

Brucella melitensis Vaccines: A Systematic Review


Abstract
:1. Introduction
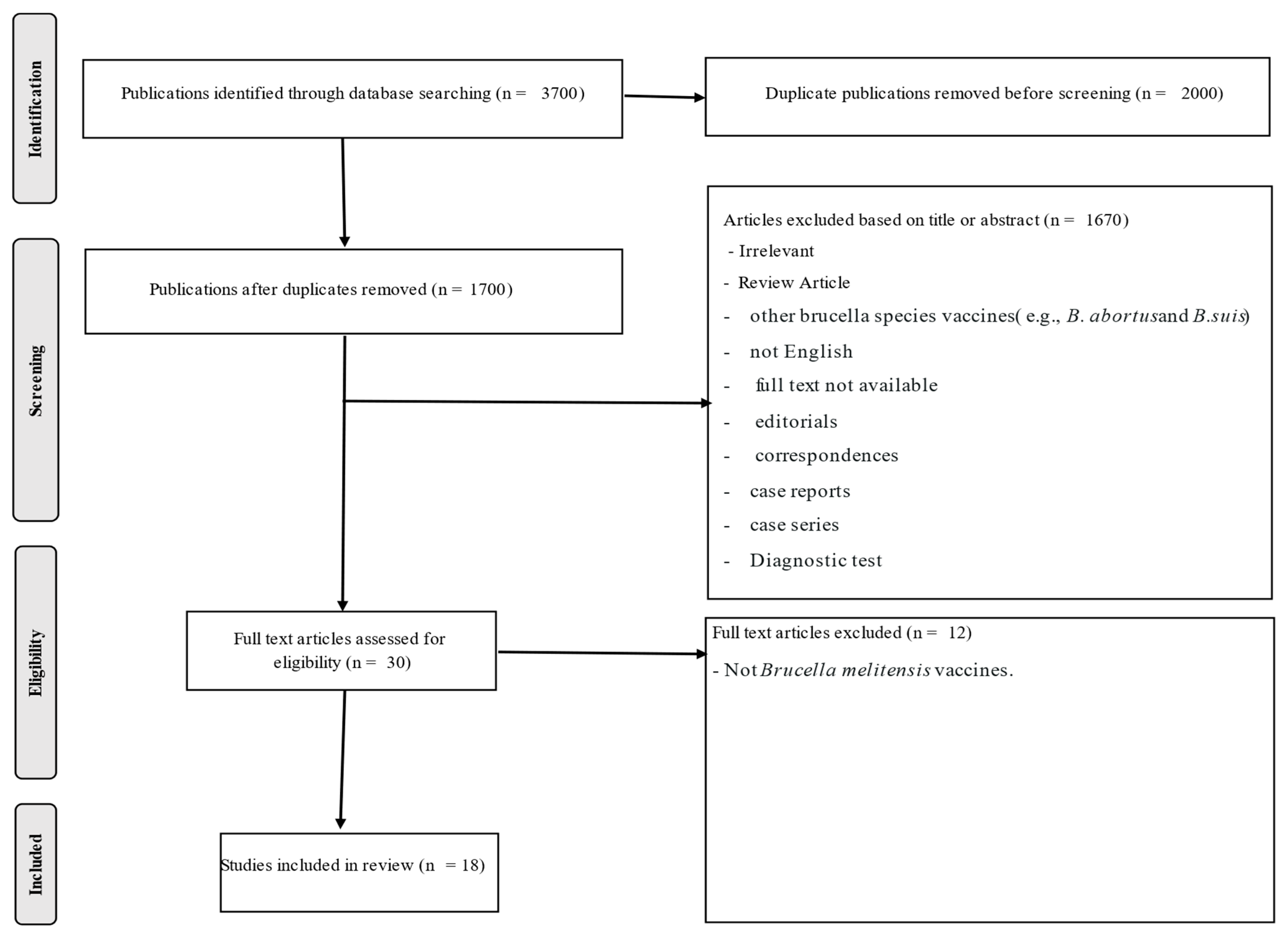
2. Materials and Methods
2.1. Search Strategy
2.2. Study Selection
2.3. Screening and Data Extraction
2.4. Quality Assessment
3. Results
3.1. Study Characteristics
3.2. Brucella melitensis Vaccines in Small Ruminants
4. Discussion
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Welburn, S.C.; Beange, I.; Ducrotoy, M.J.; Okello, A.L. The Neglected Zoonoses—The Case for Integrated Control and Advocacy. Clin. Microbiol. Infect. 2015, 21, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Godfroid, J.; DeBolle, X.; Roop, R.M.; O’Callaghan, D.; Tsolis, R.M.; Baldwin, C.; Santos, R.L.; McGiven, J.; Olsen, S.; Nymo, I.H.; et al. The quest for a true One Health perspective of brucellosis. Rev. Sci. Tech. 2014, 33, 521–538. [Google Scholar] [CrossRef] [PubMed]
- Muendo, E.N.; Mbatha, P.M.; Macharia, J.; Abdoel, T.H.; Janszen, P.V.; Pastoor, R.; Smits, H.L. Infection of cattle in Kenya with Brucella abortus biovar 3 and Brucella melitensis biovar 1 genotypes. Trop. Anim. Health Prod. 2012, 44, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Gwida, M.; El-Gohary, A.; Melzer, F.; Khan, I.; Rösler, U.; Neubauer, H. Brucellosis in camels. Res. Vet. Sci. 2012, 92, 351–355. [Google Scholar] [CrossRef]
- Mick, V.; Carrou, G.L.; Corde, Y.; Game, Y.; Jay, M.; Garin-Bastuji, B. Brucella melitensis in France: Persistence in wildlife and probable spillover from alpine ibex to domestic animals. PLoS ONE 2014, 9, e94168. [Google Scholar] [CrossRef]
- Nicoletti, P. Vaccination. In Animal Brucellosis; Nielsen, K., Duncan, J.R., Eds.; CRC Press: Boca Raton, FL, USA, 1990; pp. 284–299. [Google Scholar]
- Roux, J. Brucella vaccines in humans. In Brucellosis; Madkour, M.M., Ed.; Butterworths: London, UK, 1989; pp. 244–249. [Google Scholar]
- Plommet, M.; Renoux, G.; Philppon, A.; Lorentz, C.; Gestin, J. Experimental brucellosis, Comparison of vaccine efficacy B19 and H38 vaccine. Ann. Vet. Res. 1970, 1, 189–201. [Google Scholar]
- Elberg, S.S.; Faunce, K., Jr. Immunization against Brucella infection. VI. Immunity conferred on goats by a nondependent mutant from a streptomycin-dependent mutant strain of Brucella melitensis. J. Bacteriol. 1957, 73, 211–217. [Google Scholar] [CrossRef]
- Banai, M. Control of small ruminant brucellosis by use of Brucella melitensis Rev.1 vaccine: Laboratory aspects and field observations. Vet. Microbiol. 2002, 90, 497–519. [Google Scholar] [CrossRef]
- Alton, G.G. Vaccination of goats with reduced doses of Rev.1 Brucella melitensis vaccine. Res. Vet. Sci. 1970, 2, 54–59. [Google Scholar] [CrossRef]
- Blasco, J.M.; Molina-Flores, B. Control and eradication of Brucella melitensis infection in sheep and goats. Vet. Clin. N. Am. Food Anim. Pract. 2011, 27, 95–104. [Google Scholar] [CrossRef]
