
The Use of Agro-Industrial Waste Rich in Omega-3 PUFA during the Weaning Stress Improves the Gut Health of Weaned Piglets



 ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Chemical Composition of GFS and Diets
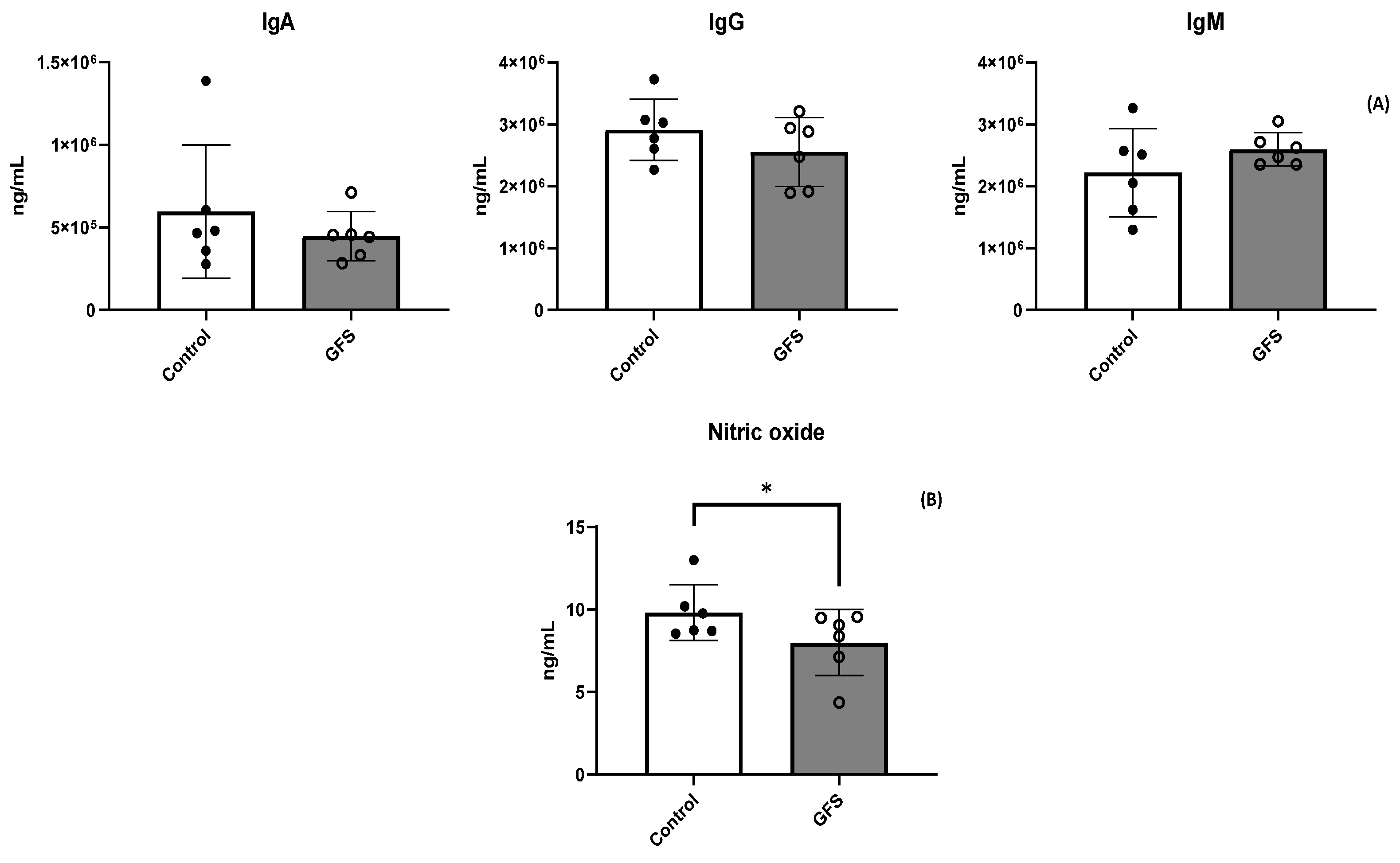
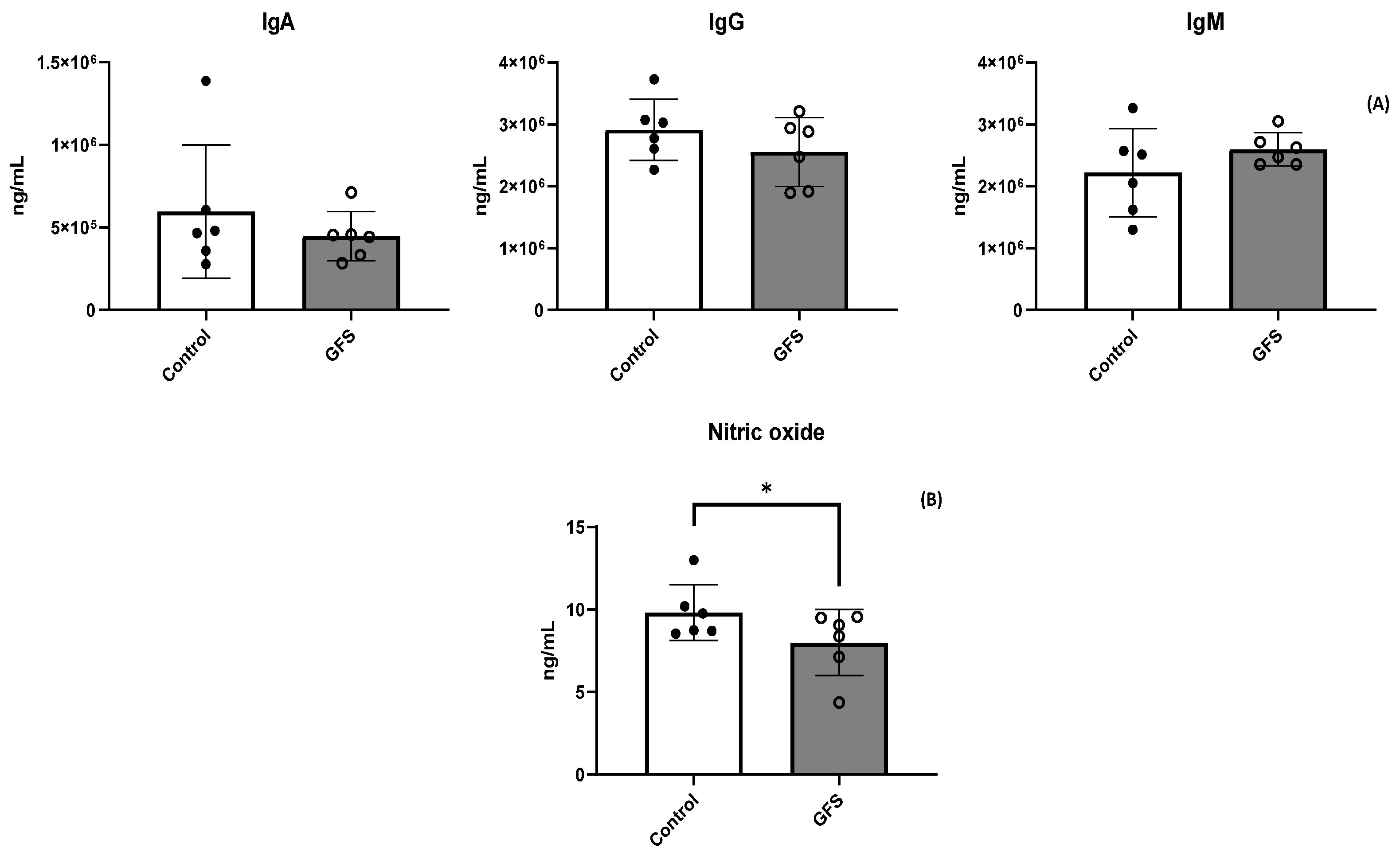
3.2. GFS Effect on Animal Performance and Health Status
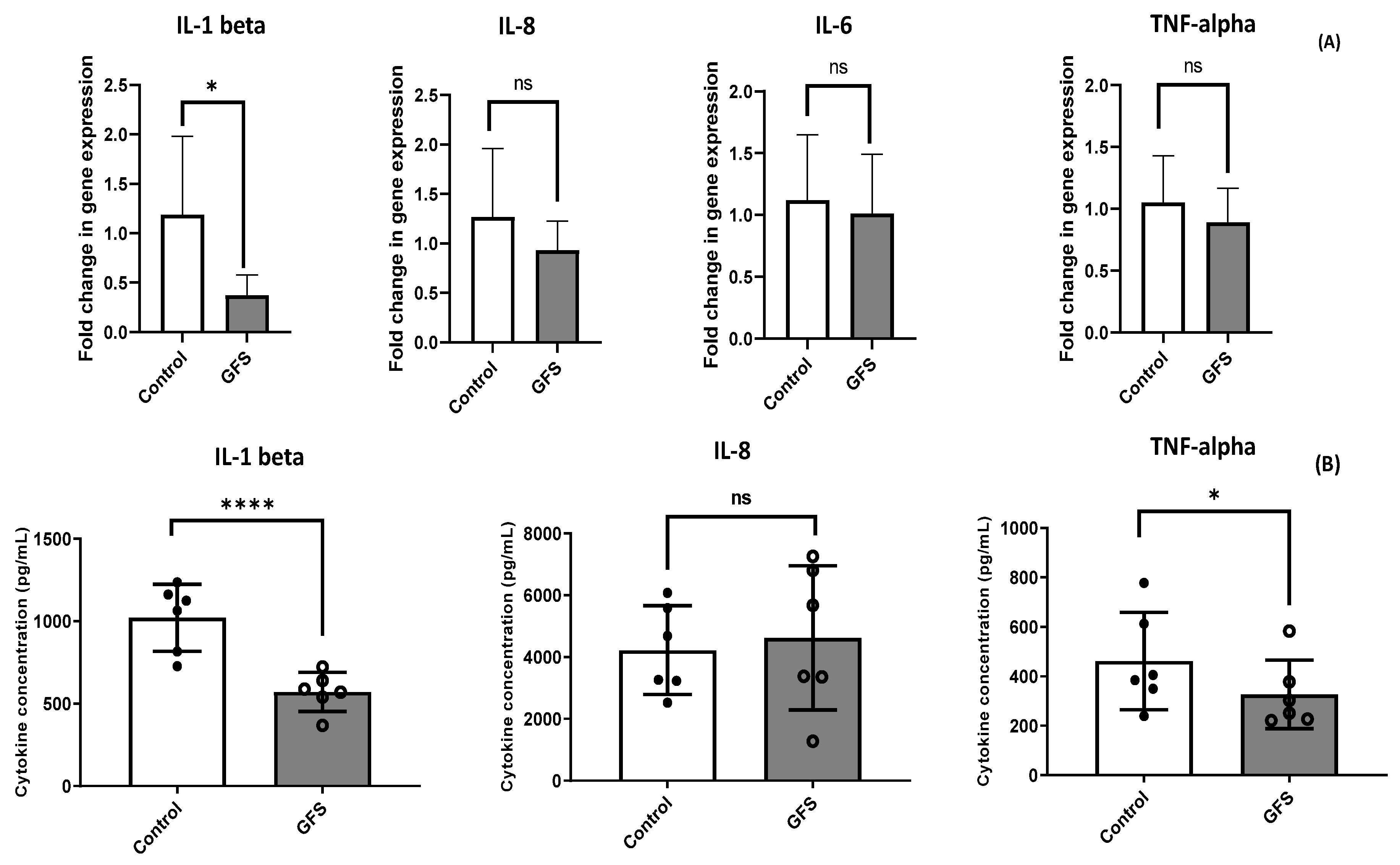
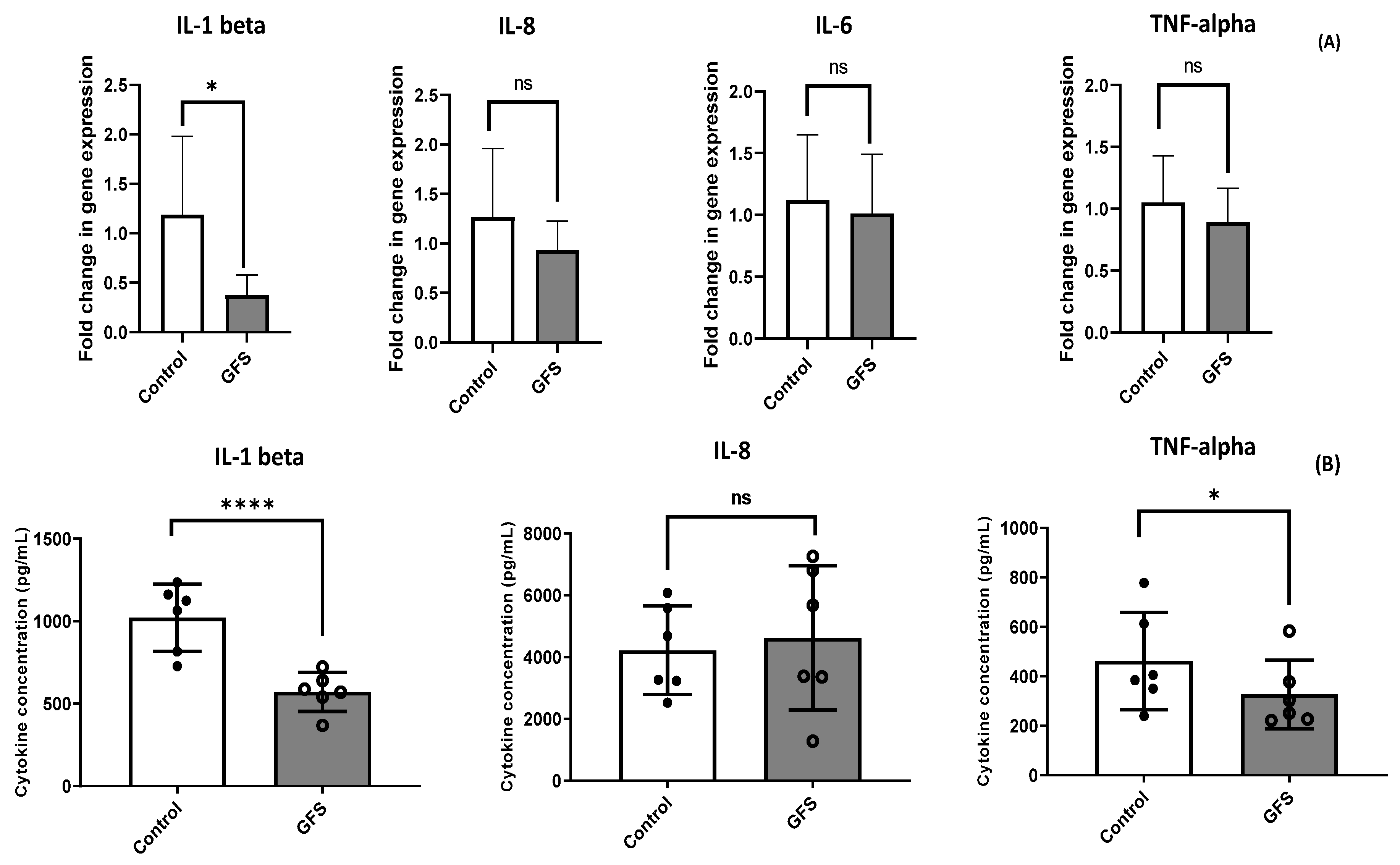
3.3. GFS Effect on Proinflammatory Cytokines
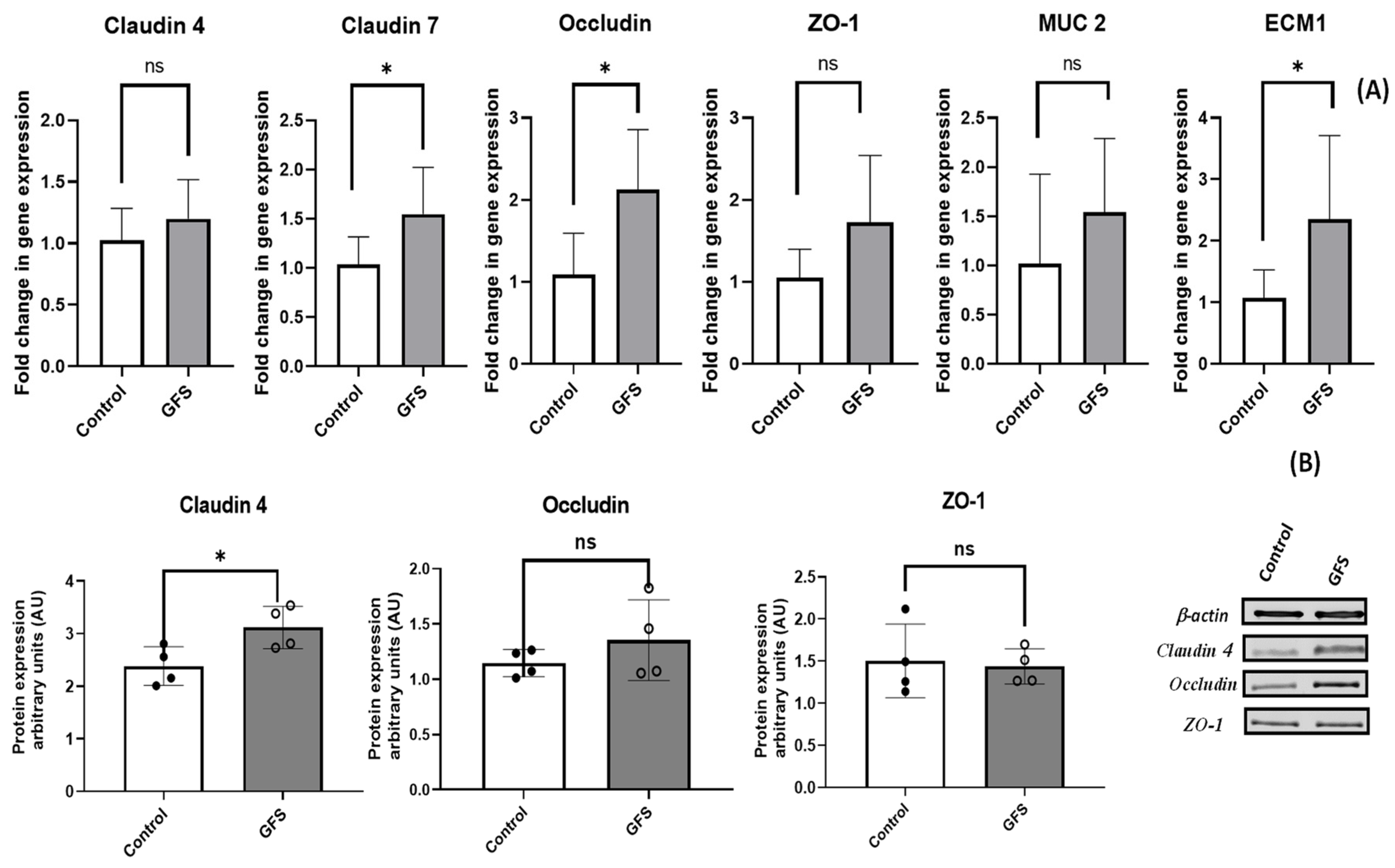
3.4. GFS Effect on Junction Proteins, Mucins and Proteins of Extracellular Matrix
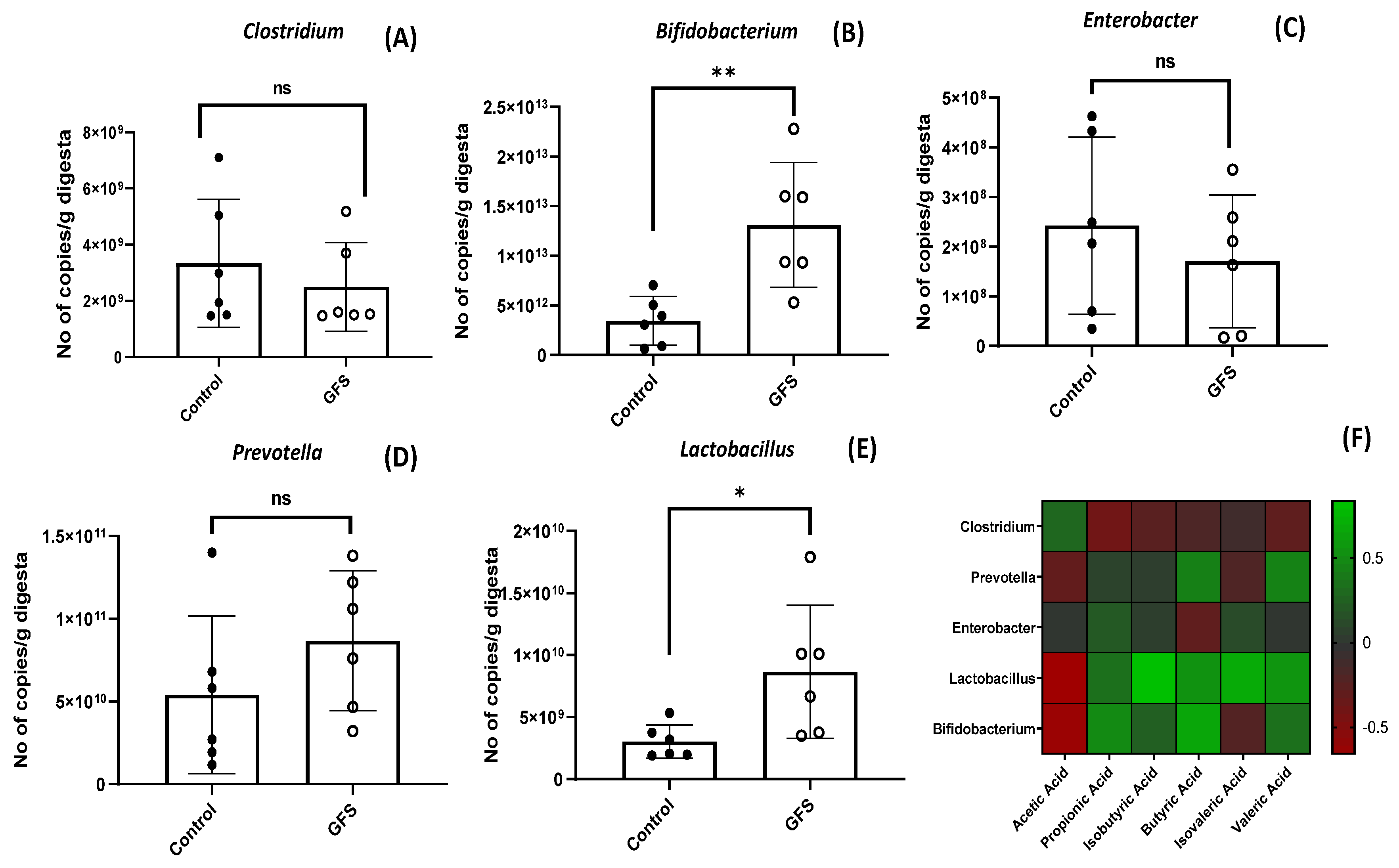
3.5. GFS Effect on Selected Microbiota Populations and on Short-Chain Fatty Acids
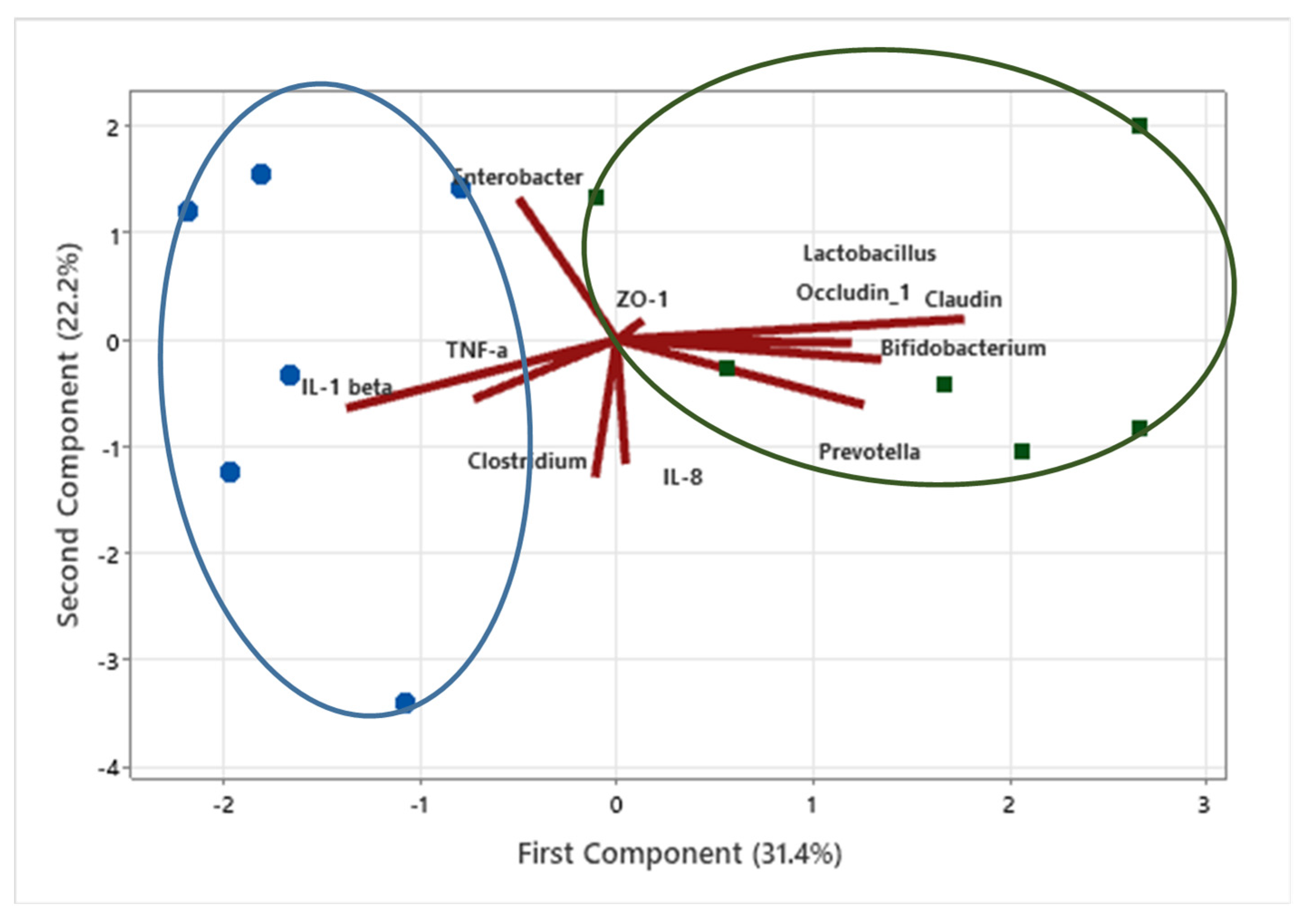
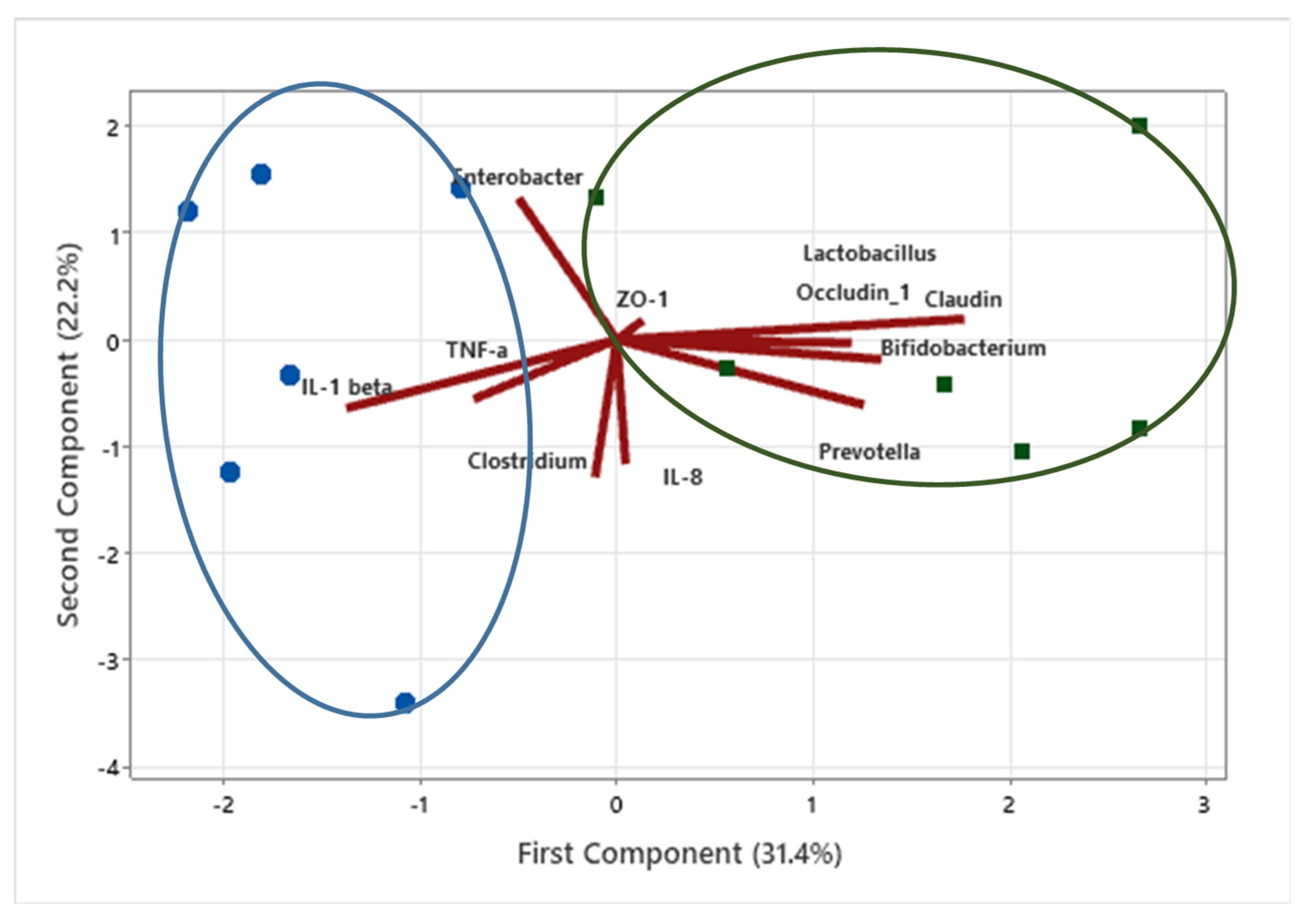
3.6. Data Clustering Using Principal Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Leeb, C.; Hegelund, L.; Edwards, S.; Mejer, H.; Roepstorff, A.; Rousing, T.; Sundrum, A.; Bonde, M. Animal health, welfare and production problems in organic weaner pigs. Org. Agric. 2014, 4, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Jayaraman, B.; Nyachoti, C.M. Husbandry practices and gut health outcomes in weaned piglets: A review. Anim. Nutr. 2017, 3, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Duarte, M.E.; Sevarolli Loftus, A.; Kim, S.W. Intestinal health of pigs upon weaning: Challenges and nutritional intervention. Front. Vet. 2021, 8, 628258. [Google Scholar] [CrossRef]
- Moeser, A.J.; Pohl, C.S.; Rajput, M. Weaning stress and gastrointestinal barrier development: Implications for lifelong gut health in pigs. Anim. Nutr. 2017, 3, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Blavi, L.; Solà-Oriol, D.; Llonch, P.; López-Vergé, S.; Martín-Orúe, S.M.; Pérez, J.F. Management and feeding strategies in early life to increase piglet performance and welfare around weaning: A review. Animals 2021, 11, 302. [Google Scholar] [CrossRef] [PubMed]
- Wensley, M.R.; Tokach, M.D.; Woodworth, J.C.; Goodband, R.D.; Gebhardt, J.T.; DeRouchey, J.M.; McKilligan, D. Maintaining continuity of nutrient intake after weaning. Ii. Review of post-weaning strategies. Transl. Anim. Sci. 2021, 5, txab022. [Google Scholar] [CrossRef] [PubMed]
- Lallès, J.-P.; Bosi, P.; Smidt, H.; Stokes, C.R. Nutritional management of gut health in pigs around weaning. Proc. Nutr. Soc. 2007, 66, 260–268. [Google Scholar] [CrossRef]
- Cheng, V.J.; Bekhit Ael, D.; Sedcole, R.; Hamid, N. The impact of grape skin bioactive functionality information on the acceptability of tea infusions made from wine by-products. J. Food Sci. 2010, 75, S167–S172. [Google Scholar] [CrossRef]
