

Transforming Rhodotorula sp. Biomass to Active Biologic Compounds for Poultry Nutrition


 ,
, 
Abstract
1. Introduction
2. Rhodotorula sp. General Aspects
3. Factors Affecting Pigmentation
3.1. Yeasts Nutrition
3.2. Yeasts Fermentation Conditions
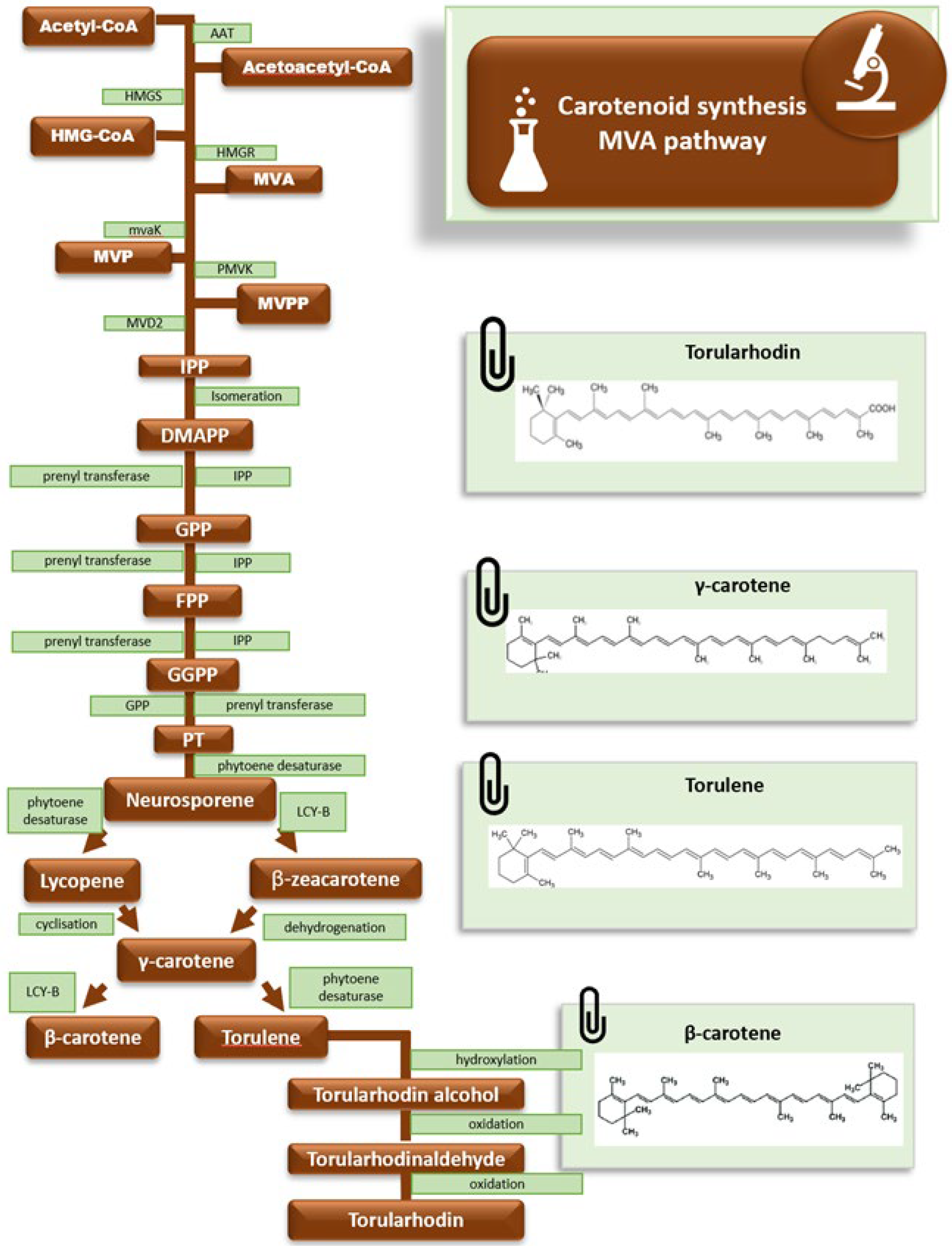
4. Yeasts Pigment Extraction and Quantification
5. Yeast Carotenoids in Poultry Nutrition
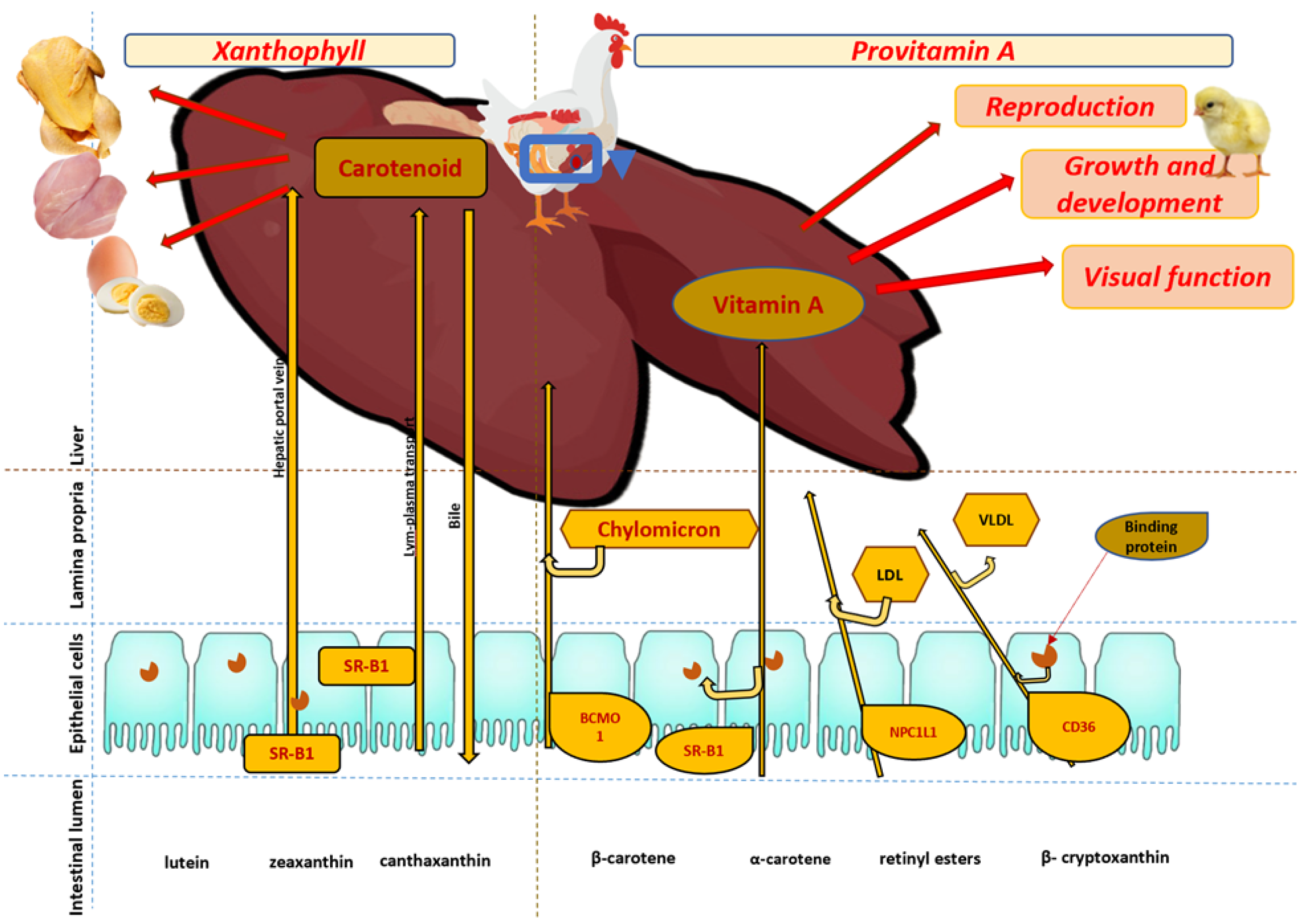
5.1. Retinol Requirements and Retinol Precursors in Poultry
5.2. Carotenoid Absorption in Poultry
5.3. Poultry Feed Sensorial Additives
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Novoveská, L.; Ross, M.E.; Stanley, M.S.; Pradelles, R.; Wasiolek, V.; Sassi, J.F. Microalgal Carotenoids: A Review of Production, Current Markets, Regulations, and Future Direction. Mar. Drugs 2019, 17, 640. [Google Scholar] [CrossRef]
- Hempel, J.; Schädle, C.N.; Leptihn, S.; Carle, R.; Schweiggert, R.M. Structure Related Aggregation Behavior of Carotenoids and Carotenoid Esters. J. Photochem. Photobiol. A Chem. 2016, 317, 161–174. [Google Scholar] [CrossRef]
- Kiokias, S.; Proestos, C.; Varzakas, T. A Review of the Structure, Biosynthesis, Absorption of Carotenoids-Analysis and Properties of Their Common Natural Extracts. Curr. Res. Nutr. Food Sci. 2016, 4, 25–37. [Google Scholar] [CrossRef]
- Swapnil, P.; Meena, M.; Singh, S.K.; Dhuldhaj, U.P.; Harish; Marwal, A. Vital Roles of Carotenoids in Plants and Humans to Deteriorate Stress with Its Structure, Biosynthesis, Metabolic Engineering and Functional Aspects. Curr. Plant Biol. 2021, 26, 100203. [Google Scholar] [CrossRef]
- Maslova, T.G.; Markovskaya, E.F.; Slemnev, N.N. Functions of Carotenoids in Leaves of Higher Plants (Review). Biol. Bull. Rev. 2021, 11, 476–487. [Google Scholar] [CrossRef]
- Zurak, D.; Slovenec, P.; Janječić, Z.; Bedeković, D.B.; Pintar, J.; Kljak, K. Overview on Recent Findings of Nutritional and Non-Nutritional Factors Affecting Egg Yolk Pigmentation. Worlds. Poult. Sci. J. 2022, 78, 531–560. [Google Scholar] [CrossRef]
- Nabi, F.; Arain, M.A.; Rajput, N.; Alagawany, M.; Soomro, J.; Umer, M.; Soomro, F.; Wang, Z.; Ye, R.; Liu, J. Health Benefits of Carotenoids and Potential Application in Poultry Industry: A Review. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1809–1818. [Google Scholar] [CrossRef]
- Shojadoost, B.; Yitbarek, A.; Alizadeh, M.; Kulkarni, R.R.; Astill, J.; Boodhoo, N.; Sharif, S. Centennial Review: Effects of Vitamins A, D, E, and C on the Chicken Immune System. Poult. Sci. 2021, 100, 100930. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Meng, J.; Zhang, L.; Bao, J.; Shi, W.; Li, Q.; Wang, X. Shudi Erzi San Relieves Ovary Aging in Laying Hens. Poult. Sci. 2022, 101, 102033. [Google Scholar] [CrossRef]
- Rapoport, A.; Guzhova, I.; Bernetti, L.; Buzzini, P.; Kieliszek, M.; Kot, A.M. Carotenoids and Some Other Pigments from Fungi and Yeasts. Metabolites 2021, 11, 92. [Google Scholar] [CrossRef] [PubMed]
- Scientific Committee on Animal Nutrition. Opinion on the Use of Canthaxanthin in Feedingstuffs for Salmon and Trout, Laying Hens, and Other Poultry; European Commission: Brussels, Belgium, 2002; pp. 1–29. [Google Scholar]
- Muley, S.; Nandgude, T.; Poddar, S. Extrusion–Spheronization a Promising Pelletization Technique: In-Depth Review. Asian J. Pharm. Sci. 2016, 11, 684–699. [Google Scholar] [CrossRef]
- Katoch, R. Effect of Harvest and Storage on Forage Quality. In Nutritional Quality Management of Forages in the Himalayan Region; Katoch, R., Ed.; Springer: Singapore, 2022; pp. 547–555. ISBN 978-981-16-5437-4. [Google Scholar]
- Steven, R.; Humaira, Z.; Natanael, Y.; Dwivany, F.M.; Trinugroho, J.P.; Dwijayanti, A.; Kristianti, T.; Tallei, T.E.; Emran, T.B.; Jeon, H.; et al. Marine Microbial-Derived Resource Exploration: Uncovering the Hidden Potential of Marine Carotenoids. Mar. Drugs 2022, 20, 352. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 6 July 2022).