- Blasco, J.M. A review of the use of B. melitensis Rev.1 vaccine in adult sheep and goats. Prev. Vet. Med. 1997, 31, 275–283. [Google Scholar] [CrossRef]
- Vershilova, P. The use of live vaccine for vaccination of human beings against brucellosis in the USSR. Bull. World Health Organ. 1961, 24, 85. [Google Scholar]
- Pappas, G. The changing Brucella ecology: Novel reservoirs, new threats. Int. J. Antimicrob. Agents 2010, 36, S8–S11. [Google Scholar] [CrossRef] [PubMed]
- FAO/WHO/OIE. Round Table on the Use of Rev.1 Vaccine in Small Ruminants and Cattle; Garin-Bastuji, B., Benkirane, A., Eds.; CNEVA: Paris, France, 1995. [Google Scholar]
- Pasquevich, K.A.; Estein, S.M.; Samartino, C.G.; Zwerdling, A.; Coria, L.M.; Barrionuevo, P.; Fossati, C.; Giambartolomei, G.; Cassataro, J. Immunization with recombinant Brucella species outer membrane protein Omp16 or Omp19 in adjuvant induces specific CD4+ and CD8+ T cells as well as systemic and oral protection against Brucella abortus infection. Infect. Immun. 2009, 77, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, 71. [Google Scholar] [CrossRef] [PubMed]
- Institute, J. The Joanna Briggs Institute Critical Appraisal Tools for Use in JBI Systematic Reviews Checklist for Analytical Cross-Sectional Studies; The Joanna Briggs Institute: Adelaide, Australia, 2017. [Google Scholar]
- Zhang, J.; Guo, F.; Chen, C.; Li, Z.; Zhang, H.; Wang, Y.; Zhang, K.; Du, G.; Li, Y.; Wang, J.; et al. Brucella melitensis 16 M Δhfq attenuation confers protection against wild-type challenge in BALB/c mice. Microbiol. Immunol. 2013, 57, 502–510. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, J.; Zhang, K.; Fu, Q.; Wang, Z.; Li, T.; Zhang, H.; Guo, F.; Chen, C. Brucella melitensis 16MΔTcfSR as a potential live vaccine allows for the differentiation between natural and vaccinated infection. Exp. Ther. Med. 2015, 10, 1182–1188. [Google Scholar] [CrossRef]
- Zhang, J.; Yin, S.; Yi, D.; Zhang, H.; Li, Z.; Guo, F.; Chen, C.; Fang, W.; Wang, J. The Brucella melitensis M5-90ΔmanB live vaccine candidate is safer than M5-90 and confers protection against wild-type challenge in BALB/c mice. Microb. Pathog. 2017, 112, 148–155. [Google Scholar] [CrossRef]
- Verdiguel-Fernández, L.; Oropeza-Navarro, R.; Ortiz, A.; Robles-Pesina, M.; Ramírez-Lezama, J.; Castañeda-Ramírez, A.; Verdugo-Rodríguez, A. Brucella melitensis omp31 mutant is attenuated and confers protection against virulent Brucella melitensis challenge in BALB/c mice. J. Microbiol. Biotechnol. 2020, 30, 497–504. [Google Scholar] [CrossRef]
- Li, Z.; Wang, S.; Zhang, H.; Xi, L.; Zhang, J.; Zhang, X.; Zhou, Q.; Yi, J.; Li, M.; Zhang, W.; et al. Development and evaluation of in murine model, of an improved live-vaccine candidate against brucellosis from to Brucella melitensis vjbR deletion mutant. Microb. Pathog. 2018, 124, 250–257. [Google Scholar] [CrossRef]
