BoiCesA Affects the Leaf Anatomy, Cellulose Content and Salt Tolerance in Broccoli. Sci. Rep. 2017, 7, 41397. [Google Scholar] [CrossRef]
- Ye, Y.; Wang, S.; Wu, K.; Ren, Y.; Jiang, H.; Chen, J.; Tao, L.; Fu, X.; Liu, B.; Wu, Y. A Semi-Dominant Mutation in OsCESA9 Improves Salt Tolerance and Favors Field Straw Decay Traits by Altering Cell Wall Properties in Rice. Rice 2021, 14, 19. [Google Scholar] [CrossRef]
- Cao, H.; Guo, S.; Xu, Y.; Jiang, K.; Jones, A.M.; Chong, K. Reduced Expression of a Gene Encoding a Golgi Localized Monosaccharide Transporter (OsGMST1) Confers Hypersensitivity to Salt in Rice (Oryza Sativa). J. Exp. Bot. 2011, 62, 4595–4604. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Gene Rename | Gene Accession | Chromosome | Location | Amino Acid | MW (kDa) | pI | |
|---|---|---|---|---|---|---|---|---|
| Start | End | |||||||
| 1 | MsCslD1 | MsG0180001606.01 | Chr1 | 24,040,717 | 24,045,076 | 1146 | 128.34 | 5.82 |
| 2 | MsCslD2 | MsG0180001607.01 | Chr1 | 24,051,752 | 24,056,169 | 1137 | 127.31 | 8.22 |
| 3 | MsCslA1 | MsG0180003417.01 | Chr1 | 62,051,942 | 62,059,793 | 488 | 56.27 | 8.79 |
| 4 | MsCslA2 | MsG0180003418.01 | Chr1 | 62,061,668 | 62,066,995 | 440 | 50.81 | 9.08 |
| 5 | MsCslA3 | MsG0180003419.01 | Chr1 | 62,073,889 | 62,083,620 | 538 | 62.18 | 8.78 |
| 6 | MsCslA4 | MsG0180003420.01 | Chr1 | 62,091,072 | 62,096,754 | 492 | 56.87 | 9.39 |
| 7 | MsCslG1 | MsG0180004053.01 | Chr1 | 72,523,965 | 72,527,724 | 711 | 81.75 | 6.82 |
| 8 | MsCslA5 | MsG0180005031.01 | Chr1 | 85,959,191 | 85,963,009 | 530 | 60.97 | 8.89 |
| 9 | MsCesA1 | MsG0280006922.01 | Chr2 | 7,998,792 | 8,011,529 | 1572 | 175.18 | 6.02 |
| 10 | MsCesA2 | MsG0280008107.01 | Chr2 | 25,539,931 | 25,545,645 | 1044 | 118.98 | 7.20 |
| 11 | MsCslD3 | MsG0280008146.01 | Chr2 | 26,016,637 | 26,025,349 | 1178 | 133.37 | 8.13 |
| 12 | MsCslB1 | MsG0280010585.01 | Chr2 | 71,770,428 | 71,788,821 | 371 | 41.98 | 8.18 |
| 13 | MsCslB2 | MsG0280010586.01 | Chr2 | 71,792,138 | 71,802,870 | 498 | 55.55 | 8.02 |
| 14 | MsCslB3 | MsG0280010587.01 | Chr2 | 71,809,645 | 71,814,024 | 698 | 79.32 | 7.94 |
| 15 | MsCesA3 | MsG0380012697.01 | Chr3 | 22,927,630 | 22,933,990 | 774 | 87.01 | 6.09 |
| 16 | MsCslE1 | MsG0380014552.01 | Chr3 | 56,426,859 | 56,438,397 | 833 | 96.37 | 6.73 |
| 17 | MsCslE2 | MsG0380014553.01 | Chr3 | 56,444,913 | 56,453,340 | 419 | 47.72 | 8.65 |
| 18 | MsCslE3 | MsG0380014558.01 | Chr3 | 56,484,562 | 56,490,490 | 595 | 68.10 | 8.54 |
| 19 | MsCslE4 | MsG0380014561.01 | Chr3 | 56,507,839 | 56,510,348 | 476 | 53.93 | 6.05 |
| 20 | MsCslC1 | MsG0380017240.01 | Chr3 | 93,932,951 | 93,936,593 | 702 | 80.35 | 8.03 |
| 21 | MsCesA4 | MsG0380017435.01 | Chr3 | 96,225,074 | 96,231,632 | 1089 | 122.97 | 6.56 |