- Rockenbach, I.I.; Gonzaga, L.V.; Rizelio, V.M.; Gonçalves, A.E.d.S.S.; Genovese, M.I.; Fett, R. Phenolic compounds and antioxidant activity of seed and skin extracts of red grape (vitis vinifera and vitis labrusca) pomace from brazilian winemaking. Food Res. Int. 2011, 44, 897–901. [Google Scholar] [CrossRef]
- Khdair, A.; Abu-Rumman, G. Sustainable environmental management and valorization options for olive mill byproducts in the middle east and north africa (mena) region. Processes 2020, 8, 671. [Google Scholar] [CrossRef]
- Romero, C.; Medina, E.; Mateo, M.; Brenes, M. New by-products rich in bioactive substances from the olive oil mill processing. J. Sci. Food Agric. 2017, 98, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Grosu, I.; Pistol, G.; Marin, D.; Cismileanu, A.; Palade, M.; Taranu, I. Effects of dietary grape seed meal bioactive compounds on the colonic microbiota of weaned piglets with dextran sodium sulfate-induced colitis used as an inflammatory model. Front. Vet. Sci. 2020, 7, 31. [Google Scholar] [CrossRef] [Green Version]
- Viveros, A.; Chamorro, S.; Pizarro, M.; Arija, I.; Centeno, C.; Brenes, A. Effects of dietary polyphenol-rich grape products on intestinal microflora and gut morphology in broiler chicks. Poult. Sci. 2011, 90, 566–578. [Google Scholar] [CrossRef]
- Abedi, E.; Sahari, M.A. Long-chain polyunsaturated fatty acid sources and evaluation of their nutritional and functional properties. Food Sci. Nutr. 2014, 2, 443–463. [Google Scholar] [CrossRef]
- Shaikh, S.R.; Edidin, M. Polyunsaturated fatty acids and membrane organization: Elucidating mechanisms to balance immunotherapy and susceptibility to infection. Chem. Phys. Lipids 2008, 153, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Marion-Letellier, R.; Savoye, G.; Beck, P.L.; Panaccione, R.; Ghosh, S. Polyunsaturated fatty acids in inflammatory bowel diseases: A reappraisal of effects and therapeutic approaches. Inflamm. Bowel Dis. 2013, 19, 650–661. [Google Scholar] [CrossRef]
- Taranu, I.; Habeanu, M.; Gras, M.A.; Pistol, G.C.; Lefter, N.; Palade, M.; Ropota, M.; Sanda Chedea, V.; Marin, D.E. Assessment of the effect of grape seed cake inclusion in the diet of healthy fattening-finishing pigs. J. Anim. Physiol. Anim. Nutr. 2018, 102, e30–e42. [Google Scholar] [CrossRef] [Green Version]
