
Predicting the Occurrence and Risk Damage Caused by the Two-Spotted Spider Mite Tetranychus urticae (Koch) in Dry Beans (Phaseolus vulgaris L.) Combining Rate and Heat Summation Models for Digital Decisions Support

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
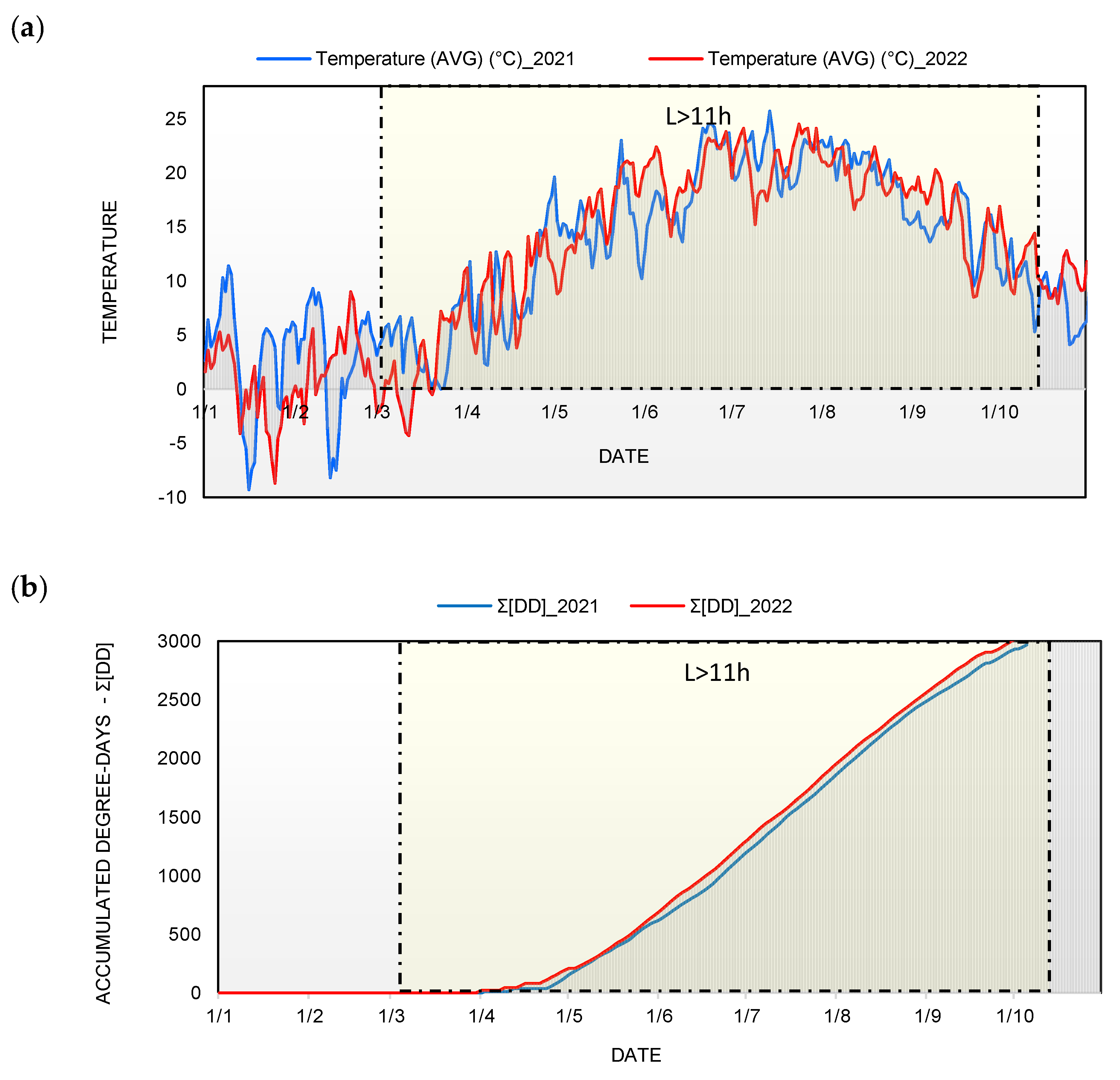
2.1. Study Site, Weather Data, and Heat Summations
2.2. T. urticae Leaf Infestation, Sampling, and Damage Assessment
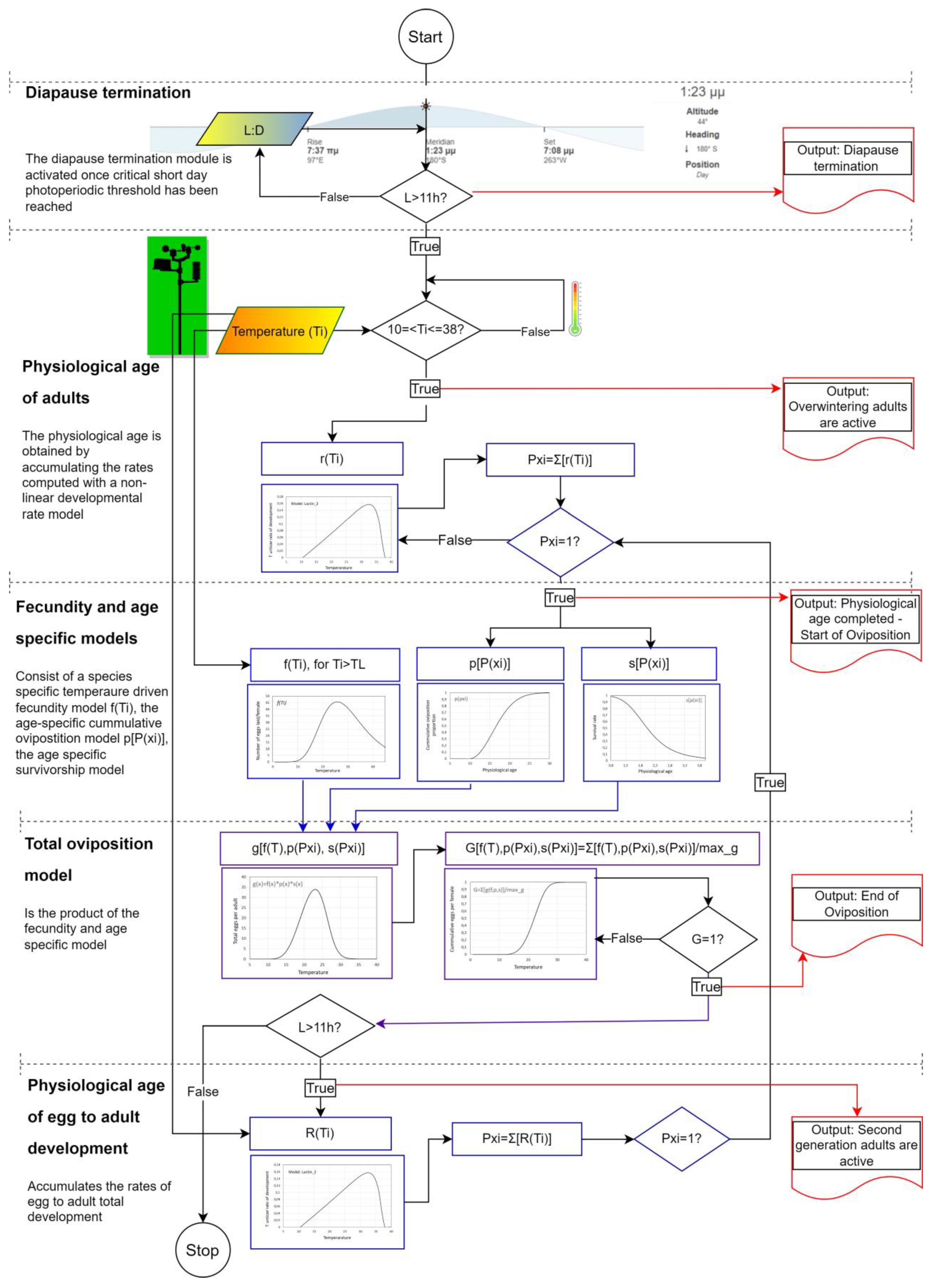
2.3. Modeling the Occurrence of T. urticae
2.3.1. Rate Summation Modeling of T. urticae
2.3.2. Physiological Age