| 22 | MsCslB4 | MsG0480020393.01 | Chr4 | 40,435,695 | 40,441,148 | 380 | 43.20 | 8.72 |
| 23 | MsCslA6 | MsG0480020597.01 | Chr4 | 44,402,146 | 44,408,477 | 542 | 62.53 | 9.07 |
| 24 | MsCslD4 | MsG0480021653.01 | Chr4 | 61,289,304 | 61,292,141 | 719 | 81.45 | 8.86 |
| 25 | MsCslD5 | MsG0480021859.01 | Chr4 | 65,279,346 | 65,280,787 | 454 | 51.10 | 8.34 |
| 26 | MsCesA5 | MsG0480023097.01 | Chr4 | 80,793,999 | 80,812,063 | 1362 | 151.46 | 5.64 |
| 27 | MsCesA6 | MsG0480023279.01 | Chr4 | 83,142,696 | 83,149,308 | 1098 | 124.09 | 6.37 |
| 28 | MsCesA7 | MsG0480023418.01 | Chr4 | 84,957,277 | 84,963,896 | 1098 | 124.08 | 6.37 |
| 29 | MsCslD6 | MsG0580025441.01 | Chr5 | 18,677,897 | 18,682,489 | 1122 | 126.02 | 5.8 |
| 30 | MsCslC2 | MsG0680030621.01 | Chr6 | 6,190,384 | 6,194,874 | 641 | 74.24 | 8.95 |
| 31 | MsCslA7 | MsG0780041074.01 | Chr7 | 86,131,839 | 86,141,384 | 1154 | 130.89 | 8.59 |
| 32 | MsCesA8 | MsG0780041211.01 | Chr7 | 88,265,473 | 88,272,679 | 1049 | 118.17 | 8.00 |
| 33 | MsCslD7 | MsG0780041340.01 | Chr7 | 89,777,194 | 89,781,609 | 1171 | 130.94 | 8.22 |
| 34 | MsCslD8 | MsG0780041693.01 | Chr7 | 94,013,062 | 94,017,095 | 1050 | 118.13 | 7.91 |
| 35 | MsCslC3 | MsG0880042124.01 | Chr8 | 4,096,083 | 4,102,789 | 698 | 80.08 | 8.92 |
| 36 | MsCesA9 | MsG0880043508.01 | Chr8 | 25,944,537 | 25,952,543 | 1075 | 120.77 | 7.69 |
| 37 | MsCesA10 | MsG0880044994.01 | Chr8 | 52,543,467 | 52,549,606 | 1002 | 114.28 | 8.92 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sod, B.; Xu, L.; Liu, Y.; He, F.; Xu, Y.; Li, M.; Yang, T.; Gao, T.; Kang, J.; Yang, Q.; et al. Genome-Wide Identification and Expression Analysis of the CesA/Csl Gene Superfamily in Alfalfa (Medicago sativa L.). Agriculture 2023, 13, 1658. https://doi.org/10.3390/agriculture13091658
Sod B, Xu L, Liu Y, He F, Xu Y, Li M, Yang T, Gao T, Kang J, Yang Q, et al. Genome-Wide Identification and Expression Analysis of the CesA/Csl Gene Superfamily in Alfalfa (Medicago sativa L.). Agriculture. 2023; 13(9):1658. https://doi.org/10.3390/agriculture13091658
Chicago/Turabian StyleSod, Bilig, Lei Xu, Yajiao Liu, Fei He, Yanchao Xu, Mingna Li, Tianhui Yang, Ting Gao, Junmei Kang, Qingchuan Yang, and et al. 2023. "Genome-Wide Identification and Expression Analysis of the CesA/Csl Gene Superfamily in Alfalfa (Medicago sativa L.)" Agriculture 13, no. 9: 1658. https://doi.org/10.3390/agriculture13091658
APA StyleSod, B., Xu, L., Liu, Y., He, F., Xu, Y., Li, M., Yang, T., Gao, T., Kang, J., Yang, Q., & Long, R. (2023). Genome-Wide Identification and Expression Analysis of the CesA/Csl Gene Superfamily in Alfalfa (Medicago sativa L.). Agriculture, 13(9), 1658. https://doi.org/10.3390/agriculture13091658