- Taranu, I.; Marin, D.E.; Palade, M.; Pistol, G.C.; Chedea, V.S.; Gras, M.A.; Rotar, C. Assessment of the efficacy of a grape seed waste in counteracting the changes induced by aflatoxin b1 contaminated diet on performance, plasma, liver and intestinal tissues of pigs after weaning. Toxicon Off. J. Int. Soc. Toxinology 2019, 162, 24–31. [Google Scholar] [CrossRef]
- Marin, D.E.; Pistol, G.C.; Gras, M.; Palade, M.; Taranu, I. A comparison between the effects of ochratoxin a and aristolochic acid on the inflammation and oxidative stress in the liver and kidney of weanling piglets. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2018, 391, 1147–1156. [Google Scholar] [CrossRef]
- Marin, D.E.; Bulgaru, C.V.; Anghel, C.A.; Pistol, G.C.; Dore, M.I.; Palade, M.L.; Taranu, I. Grape seed waste counteracts aflatoxin b1 toxicity in piglet mesenteric lymph nodes. Toxins 2020, 12, 800. [Google Scholar] [CrossRef]
- Pistol, G.C.; Bulgaru, C.V.; Marin, D.E.; Oancea, A.G.; Taranu, I. Dietary grape seed meal bioactive compounds alleviate epithelial dysfunctions and attenuates inflammation in colon of dss-treated piglets. Foods 2021, 10, 530. [Google Scholar] [CrossRef]
- Pistol, G.C.; Marin, D.E.; Rotar, M.C.; Ropota, M.; Taranu, I. Bioactive compounds from dietary whole grape seed meal improved colonic inflammation via inhibition of mapks and nf-kb signaling in pigs with dss induced colitis. J. Funct. Foods 2020, 66, 103708. [Google Scholar] [CrossRef]
- Grosu, I.; Cismileanu, A.E.; Marin, D.E.; Taranu, I. The effects of a low dose ota exposure on weanling piglet gut microbiota. Arch. Zootech. 2021, 24, 31–43. [Google Scholar] [CrossRef]
- Parolini, C. Effects of fish n-3 pufas on intestinal microbiota and immune system. Mar. Drugs 2019, 17, 374. [Google Scholar] [CrossRef] [Green Version]
- Hess, H.A.; Corl, B.A.; Lin, X.; Jacobi, S.K.; Harrell, R.J.; Blikslager, A.T.; Odle, J. Enrichment of intestinal mucosal phospholipids with arachidonic and eicosapentaenoic acids fed to suckling piglets is dose and time dependent. J. Nutr. 2008, 138, 2164–2171. [Google Scholar] [CrossRef] [Green Version]
- Lauridsen, C. Effects of dietary fatty acids on gut health and function of pigs pre- and post-weaning. J. Anim. Sci. 2020, 98, skaa086. [Google Scholar] [CrossRef]
- Tanghe, S.; De Smet, S. Does sow reproduction and piglet performance benefit from the addition of n-3 polyunsaturated fatty acids to the maternal diet? Vet. J. 2013, 197, 560–569. [Google Scholar] [CrossRef] [PubMed]
