Melatonin Secretion in Regulating the Circadian Rhythms of Reproduction in Goose (Anser cygnoides)


 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Design and Sample Collection
2.3. Reproductive Performance and Immunohistochemical Observations
2.4. RNA Extraction and Quantitative RT-PCR
2.5. Measurement of Serum Melatonin Hormone Concentrations
2.6. Statistical Analysis
3. Results
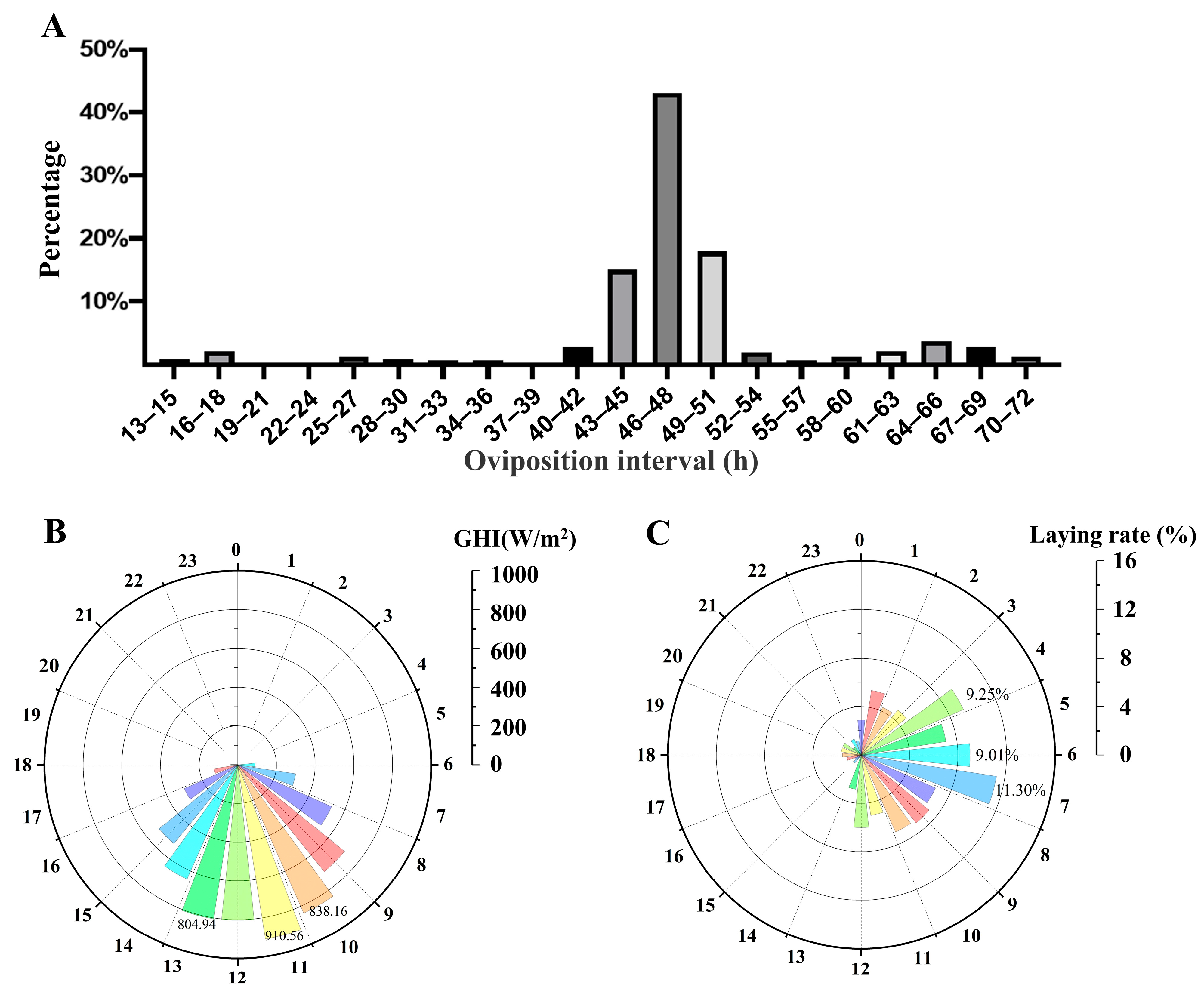
3.1. The Temporal Dynamics of Egg-Laying Preference in Goose
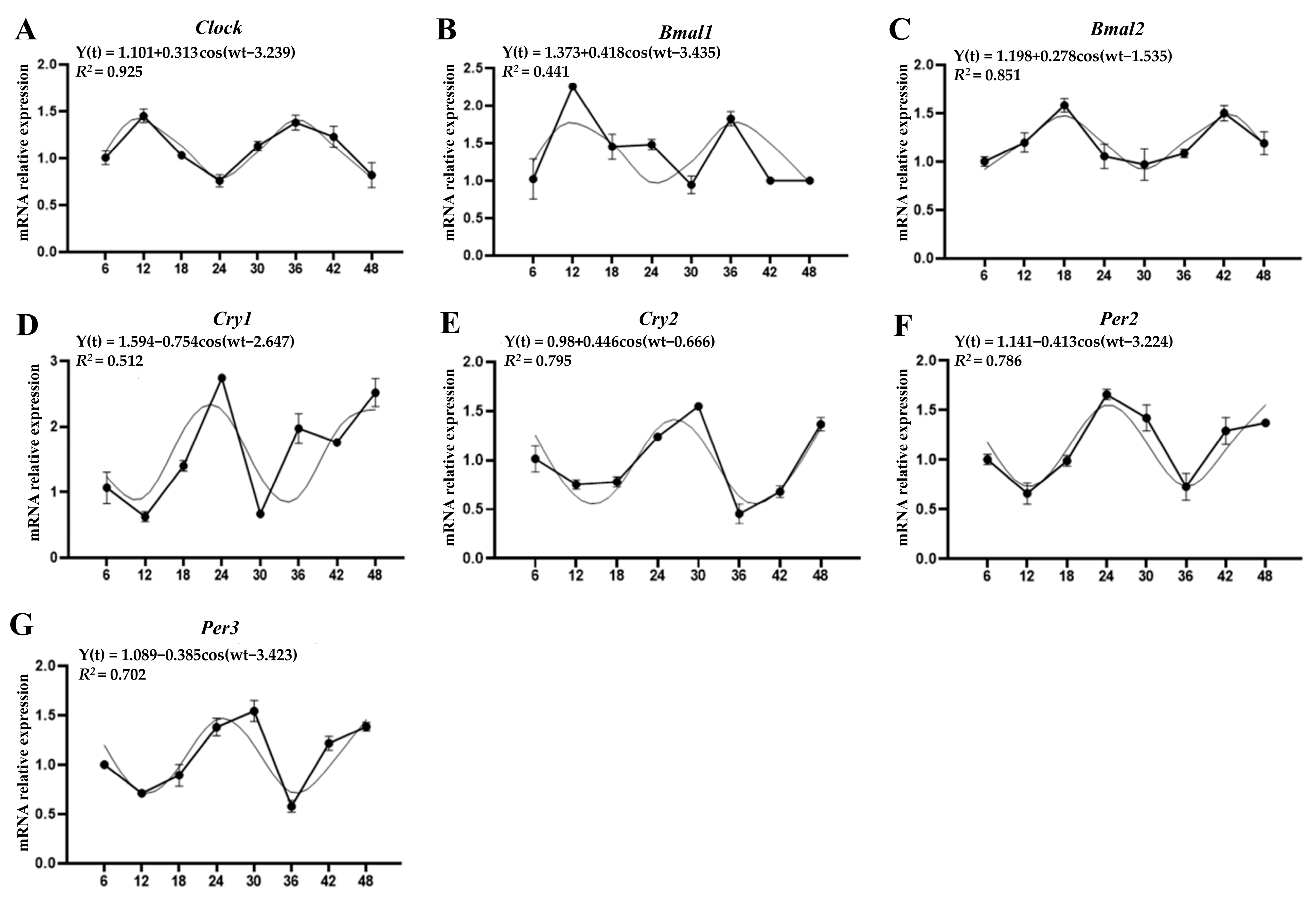
3.2. Circadian Expression of Clock Genes in Pineal Glands
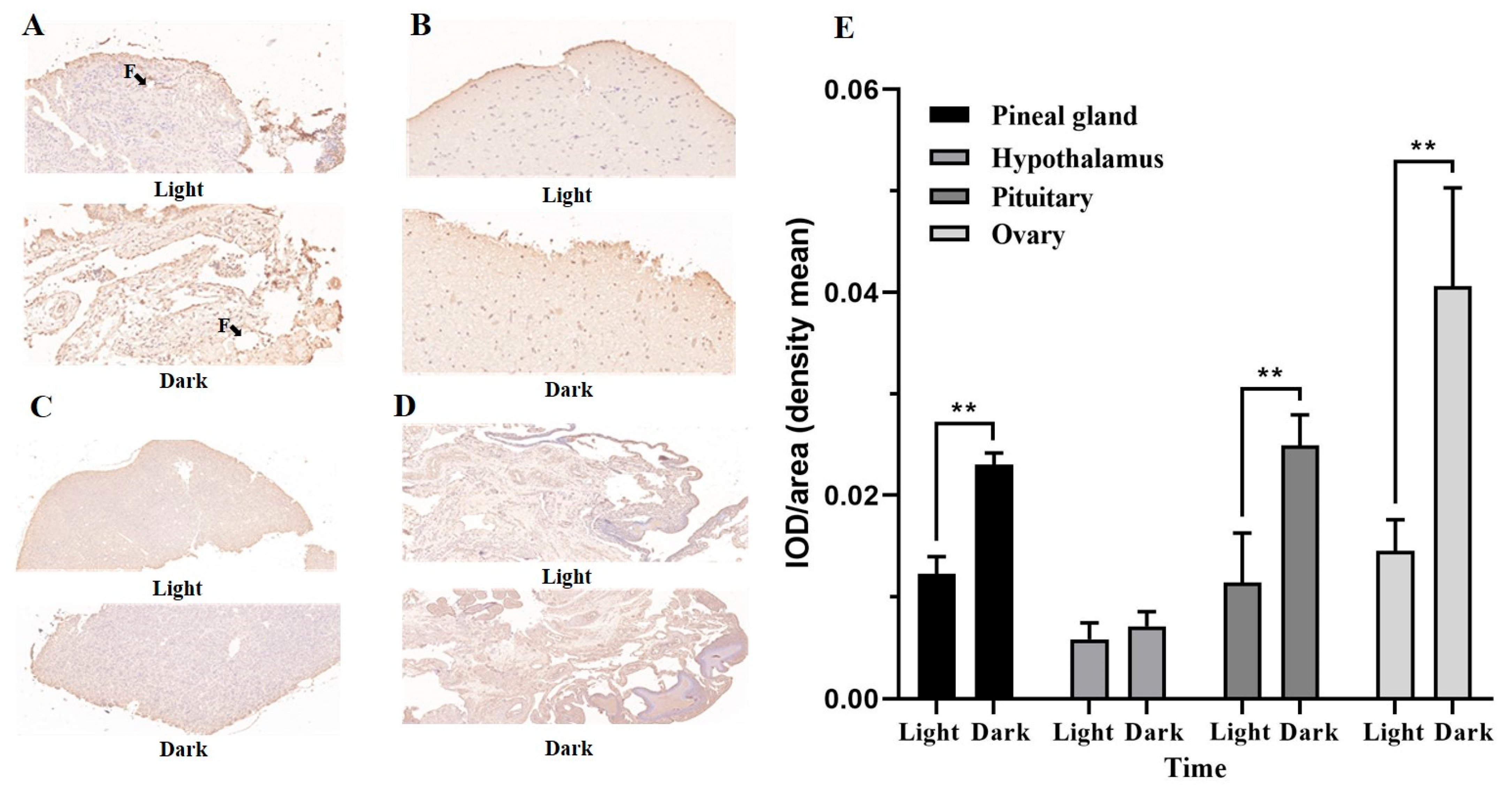
3.3. The Dynamics of Melatonin Secretion and Immunohistochemical Localization
3.4. Tissue Distribution and Circadian Expression of Mtnrs Genes
3.5. Circadian Oscillation of the Reproduction-Related Genes in HPG Axis
3.6. Correlation Analysis of Melatonin Hormone and Reproductive-Related Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Whylings, J.; Rigney, N.; de Vries, G.J.; Petrulis, A. Reduction in vasopressin cells in the suprachiasmatic nucleus in mice increases anxiety and alters fluid intake. Horm. Behav. 2021, 133, 104997. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Yuan, T.; Fu, Y.; Niu, D.; Chen, W.; Chen, L.; Lu, L. Seasonal differences in the transcriptome profile of the Zhedong white goose (Anser cygnoides) pituitary gland. Poult. Sci. 2021, 100, 1154–1166. [Google Scholar] [CrossRef] [PubMed]
- Bécot, L.; Bédère, N.; Coton, J.; Burlot, T.; Le Roy, P. Nest preference and laying duration traits to select against floor eggs in laying hens. Genet. Sel. Evol. GSE 2023, 55, 8. [Google Scholar] [CrossRef]
- Sharp, P.J.; Dawson, A.; Lea, R.W. Control of luteinizing hormone and prolactin secretion in birds. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1998, 119, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Patterson, P.H. The Relationship of Oviposition Time and Egg Characteristics to the Daily Light: Dark Cycle. J. Appl. Poult. Res. 1997, 6, 381–390. [Google Scholar] [CrossRef]