2.3.3. Temperature-Dependent Fecundity Model
2.3.4. Age-Specific Cumulative Oviposition and Survivorship Rate Models
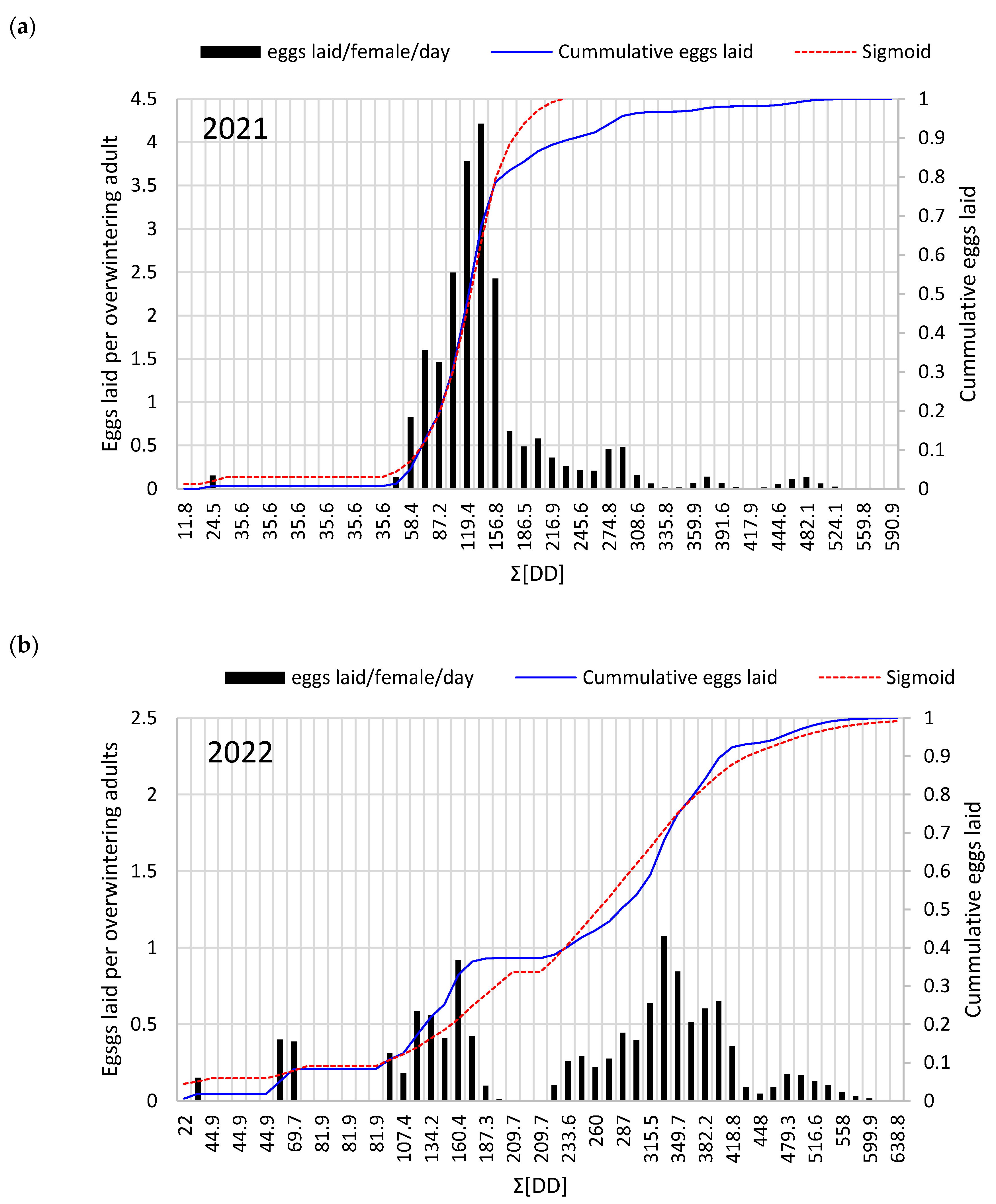
2.3.5. Total Oviposition Model and Cumulative Eggs Laid by T. urticae
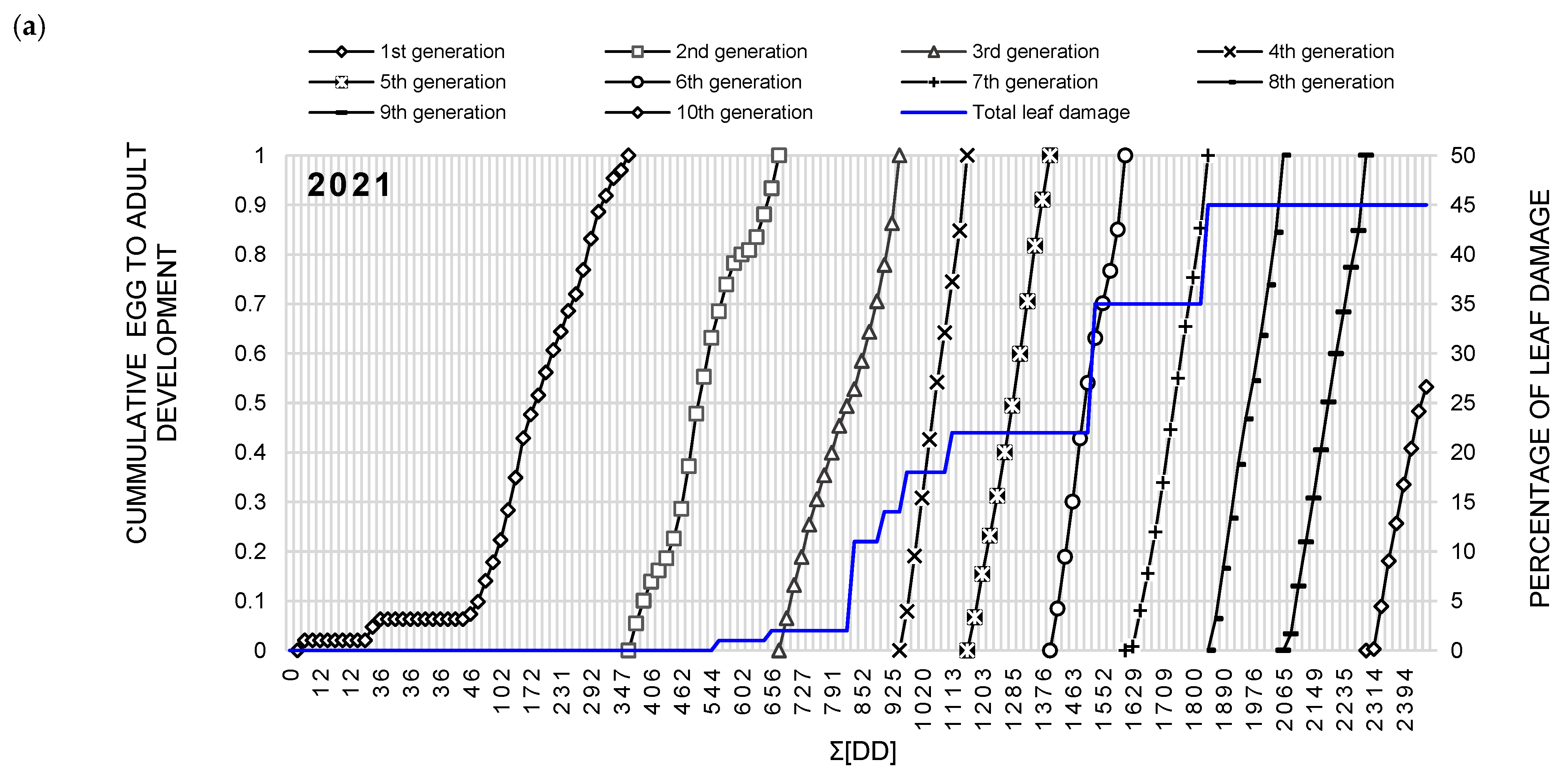
2.3.6. Heat Summation Modeling of T. urticae
3. Results
3.1. Suitability of Weather Conditions and Heat Summations
3.2. Overall Model Struction and Model Flow
3.3. T. urticae Diapause Termination and Female Total Oviposition Patterns
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Uebersax, M.A.; Cichy, K.A.; Gomez, F.E.; Porch, T.G.; Heitholt, J.; Osorno, J.M.; Kamfwa, K.; Snapp, S.S.; Bales, S. Dry