- Wei, Z.; Huang, Q. Assembly of Protein-Polysaccharide Complexes for Delivery of Bioactive Ingredients: A Perspective Paper. J. Agric. Food Chem. 2019, 67, 1344–1352. [Google Scholar] [CrossRef] [PubMed]
- FutureMarketInsights Carotenoids Pigment Market Size, Industry Share & Trends—2032. Available online: https://www.futuremarketinsights.com/reports/carotenoids-pigment-market (accessed on 6 July 2022).
- Pailliè-Jiménez, M.E.; Stincone, P.; Brandelli, A. Natural Pigments of Microbial Origin. Front. Sustain. Food Syst. 2020, 4, 1–8. [Google Scholar] [CrossRef]
- Zhang, Z.; Pang, Z.; Xu, S.; Wei, T.; Song, L.; Wang, G.; Zhang, J.; Yang, X. Improved Carotenoid Productivity and COD Removal Efficiency by Co-Culture of Rhodotorula glutinis and Chlorella Vulgaris Using Starch Wastewaters as Raw Material. Appl. Biochem. Biotechnol. 2019, 189, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.Q.; Chen, G.; Chen, S.N.; Zhu, H.L.; Xiong, W.N.; Xu, M.; Jian, S.P. Metabolic Changes of Neurospora Crassa in the Presence of Oleic Acid for Promoting Lycopene Production. J. Biosci. Bioeng. 2021, 132, 148–153. [Google Scholar] [CrossRef]
- Higa, Y.; Kim, Y.S.; Altaf-Ul-Amin, M.; Huang, M.; Ono, N.; Kanaya, S. Divergence of Metabolites in Three Phylogenetically Close Monascus Species (M. pilosus, M. ruber, and M. purpureus) Based on Secondary Metabolite Biosynthetic Gene Clusters. BMC Genom. 2020, 21, 679. [Google Scholar] [CrossRef]
- Sevgili, A.; Erkmen, O. Improved Lycopene Production from Different Substrates by Mated Fermentation of Blakeslea Trispora. Foods 2019, 8, 120. [Google Scholar] [CrossRef]
- Kumar, A.; Prajapati, S.; Nikhil; Nandan, S.; Neogi, T.G. Industrially Important Pigments from Different Groups of Fungi. In Recent Advancement in White Biotechnology through Fungi: Volume 2: Perspective for Value-Added Products and Environments; Yadav, A.N., Singh, S., Mishra, S., Gupta, A., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 285–301. ISBN 978-3-030-14846-1. [Google Scholar]
- Gurler, H.N.; Yilmazer, C.; Erkan, S.B.; Ozcan, A.; Yatmaz, E.; Öziyci, H.R.; Karhan, M.; Turhan, I. Applicability of Recombinant Aspergillus Sojae Crude Mannanase Enzyme in Carrot Juice Production. J. Food Process. Preserv. 2021, 45, 1–11. [Google Scholar] [CrossRef]
- Dai, Z.B.; Wang, X.; Li, G.H. Secondary Metabolites and Their Bioactivities Produced by Paecilomyces. Molecules 2020, 25, 5077. [Google Scholar] [CrossRef]
- Brendler, T.; Williamson, E.M. Astaxanthin: How Much Is Too Much? A Safety Review. Phyther. Res. 2019, 33, 3090–3111. [Google Scholar] [CrossRef] [PubMed]
- Behera, H.T.; Mojumdar, A.; Nivedita, S.; Ray, L. Microbial Pigments: Secondary Metabolites with Multifaceted Roles. In Microbial Polymers: Applications and Ecological Perspectives; Vaishnav, A., Choudhary, D.K., Eds.; Springer: Singapore, 2021; pp. 631–654. ISBN 978-981-16-0045-6. [Google Scholar]
- Kongkapan, T.S. EBSCOhost|150240033|Isolation and Production of Prodigiosin and Cycloprodigiosin from Marine Sponges-Associated Bacteria of the Andaman Coast of Thailand. Available online: https://web.p.ebscohost.com/abstract?direct=true&prfile=ehost&scope=site&authtype=crawler&jrnl=01253395&AN=150240033&h=%2FW6SE%2B0dta8HX%2FH2YoiP3x1oGra92MNYRHfuRc7w40%2FiyjfEyiRRtI7YSlVGOjbH%2BX%2FTimicqZpuZGLb7KfnNw%3D%3D&crl=c&resultNs=AdminWebAuth&resultLocal=ErrCrlNotAuth&crlhashurl=login.aspx%3Fdirect%3Dtrue%26profile%3Dehost%26scope%3Dsite%26authtype%3Dcrawler%26jrnl%3D01253395%26AN%3D150240033 (accessed on 6 July 2022).