- Wolc, A.; Bednarczyk, M.; Lisowski, M.; Szwaczkowski, T. Genetic relationships among time of egg formation, clutch traits and traditional selection traits in laying hens. J. Anim. Feed Sci. 2010, 19, 452–459. [Google Scholar] [CrossRef]
- Zou, K.; Asiamah, C.A.; Lu, L.L.; Liu, Y.; Pan, Y.; Chen, T.; Zhao, Z.; Su, Y. Ovarian transcriptomic analysis and follicular development of Leizhou black duck. Poult. Sci. 2020, 99, 6173–6187. [Google Scholar] [CrossRef]
- Regan, S.L.P.; Knight, P.G.; Yovich, J.L.; Leung, Y.; Arfuso, F.; Dharmarajan, A. Involvement of Bone Morphogenetic Proteins (BMP) in the Regulation of Ovarian Function. Vitam. Horm. 2018, 107, 227–261. [Google Scholar] [CrossRef]
- Yang, W.; Chen, X.; Liu, Z.; Zhao, Y.; Chen, Y.; Geng, Z. Integrated transcriptome and proteome revealed that the declined expression of cell cycle-related genes associated with follicular atresia in geese. BMC Genom. 2023, 24, 24. [Google Scholar] [CrossRef]
- Lin, J.X.; Jia, Y.D.; Zhang, C.Q. Effect of epidermal growth factor on follicle-stimulating hormone-induced proliferation of granulosa cells from chicken prehierarchical follicles. J. Zhejiang Univ. Sci. B 2011, 12, 875–883. [Google Scholar] [CrossRef]
- Zhu, G.; Fang, C.; Li, J.; Mo, C.; Wang, Y.; Li, J. Transcriptomic Diversification of Granulosa Cells during Follicular Development in Chicken. Sci. Rep. 2019, 9, 5462. [Google Scholar] [CrossRef]
- Okubo, K.; Nagahama, Y. Structural and functional evolution of gonadotropin-releasing hormone in vertebrates. Acta Physiol. 2008, 193, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Xu, B.; Wang, W. Effects of luteinizing hormone and follicle stimulating hormone on the developmental competence of porcine preantral follicle oocytes grown in vitro. J. Assist. Reprod. Genet. 2007, 24, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, M.M.; Rossi, B.V.; Hagenauer, M.H.; Lee, T.M. Characterization of the estrous cycle in Octodon degus. Biol. Reprod. 2011, 84, 664–671. [Google Scholar] [CrossRef]
- Livadas, S.; Chrousos, G.P. Molecular and Environmental Mechanisms Regulating Puberty Initiation: An Integrated Approach. Front. Endocrinol. 2019, 10, 828. [Google Scholar] [CrossRef] [PubMed]