- Gabler, N.K.; Radcliffe, J.S.; Spencer, J.D.; Webel, D.M.; Spurlock, M.E. Feeding long-chain n-3 polyunsaturated fatty acids during gestation increases intestinal glucose absorption potentially via the acute activation of ampk. J. Nutr. Biochem. 2009, 20, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Brendemuhl, J.H.; Jeong, K.C.; Badinga, L. Effects of dietary omega-3 polyunsaturated fatty acids on growth and immune response of weanling pigs. J. Anim. Sci. Technol. 2014, 56, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.Y.; Yun, H.M.; Kim, I.H. Effects of dietary n-6/n-3 polyunsaturated fatty acids ratio on growth performance, apparent digestibility, blood lipid profiles, fecal microbiota, and meat quality in finishing pigs. Can. J. Anim. Sci. 2020, 100, 272–281. [Google Scholar]
- Council, N.R. Nutrient Requirements of Swine: Eleventh Revised Edition; The National Academies Press: Washington, DC, USA, 2012; p. 420. [Google Scholar]
- Takiishi, T.; Fenero, C.I.M.; Câmara, N.O.S. Intestinal barrier and gut microbiota: Shaping our immune responses throughout life. Tissue Barriers 2017, 5, e1373208. [Google Scholar] [CrossRef]
- Pié, S.; Lallès, J.P.; Blazy, F.; Laffitte, J.; Sève, B.; Oswald, I.P. Weaning is associated with an upregulation of expression of inflammatory cytokines in the intestine of piglets. J. Nutr. 2004, 134, 641–647. [Google Scholar] [CrossRef] [Green Version]
- Simopoulos, A.P. Omega-3 fatty acids in inflammation and autoimmune diseases. J. Am. Coll. Nutr. 2002, 21, 495–505. [Google Scholar] [CrossRef]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes. Nutrients 2010, 2, 355–374. [Google Scholar] [CrossRef] [Green Version]
- Giacobbe, J.; Benoiton, B.; Zunszain, P.; Pariante, C.M.; Borsini, A. The anti-inflammatory role of omega-3 polyunsaturated fatty acids metabolites in pre-clinical models of psychiatric, neurodegenerative, and neurological disorders. Front. Psychiatry 2020, 11, 122. [Google Scholar] [CrossRef]
- McAfee, J.M.; Kattesh, H.G.; Lindemann, M.D.; Voy, B.H.; Kojima, C.J.; Burdick Sanchez, N.C.; Carroll, J.A.; Gillespie, B.E.; Saxton, A.M. Effect of omega-3 polyunsaturated fatty acid (n-3 pufa) supplementation to lactating sows on growth and indicators of stress in the postweaned pig12. J. Anim. Sci. 2019, 97, 4453–4463. [Google Scholar] [CrossRef]
- Lee, S.I.; Kang, K.S. Omega-3 fatty acids modulate cyclophosphamide induced markers of immunosuppression and oxidative stress in pigs. Sci. Rep. 2019, 9, 2684. [Google Scholar] [CrossRef]
- Uddin, M.K.; Hasan, S.; Mahmud, M.R.; Peltoniemi, O.; Oliviero, C. In-feed supplementation of resin acid-enriched composition modulates gut microbiota, improves growth performance, and reduces post-weaning diarrhea and gut inflammation in piglets. Animals 2021, 11, 2511. [Google Scholar] [CrossRef]