beans (Phaseolus vulgaris L.) as a vital component of sustainable agriculture and food security—A review. Legume Sci. 2022, 5, e155. [Google Scholar] [CrossRef]
- Papathanasiou, F.; Ninou, E.; Mylonas, I.; Baxevanos, D.; Papadopoulou, F.; Avdikos, I.; Sistanis, I.; Koskosidis, A.; Vlachostergios, D.N.; Stefanou, S.; et al. The Evaluation of Common Bean (Phaseolus vulgaris L.) Genotypes under Water Stress Based on Physiological and Agronomic Parameters. Plants 2022, 11, 2432. [Google Scholar] [CrossRef] [PubMed]
- Hellenic Statistical Authority. 2019. Available online: https://www.statistics.gr/ (accessed on 1 October 2022).
- Vakali, C.; Baxevanos, D.; Vlachostergios, D.N.; Tamoutsidis, E.; Papathanasiou, F.; Papadopoulos, I. Genetic characterization of agronomic, physiochemical, and quality parameters of dry bean landraces under low-input farming. J. Agric. Sci. Technol. 2017, 19, 757–767. [Google Scholar]
- Prespa: Agriculture and Environment, Society for the Protection of Prespa (SPP). 2009. Available online: https://www.spp.gr/oemn%20en.pdf (accessed on 5 January 2023).