- Nimz, M.; Spitschak, M.; Schneider, F.; Fürbass, R.; Vanselow, J. Down-regulation of genes encoding steroidogenic enzymes and hormone receptors in late preovulatory follicles of the cow coincides with an accumulation of intrafollicular steroids. Domest. Anim. Endocrinol. 2009, 37, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Rathgeber, B.; McLean, N.; MacIsaac, J. Providing colored photoperiodic light stimulation during incubation: 2. Effects on early posthatch growth, immune response, and production performance in broiler chickens. Poult. Sci. 2021, 100, 101328. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Sun, Y.; Li, Y.; Fan, J.; Zong, Y.; Isa, A.M.; Shi, L.; Wang, Y.; Ni, A.; Ge, P.; et al. Monochromatic green light stimulation during incubation shortened the hatching time via pineal function in White Leghorn eggs. J. Anim. Sci. Biotechnol. 2021, 12, 17. [Google Scholar] [CrossRef]
- Csernus, V.; Mess, B. Biorhythms and pineal gland. Neuro Endocrinol. Lett. 2003, 24, 404–411. [Google Scholar]
- Wan, L.; Shi, X.Y.; Ge, W.R.; Sun, Y.L.; Zhang, S.; Wang, J.; Hu, L.Y.; Zou, L.P.; Yang, G. The Instigation of the Associations Between Melatonin, Circadian Genes, and Epileptic Spasms in Infant Rats. Front. Neurol. 2020, 11, 497225. [Google Scholar] [CrossRef]
- Nagy, A.D.; Kommedal, S.; Seomangal, K.; Csernus, V.J. Circadian expression of clock genes clock and Cry1 in the embryonic chicken pineal gland. Ann. N. Y. Acad. Sci. 2009, 1163, 484–487. [Google Scholar] [CrossRef] [PubMed]
- Martyniuk, K.; Hanuszewska, M.; Lewczuk, B. Metabolism of Melatonin Synthesis-Related Indoles in the Turkey Pineal Organ and Its Modification by Monochromatic Light. Int. J. Mol. Sci. 2020, 21, 9750. [Google Scholar] [CrossRef] [PubMed]
- Gwinner, E.; Brandstätter, R. Complex bird clocks. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2001, 356, 1801–1810. [Google Scholar] [CrossRef] [PubMed]
- McMullan, C.J.; Schernhammer, E.S.; Rimm, E.B.; Hu, F.B.; Forman, J.P. Melatonin secretion and the incidence of type 2 diabetes. JAMA 2013, 309, 1388–1396. [Google Scholar] [CrossRef] [PubMed]