- Arenas-Gamboa, A.; Rice-Ficht, A.; Kahl-McDonagh, M.; Ficht, T. Protective efficacy and safety of Brucella melitensis 16MΔmucR against intraperitoneal and aerosol challenge in BALB/c mice. Infect. Immun. 2011, 79, 3653–3658. [Google Scholar] [CrossRef]
- Clapp, B.; Skyberg, J.A.; Yang, X.; Thornburg, T.; Walters, N.; Pascual, D.W. Protective live oral brucellosis vaccines stimulate Th1 and Th17 cell responses. Infect. Immun. 2011, 79, 4165–4174. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, T.; Zhang, J.; Li, Z.; Zhang, Y.; Wang, Z.; Feng, H.; Wang, Y.; Chen, C.; Zhang, H. The Brucella melitensis M5-90 phosphoglucomutase (PGM) mutant is attenuated and confers protection against wild-type challenge in BALB/c mice. World J. Microbiol. Biotechnol. 2016, 32, 58. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-Q.; Shi, J.-X.; Fu, W.-D.; Zhang, Y.; Zhang, J.; Wang, Z.; Li, T.; Chen, C.; Guo, F.; Zhang, H. A Brucella melitensis M5-90 wboA deletion strain is attenuated and enhances vaccine efficacy. Mol. Immunol. 2015, 66, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Metwally, S.; Viljoen, G.; Idrissi, A. Veterinary Vaccines: Principles and Applications; John Wiley & Sons: Hoboken, NJ, USA, 2021; p. 298. [Google Scholar]
- Shojaei, M.; Tahmoorespur, M.; Soltani, M.; Sekhavati, M.H. Immunogenicity evaluation of plasmids encoding Brucella melitensis Omp25 and Omp31 antigens in BALB/c mice. Iran. J. Basic Med. Sci. 2018, 21, 957. [Google Scholar] [CrossRef]
- Karevan, G.; Ahmadi, K.; Taheri, R.A.; Fasihi-Ramandi, M. Immunogenicity of glycine nanoparticles containing a chimeric antigen as Brucella vaccine candidate. Clin. Exp. Vaccine Res. 2021, 10, 35. [Google Scholar] [CrossRef] [PubMed]
- Abkar, M.; Fasihi-Ramandi, M.; Kooshki, H.; Lotfi, A.S. Intraperitoneal immunization with Urease loaded N-trimethyl Chitosan nanoparticles elicits high protection against Brucella melitensis and Brucella abortus infections. Immunol. Lett. 2018, 199, 53–60. [Google Scholar] [CrossRef]
- Sadeghi, Z.; Fasihi-Ramandi, M.; Azizi, M.; Bouzari, S. Mannosylated chitosan nanoparticles loaded with FliC antigen as a novel vaccine candidate against Brucella melitensis and Brucella abortus infection. J. Biotechnol. 2020, 310, 89–96. [Google Scholar] [CrossRef]
- Abkar, M.; Fasihi-Ramandi, M.; Kooshki, H.; Lotfi, A.S. Oral immunization of mice with Omp31-loaded N-trimethyl chitosan nanoparticles induces high protection against Brucella melitensis infection. Int. J. Nanomed. 2017, 12, 8769. [Google Scholar] [CrossRef]
- Maleki, M.; Salouti, M.; Shafiee Ardestani, M.; Talebzadeh, A. Preparation of a nanovaccine against Brucella melitensis M16 based on PLGA nanoparticles and oligopolysaccharide antigen. Artif. Cells Nanomed. Biotechnol. 2019, 47, 4248–4256. [Google Scholar] [CrossRef]