- Elkenawy, N.M.; Yassin, A.S.; Elhifnawy, H.N.; Amin, M.A. Optimization of Prodigiosin Production by Serratia Marcescens Using Crude Glycerol and Enhancing Production Using Gamma Radiation. Biotechnol. Rep. 2017, 14, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Mussagy, C.U. Production of Carotenoids by Yeast Rhodotorula glutinis CCT-2186 and Their Extraction Using Alternative Solvents. Ph.D. Thesis, Universidade Estadual Paulista/Biociências e Biotecnologia Aplicadas à Farmácia, São Paulo, Brazil, 2021. [Google Scholar]
- Barredo, J.L.; García-Estrada, C.; Kosalkova, K.; Barreiro, C. Biosynthesis of Astaxanthin as a Main Carotenoid in the Heterobasidiomycetous Yeast Xanthophyllomyces dendrorhous. J. Fungi 2017, 3, 44. [Google Scholar] [CrossRef] [PubMed]
- Landolfo, S.; Chessa, R.; Zara, G.; Zara, S.; Budroni, M.; Mannazzu, I. Rhodotorula mucilaginosa C2.5t1 Modulates Carotenoid Content and Car Genes Transcript Levels to Counteract the pro-Oxidant Effect of Hydrogen Peroxide. Microorganisms 2019, 7, 316. [Google Scholar] [CrossRef] [PubMed]
- Chrissian, C.; Lin, C.P.C.; Camacho, E.; Casadevall, A.; Neiman, A.M.; Stark, R.E. Unconventional Constituents and Shared Molecular Architecture of the Melanized Cell Wall of c. Neoformans and Spore Wall of S. cerevisiae. J. Fungi 2020, 6, 329. [Google Scholar] [CrossRef] [PubMed]
- Pino-Maureira, N.L.; González-Saldía, R.R.; Capdeville, A.; Srain, B. Rhodotorula Strains Isolated from Seawater That Can Biotransform Raw Glycerol into Docosahexaenoic Acid (Dha) and Carotenoids for Animal Nutrition. Appl. Sci. 2021, 11, 2824. [Google Scholar] [CrossRef]
- Tiwari, S.; Baghela, A.; Libkind, D. Rhodotorula sampaioana f.a., sp. nov., a Novel Red Yeast of the Order Sporidiobolales Isolated from Argentina and India. Antonie van Leeuwenhoek 2021, 114, 1237–1244. [Google Scholar] [CrossRef]
- Ashok, G.; Mohan, U.; Boominathan, M.; Ravichandiran, V.; Viswanathan, C.; Senthilkumar, V. Natural Pigments from Filamentous Fungi: Production and Applications. In Industrially Important Fungi for Sustainable Development: Volume 2: Bioprospecting for Biomolecules; Abdel-Azeem, A.M., Yadav, A.N., Yadav, N., Sharma, M., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 651–678. ISBN 978-3-030-85603-8. [Google Scholar]
- Allahkarami, S.; Sepahi, A.A.; Hosseini, H.; Razavi, M.R. Isolation and Identification of Carotenoid-Producing Rhodotorula sp. from Pinaceae Forest Ecosystems and Optimization of in Vitro Carotenoid Production. Biotechnol. Rep. 2021, 32, e00687. [Google Scholar] [CrossRef]
- De Jong, A.W.; Hagen, F. Attack, Defend and Persist: How the Fungal Pathogen Candida Auris Was Able to Emerge Globally in Healthcare Environments. Mycopathologia 2019, 184, 353–365. [Google Scholar] [CrossRef]
- Kot, A.M.; Błażejak, S.; Kieliszek, M.; Gientka, I.; Bryś, J. Simultaneous Production of Lipids and Carotenoids by the Red Yeast Rhodotorula from Waste Glycerol Fraction and Potato Wastewater. Appl. Biochem. Biotechnol. 2019, 189, 589–607. [Google Scholar] [CrossRef]
- Gosalawit, C.; Imsoonthornruksa, S.; Gilroyed, B.H.; Mcnea, L.; Boontawan, A.; Ketudat-Cairns, M. The Potential of the Oleaginous Yeast Rhodotorula Paludigena CM33 to Produce Biolipids. J. Biotechnol. 2021, 329, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Okoro, O.V.; Gholipour, A.R.; Sedighi, F.; Shavandi, A.; Hamidi, M. Optimization of Exopolysaccharide (Eps) Production by Rhodotorula mucilaginosa sp. Gums16. Chemengineering 2021, 5, 39. [Google Scholar] [CrossRef]
- Maharana, A.K.; Singh, S.M. A Cold and Organic Solvent Tolerant Lipase Produced by Antarctic Strain Rhodotorula sp. Y-23. J. Basic Microbiol. 2018, 58, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Bonadio, M.D.P.; de Freita, L.A.; Mutton, M.J.R. Carotenoid Production in Sugarcane Juice and Synthetic Media Supplemented with Nutrients by Rhodotorula rubra L02. Braz. J. Microbiol. 2018, 49, 872–878. [Google Scholar] [CrossRef]
- Zhong, Z.; Zhang, X.; Wang, X.; Fu, S.; Wu, S.; Lu, X.; Ren, C.; Han, X.; Yang, G. Soil Bacteria and Fungi Respond Differently to Plant Diversity and Plant Family Composition during the Secondary Succession of Abandoned Farmland on the Loess Plateau, China. Plant Soil 2020, 448, 183–200. [Google Scholar] [CrossRef]
- Mussagy, C.U.; Khan, S.; Kot, A.M. Current Developments on the Application of Microbial Carotenoids as an Alternative to Synthetic Pigments. Crit. Rev. Food Sci. Nutr. 2021, 62, 6932–6946. [Google Scholar] [CrossRef]
- Čertík, M.; Klempová, T.; Guothová, L.; Mihálik, D.; Kraic, J. Biotechnology for the Functional Improvement of Cereal-Based Materials Enriched with PUFA and Pigments. Eur. J. Lipid Sci. Technol. 2013, 115, 1247–1256. [Google Scholar] [CrossRef]
- Tkáčová, J.; Čaplová, J.; Klempová, T.; Čertík, M. Correlation between Lipid and Carotenoid Synthesis in Torularhodin-Producing Rhodotorula glutinis. Ann. Microbiol. 2017, 67, 541–551. [Google Scholar] [CrossRef]
- Chaturvedi, S.; Bhattacharya, A.; Khare, S.K. Trends in Oil Production from Oleaginous Yeast Using Biomass: Biotechnological Potential and Constraints. Appl. Biochem. Microbiol. 2018, 54, 361–369. [Google Scholar] [CrossRef]
- Rodriguez-Concepcion, M.; Avalos, J.; Bonet, M.L.; Boronat, A.; Gomez-Gomez, L.; Hornero-Mendez, D.; Limon, M.C.; Meléndez-Martínez, A.J.; Olmedilla-Alonso, B.; Palou, A.; et al. A Global Perspective on Carotenoids: Metabolism, Biotechnology, and Benefits for Nutrition and Health. Prog. Lipid Res. 2018, 70, 62–93. [Google Scholar] [CrossRef]
- Banerjee, A.; Sharma, T.; Nautiyal, A.K.; Dasgupta, D.; Hazra, S.; Bhaskar, T.; Ghosh, D. Scale-up Strategy for Yeast Single Cell Oil Production for Rhodotorula Mucilagenosa IIPL32 from Corn Cob Derived Pentosan. Bioresour. Technol. 2020, 309, 123329. [Google Scholar] [CrossRef]
- Khot, M.; Ghosh, D. Lipids of Rhodotorula mucilaginosa IIPL32 with Biodiesel Potential: Oil Yield, Fatty Acid Profile, Fuel Properties. J. Basic Microbiol. 2017, 57, 345–352. [Google Scholar] [CrossRef] [PubMed]
- da Costa, W.A.; de Araújo Padilha, C.E.; de Oliveira, S.D.; da Silva, F.L.H.; Silva, J.; Ancântara, M.A.; Ferrari, M.; dos Santos, E.S. Oil-Lipids, Carotenoids and Fatty Acids Simultaneous Production by Rhodotorula mucilaginosa Cct3892 Using Sugarcane Molasses as Carbon Source. Braz. J. Food Technol. 2020, 23, 1–11. [Google Scholar] [CrossRef]
- Lario, L.D.; Pillaca-Pullo, O.S.; Durães Sette, L.; Converti, A.; Casati, P.; Spampinato, C.; Pessoa, A. Optimization of Protease Production and Sequence Analysis of the Purified Enzyme from the Cold Adapted Yeast Rhodotorula mucilaginosa CBMAI 1528. Biotechnol. Rep. 2020, 28, e00546. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, P.M.G.; de Morais, T.P.; de Andrade Silva, C.A.; da Silva Santos, F.R.; Garcia, N.F.L.; Fonseca, G.G.; Leite, R.S.R.; da Paz, M.F. Biochemical Characterization and Evaluation of Invertases Produced from Saccharomyces Cerevisiae CAT-1 and Rhodotorula mucilaginosa for the Production of Fructooligosaccharides. Prep. Biochem. Biotechnol. 2018, 48, 506–513. [Google Scholar] [CrossRef]
- Chaturvedi, S.; Gupta, A.K.; Bhattacharya, A.; Dutta, T.; Nain, L.; Khare, S.K. Overexpression and Repression of Key Rate-Limiting Enzymes (Acetyl CoA Carboxylase and HMG Reductase) to Enhance Fatty Acid Production from Rhodotorula mucilaginosa. J. Basic Microbiol. 2021, 61, 4–14. [Google Scholar] [CrossRef]
- Gong, G.; Zhang, X.; Tan, T. Simultaneously Enhanced Intracellular Lipogenesis and β-Carotene Biosynthesis of Rhodotorula glutinis by Light Exposure with Sodium Acetate as the Substrate. Bioresour. Technol. 2020, 295, 122274. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, D.; Yang, Q.; Wang, P. Enhancing Carotenoid Production in Rhodotorula mucilaginosa KC8 by Combining Mutation and Metabolic Engineering. Ann. Microbiol. 2017, 67, 425–431. [Google Scholar] [CrossRef]
- Park, H.; Kwak, M.; Seo, J.W.; Ju, J.H.; Heo, S.Y.; Park, S.M.; Hong, W.K. Enhanced Production of Carotenoids Using a Thraustochytrid Microalgal Strain Containing High Levels of Docosahexaenoic Acid-Rich Oil. Bioprocess Biosyst. Eng. 2018, 41, 1355–1370. [Google Scholar] [CrossRef]
- Liao, P.; Hemmerlin, A.; Bach, T.J.; Chye, M.L. The Potential of the Mevalonate Pathway for Enhanced Isoprenoid Production. Biotechnol. Adv. 2016, 34, 697–713. [Google Scholar] [CrossRef]