- Nishiwaki-Ohkawa, T.; Yoshimura, T. Molecular basis for regulating seasonal reproduction in vertebrates. J. Endocrinol. 2016, 229, R117–R127. [Google Scholar] [CrossRef] [PubMed]
- Natesan, A.; Geetha, L.; Zatz, M. Rhythm and soul in the avian pineal. Cell Tissue Res. 2002, 309, 35–45. [Google Scholar] [CrossRef]
- Bell-Pedersen, D.; Cassone, V.M.; Earnest, D.J.; Golden, S.S.; Hardin, P.E.; Thomas, T.L.; Zoran, M.J. Circadian rhythms from multiple oscillators: Lessons from diverse organisms. Nat. Rev. Genet. 2005, 6, 544–556. [Google Scholar] [CrossRef]
- Ciani, E.; Fontaine, R.; Maugars, G.; Mizrahi, N.; Mayer, I.; Levavi-Sivan, B.; Weltzien, F.A. Melatonin receptors in Atlantic salmon stimulate cAMP levels in heterologous cell lines and show season-dependent daily variations in pituitary expression levels. J. Pineal Res. 2019, 67, e12590. [Google Scholar] [CrossRef]
- Dubocovich, M.L.; Markowska, M. Functional MT1 and MT2 melatonin receptors in mammals. Endocrine 2005, 27, 101–110. [Google Scholar] [CrossRef]
- Yu, K.; Deng, S.L.; Sun, T.C.; Li, Y.Y.; Liu, Y.X. Melatonin Regulates the Synthesis of Steroid Hormones on Male Reproduction: A Review. Molecules 2018, 23, 447. [Google Scholar] [CrossRef]
- Feng, P.; Zhao, W.; Xie, Q.; Zeng, T.; Lu, L.; Yang, L. Polymorphisms of melatonin receptor genes and their associations with egg production traits in Shaoxing duck. Asian-Australas. J. Anim. Sci. 2018, 31, 1535–1541. [Google Scholar] [CrossRef]
- Wang, Y.; Peng, K.M.; Li, J.L.; Song, H.; Li, S.H.; Wei, L.; Wang, J.X. Ultrastructure and melatonin 1a receptor distribution in the ovaries of African ostrich chicks. Cytotechnology 2008, 56, 187–195. [Google Scholar] [CrossRef][Green Version]
- Huang, Y.M.; Shi, Z.D.; Liu, Z.; Liu, Y.; Li, X.W. Endocrine regulations of reproductive seasonality, follicular development and incubation in Magang geese. Anim. Reprod. Sci. 2008, 104, 344–358. [Google Scholar] [CrossRef]
- Zhang, X.; Kang, B.; Zhang, L.N.; Guo, J.R.; Jiang, D.M.; Ji, H.; Zhen, L.; Yang, H.M. Gene expression profiles of LH, prolactin and their receptors in female Zi geese (Anser cygnoides) during development. Folia Biol. 2013, 61, 59–64. [Google Scholar] [CrossRef][Green Version]