- Golshani, M.; Amani, M.; Amirzadeh, F.; Nazeri, E.; Siadat, S.D.; Nejati-Moheimani, M.; Arsang, A.; Bouzari, S. Evaluation of Poly (I: C) and combination of CpG ODN plus Montanide ISA adjuvants to enhance the efficacy of outer membrane vesicles as an acellular vaccine against Brucella melitensis infection in mice. Int. Immunopharmacol. 2020, 84, 106573. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Wang, Q.; Wang, Y.; Xu, Y.; Peng, D.; Huang, H.; Hu, L.; Wei, K.; Zhu, R. Comparison of immune effects between Brucella recombinant Omp10-Omp28-L7/L1 proteins expressed in eukaryotic and prokaryotic systems. Front. Vet. Sci. 2020, 7, 576. [Google Scholar] [CrossRef] [PubMed]
- Ficht, T.A.; Kahl-McDonagh, M.M.; Arenas-Gamboa, A.M.; Rice-Ficht, A.C. Brucellosis: The case for live, attenuated vaccines. Vaccine 2009, 27, D40–D43. [Google Scholar] [CrossRef]
- Fu, S.; Xu, J.; Li, X.; Xie, Y.; Qiu, Y.; Du, X.; Yu, S.; Bai, Y.; Chen, Y.; Wang, T.; et al. Immunization of mice with recombinant protein CobB or AsnC confers protection against Brucella abortus infection. PLoS ONE 2012, 7, e29552. [Google Scholar] [CrossRef] [PubMed]
- Laine, C.G.; Johnson, V.E.; Scott, H.; Arenas-Gamboa, A.M. Global Estimate of Human Brucellosis Incidence. Emerg. Infect. Dis. 2023, 29, 1789–1797. [Google Scholar] [CrossRef]
- Franco, M.; Mulder, M.; Gilman, R.; Smits, H. Human brucellosis. Lancet Infect. Dis. 2007, 7, 775–786. [Google Scholar] [CrossRef]
- Jacob, J.M.; Curtiss, R. Characterization of Brucella abortus S19 as a challenge strain for use in a mouse model of brucellosis. Microbes Infect. 2021, 23, 104809. [Google Scholar] [CrossRef]
- Mantur, B.G.; Amarnath, S.K.; Shinde, R.S. Review of clinical and labora-tory features of human Brucellosis. Indian J. Med. Microbiol. 2007, 25, 188–202. [Google Scholar] [CrossRef]
- Shford, D.A.; di Pietra, J.; Lingappa, J.; Woods, C.; Noll, H.; Neville, B.; Weyant, R.; Bragg, S.L.; Spiegel, R.A.; Tappero, J.; et al. Adverse events in humans associated with accidentalexposure to the livestock brucellosis vaccine RB51. Vaccine 2004, 22, 3435–3439. [Google Scholar] [CrossRef]
- Schurig, G.G.; Sriranganathan, N.; Corbel, M.J. Brucellosisvaccines: Past, present and future. Vet. Microbiol. 2002, 90, 479–496. [Google Scholar] [CrossRef]
- Alton, G.G. Rev.1 and H38 Brucella melitensis vaccines. In Brucella Melitensis; Verger, J.M., Plommet, M., Eds.; Martinus Nijhoff: Dordrecht, The Netherlands, 1985; pp. 215–227. [Google Scholar]
- Alton, G.G.; Jones, L.M.; Pietz, D.E. Laboratory techniques in brucellosis. In World Health Organisation Technical Monograph; Series No. 55; WHO: Geneva, Switzerland, 1975. [Google Scholar]
- Blasco, J.M.; Marin, C.M.; Barberan, M.; Moriyon, I.; Diaz, R. Immunization with Brucella melitensis Rev.1 against Brucella ovis infection of rams. Vet. Microbiol. 1987, 14, 381–392. [Google Scholar] [CrossRef]