- Adkins, Y.; Kelley, D.S. Mechanisms underlying the cardioprotective effects of omega-3 polyunsaturated fatty acids. J. Nutr. Biochem. 2010, 21, 781–792. [Google Scholar] [CrossRef]
- Liu, F.; Smith, A.D.; Solano-Aguilar, G.; Wang, T.T.Y.; Pham, Q.; Beshah, E.; Tang, Q.; Urban, J.F.; Xue, C.; Li, R.W. Mechanistic insights into the attenuation of intestinal inflammation and modulation of the gut microbiome by krill oil using in vitro and in vivo models. Microbiome 2020, 8, 83. [Google Scholar] [CrossRef]
- Liu, Y. Fatty acids, inflammation and intestinal health in pigs. J. Anim. Sci. Biotechnol. 2015, 6, 41. [Google Scholar] [CrossRef] [Green Version]
- Mankertz, J.; Tavalali, S.; Schmitz, H.; Mankertz, A.; Riecken, E.O.; Fromm, M.; Schulzke, J.D. Expression from the human occludin promoter is affected by tumor necrosis factor alpha and interferon gamma. Journal of cell science 2000, 113 Pt 11, 2085–2090. [Google Scholar] [CrossRef]
- Zolotarevsky, Y.; Hecht, G.; Koutsouris, A.; Gonzalez, D.E.; Quan, C.; Tom, J.; Mrsny, R.J.; Turner, J.R. A membrane-permeant peptide that inhibits mlc kinase restores barrier function in in vitro models of intestinal disease. Gastroenterology 2002, 123, 163–172. [Google Scholar] [CrossRef]
- Zeisel, M.B.; Dhawan, P.; Baumert, T.F. Tight junction proteins in gastrointestinal and liver disease. Gut 2019, 68, 547–561. [Google Scholar] [CrossRef]
- Hu, C.H.; Xiao, K.; Luan, Z.S.; Song, J. Early weaning increases intestinal permeability, alters expression of cytokine and tight junction proteins, and activates mitogen-activated protein kinases in pigs. J. Anim. Sci. 2013, 91, 1094–1101. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Zhang, Q.; Zhang, M.; Wang, C.; Zhu, Z.; Li, N.; Li, J. Effect of n-3 polyunsaturated fatty acids on membrane microdomain localization of tight junction proteins in experimental colitis. FEBS J. 2008, 275, 411–420. [Google Scholar] [CrossRef]
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Van de Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut microbiota dysbiosis in postweaning piglets: Understanding the keys to health. Trends Microbiol. 2017, 25, 851–873. [Google Scholar] [CrossRef]
- Guevarra, R.B.; Hong, S.H.; Cho, J.H.; Kim, B.-R.; Shin, J.; Lee, J.H.; Kang, B.N.; Kim, Y.H.; Wattanaphansak, S.; Isaacson, R.E.; et al. The dynamics of the piglet gut microbiome during the weaning transition in association with health and nutrition. J. Anim. Sci. Biotechnol. 2018, 9, 54. [Google Scholar] [CrossRef] [Green Version]