- Oh, G.; Koncevičius, K.; Ebrahimi, S.; Carlucci, M.; Groot, D.E.; Nair, A.; Zhang, A.; Kriščiūnas, A.; Oh, E.S.; Labrie, V.; et al. Circadian oscillations of cytosine modification in humans contribute to epigenetic variability, aging, and complex disease. Genome Biol. 2019, 20, 2. [Google Scholar] [CrossRef]
- Kobayashi, H.; Oishi, K.; Hanai, S.; Ishida, N. Effect of feeding on peripheral circadian rhythms and behaviour in mammals. Genes Cells 2004, 9, 857–864. [Google Scholar] [CrossRef]
- Tischkau, S.A.; Howell, R.E.; Hickok, J.R.; Krager, S.L.; Bahr, J.M. The Luteinizing Hormone Surge Regulates Circadian Clock Gene Expression in the Chicken Ovary. Chronobiol. Int. 2011, 28, 10–20. [Google Scholar] [CrossRef]
- Zhang, Z.C.; Wang, Y.G.; Li, L.; Yin, H.D.; Li, D.Y.; Wang, Y.; Zhao, X.L.; Liu, Y.P.; Zhu, Q. Circadian clock genes are rhythmically expressed in specific segments of the hen oviduct. Poult. Sci. 2016, 95, 1653–1659. [Google Scholar] [CrossRef]
- Khan, S.; Roberts, J.; Wu, S.B. Reference gene selection for gene expression study in shell gland and spleen of laying hens challenged with infectious bronchitis virus. Sci. Rep. 2017, 7, 14271. [Google Scholar] [CrossRef]
- Bao, Q.; Yao, Y.; Weng, K.; Zheng, M.; Zhang, Y.; Zhang, Y.; Chen, G.; Xu, Q. Research Note: Comparison on laying behavior and clutch traits among Zhedong white geese (Anser cygnoides), Sichuan white geese (Anser cygnoides), and Hungarian geese (Anser anser). Poult. Sci. 2022, 101, 101594. [Google Scholar] [CrossRef]
- Morris, A.R.; Stanton, D.L.; Roman, D.; Liu, A.C. Systems Level Understanding of Circadian Integration with Cell Physiology. J. Mol. Biol. 2020, 432, 3547–3564. [Google Scholar] [CrossRef] [PubMed]
- Lagishetty, V.; Parthasarathy, P.T.; Phillips, O.; Fukumoto, J.; Cho, Y.; Fukumoto, I.; Bao, H.; Cox, R., Jr.; Galam, L.; Lockey, R.F.; et al. Dysregulation of CLOCK gene expression in hyperoxia-induced lung injury. Am. J. Physiol. Cell Physiol. 2014, 306, C999–C1007. [Google Scholar] [CrossRef]