- Damos, P.; Tsikos, E.; Louta, M.; Papathanasiou, F. Towards the development of a smart plant protection solution for improved pest management of dry beans (Phaseolus vulgaris L.) in Northern Greece Proceedings. Proceedings 2021, 68, x. [Google Scholar]
- Bolland, H.R.; Gutierrez, J.; Flechtmann, C.H.W. (Eds.) World Catalogue of the Spider Mite Family (Acari:Tetranychidae); Brill: Leiden, The Netherlands, 1998. [Google Scholar]
- Jakubowska, M.; Dobosz, R.; Zawada, D.; Kowalska, J. A Review of Crop Protection Methods against the Twospotted Spider Mite—Tetranychus urticae Koch (Acari: Tetranychidae)—With Special Reference to Alternative Methods. Agriculture 2022, 12, 898. [Google Scholar] [CrossRef]
- Ahmadi, M.; Fathipour, Y.; Kamali, K. Population growth parameters of Tetranychus urticae (Acari: Tetranychidae) on different bean varieties. J. Entomol. Soc. Iran 2007, 26, 1–10. [Google Scholar]
- Laing, J.E. Life history and life table of Tetranychus urticae Koch. Acarologia 1969, 11, 32–42. [Google Scholar]
- Puspitarini, R.D.; Fernando, I.; Rachmawati, R.; Hadi, M.S.; Rizali, A. Host plant variability affects the development and reproduction of Tetranychus urticae. Int. J. Acarol. 2021, 47, 381–386. [Google Scholar] [CrossRef]
- Gotoh, T. Termination pattern of diapauze in Tetranychus urticae Koch (Acari: Tetranychidae) in Sapporo. Appl. Entomol. Zool. 1986, 21, 480–481. [Google Scholar] [CrossRef]
- Vacante, V. The Handbook of Mites of Economic Plants: Identification, Bio-Ecology and Control; CABI International: Wallingford, UK, 2016; pp. 1–890. [Google Scholar]
- Damos, P.; Papathanasiou, F.; Tsikos, E.; Kyriakidis, T.; Louta, M. Bayesian Non-Parametric Thermal Thresholds for Helicoverpa armigera and Their Integration into a Digital Plant Protection System. Agronomy 2022, 12, 2474. [Google Scholar] [CrossRef]
- Köppen, W. Klassification der Klimate nach Temperatur, Niederschlag and Jahreslauf. Petermanns Geogr. Mitt. 1918, 64, 193–203, 243–248. [Google Scholar]
- Riahi, E.; Shishehbor, P.; Nemati, A.N.; Saeidi, Z. Temperature Effects on Development and Life TableParameters of Tetranychus urticae (Acari: Tetranychidae). J. Agr. Sci. Technol. 2013, 15, 661–672. [Google Scholar]
- Singh, R.; Mangat, N.S. Stratified Sampling. In Elements of Survey Sampling. Kluwer Texts in the Mathematical Sciences; Springer: Dordrecht, Germany, 1996; Volume 15. [Google Scholar]
- Steven, K.; Thompson, S.K. Stratified sampling. In Sampling; Wiley Series in Probability and Statistics; Walter, A.S., Wilks, S.S., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Garrett, G. In Encyclopedia of Social Measurement. 2005. Available online: https://www.sciencedirect.com/topics/mathematics/stratified-sample (accessed on 18 November 2022).
- Morrill, W.L.; Shepard, B.M. Methods for Measuring Crop Losses by Insects. In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer: Dordrecht, Germany, 2008. [Google Scholar] [CrossRef]
- Wagner, T.L.; Wu, H.I.; Feldman, R.M.; Share, P.J.H.; Coulson, R.N. Multiple-cohort Approach for Simulating Development of Insect Populations Under Variable Temperatures. Ann. Entomol. Soc. Am. 1985, 78, 691–704. [Google Scholar] [CrossRef]
- Ghosh, J.B. A rate summation model of temperature dependent development with stochastic extensions. Math. Model. 1987, 8, 170–171. [Google Scholar] [CrossRef]
- Koveos, D.S.; Kroon, A.; Veerman, A. Geographical variation of diapause intensity in the spider mite Tetranychus urticae. Physiol. Entomol. 1993, 18, 50–56. [Google Scholar] [CrossRef]
- Lactin, D.J.; Holliday, N.J.; Johnson, D.L.; Craigen, R. Improved rate model of temperature-dependentdevelopment by arthropods. Environ. Entomol. 1995, 24, 68–75. [Google Scholar] [CrossRef]