- Bardenstein, S.; Mandelboim, M.; Ficht, T.A.; Baum, M.; Banai, M. Identification of the Brucella melitensis vaccine strain Rev.1 in animals and humans in Israel by PCR analysis of the PstI site polymorphism of its omp2 gene. J. Clin. Microbiol. 2002, 40, 1475–1480. [Google Scholar] [CrossRef]
- Lantier, F.; Fensterbank, R. Kinetics of Rev.1 infection in sheep. In Brucella melitensis; Plommet, M., Verger, J.M., Eds.; Martinus Niijhoff: Dordrecht, The Netherlands, 1985; pp. 247–251. [Google Scholar]
- Van Drimmelen, C.; Horwell, F.D. Preliminary findings with the use of Brucella melitensis Rev.1 as a vaccine against brucellosis in cattle. Bull. Off. Int. Epiz. 1964, 62, 987. [Google Scholar]
- Horwell, F.D.; van Drimmelen, G.G. Brucella melitensis strain Rev.1 as a vaccine in cattle. S. Afr. Vet. Med. Assoc. 1971, 42, 233–235. [Google Scholar]
- Garcia-Carrillo, C. Comparison of Brucella melitensis Rev.1 and B. abortus strain 19 as a vaccine against brucellosis in cattle. Zentralbl. Veterinaermed. 1980, 27, 131–138. [Google Scholar] [CrossRef]
- Erasmus, J.A.; Bergh, E.C. Ovine brucellosis: Repeated vaccination with Rev.1 vaccine and the prevalence of the disease in the Winburg district. J. S. Afr. Vet. Assoc. 1985, 56, 205–208. [Google Scholar]
- Xie, X. Orally administrable brucellosis vaccine: Brucella suis strain 2 vaccine. Vaccine 1986, 4, 212–216. [Google Scholar]
- Lu, S.-L.; Zhang, J.-L. Brucellosis in China. In Brucellosis: Clinical and Laboratory Aspects; Young, E.J., Corbel, M.J., Eds.; CRC Press: Boca Raton, FL, USA, 1989; Chapter 13; pp. 173–180. [Google Scholar]
- Verger, J.M.; Grayon, M.; Zundel, E.; Lechopier, P.; Olivier-Bernardin, V. Comparison of the efficacy of Brucella suis strain 2 and Brucella melitensis Rev.1 live vaccines against a Brucella melitensis experimental infection in pregnant ewes. Vaccine 1995, 13, 7625115. [Google Scholar] [CrossRef] [PubMed]
- Gheibi, A.; Khanahmad, H.; Kashfi, K.; Sarmadi, M.; Khorramizadeh, M.R. Development of new generation of vaccines for Brucella abortus. Heliyon 2018, 4, e01079. [Google Scholar] [CrossRef] [PubMed]
- Hou, H.; Liu, X.; Peng, Q. The advances in brucellosis vaccines. Vaccine 2019, 37, 3981–3988. [Google Scholar] [CrossRef]
- Gomez, L.; Llanos, J.; Escalona, E.; Saez, D.; Alvarez, F.; Molina, R.; Flores, M.; Onate, A. Multivalent Fusion DNA Vaccine against Brucella abortus. BioMed Res. Int. 2017, 2017, 6535479. [Google Scholar] [CrossRef]
- Moreno, E.; Barquero-Calvo, E. The role of neutrophils in brucellosis. Microbiol. Mol. Biol. Rev. 2020, 84, e00048-e20. [Google Scholar] [CrossRef] [PubMed]
- Singha, H.; Mallick, A.I.; Jana, C.; Fatima, N.; Owais, M.; Chaudhuri, P. Co-immunization with interlukin-18 enhances the protective efficacy of liposomes encapsulated recombinant Cu–Zn superoxide dismutase protein against Brucella abortus. Vaccine 2011, 29, 4720–4727. [Google Scholar] [CrossRef] [PubMed]