- Helfer, G.; Fidler, A.E.; Vallone, D.; Foulkes, N.S.; Brandstaetter, R. Molecular analysis of clock gene expression in the avian brain. Chronobiol. Int. 2006, 23, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Fukada, Y.; Okano, T. Circadian clock system in the pineal gland. Mol. Neurobiol. 2002, 25, 19–30. [Google Scholar] [CrossRef]
- Keshvari, M.; Nejadtaghi, M.; Hosseini-Beheshti, F.; Rastqar, A.; Patel, N. Exploring the role of circadian clock gene and association with cancer pathophysiology. Chronobiol. Int. 2020, 37, 151–175. [Google Scholar] [CrossRef] [PubMed]
- Reppert, S.M.; Weaver, D.R. Coordination of circadian timing in mammals. Nature 2002, 418, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Teo, C.H.; Soga, T.; Parhar, I.S. Social Isolation Modulates CLOCK Protein and Beta-Catenin Expression Pattern in Gonadotropin-Inhibitory Hormone Neurons in Male Rats. Front. Endocrinol. 2017, 8, 225. [Google Scholar] [CrossRef]
- Wojtulewicz, K.; Tomaszewska-Zaremba, D.; Herman, A.P. Endotoxin-Induced Inflammation Suppresses the Effect of Melatonin on the Release of LH from the Ovine Pars Tuberalis Explants-Ex Vivo Study. Molecules 2017, 22, 1933. [Google Scholar] [CrossRef]
- Matsuda, F.; Inoue, N.; Manabe, N.; Ohkura, S. Follicular growth and atresia in mammalian ovaries: Regulation by survival and death of granulosa cells. J. Reprod. Dev. 2012, 58, 44–50. [Google Scholar] [CrossRef]
- Owino, S.; Buonfiglio, D.D.C.; Tchio, C.; Tosini, G. Melatonin Signaling a Key Regulator of Glucose Homeostasis and Energy Metabolism. Front. Endocrinol. 2019, 10, 488. [Google Scholar] [CrossRef]
- Markus, R.P.; Cecon, E.; Pires-Lapa, M.A. Immune-pineal axis: Nuclear factor κB (NF-kB) mediates the shift in the melatonin source from pinealocytes to immune competent cells. Int. J. Mol. Sci. 2013, 14, 10979–10997. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.G.; Woo, Y.S.; Park, S.W.; Seog, D.H.; Seo, M.K.; Bahk, W.M. The Neuroprotective Effects of Melatonin: Possible Role in the Pathophysiology of Neuropsychiatric Disease. Brain Sci. 2019, 9, 285. [Google Scholar] [CrossRef] [PubMed]