- Stinner, R.E.; Gutierrez, A.P.; Butler, G.D., Jr. An algorithm for temperature depended growth rare simulation. Can. Entomol. 1974, 106, 519–524. [Google Scholar]
- Wagner, T.; Hsin, W.; Sharpe, P.; Schoolfield, R.M.; Coulson, R. Modeling insect developmental rates: A literature review and application of a biophysical model. Ann. Entomol. Soc. Amer. 1984, 77, 208–220. [Google Scholar] [CrossRef]
- Vangansbeke, D.; Audenaert, J.; Nguyen, D.T.; Verhoeven, R.; Gobin, B.; Tirry, L.; De Clercq, P. Diurnal Temperature Variations Affect Development of a Herbivorous Arthropod Pest and its Predators. PLoS ONE 2015, 10, e0124898. [Google Scholar] [CrossRef]
- Farazmand, A. Effect of the temperature on development of Tetranychus urticae (Acari: Tetranychidae) feeding on cucumber leaves. Int. J. Acarol. 2020, 46, 381–386. [Google Scholar] [CrossRef]
- Kim, D.S.; Lee, J. Oviposition model of overwintering adult Tetranychus urticae (Acari: Tetranychidae) and mite phenology on the ground cover in apple orchards. Exp. Appl. Acarol. 2003, 31, 191–2008. [Google Scholar] [CrossRef]
- Curry, C.L.; Feldman, R.M. Mathematical Foundations of Population Dynamics; Texas A&M University Press: College Station, TX, USA, 1987; 240p. [Google Scholar]
- Kim, J.S.; Jung, C.; Lee, J.H. Parameter estimation fort the temperature-dependent development model of Tetranychus urticae Koch: Immature development. J. Asia-Pac. Entomol. 2001, 4, 123–129. [Google Scholar] [CrossRef]
- Mack, T.P.; Smith, J.W., Jr. Modeling insect recruitment. In Basics of Insect Modeling; Goodenough, J.L., McKinion, J.M., Eds.; American Society of Agricultural Engineers: St. Joseph, MI, USA, 1992; pp. 155–169. [Google Scholar]
- Mack, T.P.; Smith, J.W., Jr.; Reed, R.B. Mathematical model of the population dynamics of the lesser cornstalk borer, Elasmopalpus lignosellus (Lepidoptera: Pyralidae). Ecol. Modell. 1987, 39, 267–286. [Google Scholar] [CrossRef]
- Birley, M. The estimation of insect density and instar survivorship functions from census data. J. Anim. Ecol. 1977, 46, 497–510. [Google Scholar] [CrossRef]
- Clements, A.N.; Paterson, G.D. The analysis of mortality and survival rates in wild populations of mosquitoes. J. Appl. Ecol. 1981, 18, 373–399. [Google Scholar] [CrossRef]
- Readshaw, J.L.; van Gerwen, A.C.M. Age-specific survival, fecundity and fertility of the adult blow fly in relation to crowding, protein food and population cycles. J. Anim. Ecol. 1983, 52, 879–887. [Google Scholar] [CrossRef]
- Bartlett, P.W.; Murray, A.W.A. Modeling adult survival in the laboratory of diapause and nondiapause colorado beetle, Leptinotarsa decemlineata (Coleoptera: Chrysomelidae) from Normandy, France. Ann. Appl. Biol. 1986, 108, 487–501. [Google Scholar] [CrossRef]
- Madden, L.V.; Nault, L.R.; Heady, S.E.; Styer, W.E. Effect of temperature on the population dynamics of three Dalbulus leafhopper species. Ann. Appl. Biol. 1986, 108, 475–485. [Google Scholar] [CrossRef]
- Marchiorio, C.A.; Foerster, L.A. Modeling of Plutella xylostella L. (Lepidoptera: Plutellidae): Climate change may modify pest incidence levels. Bul. Entomol. Res. 2012, 102, 489–496. [Google Scholar] [CrossRef]
- Choi, K.S.; Samayoa, A.C.; Hwang, S.Y.; Huang, Y.B.; Ahn, J.J. Thermal effect on the fecundity and longevity of Bactrocera dorsalis adult and their improved oviposition model. PLoS ONE 2020, 15, e0235910. [Google Scholar] [CrossRef]
- Damos, P.; Savopoulou-Soultani, M. Development and statistical evaluation of models in forecasting major lepidopterous peach pest complex for integrated pest management programs. Crop Prot. 2010, 29, 1190–1199. [Google Scholar] [CrossRef]
- Goodsman, D.W.; Aukema, B.H.; McDowell, N.G.; Middleton, R.S.; Xu, C. Incorporating variability in simulations of seasonally forced phenology using integral projection models. Ecol. Evol. 2018, 8, 162–175. [Google Scholar] [CrossRef]