- Duarte, M.E.; Kim, S.W. Intestinal microbiota and its interaction to intestinal health in nursery pigs. Anim. Nutr. 2022, 8, 169–184. [Google Scholar] [CrossRef]
- Gryaznova, M.V.; Dvoretskaya, Y.D.; Syromyatnikov, M.Y.; Shabunin, S.V.; Parshin, P.A.; Mikhaylov, E.V.; Strelnikov, N.A.; Popov, V.N. Changes in the microbiome profile in different parts of the intestine in piglets with diarrhea. Animals 2022, 12, 320. [Google Scholar] [CrossRef]
- Fu, Y.; Wang, Y.; Gao, H.; Li, D.; Jiang, R.; Ge, L.; Tong, C.; Xu, K. Associations among dietary omega-3 polyunsaturated fatty acids, the gut microbiota, and intestinal immunity. Mediat. Inflamm. 2021, 2021, 8879227. [Google Scholar] [CrossRef] [PubMed]
- Wellington, V.N.A.; Sundaram, V.L.; Singh, S.; Sundaram, U. Dietary supplementation with vitamin d, fish oil or resveratrol modulates the gut microbiome in inflammatory bowel disease. Int. J. Mol. Sci. 2022, 23, 206. [Google Scholar] [CrossRef] [PubMed]
- Ciorba, M.A. A gastroenterologist’s guide to probiotics. Clin. Gastroenterol. Hepatol. 2012, 10, 960–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, S.; Gadonna-Widehem, P.; Rome, V.; Hamoudi, D.; Rhazi, L.; Lakhal, L.; Larcher, T.; Bahi-Jaber, N.; Pinon-Quintana, A.; Guyonvarch, A.; et al. Characterisation of early-life fecal microbiota in susceptible and healthy pigs to post-weaning diarrhoea. PLoS ONE 2017, 12, e0169851. [Google Scholar] [CrossRef] [PubMed]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short chain fatty acids (scfas)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Zhou, X.; Zhao, J.; Wang, Z.; Ye, H.; Pi, Y.; Che, D.; Han, D.; Zhang, S.; Wang, J. Sources of dietary fiber affect the scfa production and absorption in the hindgut of growing pigs. Front. Nutr. 2022, 8, 719935. [Google Scholar] [CrossRef]
- Amiri, P.; Hosseini, S.A.; Ghaffari, S.; Tutunchi, H.; Ghaffari, S.; Mosharkesh, E.; Asghari, S.; Roshanravan, N. Role of butyrate, a gut microbiota derived metabolite, in cardiovascular diseases: A comprehensive narrative review. Front. Pharmacol. 2021, 12, 837509. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GFS Mixture | |
|---|---|
| Dry matter (DM) % | 93.19 |
| Crude protein (CP) % | 24.6 |
| Ether extract (EE) % | 11.68 |
| Crude fibre (CF) % | 19 |
| Ash % | 4.26 |
| Items | Control Diet (%) | GFS Diet (%) |
|---|---|---|
| Corn | 68.46 | 63.51 |
| Soybean meal | 19 | 14 |
| Flax meal | - | 5 |
| Grapeseed meal | - | 3.75 |
| Sea buckthorn meal | - | 1.25 |
| Corn gluten | 4 | 4 |
| Milk replacer | 5 | 5 |
| L-lysine-HCl | 0.31 | 0.40 |
| Methionine | 0.10 | 0.12 |
| Limestone | 1.57 | 1.58 |