- Arendt, J. Melatonin and human rhythms. Chronobiol. Int. 2006, 23, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Portugal, S.J.; White, C.R.; Frappell, P.B.; Green, J.A.; Butler, P.J. Impacts of “supermoon” events on the physiology of a wild bird. Ecol. Evol. 2019, 9, 7974–7984. [Google Scholar] [CrossRef] [PubMed]
- Valenti, S.; Thellung, S.; Florio, T.; Giusti, M.; Schettini, G.; Giordano, G. A novel mechanism for the melatonin inhibition of testosterone secretion by rat Leydig cells: Reduction of GnRH-induced increase in cytosolic Ca2+. J. Mol. Endocrinol. 1999, 23, 299–306. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Content | Item | Content |
|---|---|---|---|
| Metabolic energy (MJ/kg) | 11.29 | Zinc (mg/kg) | 33 |
| Crude protein (%) | 16 | Calcium (g/kg) | 45 |
| Corn powder (g/kg) | 403 | Manganese (mg/kg) | 55 |
| Crushed wheat (g/kg) | 250 | Available phosphorus (g/kg) | 35 |
| Bean cake powder (g/kg) | 135 | Vitamin A (IU/kg) | 4409 |
| Green hay powder (g/kg) | 127 | Vitamin D3 (IU/kg) | 661 |
| Salt (g/kg) | 5 |
| Gene Name | Accession Number | Primer Sequence (5′-3′) | PCR Product (bp) |
|---|---|---|---|
| Mel-1a | XM_048078352.1 | F: TCATGCACGTTTGCACAGTC | 169 |
| R: GTGGTCTCAGTCTGGGGTTG | |||
| Mel-1b | XM_013178069.2 | F: CACGGTGGTGGACATCT | 139 |
| R: CAGTGGGTATGGATACAAGG | |||
| Mel-1c | XM_048075512.1 | F: CAGATAAGTGGGTTCCTGATGGG | 103 |
| R: ACCGAAGGCTGTGGCAGATGTAG | |||
| Clock | XM_048049947.1 | F: CAGAAAGACGGCGAGAAC | 155 |
| R: GGAGGAAGCAGATGGTGT | |||
| Bmal1 | XM_013189028.2 | F: CCAGGTGGAACAAGAGC | 146 |
| R: GATGTATCAGGCGGTGG | |||
| Bmal2 | XM_048050182.1 | F: GTTTTACGGATGGCTGTG | 127 |
| R: CTGCTGCCCTAAGGATTA | |||
| Cry1 | XM_048057002.1 | F: TGGCTGTTTGTCTTGTCG | 145 |
| R: CCGTGGATTGTTAGTCGC | |||
| Cry2 | XM_048065130.1 | F: GGCCATCATTAGCCGTAT | 106 |
| R: CACATCATCGTGGTTCTCC | |||
| Per2 | XM_048062937.1 | F: GGCTTCTCCACCTTTATTCC | 141 |
| R: TTGCTGCCTATGGCTCCT | |||
| Per3 | XM_048067384.1 | F: CGTCCTGTGAATGCTCTG | 114 |
| R: CTTGTCCACGTAGTTCTGCT | |||
| GnRH a | DQ023158 | F: GAAGATCTTGGTCGGTGTCCTCCTGT | 262 |
| R: AATCTCCTTTCTTCTGGCTTCTCCTTC | |||
| FSHβ | XM_013177587.2 | F: CACCAGTATCATCCGTTCAGC | 153 |