- Jeppson, L.R.; Keifer, H.H.; Baker, E.W. Mites Injurious to Economic Plants; University of California Press: Berkeley, CA, USA, 1975. [Google Scholar]
- Takafuji, A.; Ozawa, A.; Nemoto, H.; Gotoh, T. Spider Mites of Japan: Their Biology and Control. Exp. Appl. Acarol. 2000, 24, 319–335. [Google Scholar] [CrossRef]
- Pakyari, H.; Amir-Maafi, M.; Dong-Soon, K.; Enkegaard, A. Oviposition Model of Scolothrips longicornis Fed on Two-Spotted Spider Mite. Acad. J. Entomol. 2012, 5, 65–72. [Google Scholar]
- Herbert, H.J. Biology, life tables, and innate capacity for increase of the twospotted spider mite, Tetranychus urticae (Acarina: Tetranychidae). Can. Ent. 1982, 113, 371–378. [Google Scholar] [CrossRef]
- Carey, J.R.; Bradley, J.W. Developmental rates, vital schedules, sex ratios, and life tables for Tetranychus urticae, T. turkestani and T. pacificus (Acarina: Tetranychidae) on cotton. Acarologia 1982, 23, 333–345. [Google Scholar]
- Karami-Jamour, T.; Shishehbor, P. Development and life table parameters of Tetranychus turkestani (Acarina: Tetranychidae) at different constant temperatures. Acarologia 2012, 52, 113–122. [Google Scholar] [CrossRef]
- Ju, K.; Sangkoo, L.; JeongMan, K.; YoungRip, K.; TaeHeung, K.; Jisoo, K. Effect of Temperature on Development and Life Table Parameters of Tetranychus urticae Koch (Acari:Tetranychide) Reared on Eggplants. Korean J. Appl. Entomol. 2008, 47, 163–168. [Google Scholar]
- Rott, A.S.; Ponsonby, D.J. The effects of temperature, relative humidity and hostplant on the behaviour of Stethorus punctillum as apredator of the two-spotted spider mite, Tetranychus urticae. BioControl 2000, 45, 155–164. [Google Scholar] [CrossRef]
- Taghizadeh, R.; Chi, H. Demography of Tetranychus urticae (Acari: Tetranychidae) under different nitrogen regimes with estimations of confidence intervals. Crop Prot. 2022, 155, 105920. [Google Scholar] [CrossRef]
- Sohrabi, F.; Shishehbor, P. Effects of host plant and temperature on growth and reproduction of the strawberry spider mite, Tetranychus turkestani Ugarov and Nikolski (Acari: Tetranychidae). Syst. Appl. Acarol. 2008, 13, 26–32. [Google Scholar] [CrossRef]
- Chauhan, R.K.; Shukla, A. Population dynamics of two spotted spider mite, Tetranychus urticae Koch on French bean (Phaseolus vulgaris L.). Int. J. Plant Prot. 2016, 9, 536–539. [Google Scholar] [CrossRef]
- Osman, M.A.; Zamzam, M.; Dhafar, A.; Alqahtani, A.M. Biological responses of the two-spotted spider mite, Tetranychus urticae to different host plant, Arch. Phytopath. Plant Prot. 2019, 52, 1229–1238. [Google Scholar]
- Saeid, S.; Najafabadi, M. Comparative biology and fertility life tables of Tetranychus urticae Koch (Acari: Tetranychidae) on different common bean cultivars, Int. J. Acarol. 2012, 38, 706–714. [Google Scholar]
- Rao, K.S.; Vishnupriya, R.; Ramaraju, K.; Poornima, K. Effect of abiotic factors on the population dynamics of two spotted spider mite, Tetranychus urticae Koch and its predatory mite, Neoseiulus longispinosus (Evans) in Brinjal Ecosystem. J. Exp. Zool. 2018, 21, 797–800. [Google Scholar]
- Kanika, R.G.; Geroh, M. Impact of Weather Parameters on the Population Dynamics of Tetranychus urticae Koch on Field Grown Cucumber. Ann. Biol. 2014, 30, 140–145. [Google Scholar]
- Santamaria, M.E.; Arnaiz, A.; Rosa-Diaz, I.; González-Melendi, P.; Romero-Hernandez, G.; Ojeda-Martinez, D.A.; Garcia, A.; Contreras, E.; Martinez, M.; Diaz, I. Plant Defenses Against Tetranychus urticae: Mind the Gaps. Plants 2020, 9, 464. [Google Scholar] [CrossRef]
- Louta, M.; Papathanasiou, F.; Damos, P.; Ploskas, N.; Dasygenis, M.; Kyriakidis, T.; Dimokas, N.; Balafas, V.; Chatzisavvas, A.; Karampelia, I.; et al. Intelligent Pesticide and Irrigation Management in Precision Agriculture: The case of VELOS Project. In Proceedings of the HAICTA 2022, Athens, Greece, 22–25 September 2022; pp. 91–99. [Google Scholar]