| Monocalcium phosphate | 0.35 | 0.18 |
| NaCl | 0.10 | 0.10 |
| Choline premix | 0.10 | 0.10 |
| Phytase | 0.01 | 0.01 |
| Mineral–vitamin premix * | 1 | 1 |
| Calculated nutrient content | ||
| ME (kcal/kg) | 3282.60 | 3265.70 |
| Crude protein (%) | 18.70 | 18.29 |
| Lysine (%) | 1.20 | 1.20 |
| Met + Cys (%) | 0.72 | 0.72 |
| Calcium (%) | 0,90 | 0.90 |
| Phosphorus (%) | 0.72 | 0.72 |
| Fat (%) | 2.40 | 2.70 |
| Cellulose (%) | 4.24 | 4.90 |
| Dry matter | 89.44 | 89.49 |
| g FAME */100 g Total FAME | Fatty Acids | GFS Meal Mix | Control Diet | GFS Diet |
|---|---|---|---|---|
| Caprylic | C8:0 | 0.00 | 0.04 | 0.11 |
| Capric | C10:0 | 0.02 | 0.06 | 0.04 |
| Lauric | C12:0 | 0.03 | 0.06 | 0.03 |
| Myristic | C14:0 | 0.16 | 0.16 | 0.13 |
| Pentadecanoic | C15:0 | 0.11 | 0.32 | 0.87 |
| Pentadecenoic | C15:1 | 0.05 | 0.15 | 0.71 |
| Palmitic | C16:0 | 11.99 | 15.79 | 18.33 |
| Palmitoleic | C16:1 | 3.65 | 0.37 | 1.39 |
| Heptadecanoic | C17:0 | 0.08 | 0.13 | 0.03 |
| Heptadecenoic | C17:1 | 0.04 | 0.05 | 0.14 |
| Stearic | C18:0 | 2.99 | 3.26 | 4.59 |
| Oleic cis | C18:1 | 19.94 | 34.60 | 34.31 |
| Linoleic cis | C18:2n6 | 22.94 | 41.93 | 31.49 |
| Linolenic α | C18:3n3 | 37.36 | 1.19 | 6.22 |
| Octadecatetraenoic | C18:4n3 | 0.13 | 0.50 | 0.55 |
| Eicosadienoic | C20(2n6) | 0.04 | 0.28 | 0.36 |
| Eicosatrienoic | C20(3n6) | 0.03 | 0.05 | 0.06 |
| Eicosatrienoic | C20(3n3) | 0.07 | 0.07 | 0.14 |
| Arachidonic | C20(4n6) | 0.12 | 0.20 | 0.28 |
| Eicosapentaenoic | C20(5n3) | 0.08 | 0.14 | 0.00 |
| Lignoceric | C 24:0 | 0.08 | 0.26 | 0.00 |
| Nervonic C24 (1n9) | C24 (1n9) | 0.00 | 0.26 | 0.00 |
| Other fatty acids | 0.11 | 0.12 | 0.21 | |
| Saturated fatty acids (SFA) | 15.45 | 20.09 | 24.13 | |
| Unsaturated fatty acids (UFA) | 84.44 | 82.46 | 75.65 | |
| Monounsaturated fatty acids (MUFA) | 23.68 | 38.10 | 36.55 | |
| Polyunsaturated fatty acids (PUFA) | 60.77 | 44.35 | 39.10 | |
| n3 (omega-3) | 37.64 | 1.90 | 6.91 | |
| n6 (omega-6) | 23.13 | 42.45 | 32.20 | |
| TOTAL FATTY ACIDS | 100 | 100 | 100 | |
| Parameters * | Control Diet | GFS Diet | p Value |
|---|---|---|---|
| Initial body weight, kg | 11.1 ± 1.17 | 11.3 ± 1.21 | 0.9385 |
| Final body weight, kg | 20.0 ± 3.22 | 23.5 ± 3.02 | 0.0792 |
| Average daily gain, kg | 0.384 ± 0.112 | 0.529 ± 0.140 | 0.1095 |
| Average daily feed intake, kg | 1.09 ± 0.158 | 1.00 ± 0.0501 | 0.8956 |
| Feed:gain ratio, kg:kg | 2.83 | 1.89 | 0.0681 |
| Parameters * | Control Diet | GFS Diet | p Value |
|---|---|---|---|
| Glycemia, mg/dL | 128 ± 26.1 | 132.6 ± 23.3 | 0.7855 |
| Cholesterol, mg/dL | 111 ± 17.4 | 104.9 ± 10.2 | 0.1265 |