| R: CAGTGCTATCAGTGTCACAGGTC | |||
| LH b | DQ023159 | F: GGTGTATCGCAGCCCTTTG | 133 |
| R: TATCAGAGCCACGGGGAGG | |||
| ESR1 | XM_013178332.2 | F: GCAAAGAGAGTTCCAGGATTTGT | 103 |
| R: AGCGCCAGACTAAGCCAATC | |||
| PGR | XM_013178063.2 | F: GCGTCCTCTACAAGACCGAA | 227 |
| R: GGATTCTGTATCTGGCCGCA | |||
| GAPDH | XM_013199522.2 | F: GTGGTGCAAGAGGCATTGCTGAC | 86 |
| R: GCTGATGCTCCCATGTTCGTGAT |
| Gene | Mesor ± SEM | Amplitude | Acrophase (hh:mm) | Cosine p-Value |
|---|---|---|---|---|
| Clock | 1.10 ± 0.13 | 0.313 | 12:22 | 0.001 * |
| Bmal1 | 1.37 ± 0.08 | 0.418 | 13:02 | n.s. |
| Bmal2 | 1.19 ± 0.06 | 0.278 | 17:48 | 0.001 * |
| Cry1 | 1.59 ± 0.03 | 0.754 | 22:06 | 0.046 * |
| Cry2 | 0.98 ± 0.05 | 0.446 | 02:32 | 0.003 * |
| Per2 | 1.14 ± 0.07 | 0.413 | 00:19 | 0.003 * |
| Per3 | 1.08 ± 0.05 | 0.385 | 01:05 | 0.009 * |
| Parameters | 6:00 | 12:00 | 18:00 | 24:00 |
|---|---|---|---|---|
| MT (pg/mL) | 5.093 ± 0.295 b | 4.103 ± 0.125 c | 3.997 ± 0.094 c | 6.146 ± 0.243 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bao, Q.; Liu, D.; Guo, Y.; Gu, W.; Cao, Z.; Zhang, Y.; Zhang, Y.; Xu, Q.; Chen, G. Melatonin Secretion in Regulating the Circadian Rhythms of Reproduction in Goose (Anser cygnoides). Agriculture 2023, 13, 1620. https://doi.org/10.3390/agriculture13081620
Bao Q, Liu D, Guo Y, Gu W, Cao Z, Zhang Y, Zhang Y, Xu Q, Chen G. Melatonin Secretion in Regulating the Circadian Rhythms of Reproduction in Goose (Anser cygnoides). Agriculture. 2023; 13(8):1620. https://doi.org/10.3390/agriculture13081620
Chicago/Turabian StyleBao, Qiang, Di Liu, Yujiao Guo, Wang Gu, Zhengfeng Cao, Yu Zhang, Yang Zhang, Qi Xu, and Guohong Chen. 2023. "Melatonin Secretion in Regulating the Circadian Rhythms of Reproduction in Goose (Anser cygnoides)" Agriculture 13, no. 8: 1620. https://doi.org/10.3390/agriculture13081620
APA StyleBao, Q., Liu, D., Guo, Y., Gu, W., Cao, Z., Zhang, Y., Zhang, Y., Xu, Q., & Chen, G. (2023). Melatonin Secretion in Regulating the Circadian Rhythms of Reproduction in Goose (Anser cygnoides). Agriculture, 13(8), 1620. https://doi.org/10.3390/agriculture13081620