- Moser, S.; Pichler, H. Identifying and Engineering the Ideal Microbial Terpenoid Production Host. Appl. Microbiol. Biotechnol. 2019, 103, 5501–5516. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Wang, H.; Li, L.; Zhang, N.; Ma, Y. The Stability of Vitamin A from Different Sources in Vitamin Premixes and Vitamin-Trace Mineral Premixes. Appl. Sci. 2021, 11, 3657. [Google Scholar] [CrossRef]
- Meléndez-Martínez, A.J.; Mapelli-Brahm, P.; Stinco, C.M. The Colourless Carotenoids Phytoene and Phytofluene: From Dietary Sources to Their Usefulness for the Functional Foods and Nutricosmetics Industries. J. Food Compos. Anal. 2018, 67, 91–103. [Google Scholar] [CrossRef]
- Chreptowicz, K.; Mierzejewska, J.; Tkáčová, J.; Młynek, M.; Čertik, M. Carotenoid-Producing Yeasts: Identification and Characteristics of Environmental Isolates with a Valuable Extracellular Enzymatic Activity. Microorganisms 2019, 7, 653. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.H.; Wu, B.K.; Pan, D.; Sang, L.X.; Chang, B. Beta-Carotene and Its Protective Effect on Gastric Cancer. World J. Clin. Cases 2021, 9, 6591–6607. [Google Scholar] [CrossRef] [PubMed]
- Gupta, I.; Adin, S.N.; Panda, B.P.; Mujeeb, M. β-Carotene—Production Methods, Biosynthesis from Phaffia rhodozyma, Factors Affecting Its Production during Fermentation, Pharmacological Properties: A Review. Biotechnol. Appl. Biochem. 2022, 69, 2517–2529. [Google Scholar] [CrossRef]
- Kanno, K.Y.F.; Karp, S.G.; Rodrigues, C.; de Andrade Tanobe, V.O.; Soccol, C.R.; da Costa Cardoso, L.A. Influence of Organic Solvents in the Extraction and Purification of Torularhodin from Sporobolomyces ruberrimus. Biotechnol. Lett. 2021, 43, 89–98. [Google Scholar] [CrossRef]
- Ghilardi, C.; Sanmartin Negrete, P.; Carelli, A.A.; Borroni, V. Evaluation of Olive Mill Waste as Substrate for Carotenoid Production by Rhodotorula mucilaginosa. Bioresour. Bioprocess. 2020, 7, 52. [Google Scholar] [CrossRef]
- Kot, A.M.; Blazejak, S.; Kurcz, A.; Gientka, I.; Kieliszek, M. Rhodotorula glutinis potential source of lipids, carotenoids, and enzymes for use in industries. Appl. Microbiol. Biotechnol. 2016, 100, 6103–6117. [Google Scholar] [CrossRef]
- Geoffriau, E.; Simon, P. Carrots and Related Apiaceae Crops, 2nd ed.; CABI: Wallingford, UK, 2020; pp. 1–370. [Google Scholar]
- Kowalczyk, Z.; Cupiał, M. Environmental Analysis of the Conventional and Organic Production of Carrot in Poland. J. Clean. Prod. 2020, 269, 122169. [Google Scholar] [CrossRef]
- Aftab, T.; Hakeem, K.R. Plant Micronutrients: Deficiency and Toxicity Management; Tariq Aftab, K.R.H., Ed.; Deficiency; Springer: Cham, Switzerland, 2020; ISBN 3030498565. [Google Scholar]
- Foong, L.C.; Loh, C.W.L.; Ng, H.S.; Lan, J.C.W. Recent Development in the Production Strategies of Microbial Carotenoids. World J. Microbiol. Biotechnol. 2021, 37, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bessadok, B.; Santulli, A.; Breuck, T.; Sadok, S. Species Disparity Response to Mutagenesis of Marine Yeasts for the Potential Production of Biodiesel. Biotechnol. Biofuels 2019, 12, 1–16. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, X.; Tan, T. Lipid and Carotenoid Production by Rhodotorula glutinis under Irradiation/High-Temperature and Dark/Low-Temperature Cultivation. Bioresour. Technol. 2014, 157, 149–153. [Google Scholar] [CrossRef]
- Li, C.; Xu, Y.; Li, Z.; Cheng, P.; Yu, G. Transcriptomic and metabolomic analysis reveals the potential mechanisms underlying the improvement of β-carotene and torulene production in Rhodosporidiobolus colostri under low temperature treatment. Food Res. Int. 2022, 156, 111158. [Google Scholar] [CrossRef]
- Voaides, C.; Dima, R. The Effect of Nitrogen Source on Carotenoids Production by Rhodotorula sp. Rom. Biotechnol. Lett. 2012, 17, 7570–7576. [Google Scholar]
- Kong, W.; Yang, S.; Agboyibor, C.; Chen, D.; Zhang, A.; Niu, S. Light Irradiation Can Regulate the Growth Characteristics and Metabolites Compositions of Rhodotorula mucilaginosa. J. Food Sci. Technol. 2019, 56, 5509–5517. [Google Scholar] [CrossRef]
- Bento, T.F.S.R.; Viana, V.F.M.; Carneiro, L.M.; Silva, J.P.A. Influence of Agitation and Aeration on Single Cell Oil Production by Rhodotorula glutinis from Glycerol. J. Sustain. Bioenergy Syst. 2019, 09, 29–43. [Google Scholar] [CrossRef]
- Sharma, R.; Ghoshal, G. Optimization of Carotenoids Production by Rhodotorula mucilaginosa (MTCC-1403) Using Agro-Industrial Waste in Bioreactor: A Statistical Approach; Elsevier B.V.: Amsterdam, The Netherlands, 2020; Volume 25, ISBN 9101722779173. [Google Scholar]
- Sajjad, W.; Din, G.; Rafiq, M.; Iqbal, A.; Khan, S.; Zada, S.; Ali, B.; Kang, S. Pigment Production by Cold-Adapted Bacteria and Fungi: Colorful Tale of Cryosphere with Wide Range Applications. Extremophiles 2020, 24, 447–473. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Koh, C.M.J.; Yap, S.A.; Du, M.; Hlaing, M.M.; Ji, L. Identification of Novel Genes in the Carotenogenic and Oleaginous Yeast Rhodotorula toruloides through Genome-Wide Insertional Mutagenesis. BMC Microbiol. 2018, 18, 14. [Google Scholar] [CrossRef]
- Srivastava, A.; Kalwani, M.; Chakdar, H.; Pabbi, S.; Shukla, P. Biosynthesis and Biotechnological Interventions for Commercial Production of Microalgal Pigments: A Review. Bioresour. Technol. 2022, 352, 127071. [Google Scholar] [CrossRef]
- Zhao, Y.; Guo, L.; Xia, Y.; Zhuang, X.; Chu, W. Isolation, Identification of Carotenoid-Producing Rhodotorula Sp. From Marine Environment and Optimization for Carotenoid Production. Mar. Drugs 2019, 17, 161. [Google Scholar] [CrossRef]
- Zhang, C.; Shen, H.; Zhang, X.; Yu, X.; Wang, H.; Xiao, S.; Wang, J.; Zhao, Z.K. Combined Mutagenesis of Rhodosporidium toruloides for Improved Production of Carotenoids and Lipids. Biotechnol. Lett. 2016, 38, 1733–1738. [Google Scholar] [CrossRef]
- Hernández, A.; Pérez-Nevado, F.; Ruiz-Moyano, S.; Serradilla, M.J.; Villalobos, M.C.; Martín, A.; Córdoba, M.G. Spoilage Yeasts: What Are the Sources of Contamination of Foods and Beverages? Int. J. Food Microbiol. 2018, 286, 98–110. [Google Scholar] [CrossRef]
- Lin, X.; Gao, N.; Liu, S.; Zhang, S.; Song, S.; Ji, C.; Dong, X.; Su, Y.; Zhao, Z.K.; Zhu, B. Characterization the Carotenoid Productions and Profiles of Three Rhodosporidium toruloides Mutants from Agrobacterium Tumefaciens-Mediated Transformation. Yeast 2017, 34, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Johns, A.M.B.; Love, J.; Aves, S.J. Four Inducible Promoters for Controlled Gene Expression in the Oleaginous Yeast Rhodotorula toruloides. Front. Microbiol. 2016, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Elfeky, N.; Elmahmoudy, M.; Zhang, Y.; Guo, J.L.; Bao, Y. Lipid and Carotenoid Production by Rhodotorula glutinis with a Combined Cultivation Mode of Nitrogen, Sulfur, and Aluminium Stress. Appl. Sci. 2019, 9, 2444. [Google Scholar] [CrossRef]
- Villarreal, P.; Carrasco, M.; Barahona, S.; Alcaíno, J.; Cifuentes, V.; Baeza, M. Tolerance to Ultraviolet Radiation of Psychrotolerant Yeasts and Analysis of Their Carotenoid, Mycosporine, and Ergosterol Content. Curr. Microbiol. 2016, 72, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Gong, G.; Liu, L.; Zhang, X.; Tan, T. Multi-Omics Metabolism Analysis on Irradiation-Induced Oxidative Stress to Rhodotorula glutinis. Appl. Microbiol. Biotechnol. 2019, 103, 361–374. [Google Scholar] [CrossRef]
- Cortes, A.G.; Vásquez, J.A.G.; Díaz, Y.C.A.; Castrillón, M.R. Effects of cellular stress on pigment production in Rhodotorula mucilaginosa/alborubescens AJB01 strain from the Caribbean region of Colombia. bioRxiv 2020. [Google Scholar] [CrossRef]
- Verma, G.; Anand, P.; Pandey, S.; Nagar, S. Optimization of Cultivation Conditions for Microbial Lipid Production by Rhodotorula glutinis, An Oleaginous Yeast. Biosci. Biotechnol. Res. Commun. 2019, 12, 790–797. [Google Scholar] [CrossRef]
- Elsanhoty, R.M.; Al-Turki, A.I.; Abd El-Razik, M.M. Production of Carotenoids from Rhodotorula mucilaginosa and Their Applications as Colorant Agent in Sweet Candy. J. Food Agric. Environ. 2017, 15, 21–26. [Google Scholar]
- Varmira, K.; Habibi, A.; Moradi, S.; Bahramian, E. Experimental Evaluation of Airlift Photobioreactor for Carotenoid Pigments Production by Rhodotorula rubra. Rom. Biotechnol. Lett. 2018, 23, 13843. [Google Scholar] [CrossRef]
- Nasrin, T.A.A.; Matin, M.A. Valorization of Vegetable Wastes. In Food Processing By-Products and their Utilization, First; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017; pp. 53–88. [Google Scholar] [CrossRef]
- Kaur, P.; Ghoshal, G.; Jain, A. Bio-Utilization of Fruits and Vegetables Waste to Produce β-Carotene in Solid-State Fermentation: Characterization and Antioxidant Activity. Process Biochem. 2019, 76, 155–164. [Google Scholar] [CrossRef]