| Triglycerides, mg/dL | 47.7 ± 6.92 | 57.5 ± 5.23 | 0.0721 |
| Total protein, mg/dL | 6.07 ± 0.252 | 5.90 ± 0.235 | 0.2898 |
| Albumin, g/dL | 3.30 ± 0.511 | 3.00 ± 0.0216 | 0.1449 |
| Total bilirubin, g/dL | 0.171 ± 0.0232 | 0.141 ± 0.0209 | 0.0875 |
| Creatinine, mg/dL | 1.50 ± 0.423 | 1.10 ± 0.352 | 0.1367 |
| Urea, mg/dL | 28.1 ± 5.80 | 27.6 ± 3.10 | 0.8444 |
| Phosphorus, mg/dL | 7.32 ± 1.23 | 6.79 ± 0.629 | 0.3438 |
| Calcium, mg/dL | 12.6 ± 0.914 | 12.8 ± 1.32 | 0.7670 |
| Magnesium, mg/dL | 1.60 ± 0.312 | 1.65 ± 0.159 | 0.7525 |
| Iron, μg/dL | 134 ± 40.1 | 93.2 ± 10.1 | 0.0680 |
| ALAT (TGP), U/L | 61.2 ± 12.5 | 58.7 ± 7.06 | 0.6977 |
| ASAT (TGO), U/L | 42.8 ± 6.90 | 41.3 ± 9.92 | 0.7677 |
| Alkaline phosphatase (ALK), U/L | 273 ± 52.1 | 321 ± 53.2 | 0.1530 |
| Gama glutamyl transferase (GGT), U/L | 17.5 ± 1.72 | 23.7 ± 2.86 | 0.0698 |
| SCFA/Exp Group * | Acetic Acid | Propionic Acid | Isobutyric Acid | Butyric Acid | Isovaleric Acid | Valeric Acid |
|---|---|---|---|---|---|---|
| Control | 61.33 ± 1.29 | 20.16 ± 0.78 | 1.8 ± 0.12 | 11.72 ± 0.41 | 1.88 ± 0.18 | 3.08 ± 0.22 |
| GFS | 47.48 ± 0.74 | 24.82 ± 1.21 | 2.34 ± 0.2 | 19.1 ± 0.72 | 2.12 ± 0.46 | 4.11 ± 0.19 |
| p value | <0.0001 | <0.01 | 0.27 | <0.0001 | 0.9 | 0.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marin, D.E.; Anghel, A.C.; Bulgaru, C.V.; Grosu, I.; Pistol, G.C.; Cismileanu, A.E.; Taranu, I. The Use of Agro-Industrial Waste Rich in Omega-3 PUFA during the Weaning Stress Improves the Gut Health of Weaned Piglets. Agriculture 2022, 12, 1142. https://doi.org/10.3390/agriculture12081142
Marin DE, Anghel AC, Bulgaru CV, Grosu I, Pistol GC, Cismileanu AE, Taranu I. The Use of Agro-Industrial Waste Rich in Omega-3 PUFA during the Weaning Stress Improves the Gut Health of Weaned Piglets. Agriculture. 2022; 12(8):1142. https://doi.org/10.3390/agriculture12081142
Chicago/Turabian StyleMarin, Daniela Eliza, Andrei Cristian Anghel, Cristina Valeria Bulgaru, Iulian Grosu, Gina Cecilia Pistol, Ana Elena Cismileanu, and Ionelia Taranu. 2022. "The Use of Agro-Industrial Waste Rich in Omega-3 PUFA during the Weaning Stress Improves the Gut Health of Weaned Piglets" Agriculture 12, no. 8: 1142. https://doi.org/10.3390/agriculture12081142
APA StyleMarin, D. E., Anghel, A. C., Bulgaru, C. V., Grosu, I., Pistol, G. C., Cismileanu, A. E., & Taranu, I. (2022). The Use of Agro-Industrial Waste Rich in Omega-3 PUFA during the Weaning Stress Improves the Gut Health of Weaned Piglets. Agriculture, 12(8), 1142. https://doi.org/10.3390/agriculture12081142