- Raghavendra, K. Weather based prediction of pests in cotton. In Proceedings of the International Conference on Computational Intelligence and Communication Networks, Bhopal, India, 14–16 November 2014. [Google Scholar]
- Shang, Y.; Zhu, Y. Research on intelligent pest prediction of based on improved artificial neural network. In Proceedings of the Chinese Automation Congress (CAC), Xi’an, China, 30 November–2 December 2018. [Google Scholar]
- Damos, P.; Tuells, J.; Caballero, P. Soft Computing of a Medically Important Arthropod Vector with Autoregressive Recurrent and Focused Time Delay Artificial Neural Networks. Insects 2021, 12, 503. [Google Scholar] [CrossRef]
- Saleem, R.M.; Kazmi, R.; Bajwa, I.S.; Ashraf, A.; Ramzan, S.; Anwar, W. IOT-Based Cotton Whitefly Prediction Using Deep Learning. Sci. Program. 2021, 2021, 8824601. [Google Scholar] [CrossRef]
- Olden, J.D.; Jackson, D.A. Illuminating the “black box”: A randomization approach for understanding variable contributions in artificial neural networks. Ecol. Model. 2002, 154, 135–150. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Degree-Days of Overwintering T. urticae Egg Laying Events | |||||
|---|---|---|---|---|---|
| Year | Start of Oviposition | First Egg Peak | Second Egg Peak | Third Egg Peak | Pooled Egg Peak |
| 2021 | 57 | 137.2 | 292.2 | 461.7 | 123.8 |
| 2022 | 58,4 | 146.5 | 332 | 479.3 | 265.7 |
| Pooled | 57.7 | 141.8 | 321.1 | 470.5 | 194.75 |
| Year | Phenological Event | T. urticae Generation | Mean | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1st | 2nd | 3rd | 4th | 5th | 6th | 7th | 8th | 9th | |||
| 2021 | Half of first generation * | 187 | 505 | 822 | 1044 | 1295 | 1513 | 1745 | 1967 | 2193 | |
| Total generation time * | 278 | 280 | 274 | 258 | 237 | 233 | 235 | 220 | 229 | 249.3 (±7.7) | |
| 2022 | Developmental threshold | 234 | 547 | 792 | 1048 | 1271 | 1519 | 1744 | 1969 | 2195 | |
| Total generation time | 300 | 257 | 250 | 239 | 225 | 252 | 224 | 239 | 256 | 249.2 (±6.7) | |
| Pooled | Developmental threshold | 201 | 526 | 807 | 1046 | 1283 | 1516 | 1744.5 | 1968 | 2194 | |
| Total generation time | 289 | 268.5 | 262 | 248.5 | 231 | 242.5 | 229.5 | 229.5 | 242.5 | 249.8 (±5) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damos, P.; Papathanasiou, F.; Tsikos, E.; Kyriakidis, T.; Louta, M. Predicting the Occurrence and Risk Damage Caused by the Two-Spotted Spider Mite Tetranychus urticae (Koch) in Dry Beans (Phaseolus vulgaris L.) Combining Rate and Heat Summation Models for Digital Decisions Support. Agriculture 2023, 13, 756. https://doi.org/10.3390/agriculture13040756
Damos P, Papathanasiou F, Tsikos E, Kyriakidis T, Louta M. Predicting the Occurrence and Risk Damage Caused by the Two-Spotted Spider Mite Tetranychus urticae (Koch) in Dry Beans (Phaseolus vulgaris L.) Combining Rate and Heat Summation Models for Digital Decisions Support. Agriculture. 2023; 13(4):756. https://doi.org/10.3390/agriculture13040756
Chicago/Turabian StyleDamos, Petros, Fokion Papathanasiou, Evaggelos Tsikos, Thomas Kyriakidis, and Malamati Louta. 2023. "Predicting the Occurrence and Risk Damage Caused by the Two-Spotted Spider Mite Tetranychus urticae (Koch) in Dry Beans (Phaseolus vulgaris L.) Combining Rate and Heat Summation Models for Digital Decisions Support" Agriculture 13, no. 4: 756. https://doi.org/10.3390/agriculture13040756
APA StyleDamos, P., Papathanasiou, F., Tsikos, E., Kyriakidis, T., & Louta, M. (2023). Predicting the Occurrence and Risk Damage Caused by the Two-Spotted Spider Mite Tetranychus urticae (Koch) in Dry Beans (Phaseolus vulgaris L.) Combining Rate and Heat Summation Models for Digital Decisions Support. Agriculture, 13(4), 756. https://doi.org/10.3390/agriculture13040756