- Zheng, X.; Hu, R.; Chen, D.; Chen, J.; He, W.; Huang, L.; Lin, C.; Chen, H.; Chen, Y.; Zhu, J.; et al. Lipid and Carotenoid Production by the Rhodosporidium Toruloides Mutant in Cane Molasses. Bioresour. Technol. 2021, 326, 124816. [Google Scholar] [CrossRef]
- Machado, W.R.C.; Da Silva, L.G.; Vanzela, E.S.L.; Del Bianchi, V.L. Evaluation of the Process Conditions for the Production of Microbial Carotenoids by the Recently Isolated Rhodotorula mucilaginosa URM 7409. Braz. J. Food Technol. 2019, 22, 1–14. [Google Scholar] [CrossRef]
- Da Silva, J.; Honorato da Silva, F.L.; Santos Ribeiro, J.E.; Nóbrega de Melo, D.J.; Santos, F.A.; Lucena de Medeiros, L. Effect of Supplementation, Temperature and PH on Carotenoids and Lipids Production by Rhodotorula mucilaginosa on Sisal Bagasse Hydrolyzate. Biocatal. Agric. Biotechnol. 2020, 30, 101847. [Google Scholar] [CrossRef]
- Azarpour, A.; Zendehboudi, S.; Mohammadzadeh, O.; Rajabzadeh, A.R.; Chatzis, I. A Review on Microalgal Biomass and Biodiesel Production through Co-Cultivation Strategy. Energy Convers. Manag. 2022, 267, 115757. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, C.; Xu, B.; Fu, J.; Du, Y.; Fang, Q.; Dong, B.; Zhao, H. Temperature Regulation of Carotenoid Accumulation in the Petals of Sweet Osmanthus via Modulating Expression of Carotenoid Biosynthesis and Degradation Genes. BMC Genom. 2022, 23, 418. [Google Scholar] [CrossRef]
- Bhosale, P.; Gadre, R.V. Manipulation of Temperature and Illumination Conditions for Enhanced β-Carotene Production by Mutant 32 of Rhodotorula glutinis. Lett. Appl. Microbiol. 2002, 34, 349–353. [Google Scholar] [CrossRef]
- Touchette, D. Metabolic Activity Assessment of Polar Microorganisms and Metabolic Characterization of the Cold-Adapted Rhodotorula Yeast JG1b; McGill University: Montréal, QC, Canada, 2020. [Google Scholar]
- Pacia, M.Z.; Pukalski, J.; Turnau, K.; Baranska, M.; Kaczor, A. Lipids, Hemoproteins and Carotenoids in Alive Rhodotorula mucilaginosa Cells under Pesticide Decomposition—Raman Imaging Study. Chemosphere 2016, 164, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Corinaldesi, C.; Barone, G.; Marcellini, F.; Dell’Anno, A.; Danovaro, R. Marine Microbial-Derived Molecules and Their Potential Use in Cosmeceutical and Cosmetic Products. Mar. Drugs 2017, 15, 118. [Google Scholar] [CrossRef] [PubMed]
- Nazzaro, F.; Fratianni, F.; d’Acierno, A.; da Cruz, A.G.; De Feo, V.; Coppola, R. Microbial Production of Metabolites for Food and Processes; Elsevier: Amsterdam, The Netherlands, 2020; ISBN 9780444643018. [Google Scholar]
- San Martin, D.; Orive, M.; Iñarra, B.; Castelo, J.; Estévez, A.; Nazzaro, J.; Iloro, I.; Elortza, F.; Zufía, J. Brewers’ Spent Yeast and Grain Protein Hydrolysates as Second-Generation Feedstuff for Aquaculture Feed. Waste Biomass Valorization 2020, 11, 5307–5320. [Google Scholar] [CrossRef]
- Zainuddin, M.F.; Fai, C.K.; Ariff, A.B.; Rios-Solis, L.; Halim, M. Current Pretreatment/Cell Disruption and Extraction Methods Used to Improve Intracellular Lipid Recovery from Oleaginous Yeasts. Microorganisms 2021, 9, 251. [Google Scholar] [CrossRef] [PubMed]
- Khot, M.; Raut, G.; Ghosh, D.; Alarcón-Vivero, M.; Contreras, D.; Ravikumar, A. Lipid Recovery from Oleaginous Yeasts: Perspectives and Challenges for Industrial Applications. Fuel 2020, 259, 116292. [Google Scholar] [CrossRef]
- Menegazzo, M.L.; Fonseca, G.G. Biomass Recovery and Lipid Extraction Processes for Microalgae Biofuels Production: A Review. Renew. Sustain. Energy Rev. 2019, 107, 87–107. [Google Scholar] [CrossRef]
- Vargas-Sinisterra, A.F.; Ramírez-Castrillón, M. Yeast Carotenoids: Production and Activity as Antimicrobial Biomolecule. Arch. Microbiol. 2021, 203, 873–888. [Google Scholar] [CrossRef]
- Martínez, J.M.; Delso, C.; Aguilar, D.E.; Álvarez, I.; Raso, J. Organic-Solvent-Free Extraction of Carotenoids from Yeast Rhodotorula glutinis by Application of Ultrasound under Pressure. Ultrason. Sonochem. 2020, 61, 104833. [Google Scholar] [CrossRef]
- Liu, Z.; van den Berg, C.; Weusthuis, R.A.; Dragone, G.; Mussatto, S.I. Strategies for an Improved Extraction and Separation of Lipids and Carotenoids from Oleaginous Yeast. Sep. Purif. Technol. 2021, 257, 117946. [Google Scholar] [CrossRef]
- Rani, A.; Saini, K.C.; Bast, F.; Mehariya, S.; Bhatia, S.K.; Lavecchia, R.; Zuorro, A. Microorganisms: A Potential Source of Bioactive Molecules for Antioxidant Applications. Molecules 2021, 26, 1142. [Google Scholar] [CrossRef]
- Kultys, E.; Kurek, M.A. Green Extraction of Carotenoids from Fruit and Vegetable Byproducts: A Review. Molecules 2022, 27, 518. [Google Scholar] [CrossRef]
- Dall’armellina, A.; Letan, M.; Duval, C.; Contino-Pépin, C. One-Pot Solvent-Free Extraction and Formulation of Lipophilic Natural Products: From Curcuma to Dried Formulations of Curcumin. Green Chem. 2021, 23, 8891–8900. [Google Scholar] [CrossRef]
- Nemer, G.; Louka, N.; Vorobiev, E.; Salameh, D.; Nicaud, J.M.; Maroun, R.G.; Koubaa, M. Mechanical Cell Disruption Technologies for the Extraction of Dyes and Pigments from Microorganisms: A Review. Fermentation 2021, 7, 36. [Google Scholar] [CrossRef]
- Guzik, P.; Kulawik, P.; Zając, M.; Migdał, W. Microwave Applications in the Food Industry: An Overview of Recent Developments. Crit. Rev. Food Sci. Nutr. 2021, 62, 7989–8008. [Google Scholar] [CrossRef] [PubMed]
- Zbyradowski, M.; Duda, M.; Wisniewska-Becker, A.; Heriyanto; Rajwa, W.; Fiedor, J.; Cvetkovic, D.; Pilch, M.; Fiedor, L. Triplet-Driven Chemical Reactivity of β-Carotene and Its Biological Implications. Nat. Commun. 2022, 13, 2474. [Google Scholar] [CrossRef]
- Alvarez, R.; de Lera, A.R. Natural Polyenic Macrolactams and Polycyclic Derivatives Generated by Transannular Pericyclic Reactions: Optimized Biogenesis Challenging Chemical Synthesis. Nat. Prod. Rep. 2021, 38, 1136–1220. [Google Scholar] [CrossRef] [PubMed]
- Torres-Montilla, S.; Rodriguez-Concepcion, M. Making Extra Room for Carotenoids in Plant Cells: New Opportunities for Biofortification. Prog. Lipid Res. 2021, 84, 101128. [Google Scholar] [CrossRef]
- Molino, A.; Mehariya, S.; Di Sanzo, G.; Larocca, V.; Martino, M.; Leone, G.P.; Marino, T.; Chianese, S.; Balducchi, R.; Musmarra, D. Recent Developments in Supercritical Fluid Extraction of Bioactive Compounds from Microalgae: Role of Key Parameters, Technological Achievements and Challenges. J. CO2 Util. 2020, 36, 196–209. [Google Scholar] [CrossRef]
- Song, B.; Lin, R.; Lam, C.H.; Wu, H.; Tsui, T.H.; Yu, Y. Recent Advances and Challenges of Inter-Disciplinary Biomass Valorization by Integrating Hydrothermal and Biological Techniques. Renew. Sustain. Energy Rev. 2021, 135, 110370. [Google Scholar] [CrossRef]
- Zhang, L.; Liao, W.; Wei, Y.; Tong, Z.; Wang, Y.; Gao, Y. Fabrication, Characterization and in Vitro Digestion of Food-Grade β-Carotene High Loaded Microcapsules: A Wet-Milling and Spray Drying Coupling Approach. LWT 2021, 151, 112176. [Google Scholar] [CrossRef]
- Mussagy, C.U.; Santos-Ebinuma, V.C.; Kurnia, K.A.; Dias, A.C.R.V.; Carvalho, P.J.; Coutinho, J.A.P.; Pereira, J.F.B. Integrative Platform for the Selective Recovery of Intracellular Carotenoids and Lipids from Rhodotorula glutinis CCT-2186 Yeast Using Mixtures of Bio-Based Solvents. Green Chem. 2020, 22, 8478–8494. [Google Scholar] [CrossRef]
- Minyuk, G.S.; Solovchenko, A.E. Express Analysis of Microalgal Secondary Carotenoids by TLC and UV-Vis Spectroscopy. In Microbial Carotenoids: Methods and Protocols; Barreiro, C., Barredo, J.-L., Eds.; Springer: New York, NY, USA, 2018; pp. 73–95. ISBN 978-1-4939-8742-9. [Google Scholar]
- Llanto Revellame, M. Generation of Biodiesel and Carotenoids from Rhodotorula glutinis Using Sweet Sorghum Juice. Master’s Thesis, Mississippi State University, Starkville, MS, USA, 2012; p. 106. [Google Scholar]
- Rostami, H.; Hamedi, H.; Yolmeh, M. Some Biological Activities of Pigments Extracted from Micrococcus Roseus (PTCC 1411) and Rhodotorula glutinis (PTCC 5257). Int. J. Immunopathol. Pharmacol. 2016, 29, 684–695. [Google Scholar] [CrossRef] [PubMed]
- Popescu, M.; Iancu, P.; Pleșu, V.; Bîldea, C.S.; Todasca, C.M. Different Spectrophotometric Methods for Simultaneous Quantification of Lycopene and β-Carotene from a Binary Mixture. LWT 2022, 160, 113238. [Google Scholar] [CrossRef]
- USP. Beta Carotene Molecule. 2019. Available online: https://www.uspnf.com/sites/default/files/usp_pdf/EN/USPNF/revisions/beta-carotene.pdf (accessed on 20 May 2023).