- Olsen, S.C. Recent developments in livestock and wildlife brucellosis vaccination. Rev. Sci. Tech. 2013, 32, 207–217. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Name of Vaccine | Type of Vaccine | Animal Models & Ages | Advantages | Disadvantages | Publication Reference Year |
|---|---|---|---|---|---|
| B. melitensis 16MΔhfq | Recombinant Vaccine | Six-week-old BALB/c female mice |
|
| 2013 [20] |
| B. melitensis 16MΔTcfSR | Recombinant Vaccine | Six-week-old BALB/c female mice |
|
| 2015 [21] |
| B. melitensis M5-90ΔmanB | Recombinant Vaccine | Seven-week-old BALB/c female mice |
|
| 2017 [22] |
| B. melitensis LVM31 mutant strain | Recombinant Vaccine | Eight-week-old BALB/c female and male mice |
|
| 2020 [23] |
| B. melitensis M5-90ΔvjbR | Recombinant Vaccine | Six-week-old BALB/c female mice |
|
| 2018 [24] |
| B. melitensis 16MΔmucR | Recombinant Vaccine | Six–eight-week-old BALB/c female mice |
|
| 2011 [25] |
| B. melitensis ΔznuA | Recombinant Vaccine | BALB/c female mice |
|
| 2011 [26] |
| B. melitensis M5-90Δpgm | Recombinant Vaccine | Six-week-old BALB/c female mice |
|
| 2016 [27] |
| B. melitensis M5-90ΔwboA | Recombinant Vaccine | Six-week-old BALB/c female mice |
|
| 2015 [28] |
| B. melitensis Rev 1 | Live B. melitensis strains | Goats |
|
| 1957 [9] |
| B. melitensis outer membrane proteins (Omp25 and Omp31) | DNA Vaccine | Seven–eight-week-old BALB/c female mice |
|
| 2018 [30] |
| B. melitensis 16M | Nanoparticle Vaccine | Six–eight-week-old BALB/c female mice |
|
| 2021 [31] |
| B. melitensis and B. abortus combined | Nanoparticle Vaccine | Four–six-week-old BALB/c female mice |
|
| 2018 [32] |
| FliC antigen-MCN | Nanoparticle Vaccine | Six–eight-week-old BALB/c female mice |
|
| 2020 [33] |
| B. melitensis Omp31 | Nanoparticle Vaccine | Four–six-week-old BALB/c female mice |
|
| 2017 [34] |
| B. melitensis 16M combined with oligopolysaccharide | Nanoparticle Vaccine | Six–eight-week-old BALB/c mice |
|
| 2019 [35] |
| Outer membrane vesicles | Subunit Vaccine | Six–eight-week-old BALB/c female mice |
|
| 2020 [36] |
| Outer membrane proteins | Subunit Vaccine | Five–six-week-old BALB/c female mice |
|
| 2020 [37] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naseer, A.; Mo, S.; Olsen, S.C.; McCluskey, B. Brucella melitensis Vaccines: A Systematic Review. Agriculture 2023, 13, 2137. https://doi.org/10.3390/agriculture13112137
Naseer A, Mo S, Olsen SC, McCluskey B. Brucella melitensis Vaccines: A Systematic Review. Agriculture. 2023; 13(11):2137. https://doi.org/10.3390/agriculture13112137
Chicago/Turabian StyleNaseer, Alnakhli, Salman Mo, Steven C. Olsen, and Brian McCluskey. 2023. "Brucella melitensis Vaccines: A Systematic Review" Agriculture 13, no. 11: 2137. https://doi.org/10.3390/agriculture13112137
APA StyleNaseer, A., Mo, S., Olsen, S. C., & McCluskey, B. (2023). Brucella melitensis Vaccines: A Systematic Review. Agriculture, 13(11), 2137. https://doi.org/10.3390/agriculture13112137