- Suarez Ruiz, C.A.; Emmery, D.P.; Wijffels, R.H.; Eppink, M.H.M.; van den Berg, C. Selective and Mild Fractionation of Microalgal Proteins and Pigments Using Aqueous Two-Phase Systems. J. Chem. Technol. Biotechnol. 2018, 93, 2774–2783. [Google Scholar] [CrossRef] [PubMed]
- Di Caprio, F.; Altimari, P.; Pagnanelli, F. Sequential Extraction of Lutein and β-Carotene from Wet Microalgal Biomass. J. Chem. Technol. Biotechnol. 2020, 95, 3024–3033. [Google Scholar] [CrossRef]
- Su, W.H.; Sun, D.W. Fourier Transform Infrared and Raman and Hyperspectral Imaging Techniques for Quality Determinations of Powdery Foods: A Review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 104–122. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Li, Z.; Li, J.; Lin, H. Raman Spectroscopic Techniques for Nondestructive Analysis of Agri-Foods: A State-of-the-Art Review. Trends Food Sci. Technol. 2021, 118, 490–504. [Google Scholar] [CrossRef]
- National Research Council. NRC Nutrient Requirements of Domestics Animals: Nutrient Requirements of Poultry; National Academy Press: Washington, DC, USA, 1994. [Google Scholar]
- Khan, M.I.; Sameen, A. Animal Sourced Foods for Developing Economies: Preservation, Nutrition, and Safety; CRC Press: Boca Raton, FL, USA, 2018; p. 311. [Google Scholar]
- Alagawany, M.; Elnesr, S.S.; Farag, M.R.; El-Naggar, K.; Madkour, M. Nutrigenomics and Nutrigenetics in Poultry Nutrition: An Updated Review. Worlds. Poult. Sci. J. 2022, 78, 377–396. [Google Scholar] [CrossRef]
- Wu, G. Management of Metabolic Disorders (Including Metabolic Diseases) in Ruminant and Nonruminant Animals; Elsevier Inc.: Amsterdam, The Netherlands, 2019; ISBN 9780128170526. [Google Scholar]
- Lima, H.J.D.; Souza, L.A.Z. Vitamin A in the Diet of Laying Hens: Enrichment of Table Eggs to Prevent Nutritional Deficiencies in Humans. Worlds. Poult. Sci. J. 2018, 74, 619–626. [Google Scholar] [CrossRef]
- Jones, M.P. Avian Clinical Pathology. Vet. Clin. N. Am. Exot. Anim. Pract. 1999, 2, 663–687. [Google Scholar] [CrossRef]
- Cioffi, C.L.; Muthuraman, P.; Raja, A.; Varadi, A.; Racz, B.; Petrukhin, K. Discovery of Bispecific Antagonists of Retinol Binding Protein 4 That Stabilize Transthyretin Tetramers: Scaffolding Hopping, Optimization, and Preclinical Pharmacological Evaluation as a Potential Therapy for Two Common Age-Related Comorbidities. J. Med. Chem. 2020, 63, 11054–11084. [Google Scholar] [CrossRef]
- Garrido, T.; Uranga, J.; Guerrero, P.; de la Caba, K. The Potential of Vegetal and Animal Proteins to Develop More Sustainable Food Packaging. In Polymers for Food Applications; Gutiérrez, T.J., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 25–59. ISBN 978-3-319-94625-2. [Google Scholar]
- Dhakal, S.P.; He, J. Microencapsulation of Vitamins in Food Applications to Prevent Losses in Processing and Storage: A Review. Food Res. Int. 2020, 137, 109326. [Google Scholar] [CrossRef]
- McAllister, T.A.; Ribeiro, G.; Stanford, K.; Wang, Y. Forage-Induced Animal Disorders. Forages 2020, 2, 839–860. [Google Scholar] [CrossRef]
- Tsatsakis, A.; Tyshko, N.V.; Docea, A.O.; Shestakova, S.I.; Sidorova, Y.S.; Petrov, N.A.; Zlatian, O.; Mach, M.; Hartung, T.; Tutelyan, V.A. The Effect of Chronic Vitamin Deficiency and Long Term Very Low Dose Exposure to 6 Pesticides Mixture on Neurological Outcomes—A Real-Life Risk Simulation Approach. Toxicol. Lett. 2019, 315, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Cobb-Vantress Cobb 500 Broiler Performance & Nutrition Supplement. 2022. Available online: https://www.cobb-vantress.com/assets/5a88f2e793/Broiler-Performance-Nutrition-Supplement.pdf/ (accessed on 20 May 2023).
- Aviagen Ross 308 Broiler: Nutrition Specifications. 2022. Available online: https://en.aviagen.com/assets/Tech_Center/Ross_Broiler/Ross-BroilerHandbook2018-EN.pdf (accessed on 20 May 2023).
- Agri-Climate Rural Environment Scheme (ACRES). 2022. Available online: https://assets.gov.ie/231749/39dd9ce0-fb95-4619-bd8e-8ff53b752d08.pdf (accessed on 20 May 2023).
- Hubbard LLC. Breeder Nutrition Guide. 2020. Available online: https://www.hubbardbreeders.com/media/premium_guide_and_nutrient_specifications.pdf (accessed on 20 May 2023).
- Frohlich, U.; Wehrle, K.; Niederer, S. Management Guide. 2022. Available online: https://www.hyline.com/filesimages/Hy-Line-Products/Hy-Line-Product-PDFs/Brown/BRN%20COM%20ENG.pdf (accessed on 20 May 2023).
- ISA Brown. 2022. Available online: https://kenanaonline.com/files/0071/71989/ISA%20Brown%20Guide-Nov.%203,2010.pdf (accessed on 20 May 2023).
- Waddell, D. Management Guide: Alternative Systems. Lohmann Tiersucht 2005, 1, 1–68. [Google Scholar]
- Hybrid Turkey. Nutrient Guidelines. May 2023. Available online: https://www.hybridturkeys.com/en/resources/commercial-management/feed-and-water/nutrient-guidelines/ (accessed on 20 May 2023).
- Ciurescu, G.; Vasilachi, A.; Grosu, H. Efficacy of Microbial Phytase on Growth Performance, Carcass Traits, Bone Mineralization, and Blood Biochemistry Parameters in Broiler Turkeys Fed Raw Chickpea (Cicer arietinum L., Cv. Burnas) Diets. J. Appl. Poult. Res. 2020, 29, 171–184. [Google Scholar] [CrossRef]
- Zampiga, M.; Tavaniello, S.; Soglia, F.; Petracci, M.; Mazzoni, M.; Maiorano, G.; Meluzzi, A.; Clavenzani, P.; Sirri, F. Comparison of 2 Commercial Turkey Hybrids: Productivity, Occurrence of Breast Myopathies, and Meat Quality Properties. Poult. Sci. 2019, 98, 2305–2315. [Google Scholar] [CrossRef]
- Fouad, A.M.; Ruan, D.; Wang, S.; Chen, W.; Xia, W.; Zheng, C. Nutritional Requirements of Meat-Type and Egg-Type Ducks: What Do We Know? J. Anim. Sci. Biotechnol. 2018, 9, 1–11. [Google Scholar] [CrossRef]
- Kokoszyński, D.; Wasilewski, R.; Saleh, M.; Piwczyński, D.; Arpášová, H.; Hrnčar, C.; Fik, M. Growth Performance, Body Measurements, Carcass and Some Internal Organs Characteristics of Pekin Ducks. Animals 2019, 9, 963. [Google Scholar] [CrossRef]
- Estienne, A.; Brossaud, A.; Ramé, C.; Bernardi, O.; Reverchon, M.; Rat, C.; Delaveau, J.; Chambellon, E.; Helloin, E.; Froment, P.; et al. Chemerin Is Secreted by the Chicken Oviduct, Accumulates in Egg Albumen and Could Promote Embryo Development. Sci. Rep. 2022, 12, 8989. [Google Scholar] [CrossRef]
- Wen, Y.; Dai, B.; Zhang, X.; Zhu, H.; Xie, C.; Xia, J.; Sun, Y.; Zhu, M.; Tong, J.; Shen, Y. Retinal Transcriptomics Analysis Reveals the Underlying Mechanism of Disturbed Emmetropization Induced by Wavelength Defocus. Curr. Eye Res. 2022, 47, 908–917. [Google Scholar] [CrossRef]
- Maoka, T. Carotenoid Metabolism in Terrestrial Animals. In Carotenoids: Biosynthetic and Biofunctional Approaches; Misawa, N., Ed.; Springer: Singapore, 2021; pp. 51–66. ISBN 978-981-15-7360-6. [Google Scholar]
- Kamal, S.; Junaid, M.; Ejaz, A.; Bibi, I.; Bigiu, N. Eye Sight and Carotenoids. In Carotenoids: Structure and Function in the Human Body; Zia-Ul-Haq, M., Dewanjee, S., Riaz, M., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 609–647. ISBN 978-3-030-46459-2. [Google Scholar]
- Grashorn, M. Feed Additives for Influencing Chicken Meat and Egg Yolk Color; Elsevier Ltd.: Amsterdam, The Netherlands, 2016; ISBN 9780081003923. [Google Scholar]
- Merhan, O. The Biochemistry and Antioxidant Properties of Carotenoids. Carotenoids 2017, 5, 5. [Google Scholar] [CrossRef]
- Nogareda, C.; Moreno, J.A.; Angulo, E.; Sandmann, G.; Portero, M.; Capell, T.; Zhu, C.; Christou, P. Carotenoid-Enriched Transgenic Corn Delivers Bioavailable Carotenoids to Poultry and Protects Them against Coccidiosis. Plant Biotechnol. J. 2016, 14, 160–168. [Google Scholar] [CrossRef]
- Yamamoto, T.; Juneja, L.R.; Hatta, H.; Kim, M. Hen Eggs: Basic and Applied Science; CRC Press: Boca Raton, FL, USA, 2018; p. 217. [Google Scholar]
- Langi, P.; Kiokias, S.; Varzakas, T.; Proestos, C. Carotenoids: From Plants to Food and Feed Industries. In Microbial Carotenoids: Methods and Protocols; Barreiro, C., Barredo, J.-L., Eds.; Springer: New York, NY, USA, 2018; pp. 57–71. ISBN 978-1-4939-8742-9. [Google Scholar]
- Council Directive 96/23/EC V. 2. Nurse Responsible for General Care This Document Is Meant Purely as a Documentation Tool and the Commission and Its Services Do Not Assume Any Liability for Its Contents. MachineryDirective 2010, 66, 1–27.
- Hill, G.E.; Johnson, D.J. The vitamin A–redox hypothesis: A biochemical basis for honest signaling via carotenoid pigmentation. Am. Nat. 2012, 180, E127–E150. [Google Scholar] [CrossRef]
- Index of Food Colours | Safefood. Available online: https://www.safefood.net/food-colour-index (accessed on 6 July 2022).
- Dansou, D.M.; Wang, H.; Nugroho, R.D.; He, W.; Zhao, Q.; Zhang, J. Assessment of Response to Moderate and High Dose Supplementation of Astaxanthin in Laying Hens. Animals 2021, 11, 1138. [Google Scholar] [CrossRef]
- Xue, X.; Fan, L.; Dong, Y.; Yuan, X.; Wang, L.; Yang, F.; Zheng, Y.; Zhao, S. Evaluation of Canthaxanthin in Eggs and Its Subsequent Dietary Risks to Chinese Consumers. Food Addit. Contam. Part A 2021, 38, 255–260. [Google Scholar] [CrossRef]
- Vranic, D.; Koricanac, V.; Milicevic, D.; Djinovic-Stojanovic, J.; Geric, T.; Lilic, S.; Petrovic, Z. Nitrite Content in Meat Products from the Serbian Market and Estimated Intake. IOP Conf. Ser. Earth Environ. Sci. 2021, 854, 012106. [Google Scholar] [CrossRef]
- Ortiz, D.; Lawson, T.; Jarrett, R.; Ring, A.; Scoles, K.L.; Hoverman, L.; Rocheford, E.; Karcher, D.M.; Rocheford, T. Biofortified Orange Corn Increases Xanthophyll Density and Yolk Pigmentation in Egg Yolks from Laying Hens. Poult. Sci. 2021, 100, 101117. [Google Scholar] [CrossRef] [PubMed]
- Erasmus, L.J.; Machpesh, G.; Coertze, R.J.; du Toit, C.J.L. Effects of Feeding System and Pre-Partum Supplementation on the β-Carotene Status of South African Holstein Cows. S. Afr. J. Anim. Sci. 2021, 51, 339–348. [Google Scholar] [CrossRef]
- Mavrommatis, A.; Zografaki, M.E.; Marka, S.; Myrtsi, E.D.; Giamouri, E.; Christodoulou, C.; Evergetis, E.; Iliopoulos, V.; Koulocheri, S.D.; Moschopoulou, G.; et al. Effect of a Carotenoid Extract from Citrus Reticulata By-Products on the Immune-Oxidative Status of Broilers. Antioxidants 2022, 11, 144. [Google Scholar] [CrossRef]
- Mishra, B.; Varjani, S.; Karthikeya Srinivasa Varma, G. Agro-Industrial By-Products in the Synthesis of Food Grade Microbial Pigments: An Eco-Friendly Alternative. In Green Bio-Processes: Enzymes in Industrial Food Processing; Parameswaran, B., Varjani, S., Raveendran, S., Eds.; Springer: Singapore, 2019; pp. 245–265. ISBN 978-981-13-3263-0. [Google Scholar]
- Luiza Koop, B.; Nascimento da Silva, M.; Diniz da Silva, F.; Thayres dos Santos Lima, K.; Santos Soares, L.; José de Andrade, C.; Ayala Valencia, G.; Rodrigues Monteiro, A. Flavonoids, Anthocyanins, Betalains, Curcumin, and Carotenoids: Sources, Classification and Enhanced Stabilization by Encapsulation and Adsorption. Food Res. Int. 2022, 153, 110929. [Google Scholar] [CrossRef]
- Ashour, E.A.; Farsi, R.M.; Alaidaroos, B.A.; Abdel-Moneim, A.M.E.; El-Saadony, M.T.; Osman, A.O.; Abou Sayed-Ahmed, E.T.; Albaqami, N.M.; Shafi, M.E.; Taha, A.E.; et al. Impacts of Dietary Supplementation of Pyocyanin Powder on Growth Performance, Carcase Traits, Blood Chemistry, Meat Quality and Gut Microbial Activity of Broilers. Ital. J. Anim. Sci. 2021, 20, 1357–1372. [Google Scholar] [CrossRef]
- Kanwugu, O.N.; Ranga Rao, A.; Ravishankar, G.A.; Glukhareva, T.V.; Kovaleva, E.G. (Eds.) Chapter 31—Astaxanthin from Bacteria as a Feed Supplement for Animals; Academic Press: Cambridge, MA, USA, 2021; pp. 647–667. ISBN 978-0-12-823304-7. [Google Scholar]
- Elwan, H.A.M.; Elnesr, S.S.; Abdallah, Y.; Hamdy, A.; El-Bogdady, A.H. Red Yeast (Phaffia rhodozyma) as a Source of Astaxanthin and Its Impacts on Productive Performance and Physiological Responses of Poultry. Worlds Poult. Sci. J. 2019, 75, 273–284. [Google Scholar] [CrossRef]
- Xu, Y.; Li, J.; Zhao, J.; Wang, W.; Griffin, J.; Li, Y.; Bean, S.; Tilley, M.; Wang, D. Hempseed as a Nutritious and Healthy Human Food or Animal Feed Source: A Review. Int. J. Food Sci. Technol. 2021, 56, 530–543. [Google Scholar] [CrossRef]
- Coulombier, N.; Jauffrais, T.; Lebouvier, N. Antioxidant Compounds from Microalgae: A Review. Mar. Drugs 2021, 19, 549. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.M.; Sun, Y.L.; Lin, C.H.; Lin, C.H.; Wu, H.T.; Lin, C.S. Cultivation and Biorefinery of Microalgae (Chlorella) sp. for Producing Biofuels and Other Byproducts: A Review. Sustain. 2021, 13, 3480. [Google Scholar] [CrossRef]
- Phan, M.A.T.; Bucknall, M.; Arcot, J. Interactive Effects of β-Carotene and Anthocyanins on Cellular Uptake, Antioxidant Activity and Anti-Inflammatory Activity in Vitro and Ex Vivo. J. Funct. Foods 2018, 45, 129–137. [Google Scholar] [CrossRef]
- Sayahi, M.; Shirali, S. The Antidiabetic and Antioxidant Effects of Carotenoids: A Review. Asian J. Pharm. Res. Health Care 2017, 9, 186–191. [Google Scholar] [CrossRef]
- Ribeiro, D.; Freitas, M.; Silva, A.M.S.; Carvalho, F.; Fernandes, E. Antioxidant and Pro-Oxidant Activities of Carotenoids and Their Oxidation Products. Food Chem. Toxicol. 2018, 120, 681–699. [Google Scholar] [CrossRef]
- Molnár, S.; Szollosi, L. Sustainability and Quality Aspects of Different Table Egg Production Systems: A Literature Review. Sustainability 2020, 12, 7884. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Microorganism | Carotenoid | Structure | Reference |
|---|---|---|---|
| Funghi | |||
| Neurospora crassa | β-carotene |  | [20] |
| Monascus sp. | Monascorubramin |  | [21] |
| Blakeslea trispora | Lycopene |  | [22] |
| Fusarium sporotrichioides | Lycopene |  | [23] |
| Aspergillus sp. | β-carotene |  | [24] |
| Pacilomyces farinosus | Anthraquinone |  | [25] |
| Bacteria | |||
| Paracoccus carotinifaciens | Astaxanthin |  | [26] |
| Staphylococcus aureus | Zeaxanthin |  | [27] |
| Zooshikella sp. | Prodigiosin |  | [28] |
| Serratia marcescens | Prodigiosin |  | [29] |
| Yeast | |||
| Rhodotorula glutinis | Torularhodin |  | [30] |
| Xanthophyllomyces dendrorhous | Astaxanthin |  | [31] |
| Rhodototula mucilaginosa | β-carotene |  | [32] |
| Saccharomyces neoformans | Melanin |  | [33] |
| Yeast Strain | Assay Conditions and Results | Reference |
|---|---|---|
| Oil←lipids→fatty acids | ||
| Rh. mucilaginosa IIPL32 | Fed-batch C/N ratio = 40; Fed-batch C/N ratio = 60; Scalability: 50 mL→50 L; lipid yield for C/N ratio fed-batch = 40: 0.4 g→1.3 g/L lipid yield for C/N ratio fed-batch = 60: 0.45 g→1.8 g/L | [50] |
| Rh. mucilaginosa IIPL32 | (Lipids as FAME) 72 h→97.23 mg/g dry cell weight; 35–55%, MUFA C18:1 and C16:1 (oleic and palmitoleic acids) | [51] |
| Rh. mucilaginosa CCT3892 | The total amount of lipids obtained in the molasses medium was similar to the synthetic medium (15.36% ± 1.36% and 16.50% ± 0.68%, respectively), thus, the production of the metabolites was higher in the molasses medium. | [52] |
| Enzymes | ||
| Rhodotorula mucilaginosa CBMAI 1528 | Aspartic protease—pepsin family | [53] |
| Rhodotorula mucilaginosa | Invertase—the invertase with greater cell-structural stability and nystose productivity | [54] |
| Rhodotorula sp. Y-23 | Lipase (Lip-Y23)—low-temperature applications | [42] |
| R. mucilaginosa Y-1 | Carboxylase—Acetyl coenzyme A carboxylase (ACC1) | [55] |
| Carotenoids | ||
| Rhodotorula glutinis | β-carotene, torularhodin | [47,56] |
| Rhodotorula mucilaginosa KC8 | β-carotene, torularhodin | [57] |
| Species | Factors Affecting Pigmentation Capacity and Productivity | Results | References | |
|---|---|---|---|---|
| Mutagenesis | ||||
| R. toruloides NP11 | Atmospheric exposure, 30 °C and plasma technique followed by chemical mutagenesis with nitrosoguanidine | R. toruloides XR-2, colonies of dark-red colored Nitrogen limitation conditions induced slower growth with high carotenoid yields | [84] | |
| Rhodotorula sp. Strains | T-DNA insertional mutagenesis gene discovery in R. toruloids | Twenty-seven mutant yeast phenotypes for lipid and carotenoid metabolism | [81] | |
| R. toruloides NP11 | An Agrobacterium tumefaciens-mediated transformation (ATMT) to change its carotenoid production and profiles | Selected three new phenotypes and mutants with different colors Characterized their carotenoid products | [86] | |
| Rhodotorula toruloides CBS 14 and NCYC 1585 | Cloning strategy; four inducible promoters for control gene expression in Rhodotorula toruloides to obtain molecular genetic tools for manipulation | Directed genetic and gene expression for carotenoid and lipid yields in Rhodotorula toruloides | [87] | |
| Cultivation medium | ||||
| Carbon | Nitrogen | |||
| Rhodotorula sp. | - | Threonine (0.1, 0.2, 0.3%) glutamic acid (0.1, 0.2, 0.3%) | Both amino acid stimulation enhanced yeast growth parameters and total carotenoid formation | [76] |
| Rhodotorula mucilaginosa | Waste from the olive oil industry (Alperujo water, AE) in different aqueous solutions at concentrations: of 5, 10, 20, and 30% | The volumetric carotenoid production significantly increases in 20 and 30% AE concentration (up to 7.3 ± 0.6 mg/L total carotenoids) | [67] | |
| Rhodotorula sp. RY1801 | Sucrose, lactose, maltose, fructose and glucose | Inorganic nitrogen: ammonium sulfate, ammonium nitrate; Organic nitrogen: yeast extract, urea | Carbon sources: glucose; carotene yield up to 962 µg/L Nitrogen sources: yeast extract; carotene yield up to 987 µg/L | [83] |
| Rhodotorula glutinis (AS 2.703) | - | Peptone (PEP), yeast extract (YE), and ammonium sulfate | The highest biomass accumulation was 12.2 g/L after 144 h (YE) | [88] |
| Rhodotorula mucilaginosa | Onion and potato (skin) | Mung bean (husk) and pea (pods) | The highest carotenoid yield was archived by using onion peel extract and mung bean (up to 717.82 μg/g) | [79] |
| Lightning conditions | ||||
| Rhodotorula mucilaginosa | Irradiation UV-C—254 nm | Metabolite production by psychro-tolerant Rhodotorula mucilaginosa produced up to 56.9 ± 3.2 (μg/g−1.dry weight) of total carotenoids | [89] | |
| R. glutinis (CGMCC No. 2258) | LED lamp’s light exposure intensities (4000 lx and 8000 lx) | The lipid and β-carotene production enhancement by using light exposure and sodium acetate componence substrate in R. glutinis | [56] | |
| R. mucilaginosa K-(1) | Two lighting shakers: Shaker 1: 1700 lx; Shaker 2: 3500 lx. Settled as 12 h dark: 12 h light | Illumination intensity increases the carotenoid yield (1700 lx); High illuminating intensity (3200 lx) inhibits yeast glucose metabolism. Thus, cell growth | [77] | |
| R. glutinis (CGMCC No. 2258) | Two groups: without and with continuous irradiation (3400 lx) | Continuous irradiation might positively affect the lipid and carotenoid content | [90] | |
| Rhodotorula mucilaginosa | Stress conditions: ultraviolet (UV) light and photoperiods | Optimum conditions for stimulating the carotenoid productivity were 1 min of UV exposure combined with 0.5 mg/L magnesium sulfate and 18:6 h lighting conditions | [91] | |
| Thermic conditions | ||||
| Rhodotorula sp. RY1801 | Incubation temperature ranges from 20 to 37 °C | Optimum incubation temperature at 28 °C | [83] | |
| Rhodotorula glutinis | Incubation temperature ranges from 25, 30, 35 to 40 °C | Optimum incubation temperature at 30 °C | [92] | |
| Rhodotorula mucilaginosa | Incubation temperatures 15, 20, 25, 28, 30, 35, and 40 °C | The most suitable temperature for culture growth and carotenoid production was 28 °C | [93] | |
| R. mucilaginosa ATCC 66034 and R. gracilis ATCC 10788 | Incubation temperature 20 °C and 28 °C | Optimum incubation temperature at 20 °C | [39] | |
| [74] | Low temperature (16 °C) treatment Control temperature (25 °C) treatment | At 16 °C, the carotenoid yield was significantly increased | [75] | |
| Aeration conditions | ||||
| Rhodotorula glutinis NRRL Y-12905 | Different conditions of agitation (150 to 250 rpm) and aeration [(2.5 to 5.0 of flask volume-to-medium volume ratio (vvm)] | Agitation and aeration at 250 rpm and 5.0 optimal conditions (high yeast cell concentration) | [78] | |
| Rhodotorula rubra PTCC 5255 | Aeration levels: 0.115, 0.345 and 0575 vvm | The optimum carotenoid concentration was found at an aeration rate of 0.469 vvm, having the substrate initial pH of 6.48, and light intensity of 1757.84 lx | [94] | |
| Rhodotorula mucilaginosa MTCC-1403 | Different conditions of agitation: 80, 110, and 140 rpm | Elevation of up to 100 μg carotenoids per g of dry biomass | [79] | |
| Metabolizable salts and microelements addition | ||||
| Rhodotorula glutinis CCT 2186 | Different experimental levels of: glucose, KH2PO4, MgSO4, NH4NO3, and pH | Combined sources of inorganic and organic nitrogen sources had high productivity yields | [45] | |
| Broiler Chicks | |||||
|---|---|---|---|---|---|
| Hybrid | 0–11 Days | 12–23 Days | 24–42 Days | References | |
| Cobb 500 | Up to13,000 | 10,000 | 10,000 | [146] | |
| Ross 308 | 13,000 | 13,000 | 13,000 | [147] | |
| Arbor acres | 13,000 | 10,000 | 10,000 | [148] | |
| Hubbard | 13,000 | 13,000 | 13,000 | [149] | |
| Laying Hens | |||||
| Hybrid | 0–6 Weeks | 7–12 Weeks | 12–18 Weeks | >18 Weeks | |
| Hy-line W36 | 5700 | 5700 | 5700 | 5700 | [150] |
| ISA chick | 15,000 | 15,000 | 13,500 | 13,500 | [151] |
| Lohmann | 10,000 | 10,000 | 10,000 | 10,000 | [152] |
| Turkeys | |||||
| Hybrid | 0–42 Weeks | 43–84 Weeks | >84 Weeks | ||
| Hybrid Grade Maker male turkey | 9000–12,000 | 9000–12,000 | 8000–11,000 | [153,154] | |
| Hybrid meat turkey | 13,500 | 12,000 | 11,000 | [155] | |
| Ducks | |||||
| Hybrid | Starter | Grower/Finisher | |||
| Longyan laying ducks | 10,000 | 8000/12,000 | [156] | ||
| Pekin | 10,000 | 10,000 | [157] | ||
| Pigment Additive 1 | Code 2 | Meat * | Egg * | Origin |
|---|---|---|---|---|
| β-Apo-8′-carotenal | E 161e | 80 | 80 | artificial |
| Cryptoxanthin | E 161b | 80 | 80 | natural |
| Lutein | E 161b | 80 | 80 | natural |
| Ethyl ester of β-apo-8′-carotenoid acid | E 160f | 80 | 80 | artificial |
| Zeaxanthin | E 161h | 80 | 80 | natural |
| Violaxanthin | E 161e | 80 | 80 | natural |
| Citranaxanthin | E 161i | - | 80 | artificial |
| Canthaxanthin | E 1601g | 25 | 8 | artificial |
| Capsanthin | E 160c | 80 | 80 | natural |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grigore, D.-M.; Ungureanu-Iuga, M.; Pogurschi, E.N.; Băbeanu, N.E. Transforming Rhodotorula sp. Biomass to Active Biologic Compounds for Poultry Nutrition. Agriculture 2023, 13, 1159. https://doi.org/10.3390/agriculture13061159
Grigore D-M, Ungureanu-Iuga M, Pogurschi EN, Băbeanu NE. Transforming Rhodotorula sp. Biomass to Active Biologic Compounds for Poultry Nutrition. Agriculture. 2023; 13(6):1159. https://doi.org/10.3390/agriculture13061159
Chicago/Turabian StyleGrigore, Daniela-Mihaela, Mădălina Ungureanu-Iuga, Elena Narcisa Pogurschi, and Narcisa Elena Băbeanu. 2023. "Transforming Rhodotorula sp. Biomass to Active Biologic Compounds for Poultry Nutrition" Agriculture 13, no. 6: 1159. https://doi.org/10.3390/agriculture13061159
APA StyleGrigore, D.-M., Ungureanu-Iuga, M., Pogurschi, E. N., & Băbeanu, N. E. (2023). Transforming Rhodotorula sp. Biomass to Active Biologic Compounds for Poultry Nutrition. Agriculture, 13(6), 1159. https://doi.org/10.3390/agriculture13061159